
Reproductive Biology of Male European Hake (Merluccius merluccius) in Central Mediterranean Sea: An Overview from Macroscopic to Molecular Investigation

 , ,
, ,  and
and 
Abstract
Simple Summary
Abstract

1. Introduction
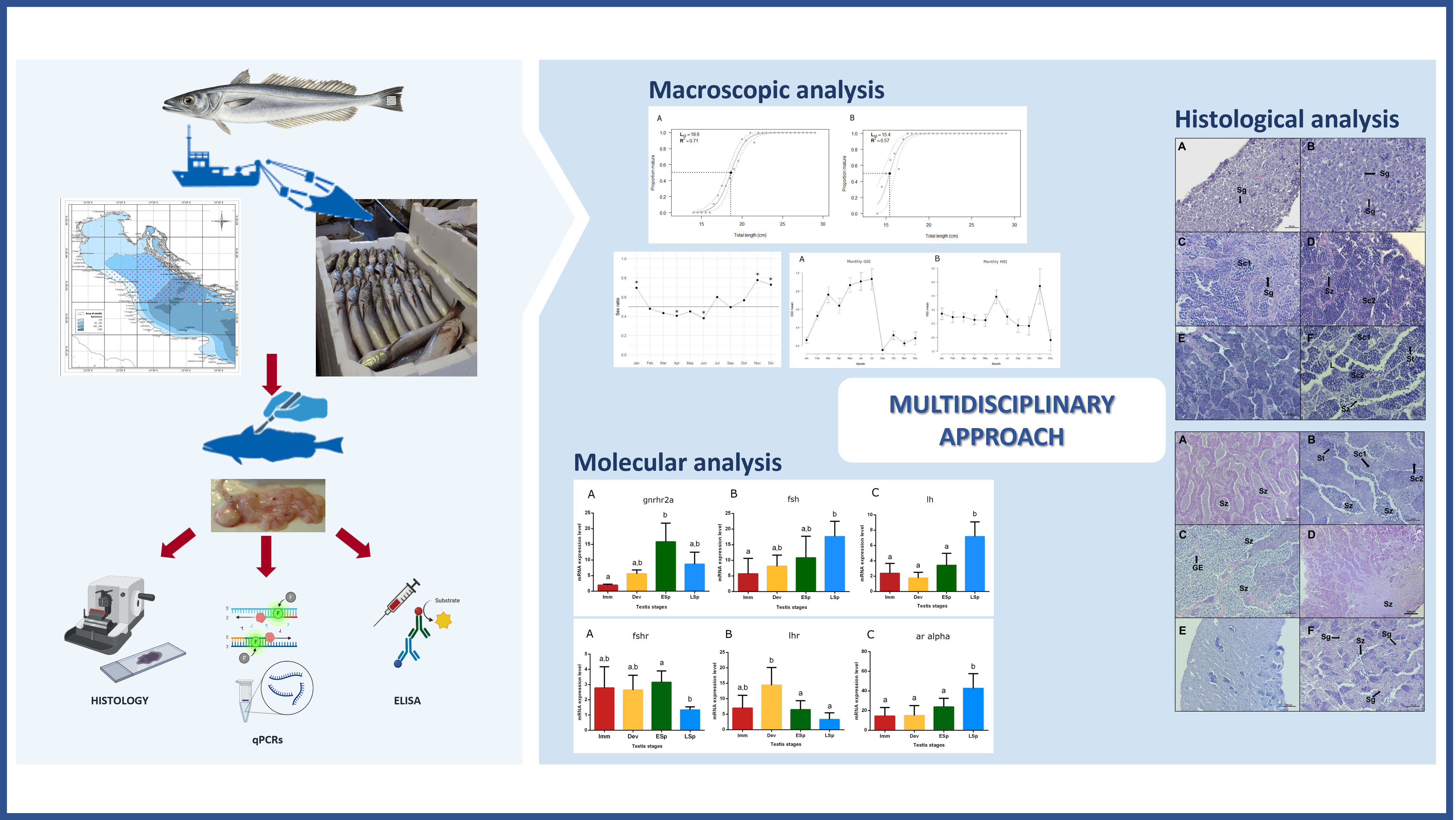
2. Materials and Methods
- -
- random specimens were collected each month from all three years by professional fishing trawlers on the landing port, then preserved on ice and transported to the laboratory for analysis. The analysis performed on these specimens was finalized for macroscopic and histological investigations (both sex for sex ratio calculation, only males for somatic indices evaluation, histological gonad assessment, and size at first maturity estimation).
- -
- three additional biological samplings were carried out by researchers onboard commercial fishing trawl vessels, during the daily fishing activity, in June, July, and September 2018, in order to dissect some male specimens selected from the catch, collect fresh tissues (pituitary and testis were preserved in RNA until later use) and drew blood immediately after death from the caudal peduncle area, as required by the protocols of hormonal and molecular analysis. The operator selected individuals in good condition and at different sizes, in order to obtain a total sample that favored the male sex and involved all length classes.
3. Results
3.1. Somatic Indices
3.2. Histology
3.3. Size at First Maturity (L50)
3.4. Gene Expression Profile
3.4.1. Pituitary Gland
3.4.2. Testis
3.5. 11-Ketotestosterone Immunoassay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bellido, J.M.; Sumaila, U.R.; Sánchez-Lizaso, J.L.; Palomares, M.L.; Pauly, D. Input versus Output Controls as Instruments for Fisheries Management with a Focus on Mediterranean Fisheries. Mar. Policy 2020, 118, 22–25. [Google Scholar] [CrossRef]
- Gomei, M.; Steenbeek, J.; Coll, M.; Claudet, J. 30 by 30: Scenarios to Recover Biodiversity and Rebuild Fish Stocks in the Mediterranean; World Wildlife Fund: Rome, Italy, 2021. [Google Scholar]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations FAO. The State of World Fisheries and Aquaculture 2020; Sustainability in Action: Rome, Italy, 2021; Volume 32, ISBN 9789251326923. [Google Scholar]
- FAO. Mediterranean and Black Sea (Major Fishing Area 37); Fisheries and Aquaculture Division: Rome, Italy, 2022. [Google Scholar]
- STECF. Scientific, Technical and Economic Committee for Fisheries (STECF)–Monitoring the Performance of the Common Fisheries Policy (STECF-Adhoc-20-01); STECF: Brussels, Belgium, 2018; ISBN 978-92-76-18115-6. [Google Scholar]
- General Fisheries Commission for the Mediterranean. The State of Mediterranean and Black Sea Fisheries 2022; FAO: Rome, Italy, 2022; ISBN 9789251373705. [Google Scholar]
- Food and Agriculture Organization of the United Nations. Review of the State of World Marine Fishery Resources; FAO: Rome, Italy, 2011; ISBN 9789251070239. [Google Scholar]
- Ungaro, N.; Mannini, P.; Vrgoc, N. Biology and Stock Assessment of Merluccius merluccius (L.) in Adriatic Sea: An Historical Review by Geographical Sub-Areas. Acta Adriat. 2003, 44, 9–20. [Google Scholar]
- Recasens, L.; Chiericoni, V.; Belcari, P. Spawning Pattern and Batch Fecundity of the European Hake (Merluccius merluccius (Linnaeus, 1758)) in the Western Mediterranean. Sci. Mar. 2008, 72, 721–732. [Google Scholar] [CrossRef]
- Sabatella, E.C.; Colloca, F.; Coppola, G.; Fiorentino, F.; Gambino, M.; Malvarosa, L.; Sabatella, R. Key Economic Characteristics of Italian Trawl Fisheries and Management Challenges. Front. Mar. Sci. 2017, 4, 371. [Google Scholar] [CrossRef]
- Grati, F.; Aladžuz, A.; Azzurro, E.; Bolognini, L.; Carbonara, P.; Çobani, M.; Domenichetti, F.; Dragičević, B.; Dulčić, J.; Durović, M.; et al. Seasonal Dynamics of Small-Scale Fisheries in the Adriatic Sea. Mediterr. Mar. Sci. 2018, 19, 21–35. [Google Scholar] [CrossRef]
- General Fisheries Commission for the Mediterranean (GFCM); Working Group on Stock Assessment of Demersal Species (WGSAD); Scientific Advisory Committee on Fisheries (SAC). 2021. Available online: https://www.fao.org/gfcm/technical-meetings/detail/en/c/1105308/ (accessed on 15 September 2022).
- Froese, R.; Pauly, D. World Wide Web electronic publication (Retrieved on: 05/12/2021). FishBase. 2021. Available online: www.fishbase.org (accessed on 4 June 2022).
- Zorica, B.; Isajlović, I.; Vrgoč, N.; Čikeš Keč, V.; Medvešek, D.; Vuletin, V.; Radonić, I.; Cvitanić, R.; Lepen Pleić, I.; Šestanović, M. Reproductive Traits of the European Hake, Merluccius merluccius (L. 1758), in the Adriatic Sea. Acta Adriat. 2021, 62, 183–198. [Google Scholar] [CrossRef]
- Domínguez-Petit, R.; Saborido-Rey, F. New bioenergetic perspective of European hake (Merluccius merluccius L.) reproductive ecology. Fish Res. 2010, 104, 83–88. [Google Scholar] [CrossRef]
- Recasens, L.; Lombarte, A.; Morales-Nin, B.; Tores, G.J. Spatiotemporal Variation in the Population Structure of the European Hake in the NW Mediterranean. J. Fish Biol. 1998, 53, 387–401. [Google Scholar] [CrossRef]
- Khoufi, W.; Ferreri, R.; Jaziri, H.; El Fehri, S.; Gargano, A.; Mangano, S.; Ben Meriem, S.; Romdhane, M.S.; Bonanno, A.; Aronica, S.; et al. Reproductive Traits and Seasonal Variability of Merluccius merluccius from the Tunisian Coast. J. Mar. Biol. Assoc. UK 2014, 94, 1545–1556. [Google Scholar] [CrossRef]
- Kahraman, A.E.; Yldz, T.; Uzer, U.; Karakulak, F.S. Sexual Maturity and Reproductive Patterns of European Hake Merluccius merluccius (Linnaeus, 1758) (Actinopterygii: Merlucciidae) from the Sea of Marmara, Turkey. Acta Zool. Bulg. 2017, 69, 385–392. [Google Scholar]
- Candelma, M.; Marisaldi, L.; Bertotto, D.; Radaelli, G.; Gioacchini, G.; Santojanni, A.; Colella, S.; Carnevali, O. Aspects of Reproductive Biology of the European Hake (Merluccius merluccius) in the Northern and Central Adriatic Sea (Gsa 17-Central Mediterranean Sea). J. Mar. Sci. Eng. 2021, 9, 389. [Google Scholar] [CrossRef]
- Carbonara, P.; Porcu, C.; Donnaloia, M.; Pesci, P.; Sion, L.; Spedicato, M.T.; Zupa, W.; Vitale, F.; Follesa, M.C. The Spawning Strategy of European Hake (Merluccius merluccius, L. 1758) across the Western and Central Mediterranean Sea. Fish. Res. 2019, 219, 105333. [Google Scholar] [CrossRef]
- The General Fisheries Commission for the Mediterranean. Establishment of Geographical Sub-Areas in the GFCM Area Amending the Resolution GFCM/31/2007/2; The General Fisheries Commission for the Mediterranea: Rome, Italy, 2009; pp. 26–30. [Google Scholar]
- Brown-Peterson, N.J.; Wyanski, D.M.; Saborido-Rey, F.; Macewicz, B.J.; Lowerre-Barbieri, S.K. A Standardized Terminology for Describing Reproductive Development in Fishes. Mar. Coast. Fish. Dyn. Manag. Ecosyst. Sci. 2011, 3, 52–70. [Google Scholar] [CrossRef]
- Candelma, M.; Valle, L.D.; Colella, S.; Santojanni, A.; Carnevali, O. Cloning, Characterization, and Molecular Expression of Gonadotropin Receptors in European Hake (Merluccius merluccius), a Multiple-Spawning Species. Fish Physiol. Biochem. 2018, 44, 895–910. [Google Scholar] [CrossRef] [PubMed]
- Okuzawa, K. Puberty in Teleosts. Fish Physiol. Biochem. 2002, 26, 31–41. [Google Scholar] [CrossRef]
- Taranger, G.L.; Carrillo, M.; Schulz, R.W.; Fontaine, P.; Zanuy, S.; Felip, A.; Weltzien, F.A.; Dufour, S.; Karlsen, Ø.; Norberg, B.; et al. Control of Puberty in Farmed Fish. Gen. Comp. Endocrinol. 2009, 165, 483–515. [Google Scholar] [CrossRef]
- Carrillo, M.; Zanuy, S.; Felip, A.; Bayarri, M.J.; Molés, G.; Gómez, A. Hormonal and Environmental Control of Puberty in Perciform Fish: The Case of Sea Bass. Ann. N. Y. Acad. Sci. 2009, 1163, 49–59. [Google Scholar] [CrossRef]
- Oliveira, M.R.; Costa, E.F.S.; Araújo, A.S.; Pessoa, E.K.R.; Chellappa, S. Sex Ratio and Length-Weight Relationship for Five Marine Fish Species from Brazil. J. Mar. Biol. Oceanogr. 2012, 1, 11–14. [Google Scholar] [CrossRef]
- Papastamatiou, Y.P.; Caselle, J.E.; Friedlander, A.M.; Lowe, C.G. Distribution, Size Frequency, and Sex Ratios of Blacktip Reef Sharks Carcharhinus melanopterus at Palmyra Atoll: A Predator-Dominated Ecosystem. J. Fish Biol. 2009, 75, 647–654. [Google Scholar] [CrossRef]
- Sun, C.L.; Chang, H.Y.; Liu, T.Y.; Yeh, S.Z.; Chang, Y.J. Reproductive Biology of the Black Marlin, Istiompax indica, off Southwestern and Eastern Taiwan. Fish. Res. 2015, 166, 12–20. [Google Scholar] [CrossRef]
- Chang, H.Y.; Sun, C.L.; Yeh, S.Z.; Chang, Y.J.; Su, N.J.; DiNardo, G. Reproductive Biology of Female Striped Marlin Kajikia audax in the Western Pacific Ocean. J. Fish Biol. 2018, 92, 105–130. [Google Scholar] [CrossRef] [PubMed]
- Manzo, C.; Cilenti, L.; Fabbrocini, A.; D’adamo, R. Population Size Structure, Growth and Reproduction of the European Anchovy (Engraulis encrasicolus, L.) in the Lagoon of Lesina (South-Western Adriatic Sea, Italy). Transit. Waters Bull. 2013, 7, 41–52. [Google Scholar] [CrossRef]
- Pešić, A.; Durović, M.; Joksimović, A.; Regner, S.; Simonović, P.; Glamuzina, B. Some Reproductive Patterns of the Sardine, Sardina pilchardus (Walb, 1792), in Boka Kotorska Bay (Montenegro, Southern Adriatic Sea). Acta Adriat. 2010, 51, 159–168. [Google Scholar]
- Park, J.J.; Kim, H.J.; Shin, S.R.; Jin, Y.G.; Kim, J.W.; Lee, J.S. Reproductive Cycle and Sexual Group Maturity Of. Fishes 2022, 7, 5026. [Google Scholar] [CrossRef]
- Somarakis, S.; Ganias, K.; Tserpes, G.; Koutsikopoulos, C. Ovarian Allometry and the Use of the Gonosomatic Index: A Case Study in the Mediterranean sardine, Sardina pilchardus. Mar. Biol. 2004, 146, 181–189. [Google Scholar] [CrossRef]
- Prager, M.H.; Saila, S.B.; Recksiek, C.W. FISHPARM: A Microcomputer Program for Parameter Estimation of Nonlinear Models by Important Note about FISHPARM Software; Old Dominion University: Norfolk, Virginia, 1989. [Google Scholar]
- Candelma, M.; Fontaine, R.; Colella, S.; Santojanni, A.; Weltzien, F.-A.; Carnevali, O. Gonadotropin Characterization, Localization and Expression in the European Hake (Merluccius merluccius); Reproduction: Cambridge, UK, 2017. [Google Scholar] [CrossRef]
- Cuisset, B.; Pradelles, P.; Kime, D.E.; Kühn, E.R.; Babin, P.; Davail, S.; Le Menn, F. Enzyme Immunoassay for 11-Ketotestosterone Using Acetylcholinesterase as Laberl: Application to the Measurement of 11-Ketotestosterone in Plasma of Siberian Sturgeon. Comp. Biochem. Physiol. Part C Pharmacol. 1994, 108, 229–241. [Google Scholar] [CrossRef]
- Rodríguez, L.; Begtashi, I.; Zanuy, S.; Carrillo, M. Long-Term Exposure to Continuous Light Inhibits Precocity in European Male Sea Bass (Dicentrarchus labrax L.): Hormonal Aspects. Gen. Comp. Endocrinol. 2005, 140, 116–125. [Google Scholar] [CrossRef]
- Cohen, J. A Coefficient of Agreement for Nominal Scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- McFadden, D. Conditional Logit Analysis of Qualitative Choice Behaviour. Front. Econom. 1974, 105–142. [Google Scholar]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Soykan, O.; Ilkyaz, A.T.; Metan, G.; Kinacigal, H.T. Age, Growth and Reproduction of European Hake (Merluccius merluccius (Linn., 1758)) in the Central Aegean Sea, Turkey. J. Mar. Biol. Assoc. UK 2011, 95, 829–837. [Google Scholar] [CrossRef]
- Sion, L.; Zupa, W.; Calculli, C.; Garofalo, G.; Hidalgo, M.; Jadaud, A.; Lefkaditou, E.; Ligas, A.; Peristeraki, P.; Bitetto, I.; et al. Spatial Distribution Pattern of European Hake, Merluccius merluccius (Pisces: Merlucciidae), in the Mediterranean Sea. Sci. Mar. 2019, 83, 21–32. [Google Scholar] [CrossRef]
- D’Iglio, C.; Porcino, N.; Savoca, S.; Profeta, A.; Perdichizzi, A.; Armeli Minicante, E.; Salvati, D.; Soraci, F.; Rinelli, P.; Giordano, D. Ontogenetic Shift and Feeding Habits of the European Hake (Merluccius Merluccius L., 1758) in Central and Southern Tyrrhenian Sea (Western Mediterranean Sea): A Comparison between Past and Present Data. Ecol. Evol. 2022, 12, e8634. [Google Scholar] [CrossRef]
- D’Iglio, C.; Famulari, S.; Albano, M.; Giordano, D.; Rinelli, P.; Capillo, G.; Spanò, N.; Savoca, S. Time-Scale Analysis of Prey Preferences and Ontogenetic Shift in the Diet of European Hake Merluccius merluccius (Linnaeus, 1758) in Southern and Central Tyrrhenian Sea. Fishes 2022, 7, 167. [Google Scholar] [CrossRef]
- Ministero Delle Politiche Agricole Alimentari e Forestali. Decreto 30 Ottobre 2007; Gazzetta Ufficiale: Rome, Italy, 2007; Volume 18, p. 125. [Google Scholar]
- Morales-Nin, B.; Moranta, J. Recruitment and Post-Settlement Growth of Juvenile Merluccius merluccius on the Western Mediterranean Shelf. Sci. Mar. 2004, 68, 399–409. [Google Scholar] [CrossRef]
- Arneri, E.; Morales-Nin, B. Aspects of the Early Life History of European Hake from the Central Adriatic. J. Fish Biol. 2000, 56, 1368–1380. [Google Scholar] [CrossRef]
- Uçkun, D.; Taşkavak, E.; Toǧulga, M. A Preliminary Study on Otolith-Total Length Relationship of the Common Hake (Merluccius merluccius L., 1758) in İzmir Bay, Aegean Sea. Pakistan J. Biol. Sci. 2006, 9, 1720–1725. [Google Scholar] [CrossRef]
- Belcari, P.; Ligas, A.; Viva, C. Age Determination and Growth of Juveniles of the European Hake, Merluccius merluccius (L., 1758), in the Northern Tyrrhenian Sea (NW Mediterranean). Fish. Res. 2006, 78, 211–217. [Google Scholar] [CrossRef]
- Murua, H.; Saborido-Rey, F. Female Reproductive Strategies of Marine Fish Species of the North Atlantic. J. Northw. Atl. Fish. Sci. 2003, 33, 23–31. [Google Scholar] [CrossRef]
- Al-Absawy, M.A.E.G. The Reproductive Biology and the Histological and Ultrastructural Characteristics in Ovaries of the Female Gadidae Fish Merluccius merluccius from the Egyptian Mediterranean Water. African J. Biotechnol. 2010, 9, 2544–2559. [Google Scholar]
- Piñeiro, C.; Saínza, M. Age Estimation, Growth and Maturity of the European Hake (Merluccius merluccius (Linnaeus, 1758)) from Iberian Atlantic Waters. J. Mar. Sci. 2003, 60, 1086–1102. [Google Scholar] [CrossRef]
- El Habouz, H.; Recasens, L.; Kifani, S.; Moukrim, A.; Bouhaimi, A.; El Ayoubi, S. Maturity and Batch Fecundity of the European Hake (Merluccius merluccius, Linnaeus, 1758) in the Eastern Central Atlantic. Sci. Mar. 2011, 75, 447–454. [Google Scholar] [CrossRef]
- Zupanovic, S.; Jardas, I. A Contribution to the Study of Biology and Population Dynamics of Adriatic Hake, Merluccius merluccius (L.). Acta Adriat. 1986, 27, 97–149. [Google Scholar]
- Ungaro, N.; Rizzi, E.; Marano, G. Note Sulla Biologia e Pesca Di Merluccius merluccius (L.) Nell’Adriatico Pugliese. Biol. Mar. Suppl. Notiziario SIBM 1993, 1, 329–334. [Google Scholar]
- Murua, H.; Motos, L. Reproductive Strategy and Spawning Activity of the European Hake Merluccius merluccius (L.) in the Bay of Biscay. J. Fish Biol. 2006, 69, 1288–1303. [Google Scholar] [CrossRef]
- Alvarez, P.; Fives, J.; Motos, L.; Santos, M. Distribution and Abundance of European Hake Merluccius merluccius (L.), Eggs and Larvae in the North East Atlantic Waters in 1995 and 1998 in Relation to Hydrographic Conditions. J. Plankton Res. 2004, 26, 811–826. [Google Scholar] [CrossRef]
- Costa, A.M. Somatic Condition, Growth and Reproduction of Hake, Merluccius merluccius L., in the Portuguese Coast. Open J. Mar. Sci. 2013, 3, 12–30. [Google Scholar] [CrossRef]
- García-Fernández, C.; Domínguez-Petit, R.; Saborido-Rey, F. The Use of Daily Growth to Analyze Individual Spawning Dynamics in an Asynchronous Population: The Case of the European Hake from the Southern Stock. Fishes 2022, 7, 208. [Google Scholar] [CrossRef]
- Korta, M.; García, D.; Santurtún, M.; Goikoetxea, N.; Andonegi, E.; Murua, H.; Álvarez, P.; Cerviño, S.; Castro, J.; Murillas, A. European Hake (Merluccius merluccius) in the Northeast Atlantic Ocean; Wiley: New York, NY, USA, 2015; ISBN 9781118568262. [Google Scholar]
- Htun-Han, M. The Reproductive Biology of the Dab Limanda limanda (L.) in the North Sea: Gonosomatic Index, Hepatosomatic Index and Condition Factor. J. Fish Biol. 1978, 13, 369–378. [Google Scholar] [CrossRef]
- Lloret, J.; Demestre, M.; Sanchez-Pardo, J. Lipid (Energy) Reserves of European Hake (Merluccius merluccius) in the North-Western Mediterranean. Vie Milieu/Life Environ. 2008, 58, 77–85. [Google Scholar]
- Sainmont, J.; Andersen, K.H.; Varpe, Ø.; Visser, A.W. Capital versus Income Breeding in a Seasonal Environment. Am. Nat. 2014, 184, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Varpe, Ø.; Jørgensen, C.; Tarling, G.A.; Fiksen, Ø. The Adaptive Value of Energy Storage and Capital Breeding in Seasonal Environments. Oikos 2009, 118, 363–370. [Google Scholar] [CrossRef]
- Ferrer Maza, D.; Lloret, J.; Muñoz, M.; Faliex, E.; Vila, S.; Sasal, P. Parasitism, Condition and Reproduction of the European Hake (Merluccius merluccius) in the Northwestern Mediterranean Sea. J. Mar. Sci. 2014, 71, 1088–1099. [Google Scholar] [CrossRef]
- Domínguez-Petit, R.; Korta, M.; Saborido-Rey, F.; Murua, H.; Sainza, M.; Piñeiro, C. Changes in Size at Maturity of European Hake Atlantic Populations in Relation with Stock Structure and Environmental Regimes. J. Mar. Syst. 2008, 71, 260–278. [Google Scholar] [CrossRef]
- West, G. Methods of Assessing Ovarian Development in Fishes:A Review. Mar. Freshw. Res. 1990, 41, 199–222. [Google Scholar] [CrossRef]
- McPherson, L.R.; Ganias, K.; Marshall, C.T. Inaccuracies in Routinely Collected Atlantic Herring (Clupea harengus) Maturity Data and Correction Using a Gonadosomatic Index Model. J. Mar. Biol. Assoc. UK 2011, 91, 1477–1487. [Google Scholar] [CrossRef]
- Flores, A.; Wiff, R.; Ganias, K.; Marshall, C.T. Accuracy of Gonadosomatic Index in Maturity Classification and Estimation of Maturity Ogive. Fish. Res. 2019, 210, 50–62. [Google Scholar] [CrossRef]
- Vrgoč, N.; Arneri, E.; Jukić-Peladić, S.; Šifner, S.K.; Mannini, P.; Marčeta, B.; Osmani, K.; Piccinetti, C.; Ungaro, N. Review of Current Knowledge on shared demersal stocks of the Adriatic Sea. In AdriaMed Technical Documents; FAO-MiPAAF Scientific Cooperation to Support Responsible Fisheries in the Adriatic Sea: Termoli, Italy, 2004; Volume 12, 91p. [Google Scholar]
- Ferreri, R.; Basilone, G.; D’Elia, M.; Traina, A.; Saborido-Rey, F.; Mazzola, S. Validation of Macroscopic Maturity Stages According to Microscopic Histological Examination for European Anchovy. Mar. Ecol. 2009, 30, 181–187. [Google Scholar] [CrossRef]
- Tomkiewicz, J.; Tybjerg, L.; Jespersen, Å. Micro- and Macroscopic Characteristics to Stage Gonadal Maturation of Female Baltic Cod. J. Fish Biol. 2003, 62, 253–275. [Google Scholar] [CrossRef]
- Marisaldi, L.; Basili, D.; Candelma, M.; Sesani, V.; Pignalosa, P.; Gioacchini, G.; Carnevali, O. Maturity Assignment Based on Histology-Validated Macroscopic Criteria: Tackling the Stock Decline of the Mediterranean Swordfish (Xiphias gladius). Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 303–314. [Google Scholar] [CrossRef]
- European Union. Regulation (EU) 2019/1241 of the European Parliament and of the Council of 20 June 2019 on the Conservation of Fisheries Resources and the Protection of Marine Ecosystems through Technical Measures. Off. J. Eur. Union 2019. Available online: http://data.europa.eu/eli/reg/2019/1241/oj (accessed on 23 October 2022).
- Anderson, C.N.K.; Hsieh, C.H.; Sandin, S.A.; Hewitt, R.; Hollowed, A.; Beddington, J.; May, R.M.; Sugihara, G. Why Fishing Magnifies Fluctuations in Fish Abundance. Nature 2008, 452, 835–839. [Google Scholar] [CrossRef] [PubMed]
- González-Martínez, D.; Madigou, T.; Mañanos, E.; Cerdá-Reverter, J.M.; Zanuy, S.; Kah, O.; Muñoz-Cueto, J.A. Cloning and Expression of Gonadotropin-Releasing Hormone Receptor in the Brain and Pituitary of the European Sea Bass: An in Situ Hybridization Study. Biol. Reprod. 2004, 70, 1380–1391. [Google Scholar] [CrossRef] [PubMed]
- Sohn, Y.C.; Yoshiura, Y.; Kobayashi, M.; Aida, K. Seasonal Changes in MRNA Levels of Gonadotropin and Thyrotropin Subunits in the Goldfish, Carassius auratus. Gen. Comp. Endocrinol. 1999, 113, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Kajimura, S.; Yoshiura, Y.; Suzuki, M.; Aida, K. CDNA Cloning of Two Gonadotropin β Subunits (GTH-Iβand -IIβ) and Their Expression Profiles during Gametogenesis in the Japanese Flounder (Paralichthys olivaceus). Gen. Comp. Endocrinol. 2001, 122, 117–129. [Google Scholar] [CrossRef]
- Mateos, J.; Querat, B.; Zanuy, S. Molecular Characterization of Sea Bass Gonadotropin Subunits (α, FSHβ, and LHβ) and Their Expression during the Reproductive Cycle. Gen. Comp. Endocrinol. 2003, 133, 216–232. [Google Scholar] [CrossRef]
- Ministero delle politiche agricole alimentari e forestali (MiPAAF). Decreto N. 229107 Del 18 Maggio 2021. Disposizioni in Materia Di Interruzione Temporanea Obbligatoria Delle Attività Di Pesca Esercitate Mediante L’Utilizzo Di Attrezzi Trainati “Reti a Strascico a Divergenti (Otb)”, “Reti Gemelle a Divergenti(OTT)” E/O “Sfogliare—Rapidi (Tbb)”. 2021, pp. 1–10. Available online: https://www.politicheagricole.it/flex/cm/pages/ServeBLOB.php/L/IT/IDPagina/17050 (accessed on 23 October 2022).












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Primer Sequence (5′-3′) | Orientation | Annealing Temperature (°C) | |
|---|---|---|---|---|---|
| Testis | b-act | EU022566 | 5′-GTCATGGACTCCGGTGATGG-3′ | forward | 60 °C |
| 5′-GAGGTAGTCTGTGAGGTCGC-3′ | reverse | 60 °C | |||
| 18S | KF986702 | 5′-GAGGCCCTGTAATTGGAATG-3′ | forward | 60 °C | |
| 5′-CGCAAGACACTCAACCAAGA-3′ | reverse | 60 °C | |||
| fshr | KY178270 | 5′-CATGGCCGTGCTCATCTTC-3′ | forward | 58 °C | |
| 5′-ATGAAGAGGAAGGGGTTGGC-3′ | reverse | 58 °C | |||
| lhr | KY178271 | 5′-GTCAGCGAGTTGGACATGGA-3′ | forward | 61 °C | |
| 5′-ATGACCCAGGTGAGAAAGCG-3′ | reverse | 61 °C | |||
| ar-alpha | ON736432 | 5′-AAGCCATACCAGGTTTCCGT-3′ | forward | 57 °C | |
| 5′-GATCAGGTCTGGAGCGAAGT-3′ | reverse | 57 °C | |||
| Pituitary | b-act | EU022566 | 5′-GTCATGGACTCCGGTGATGG-3′ | forward | 60 °C |
| gland | 5′-GAGGTAGTCTGTGAGGTCGC-3′ | reverse | 60 °C | ||
| 18S | KF986702 | 5′-GAGGCCCTGTAATTGGAATG-3′ | forward | 60 °C | |
| 5′-CGCAAGACACTCAACCAAGA-3′ | reverse | 60 °C | |||
| fshb | KX377614 | 5′-TCTGTCGCCCAGTCAACTTC-3′ | forward | 58 °C | |
| 5′-CCCACCGGACAGTCTTCAAA-3′ | reverse | 58 °C | |||
| lhb | KX377615 | 5′-CAGCGGACACTGCATCAC-3′ | forward | 60 °C | |
| 5′-ACAGTCCGGCAGCTCAAA-3′ | reverse | 60 °C | |||
| gnrh-r2a | ON736433 | 5′-CGTTCCTCAGTTGTTCCTCT-3′ | forward | 60 °C | |
| 5′-CCAGTGGGTGTCGAAGCTG-3′ | reverse | 60 °C |
| MONTH | N° of Fish | N° of Males | N° of Females | Sex Ratio (M:F) | Sex Ratio (Females/Males + Females) | χ2 | p-Value |
|---|---|---|---|---|---|---|---|
| January | 122 | 37 | 85 | 0.44:1 | 0.697 | 18.885 | 1.388 × 10−5 * |
| February | 123 | 64 | 59 | 1.08:1 | 0.480 | 0.20325 | 0.6521 |
| March | 122 | 69 | 53 | 1.30:1 | 0.434 | 2.0984 | 0.1475 |
| April | 133 | 79 | 54 | 1.46:1 | 0.406 | 4.6992 | 0.03018 * |
| May | 142 | 78 | 64 | 1.22:1 | 0.451 | 1.3803 | 0.2401 |
| June | 113 | 70 | 43 | 1.63:1 | 0.381 | 6.4513 | 0.01109 * |
| July | 95 | 38 | 57 | 0.67:1 | 0.600 | 3.80 | 0.05125 |
| September | 105 | 53 | 52 | 1.02:1 | 0.495 | 0.009524 | 0.9223 |
| October | 111 | 48 | 63 | 0.76:1 | 0.568 | 2.027 | 0.1545 |
| November | 117 | 26 | 91 | 0.29:1 | 0.778 | 36.111 | 1.864 × 10−9 * |
| December | 133 | 36 | 97 | 0.37:1 | 0.729 | 27.977 | 1.227 × 10−7 * |
| TOTAL | 1316 | 598 | 718 | 0.83:1 | 0.545 | 10.942 | 0.00094 * |
| L50 MACROSCOPIC | ||||
|---|---|---|---|---|
| Coefficient | Estimate | Std. Error (SE) | z Value | p Value |
| a | −21.08 | 3.14 | −6.73 | <0.0001 |
| b | 1.13 | 0.17 | 6.81 | <0.0001 |
| L50 HISTOLOGICAL | ||||
| Coefficient | Estimate | Std. Error (SE) | z Value | p Value |
| a | −22.13 | 6.02 | −3.68 | 0.00023 |
| b | 1.44 | 0.37 | 3.89 | 0.00010 |
| Estimation of Size at First Maturity | Likelihood Ratio Test | Cohen’s k Coefficient of Agreement | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Value | Conf. Interval | R2MF | LogLik | Chi-sq | p Value | Value | Conf. Interval | ASE | z Value | p Value | |
| Macroscopic Method | 18.6 | 18.2–19.0 | 0.71 | −59.42 | 73.63 | <0.0001 | 0.24 | 0.13–0.36 | 0.06 | 4.10 | 0.000042 |
| Histological Method | 15.4 | 14.6–16.1 | 0.57 | −22.61 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mascoli, A.; Candelma, M.; Santojanni, A.; Carnevali, O.; Colella, S. Reproductive Biology of Male European Hake (Merluccius merluccius) in Central Mediterranean Sea: An Overview from Macroscopic to Molecular Investigation. Biology 2023, 12, 562. https://doi.org/10.3390/biology12040562
Mascoli A, Candelma M, Santojanni A, Carnevali O, Colella S. Reproductive Biology of Male European Hake (Merluccius merluccius) in Central Mediterranean Sea: An Overview from Macroscopic to Molecular Investigation. Biology. 2023; 12(4):562. https://doi.org/10.3390/biology12040562
Chicago/Turabian StyleMascoli, Alessia, Michela Candelma, Alberto Santojanni, Oliana Carnevali, and Sabrina Colella. 2023. "Reproductive Biology of Male European Hake (Merluccius merluccius) in Central Mediterranean Sea: An Overview from Macroscopic to Molecular Investigation" Biology 12, no. 4: 562. https://doi.org/10.3390/biology12040562
APA StyleMascoli, A., Candelma, M., Santojanni, A., Carnevali, O., & Colella, S. (2023). Reproductive Biology of Male European Hake (Merluccius merluccius) in Central Mediterranean Sea: An Overview from Macroscopic to Molecular Investigation. Biology, 12(4), 562. https://doi.org/10.3390/biology12040562