

Live to Die Another Day: Regeneration in Diopatra aciculata Knox and Cameron, 1971 (Annelida: Onuphidae) Collected as Bait in Knysna Estuary, South Africa
Simple Summary
Abstract
1. Introduction
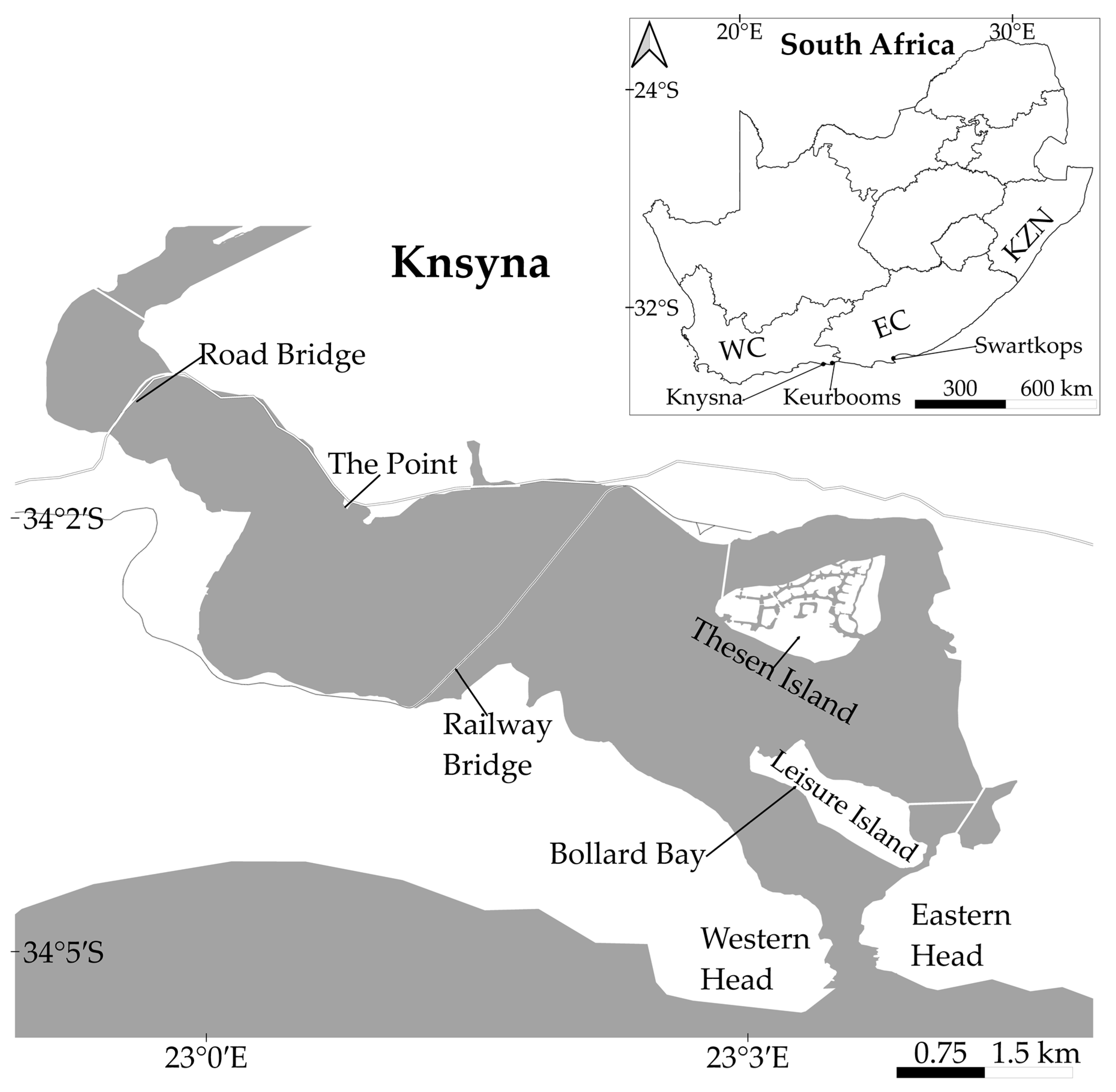
2. Materials and Methods
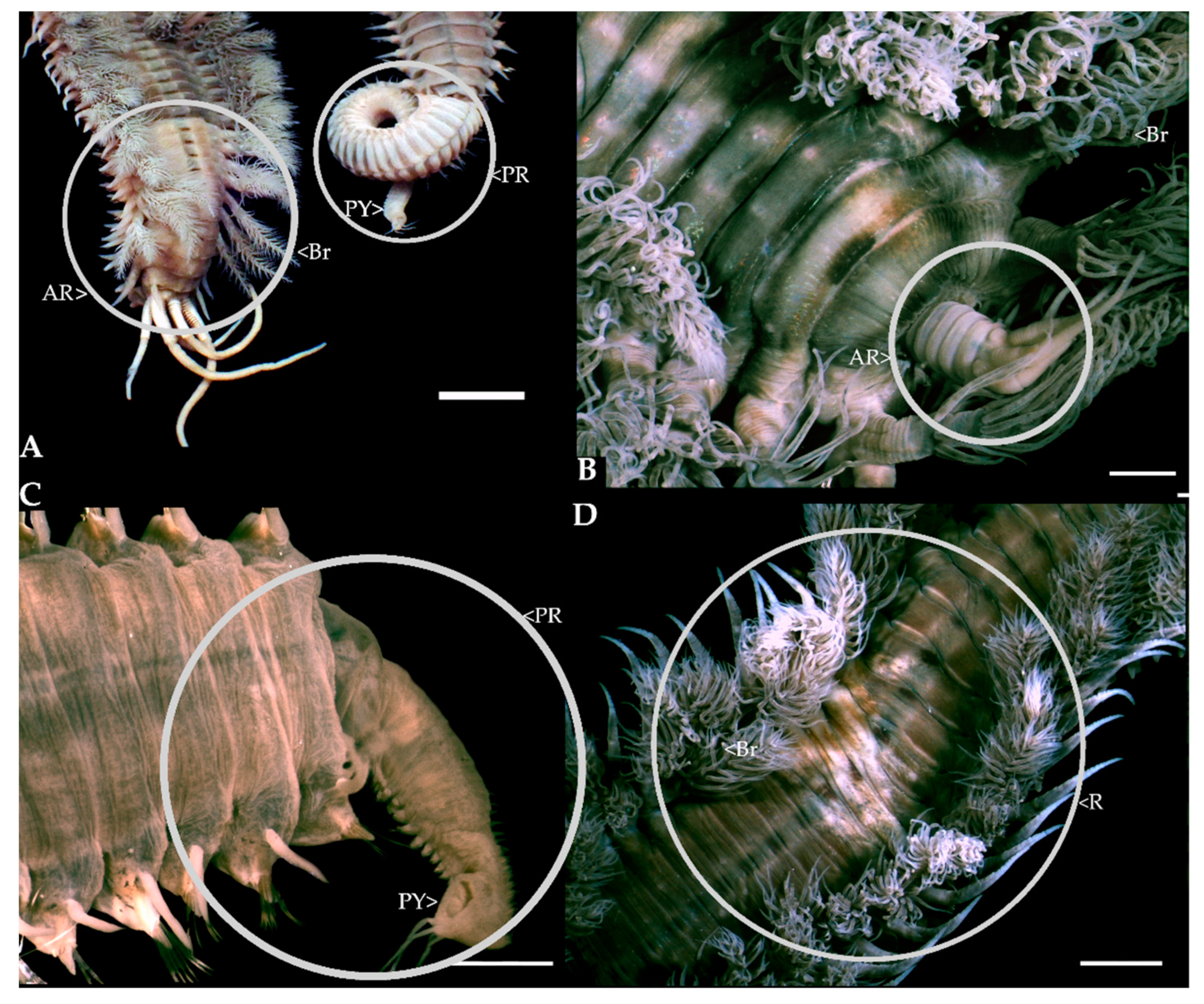
2.1. In Situ Regeneration
2.2. Interviews
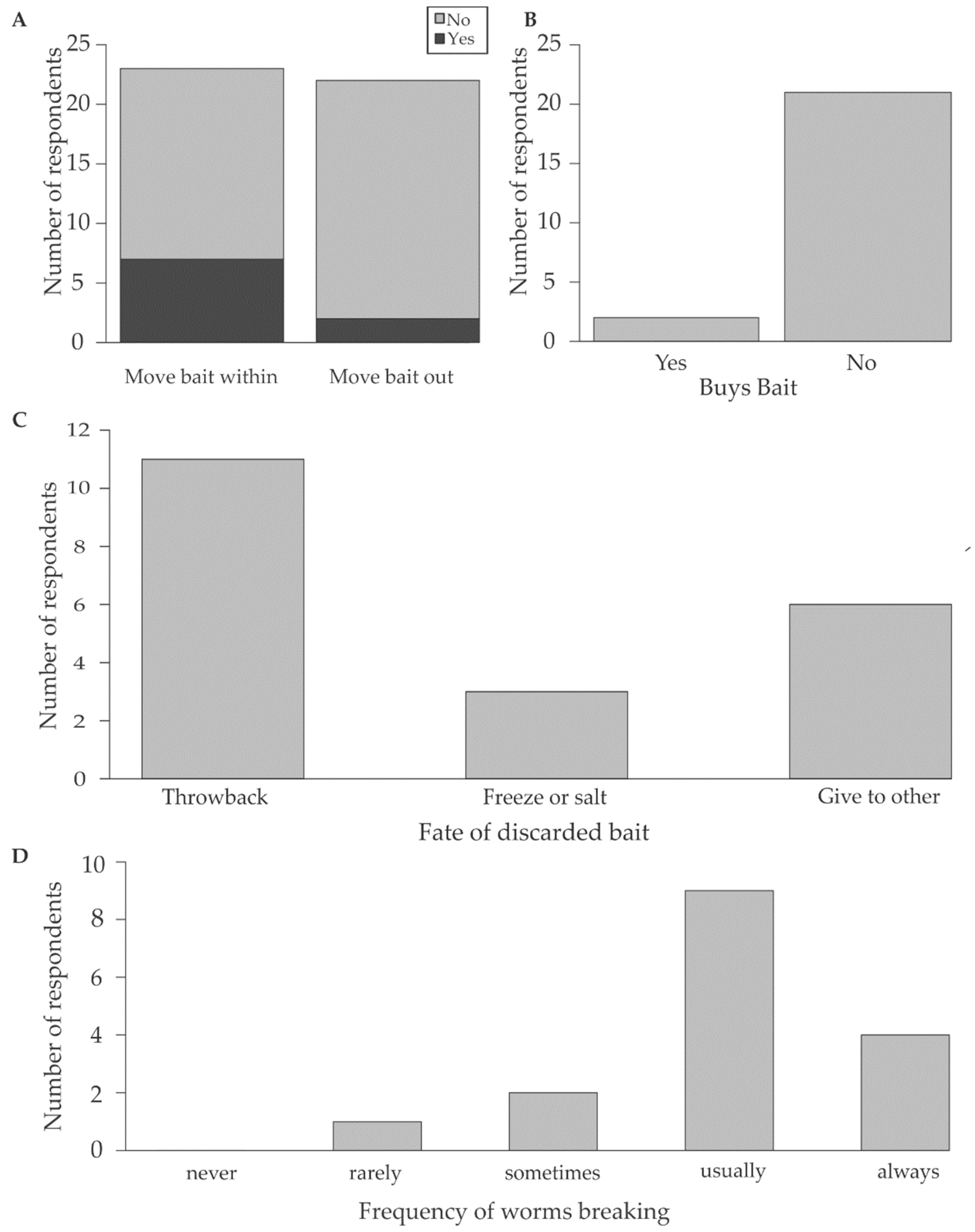
- To identify the fishermen who use D. aciculata, fishermen were asked to list their preferred bait species; only responses from those that selected D. aciculata were retained for analysis (Figure 2, Q1).
- To estimate the magnitude of potential for the regeneration of D. aciculata, fishermen were asked how many worms they collected (Figure 2, Q2).
- To assess if regeneration could lead to dispersal, respondents were asked if they moved bait within and out of the Knysna Estuary (Figure 2, Q3a), as this creates an opportunity for anthropogenic dispersal. Secondly, respondents were asked if they bought Diopatra (Figure 2, Q3b), because recreational fishermen tend to purchase bait from subsistence fishermen. As recreational fishermen tend to fish in areas away from the subsistence fisherman (i.e., from whom bait is purchased), the likelihood of anthropogenic dispersal also increases if Diopatra are purchased as bait [49]. Furthermore, many recreational fishermen fish outside of Knysna [49].
- To assess the extent to which discarding unused bait could contribute to dispersal and to maintaining population size despite harvesting, fishermen were asked if they had bait left over and if yes, how the leftover bait was processed or discarded (Figure 2, Q4). In the latter instance, we only considered the discarding of fresh, unprocessed, bait. If large enough pieces of Diopatra are thrown back (size gleaned from observational data), a potential for regeneration is created. Once the worm fragments settle and regenerate fully, naturalisation is possible.
- The fishermen were asked which portion of the worm they preferred as bait (head, middle, tail, or whole worm), together with the frequency with which D. aciculata broke during collection (never 0%, rarely 0–33%, sometimes 33–66%, usually 66–99%, always 100%) (Figure 2, Q5a and 5b). This information was used in conjunction with observations of in situ regeneration to explore if broken pieces of worm that are left behind during bait collection could regenerate and contribute to population growth or maintenance. The assumption was that if fishermen predominantly collected the portion of the worm that they preferred to use as bait, this section would predominantly be leftover and discarded, and these sections would therefore have the greatest potential to survive and, if large enough, regenerate. Additionally, the section of the worm left in the tube (i.e., usually the posterior) could also regenerate if large enough. If both anterior and posterior regeneration is possible, both portions can regenerate leading to incidental asexual reproduction.
2.3. Statistical Analysis
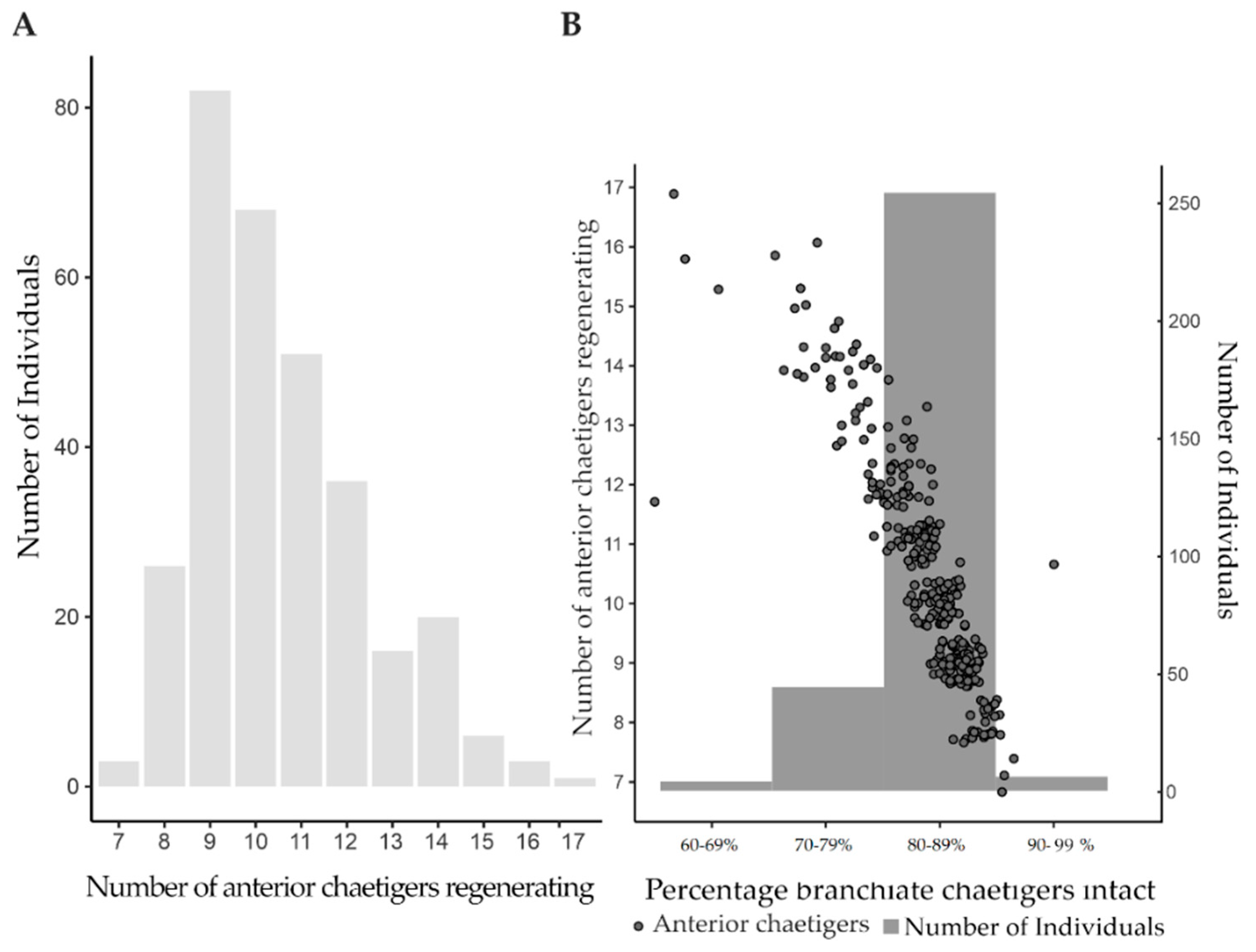
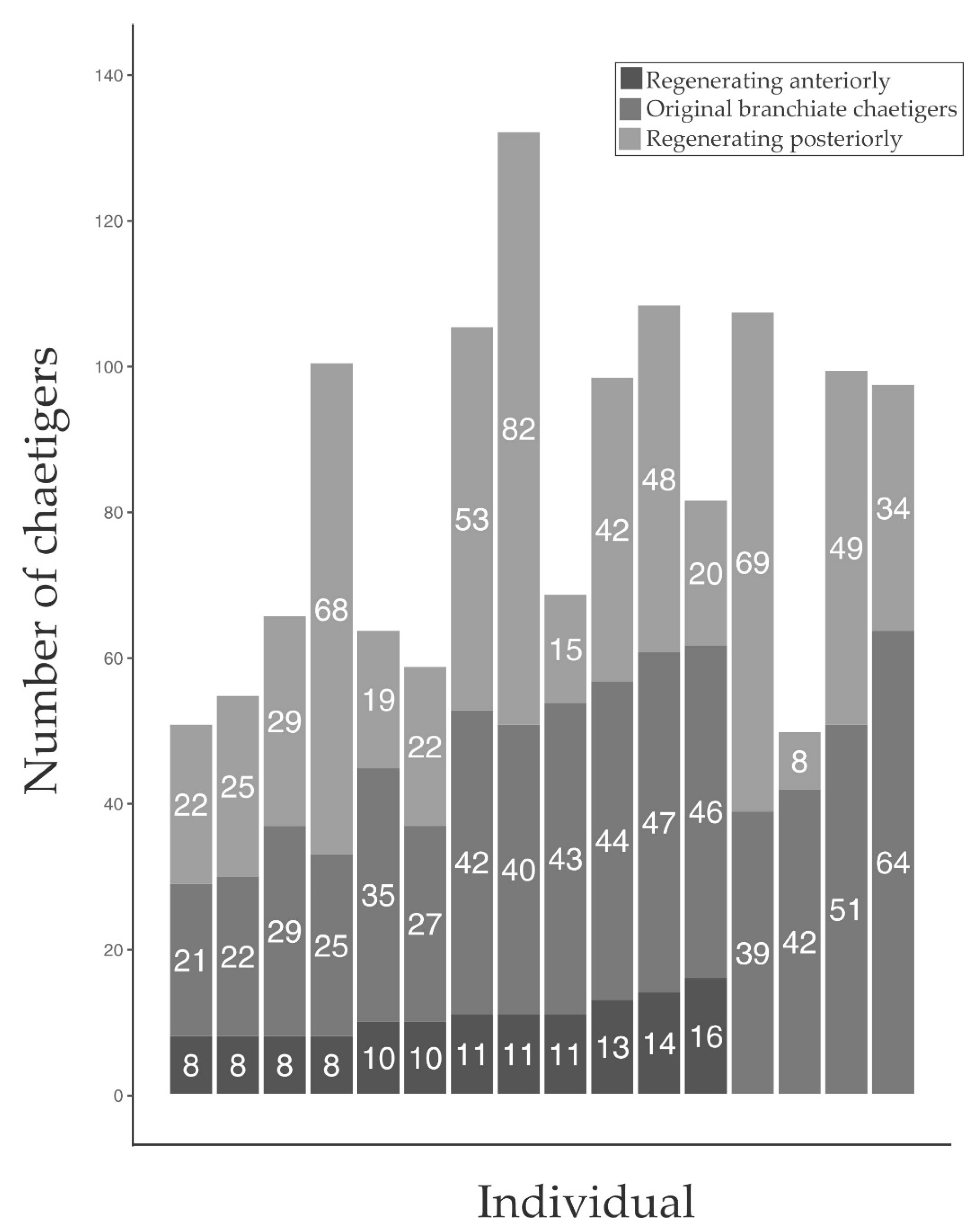
2.3.1. In Situ Regeneration
2.3.2. Interviews
3. Results
3.1. In Situ Regeneration
3.2. Fishermen Baiting Habit Survey
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cole, V.J.; Chick, R.C.; Hutchings, P.A. A Review of Global Fisheries for Polychaete Worms as a Resource for Recreational Fishers: Diversity, Sustainability and Research Needs. Rev. Fish Biol. Fish. 2018, 28, 543–565. [Google Scholar] [CrossRef]
- Glasby, C.J.; Hutchings, P.A.; Fauchald, K.; Paxton, H.; Rouse, G.W.; Russell, C.W.; Wilson, R.S. Class Polychaeta. In Fauna of Australia—Volume 4A Polychaetes & Allies; Beesley, P.L., Ross, G.J.B., Glasby, C.J., Eds.; Australian Biological Resources Study, CSIRO Publishing: Clayton, VIC, Australia, 2000; Volume 4A, pp. 4–372. [Google Scholar]
- Olive, P.J.W. Polychaeta as a World Resource: A Review of Patterns of Exploitation as Sea Angling Baits, and the Potential for Aquaculture Based Production. Mém. Mus. Natl. Hist. Nat. 1994, 162, 603–610. [Google Scholar]
- Van Der Have, T.; Broeckx, P.; Kersbergen, A. Risk Assessment of Live Bait: Alien Species in Live Bait Used by Anglers in the Netherlands; Nederlandse Voedsel- en Warenautoriteit: Wageningen, The Netherlands, 2015. [Google Scholar]
- Saito, H.; Kawai, K.; Umino, T.; Imabayashi, H. Fishing Bait Worm Supplies in Japan in Relation to Their Physiological Traits. Mem. Mus. Vic. 2014, 71, 279–287. [Google Scholar] [CrossRef]
- Safarik, M.; Redden, A.M.; Schreider, M.J. Density-Dependent Growth of the Polychaete Diopatra aciculata. Sci. Mar. 2006, 70, 337–341. [Google Scholar] [CrossRef]
- Gambi, M.C.; Castelli, A.; Giangrande, A.; Lanera, P.; Prevedelli, D.; Zunarelli Vandini, R. Polychaetes of Commercial and Applied Interest in Italy: An Overview. Mém. Mus. Natl. Hist. Nat. 1994, 162, 593–601. [Google Scholar]
- Paxton, H. Diopatra Audouin and Milne Edwards (Polychaeta: Onuphidae) from Australia, with a Discussion of Developmental Patterns in the Genus. Beagle 1993, 10, 115–154. [Google Scholar] [CrossRef]
- Paxton, H. Generic Revision and Relationships of the Family Onuphidae (Annelida: Polychaeta). Rec. Aust. Mus. 1986, 38, 1–74. [Google Scholar] [CrossRef]
- Paxton, H. Revision of the Rhamphobrachium Complex (Polychaeta: Onuphidae). Rec. Aust. Mus. 1986, 38, 75–104. [Google Scholar] [CrossRef][Green Version]
- Paxton, H. Taxonomy and Aspects of the Life History of Australian Beachworms (Polychaeta: Onuphidae). Mar. Freshw. Res. 1979, 30, 265–294. [Google Scholar] [CrossRef]
- Audouin, J.V.V.; Edwards, H.M.; Milne Edwards, H. Classification des Annélides et Description de Celles qui Habitent les Côtes de la France. Ann. Sci. Nat. Paris. Sér. 1833, 1, 187–247. [Google Scholar] [CrossRef]
- Watson, G.J.; Murray, J.M.; Schaefer, M.; Bonner, A. Bait Worms: A Valuable and Important Fishery with Implications for Fisheries and Conservation Management. Fish Fish. 2017, 18, 374–388. [Google Scholar] [CrossRef]
- Daǧli, E.; Ergen, Z.; Çinar, M.E. One-Year Observation on the Population Structure of Diopatra neapolitana Delle Chiaje (Polychaeta: Onuphidae) in Izmir Bay (Aegean Sea, Eastern Mediterranean). Mar. Ecol. 2005, 26, 265–272. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H. The Cryptogenic Bait Worm Diopatra biscayensis Fauchald et. al., 2012 (Annelida: Onuphidae)—Revisiting Its History, Biology and Ecology. Estuar. Coast Shelf Sci. 2015, 163, 22–36. [Google Scholar] [CrossRef]
- Pires, A.; Quintino, V.; Gentil, F.; Freitas, R.; Rodrigues, A.M. Reproductive Biology of a Brooding Diopatra Species: Diopatra marocensis Paxton et. al., 1995. Estuar. Coast Shelf Sci. 2012, 110, 85–92. [Google Scholar] [CrossRef]
- Fauchald, K.; Berke, S.K.; Woodin, S.A. Diopatra (Onuphidae: Polychaeta) from intertidal sediments in southwestern Europe. Zootaxa 2012, 3395, 47–58. [Google Scholar] [CrossRef]
- Knox, G.A.; Cameron, D.B. Port Phillip Bay Survey Part 2. Polychaeta. Mem. Mus. Vic. 1971, 32, 21–41. [Google Scholar] [CrossRef]
- Paxton, H. Diopatra Audouin and Milne Edwards (Polychaeta: Onuphidae) From Thailand. Phuket Mar. Biol. Cent. Spec. Publ. 2002, 24, 101–114. [Google Scholar] [CrossRef]
- Elgetany, A.H.; van Rensburg, H.; Hektoen, M.; Matthee, C.; Budaeva, N.; Simon, C.A.; Struck, T.H. Species Delineation in the Speciation Grey Zone—The Case of Diopatra (Annelida, Onuphidae). Zool. Scr. 2020, 49, 516–534. [Google Scholar] [CrossRef]
- Van Rensburg, H.; Matthee, C.A.; Simon, C.A. Moonshine Worms (Diopatra aciculata: Onuphidae, Annelida) in the Knysna Estuary, South Africa; Taxonomy and Distribution. J. Mar. Biol. Assoc. 2020, 100, 897–907. [Google Scholar] [CrossRef]
- Simon, C.; Kara, J.; du Toit, A.; van Rensburg, H.; Naidoo, C.; Matthee, C.A. Reeling Them in: Taxonomy of Marine Annelids Used as Bait by Anglers in the Western Cape Province, South Africa. PeerJ 2021, 9, e11847. [Google Scholar] [CrossRef]
- Simon, C.A.; Du Toit, A.N.; Lamberth, S.J.; Branch, G.M. Standardising English and Afrikaans Common Names for Polychaetes Harvested as Bait in South Africa. Afr. Zool. 2022, 57, 75–89. [Google Scholar] [CrossRef]
- Day, J.H. A Guide to Marine Life on South African Shores; Balkema: Cape Town, South Africa, 1969. [Google Scholar]
- Macnae, W. The Ecology of Plants and Animals in the Intertidal Regions of the Zwartkops Estuary, Near Port Elizabeth, South Africa. J. Ecol. 1957, 45, 361. [Google Scholar] [CrossRef]
- Day, J.H.; Millard, N.A.H.; Harrison, A.D. The Ecology of South African Estuaries. Trans. R. Soc. S. Afr. 1951, 33, 367–413. [Google Scholar] [CrossRef]
- Heydorn, A.E.F.; Grindley, J.R.; Morant, P.D. Estuaries of the Cape. In Part II, Synopses of Available Information on Individual Systems; National Research Institute for Oceanology, Council for Scientific and Industrial Research: Stellenbosch, South Africa, 1980. [Google Scholar]
- Fielding, P.J. The Subsistence Bait Fishery in the Swartkops Estuary, Port Elizabeth, Eastern Cape; Zwartkops Conservancy: Nelson Mandela Bay Municipality, South Africa, 2007. [Google Scholar]
- Van der Westhuizen, H.C.; Marais, J.F.K. Stomach Content Analyses of Pomadasys commersonni from the Swartkops Estuary (Pisces: Pomadasyidae). Afr. Zool. 1977, 12, 500–504. [Google Scholar] [CrossRef]
- Napier, V.R.; Turpie, J.K.; Clark, B.M. Value and Management of the Subsistence Fishery at Knysna Estuary, South Africa. Afr. J. Mar. Sci. 2009, 31, 297–310. [Google Scholar] [CrossRef]
- Simon, C.; Du Toit, A.N.; Smith, M.K.S.; Claassens, L.; Smith, F.; Smith, P. Bait Collecting by Subsistence and Recreational Fishers in Knysna Estuary May Impact Management and Conservation. Afr. Zool. 2019, 54, 91–103. [Google Scholar] [CrossRef]
- Zajac, R.N. the Effects of Sublethal Predation on Reproduction in the Spionid Polychaete Polydora ligni Webster. J. Exp. Mar. Biol. Ecol. 1985, 88, 1–19. [Google Scholar] [CrossRef]
- Hentschel, B.T.; Harper, N.S. Effects of Simulated Sublethal Predation on the Growth and Regeneration Rates of a Spionid Polychaete in Laboratory Flumes. Mar. Biol. 2006, 149, 1175–1183. [Google Scholar] [CrossRef]
- Lindsay, S.; Jackson, J.L.; He, S. Anterior Regeneration in the Spionid Polychaetes Dipolydora quadrilobata and Pygospio elegans. Mar. Biol. 2007, 150, 1161–1172. [Google Scholar] [CrossRef]
- Berke, S.K.; Cruz, V.; Osman, R.W. Sublethal Predation and Regeneration in Two Onuphid Polychaetes: Patterns and Implications. Biol. Bull. 2009, 217, 242–252. [Google Scholar] [CrossRef]
- Pires, A.; Freitas, R.; Quintino, V.; Rodrigues, A.M. Can Diopatra neapolitana (Annelida: Onuphidae) Regenerate Body Damage Caused by Bait Digging or Predation? Estuar. Coast Shelf Sci. 2012, 110, 36–42. [Google Scholar] [CrossRef]
- Bely, A.E. Distribution of Segment Regeneration Ability in the Annelida. Integr. Comp. Biol. 2006, 46, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Rouse, G.W. Annelida (Segmented Worms). In eLS; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2002. [Google Scholar] [CrossRef]
- Kostyuchenko, R.P.; Kozin, V.V. Comparative Aspects of Annelid Regeneration: Towards Understanding the Mechanisms of Regeneration. Genes 2021, 12, 1148. [Google Scholar] [CrossRef] [PubMed]
- Balavoine, G. Segment Formation in Annelids: Patterns, Processes and Evolution. Int. J. Dev. Biol. 2014, 58, 469–483. [Google Scholar] [CrossRef]
- Gazave, E.; Béhague, J.; Laplane, L.; Guillou, A.; Préau, L.; Demilly, A.; Balavoine, G.; Vervoort, M. Posterior Elongation in the Annelid Platynereis dumerilii Involves Stem Cells Molecularly Related to Primordial Germ Cells. Dev. Biol. 2013, 382, 246–267. [Google Scholar] [CrossRef]
- Gibson, G.D.; Harvey, J.M.L. Morphogenesis During Asexual Reproduction in Pygospio elegans Claparede (Annelida, Polychaeta). Biol. Bull. 2000, 199, 41–49. [Google Scholar] [CrossRef]
- David, A.A.; Williams, J.D.; Simon, C.A. A New Record of a Cryptogenic Dipolydora Species (Annelida: Spionidae) in South Africa. J. Mar. Biol. Assoc. 2021, 101, 271–278. [Google Scholar] [CrossRef]
- Pires, A.; Paxton, H.; Quintino, V.; Rodrigues, A.M. Diopatra (Annelida: Onuphidae) Diversity in European Waters with the Description of Diopatra micrura, New Species. Zootaxa 2010, 2395, 17. [Google Scholar] [CrossRef]
- Otegui, M.B.P.; Brauko, K.M.; Pagliosa, P.R. Matching Ecological Functioning with Polychaete Morphology: Consistency Patterns along Sedimentary Habitats. J. Sea Res. 2016, 114, 13–21. [Google Scholar] [CrossRef]
- Schoeman, S.; (Stellenbosch University, Stellenbosch, South Africa). Personal Communication, 2021.
- Van Rensburg, H. (Stellenbosch University, Stellenbosch, South Africa). Unpublished Data, 2018.
- Hodgson, A.N.; Allanson, B.R.; Cretchley, R.; Allanson, B.R. The Exploitation of Upogebia africana (Crustacea: Thalassinidae) for Bait in the Knysna Estuary. Trans. R. Soc. S. Afr. 2000, 55, 197–204. [Google Scholar] [CrossRef]
- Smit, M.K.; (SANParks Garden Route National Park, Rondevlei Office, Sedgefield, South Africa). Personal Communication, 2022.
- SANParks Garden Route National Park, Knysna, South Africa. Unpublished Report, 2022.
- Kilian, J.V.; Klauda, R.J.; Widman, S.; Kashiwagi, M.; Bourquin, R.; Weglein, S.; Schuster, J. An Assessment of a Bait Industry and Angler Behavior as a Vector of Invasive Species. Biol. Invasions 2012, 14, 1469–1481. [Google Scholar] [CrossRef]
- Whitfield, A.K. A Characterization of Southern African Estuarine Systems. S. Afr. J. Aquat. Sci. 1992, 18, 89–103. [Google Scholar] [CrossRef]
- Barnes, R.S.K. Patterns of Seagrass Macrobenthic Biodiversity in the Warm-Temperate Knysna Estuarine Bay, Western Cape: A Review. Aquat. Ecol. 2021, 55, 327–345. [Google Scholar] [CrossRef]
- Largier, J.L.; Attwood, C.; Harcourt-Baldwin, J.L. The Hydrographic Character of the Knysna Estuary. Trans. R. Soc. S. Afr. 2000, 55, 107–122. [Google Scholar] [CrossRef]
- Perez-Hurtado, A.; Goss-Custard, J.D.; Garcia, F. The Diet of Wintering Waders in Cádiz Bay, Southwest Spain. Bird Study 2010, 44, 45–52. [Google Scholar] [CrossRef]
- Bailey-Brock, J.H. Ecology of the Tube-Building Polychaete Diopatra leuckarti Kinberg, 1865 (Onuphidae) in Hawaii: Community Structure, and Sediment Stabilizing Properties. Zool. J. Linn. Soc. 1984, 80, 191–199. [Google Scholar] [CrossRef]
- Rouse, G.; Pleijel, F. Polychaetes; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Department of Agriculture Forestries and Fisheries, (DAFF). Marine Recreational Activity Information Brochure. 2017. Available online: https://www.treedomvillas.co.za/portals/0/articles/RecreationalFishingBrochure2017-18.pdf (accessed on 23 November 2022).
- Schoeman, S.; Simon, A.C.; Van Wyk, H. (Stellenbosch University, Stellenbosch, South Africa). Manuscript in preparation, 2023.
- Anon. Socio-Economic Profile: Knysna Municipality, Knysna, South Africa. 2021. Available online: https://www.knysna.gov.za/wp-content/uploads/2022/03/2021-SEP-LG-Knysna-Municipality.pdf (accessed on 20 November 2022).
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.; Richardson, D.M. A Proposed Unified Framework for Biological Invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Percentage Preferring Diopatra | Number of Bait Collection Opportunities per Year | Number Using Diopatra (2) | Number Caught per Year (3) | Number Discarded per Year (4) | Number of Worms Breaking per Year (5) | Number Capable of Regeneration (6) |
|---|---|---|---|---|---|---|
| 12% * | 19,954 | 2394 | 23,945 | 11,972 | 15,804–23,705 | 1796 |
| 32% ** | 19,954 | 6385 | 63,853 | 31,926 | 42,143–63,214, | 4789 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schoeman, S.; Simon, C.A. Live to Die Another Day: Regeneration in Diopatra aciculata Knox and Cameron, 1971 (Annelida: Onuphidae) Collected as Bait in Knysna Estuary, South Africa. Biology 2023, 12, 483. https://doi.org/10.3390/biology12030483
Schoeman S, Simon CA. Live to Die Another Day: Regeneration in Diopatra aciculata Knox and Cameron, 1971 (Annelida: Onuphidae) Collected as Bait in Knysna Estuary, South Africa. Biology. 2023; 12(3):483. https://doi.org/10.3390/biology12030483
Chicago/Turabian StyleSchoeman, Stephanie, and Carol A. Simon. 2023. "Live to Die Another Day: Regeneration in Diopatra aciculata Knox and Cameron, 1971 (Annelida: Onuphidae) Collected as Bait in Knysna Estuary, South Africa" Biology 12, no. 3: 483. https://doi.org/10.3390/biology12030483
APA StyleSchoeman, S., & Simon, C. A. (2023). Live to Die Another Day: Regeneration in Diopatra aciculata Knox and Cameron, 1971 (Annelida: Onuphidae) Collected as Bait in Knysna Estuary, South Africa. Biology, 12(3), 483. https://doi.org/10.3390/biology12030483