

Cryptic mtDNA Diversity of Diopatra cuprea (Onuphidae, Annelida) in the Northwestern Atlantic Ocean

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
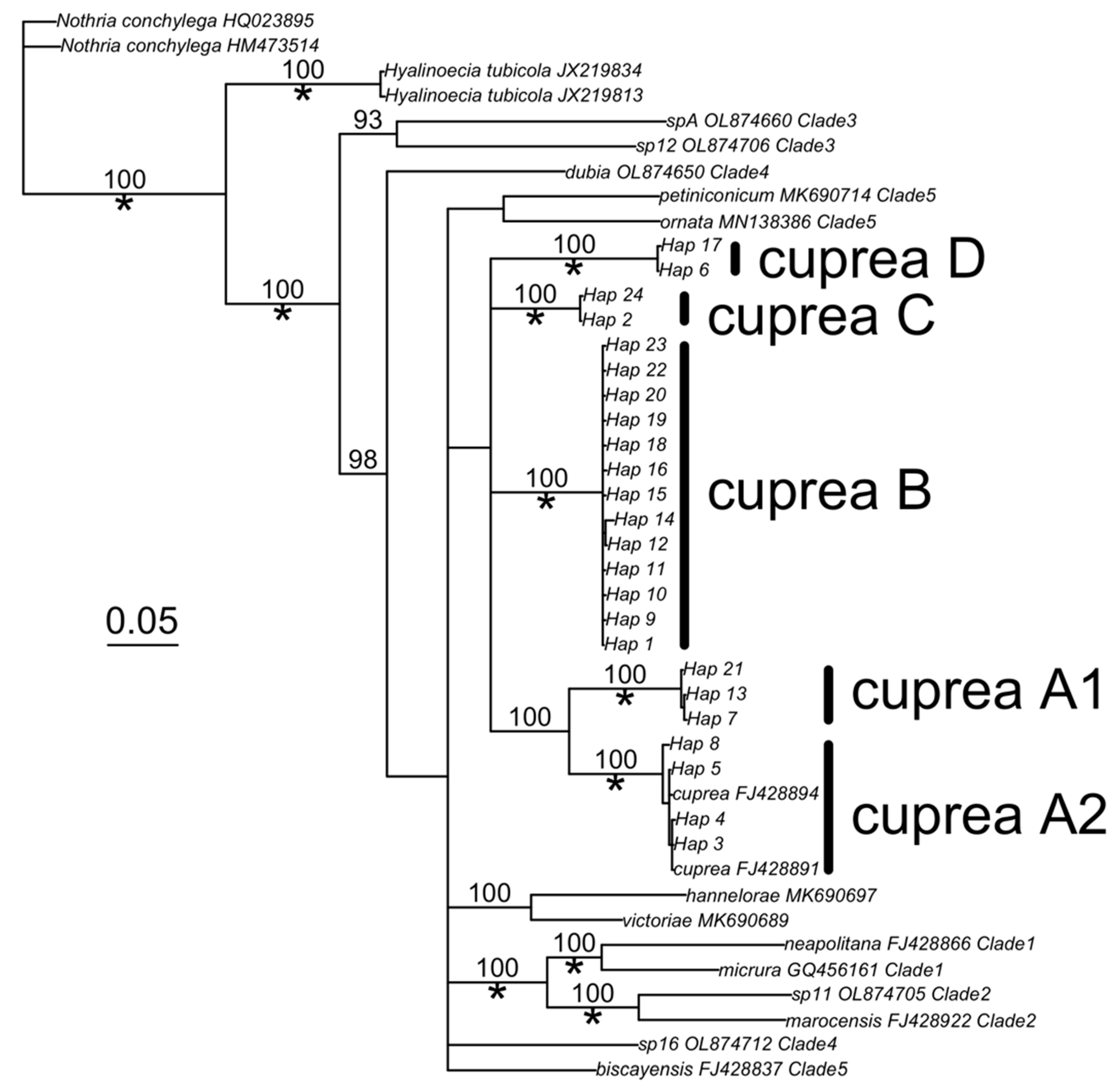
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andrade, S.C.S.; Novo, M.; Kawauchi, G.Y.; Worsaae, K.; Pleijel, F.; Giribet, G.; Rouse, G.W. Articulating “Archiannelids”: Phylogenomics and Annelid Relationships, with Emphasis on Meiofaunal Taxa. Mol. Biol. Evol. 2015, 32, 2860–2875. [Google Scholar] [CrossRef] [PubMed]
- Carr, C.M.; Hardy, S.M.; Brown, T.M.; Macdonald, T.A.; Hebert, P.D.N. A tri-oceanic perspective: DNA barcoding reveals geographic structure and cryptic diversity in Canadian polychaetes. PLoS ONE 2011, 6, e22232. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, P.; Kupriyanova, E. Cosmopolitan polychaetes—Fact or fiction? Personal and historical perspectives. Invertebr. Syst. 2018, 32, 1–9. [Google Scholar] [CrossRef]
- Weigert, A.; Bleidorn, C. Current status of annelid phylogeny. Org. Divers. Evol. 2016, 16, 345–362. [Google Scholar] [CrossRef]
- Day, J.H. Polychaeta of Southern Africa Parts 1: Errantia and 2: Sedentaria; Trustees of the British Museum (Natural History): London, UK, 1967. [Google Scholar]
- Hartman, O. Polychaetous Annelids. Part V. Eunicea. In Allan Hancock Pacific Expeditions; The University of Southern California Press: Los Angeles, CA, USA, 1944; Volume 10, pp. 1–237. Available online: https://www.biodiversitylibrary.org/page/4679950 (accessed on 15 March 2023).
- Kirkegaard, J.B. The Polychaeta of West Africa Part II. Errant Species. 2. Nephtyidae to Dorvilleidae. Atlantide Rep. 1988, 14, 7–89. [Google Scholar]
- Paxton, H. Diopatra Audouin and Milne Edwards (Polychaeta: Onuphidae) from Australia, with a discussion of developmental patterns in the genus. Beagle Rec. North. Territ. Mus. Arts Sci. 1993, 10, 115–154. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H. The cryptogenic bait worm Diopatra biscayensis Fauchald et al., 2012 (Annelida: Onuphidae)—Revisiting its history, biology and ecology. Estuar. Coast. Shelf Sci. 2015, 163, 22–36. [Google Scholar] [CrossRef]
- Budaeva, N.; Fauchald, K. Diopatra tuberculantennata, a new species of Onuphidae (Polychaeta) from Belize with a key to onuphids from the Caribbean Sea. Zootaxa 2008, 1795, 29045. [Google Scholar] [CrossRef]
- Elgetany, A.H.; van Rensburg, H.; Hektoen, M.; Matthee, C.; Budaeva, N.; Simon, C.A.; Struck, T.H. Species delineation in the speciation grey zone—The case of Diopatra (Annelida, Onuphidae). Zool. Scr. 2020, 49, 516–534. [Google Scholar] [CrossRef]
- Hektoen, M.M.; Willassen, E.; Budaeva, N. Phylogeny and Cryptic Diversity of Diopatra (Onuphidae, Annelida) in the East Atlantic. Biology 2022, 11, 327. [Google Scholar] [CrossRef]
- Paxton, H. The Diopatra chiliensis confusion—Redescription of D. chiliensis (Polychaeta, Onuphidae) and implicated species. Zool. Scr. 1998, 27, 31–48. [Google Scholar] [CrossRef]
- Paxton, H. Diopatra nishii, a new brooding species of Onuphidae (Annelida) from Japan. Mar. Biodivers. 2015, 45, 711–717. [Google Scholar] [CrossRef]
- Paxton, H. A new species of Diopatra (Annelida: Onuphidae) from Namibia, southwestern Africa. Mar. Biodivers. 2016, 46, 889–895. [Google Scholar] [CrossRef]
- Paxton, H.; Arias, A. Unveiling a surprising diversity of the genus Diopatra Audouin & Milne Edwards, 1833 (Annelida: Onuphidae) in the Macaronesian region (eastern North Atlantic) with the description of four new species. Zootaxa 2017, 4300, 505. [Google Scholar]
- Paxton, H.; Fadlaoui, S.; Lechapt, J.P. Diopatra marocensis, a New Brooding Species of Onuphidae (Annelida, Polychaeta). J. Mar. Biol. Assoc. United Kingd. 1995, 75, 949–955. [Google Scholar] [CrossRef]
- Pires, A.; Paxton, H.; Quintino, V.; Rodrigues, A.M. Diopatra (Annelida: Onuphidae) diversity in European waters with the description of Diopatra micrura, new species. Zootaxa 2010, 2395, 17–33. [Google Scholar] [CrossRef]
- World Polychaeta Database. Diopatra cuprea (Bosc, 1802). Read, G., Fauchald, K., Eds.; Accessed through: World Register of Marine Species. 2023. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=157339 (accessed on 15 March 2023).
- Seixas, V.C.; Steiner, T.M.; Solé-Cava, A.M.; Amaral, A.C.Z.; Paiva, P.C. Hidden diversity within the Diopatra cuprea complex (Annelida: Onuphidae): Morphological and genetics analyses reveal four new species in the south-west Atlantic. Zool. J. Linn. Soc. 2021, 191, 637–671. [Google Scholar] [CrossRef]
- Berke, S.K. Biogeographic variability in ecosystem engineering: Patterns in the abundance and behavior of the tube-building polychaete Diopatra cuprea. Mar. Ecol. Prog. Ser. 2012, 447, 1–13. [Google Scholar] [CrossRef]
- Berke, S. A Review of Diopatra Ecology: Current Knowledge, Open Questions, and Future Threats for an Ecosystem Engineering Polychaete. Biology 2022, 11, 1485. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.S.; Gurgel, C.F.D.; Fredericq, S.; McGlathery, K.J. Gracilaria vermiculophylla (Rhodophyta, Gracilariales) in Hog Island Bay, Virginia: A cryptic alien and invasive macroalga and taxonomic correction. J. Phycol. 2005, 42, 139–141. [Google Scholar] [CrossRef]
- Keller, E.L.; Berke, S.K.; Needham, C.N.; Salerno, C.R. A double-edged sword: Infaunal responses to Agarophyton vermiculophyllum in the mid-Atlantic United States. Estuaries Coasts J. ERF 2019, 42, 1924–1937. [Google Scholar] [CrossRef]
- Kollars, N.M.; Byers, J.E.; Sotka, E.E. Invasive décor: An association between a native decorator worm and a non-native seaweed can be mutualistic. Mar. Ecol. Prog. Ser. 2016, 545, 135–145. [Google Scholar] [CrossRef]
- Mott, A.W.; Krueger-Hadfield, S.A.; Blakeslee, A.M.H.; Fowler, A.E. Native tube-building polychaete prefers to anchor non-native alga over other macrophytes. Oecologia 2022, 198, 967–980. [Google Scholar] [CrossRef]
- Thomsen, M.S.; McGlathery, K. Facilitation of macroalgae by the sedimentary tube forming polychaete Diopatra cuprea. Estuar. Coast. Shelf Sci. 2005, 62, 63–73. [Google Scholar] [CrossRef]
- Pollock, L.W. A Practical Guide to the Marine Animals of Northeastern North America; Rutgers University Press: New Brunswick, NJ, USA; London, UK, 1998; p. 367. Available online: http://books.google.com/books?id=i1AmT31cuR4C (accessed on 15 March 2023).
- Berke, S.K.; Mahon, A.R.; Lima, F.P.; Halanych, K.M.; Wethey, D.S.; Woodin, S.A. Range Shifts and Species Diversity in Marine Ecosystem Engineers: Patterns and Predictions for European Sedimentary Habitats. Glob. Ecol. Biogeogr. 2010, 19, 223–232. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. ape 5.0, an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 15 March 2023).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Archer, F.I.; Adams, P.E.; Schneiders, B.B. strataG: An R package for manipulating, summarizing and analysing population genetic data. Mol. Ecol. Resour. 2016, 17, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Leray, M.; Paulay, G.; Mulcahy, D.G. Diopatra Ornata Voucher BMBM0561 Cytochrome Oxidase Subunit 1 (COI) Gene, Partial cds; Mitochondrial. 2019. Available online: https://www.ncbi.nlm.nih.gov/nuccore/MN138386 (accessed on 15 March 2023).
- Harper, K.E.; Scheinberg, L.A.; Boyer, K.E.; Sotka, E.E. Global distribution of cryptic native, introduced and hybrid lineages in the widespread estuarine amphipod Ampithoe valida. Conserv. Genet. 2022, 23, 791–806. [Google Scholar] [CrossRef]
- Bors, E.K.; Rowden, A.A.; Maas, E.W.; Clark, M.R.; Shank, T.M. Patterns of Deep-Sea Genetic Connectivity in the New Zealand Region: Implications for Management of Benthic Ecosystems. PLoS ONE 2012, 7, e49474. [Google Scholar] [CrossRef]
- Avise, J.C. Molecular Population Structure and the Biogeographic History of a Regional Fauna: A Case History with Lessons for Conservation Biology. Oikos 1992, 63, 62–76. [Google Scholar] [CrossRef]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Soltis, D.E.; Morris, A.B.; McLachlan, J.S.; Manos, P.S.; Soltis, P.S. Comparative phylogeography of unglaciated eastern North America. Mol. Ecol. 2006, 15, 4261–4293. [Google Scholar] [CrossRef] [PubMed]
- Wares, J.P. Community genetics in the Northwestern Atlantic intertidal. Mol. Ecol. 2002, 11, 1131–1144. [Google Scholar] [CrossRef]
- Hilbish, T.J. Population genetics of marine species: The interaction of natural selection and historically differentiated populations. J. Exp. Mar. Biol. Ecol. 1996, 200, 67–83. [Google Scholar] [CrossRef]
- Zerebecki, R.A.; Sotka, E.E.; Hanley, T.C.; Bell, K.L.; Gehring, C.; Nice, C.C.; Richards, C.L.; Hughes, A.R. Repeated Genetic and Adaptive Phenotypic Divergence across Tidal Elevation in a Foundation Plant Species. Am. Nat. 2021, 198, E152–E169. [Google Scholar] [CrossRef]
- Galaska, M.P.; Wethey, D.S.; Arias, A.; Dubois, S.F.; Halanych, K.M.; Woodin, S.A. The Impact of Aquaculture on the Genetics and Distribution of the Onuphid Annelid Diopatra Biscayensis. Ecol. Evol. 2021, 11, 6184–6194. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| ID | Population | Region | Latitude | Longitude | Collector | Total (n) | A1 (n) | A2 (n) | B (n) | C (n) | D (n) | Nucleotide Diversity (mean) | Unique Haplotypes (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FL.ST | Saint Teresa Beach FL | FL-Gulf | 29.922381 | −84.474212 | Bell | 11 | 9 | 1 | 1 | 0 | 0 | 0.052 | 0.545 |
| FL.FP | Fort Pierce FL | FL-Atlantic | 27.4575 | −80.306 | Berke | 7 | 0 | 7 | 0 | 0 | 0 | 0.000 | 0.143 |
| FL.CL | Chicken Island FL | FL-Atlantic | 29.03239 | −80.91479 | Berke | 4 | 0 | 4 | 0 | 0 | 0 | 0.000 | 0.250 |
| FL.AM | Amelia Island Florida | FL-SC-NC-VA | 30.69617 | −81.46038 | Berke | 5 | 0 | 0 | 5 | 0 | 0 | 0.002 | 0.600 |
| SC.CH | Charleston SC | FL-SC-NC-VA | 32.75106 | −79.90281 | Berke | 33 | 0 | 1 | 31 | 0 | 1 | 0.018 | 0.303 |
| SC.BA | North Inlet SC | FL-SC-NC-VA | 33.341 | −79.165 | Berke | 7 | 0 | 0 | 7 | 0 | 0 | 0.000 | 0.143 |
| NC.BE | Beaufort NC | FL-SC-NC-VA | 34.72195 | −76.687 | Berke, Bell | 35 | 1 | 0 | 29 | 4 | 1 | 0.044 | 0.171 |
| VA.WA | Wachapreague VA | FL-SC-NC-VA | 37.601553 | −75.686788 | Berke | 30 | 0 | 1 | 28 | 0 | 1 | 0.020 | 0.167 |
| MA.BA | Barnstable Harbor MA | MA | 41.70911 | −70.30297 | Berke | 14 | 0 | 0 | 0 | 0 | 14 | 0.000 | 0.071 |
| MA.WE | Wellfleet MA | MA | 41.929924 | −70.073089 | Bell | 4 | 0 | 0 | 1 | 0 | 3 | 0.076 | 0.500 |
| MA.DU | Duxbury MA | MA | 42.04647 | −70.65025 | Berke | 3 | 0 | 0 | 1 | 0 | 2 | 0.101 | 0.667 |
| Species | CladeID-Hektoen | Accession Num | Reference |
|---|---|---|---|
| Diopatra_biscayensis | 5 | FJ428837 | [29] |
| Diopatra_ornata | 5 | MN138386 | [37] |
| Diopatra_petiniconicum | 5 | MK690714 | [31] |
| Diopatra_cuprea | 5 | FJ428891 | [29] |
| Diopatra_cuprea | 5 | FJ428894 | [29] |
| Diopatra_victoriae | 5 | MK690689 | [31] |
| Diopatra_hannelorae | 5 | MK690697 | [31] |
| Diopatra_marocensis | 2 | FJ428922 | [29] |
| Diopatra sp11 | 2 | OL874705 | [38] |
| Diopatra_micrura | 1 | GQ456161 | [16] |
| Diopatra_neapolitana | 1 | FJ428866 | [29] |
| Diopatra sp12 | 3 | OL874706 | [6] |
| Diopatra spA | 3 | OL874660 | [6] |
| Diopatra_dubia | 4 | OL874650 | [6] |
| Diopatra sp16 | 4 | OL874712 | [6] |
| Nothria_conchylega | Outgroup | HM473514 | [10] |
| Nothria_conchylega | Outgroup | HQ023895 | [10] |
| Hyalinoecia_tubicola | Outgroup | JX219813 | [39] |
| Hyalinoecia_tubicola | Outgroup | JX219834 | [39] |
| A1 | A2 | B | C | D | |
|---|---|---|---|---|---|
| A1 | - | ||||
| A2 | 0.146 | - | |||
| B | 0.178 | 0.169 | - | ||
| C | 0.167 | 0.169 | 0.151 | - | |
| D | 0.183 | 0.205 | 0.172 | 0.192 | - |
| FL.ST | FL.FP | FL.CL | FL.AM | SC.CH | SC.BA | NC.BE | VA.WA | MA.BA | MA.WE | MA.DU | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| FL.ST | - | 0.002 | 0.007 | 0.002 | 0.002 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.007 |
| FL.FP | 0.721 | - | NA | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.003 | 0.009 |
| FL.CL | 0.669 | NA | - | 0.004 | 0.005 | 0.001 | 0.001 | 0.001 | 0.001 | 0.034 | 0.030 |
| FL.AM | 0.783 | 1.000 | 1.000 | - | 1.000 | 0.341 | 0.356 | 0.284 | 0.001 | 0.028 | 0.085 |
| SC.CH | 0.806 | 1.000 | 1.000 | −0.208 | - | 0.691 | 0.382 | 0.895 | 0.001 | 0.024 | 0.062 |
| SC.BA | 0.825 | 0.899 | 0.890 | −0.095 | −0.068 | - | 0.104 | 0.880 | 0.001 | 0.001 | 0.017 |
| NC.BE | 0.704 | 0.783 | 0.766 | −0.043 | −0.013 | 0.023 | - | 0.200 | 0.001 | 0.001 | 0.021 |
| VA.WA | 0.810 | 0.891 | 0.881 | −0.094 | −0.067 | −0.033 | 0.019 | - | 0.001 | 0.003 | 0.023 |
| MA.BA | 0.879 | 1.000 | 1.000 | 1.000 | 1.000 | 0.922 | 0.822 | 0.914 | - | 0.233 | 0.169 |
| MA.WE | 0.667 | 0.867 | 0.809 | 0.709 | 0.761 | 0.766 | 0.600 | 0.744 | 0.336 | - | 1.000 |
| MA.DU | 0.635 | 0.858 | 0.784 | 0.638 | 0.708 | 0.714 | 0.518 | 0.687 | 0.512 | −0.388 | - |
| FL-Gulf | FL-Atlantic | FL-SC-NC | MA | |
|---|---|---|---|---|
| FL-Gulf | - | 0.001 | 0.001 | 0.001 |
| FL-Atlantic | 0.767 | - | 0.001 | 0.001 |
| FL-SC-NC-VA | 0.812 | 0.855 | - | 0.001 |
| MA | 0.794 | 0.911 | 0.825 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sotka, E.E.; Bell, T.; Berke, S. Cryptic mtDNA Diversity of Diopatra cuprea (Onuphidae, Annelida) in the Northwestern Atlantic Ocean. Biology 2023, 12, 521. https://doi.org/10.3390/biology12040521
Sotka EE, Bell T, Berke S. Cryptic mtDNA Diversity of Diopatra cuprea (Onuphidae, Annelida) in the Northwestern Atlantic Ocean. Biology. 2023; 12(4):521. https://doi.org/10.3390/biology12040521
Chicago/Turabian StyleSotka, Erik E., Tina Bell, and Sarah Berke. 2023. "Cryptic mtDNA Diversity of Diopatra cuprea (Onuphidae, Annelida) in the Northwestern Atlantic Ocean" Biology 12, no. 4: 521. https://doi.org/10.3390/biology12040521
APA StyleSotka, E. E., Bell, T., & Berke, S. (2023). Cryptic mtDNA Diversity of Diopatra cuprea (Onuphidae, Annelida) in the Northwestern Atlantic Ocean. Biology, 12(4), 521. https://doi.org/10.3390/biology12040521