
Inputs and Outputs of the Mammalian Circadian Clock
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
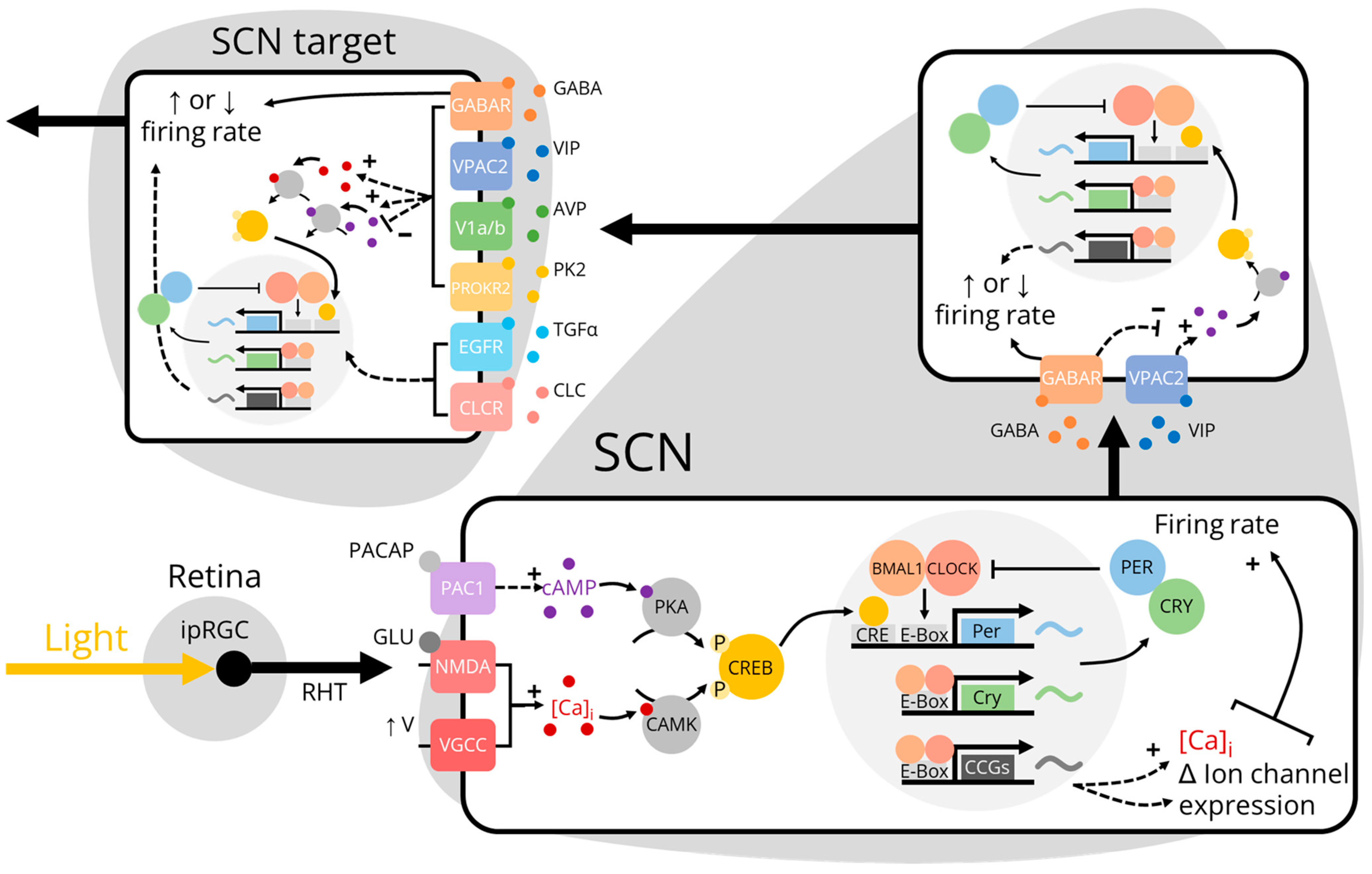
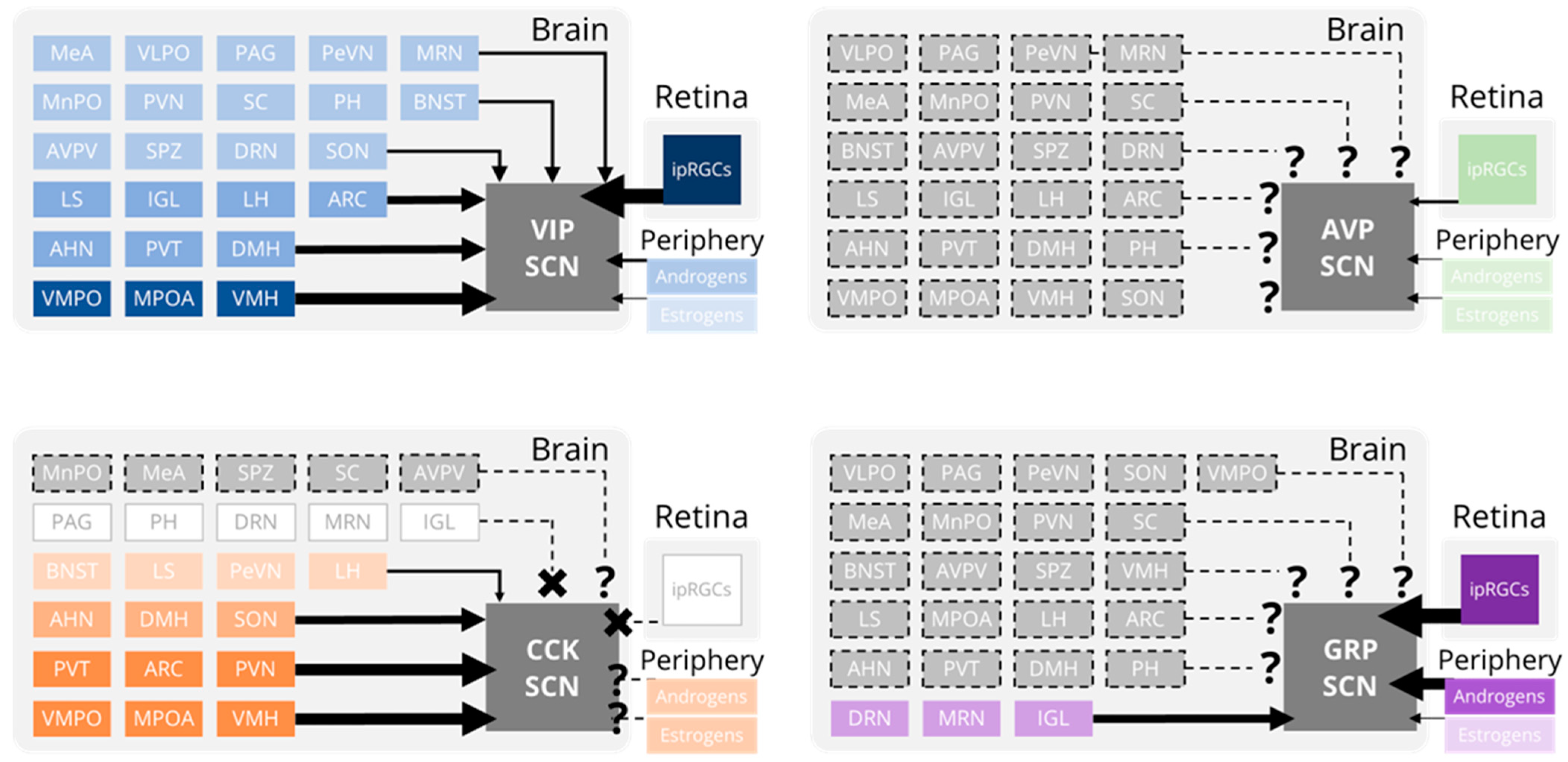
2. Inputs to the SCN: Afferent Projections
3. Inputs to the SCN: Hormones
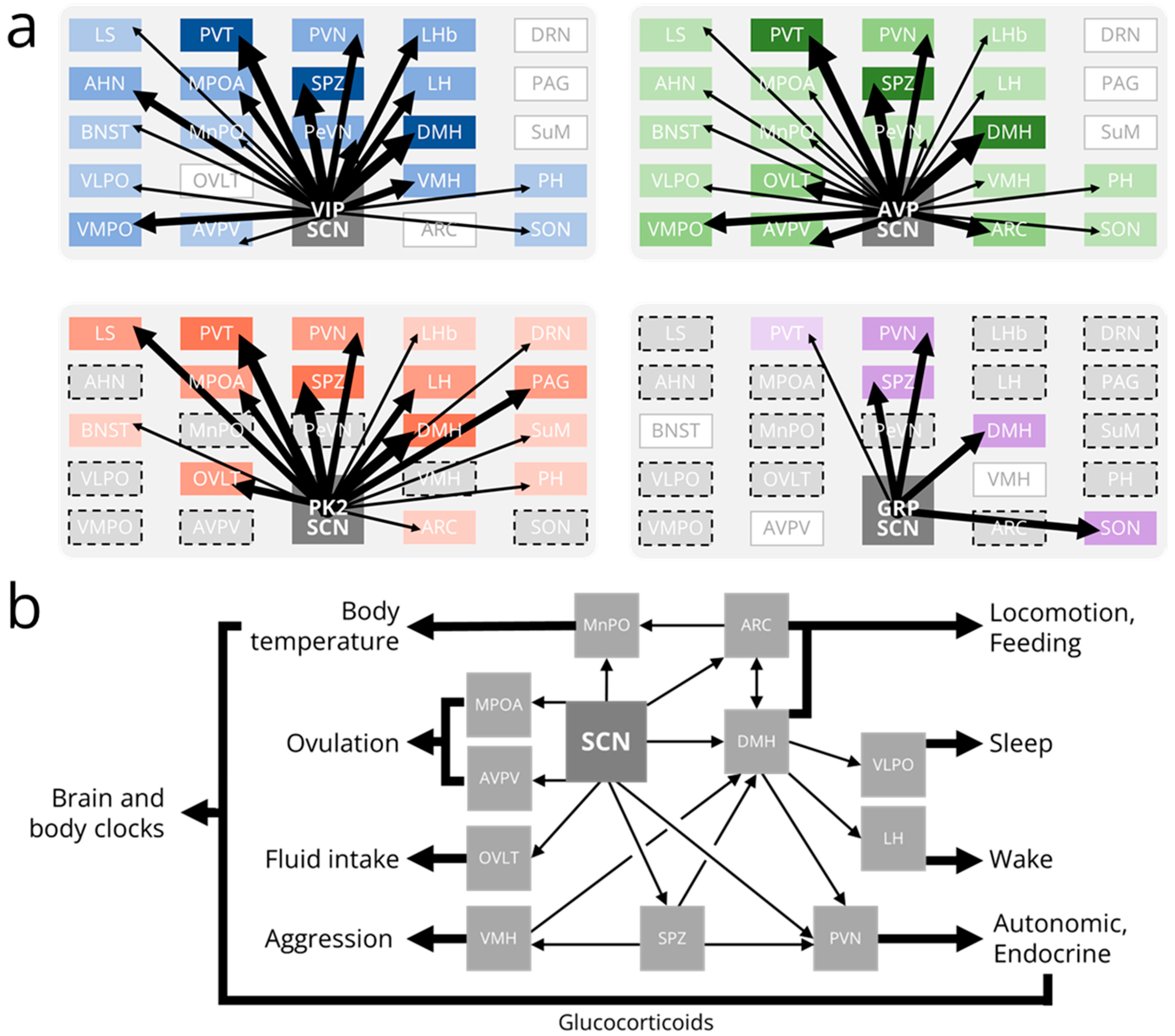
4. Outputs from the SCN: Efferent Projections
5. Outputs from the SCN: Physiological Rhythms
6. Outputs from the SCN: Behavioral Rhythms
7. Outputs from the SCN: Non-Synaptic Signaling
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Silver, R.; Rainbow, M. The Suprachiasmatic Nucleus and the Circadian Timekeeping System of the Body. In Neuroscience in the 21st Century: From Basic to Clinical; Pfaff, D.W., Ed.; Springer: New York, NY, USA, 2013; pp. 1847–1888. ISBN 9781461419976. [Google Scholar]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. Generation of Circadian Rhythms in the Suprachiasmatic Nucleus. Nat. Rev. Neurosci. 2018, 19, 453–469. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.Y.; Speh, J.C. GABA Is the Principal Neurotransmitter of the Circadian System. Neurosci. Lett. 1993, 150, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Van den Pol, A.N. The Hypothalamic Suprachiasmatic Nucleus of Rat: Intrinsic Anatomy. J. Comp. Neurol. 1980, 191, 661–702. [Google Scholar] [CrossRef] [PubMed]
- Ono, D.; Honma, K.-I.; Honma, S. Roles of Neuropeptides, VIP and AVP, in the Mammalian Central Circadian Clock. Front. Neurosci. 2021, 15, 650154. [Google Scholar] [CrossRef] [PubMed]
- Abrahamson, E.E.; Moore, R.Y. Suprachiasmatic Nucleus in the Mouse: Retinal Innervation, Intrinsic Organization and Efferent Projections. Brain Res. 2001, 916, 172–191. [Google Scholar] [CrossRef] [PubMed]
- Kofuji, P.; Mure, L.S.; Massman, L.J.; Purrier, N.; Panda, S.; Engeland, W.C. Intrinsically Photosensitive Retinal Ganglion Cells (ipRGCs) Are Necessary for Light Entrainment of Peripheral Clocks. PLoS ONE 2016, 11, e0168651. [Google Scholar] [CrossRef]
- Schwartz, W.J.; Tavakoli-Nezhad, M.; Lambert, C.M.; Weaver, D.R.; de la Iglesia, H.O. Distinct Patterns of Period Gene Expression in the Suprachiasmatic Nucleus Underlie Circadian Clock Photoentrainment by Advances or Delays. Proc. Natl. Acad. Sci. USA 2011, 108, 17219–17224. [Google Scholar] [CrossRef]
- Lindberg, P.T.; Mitchell, J.W.; Burgoon, P.W.; Beaulé, C.; Weihe, E.; Schäfer, M.K.-H.; Eiden, L.E.; Jiang, S.Z.; Gillette, M.U. Pituitary Adenylate Cyclase-Activating Peptide (PACAP)-Glutamate Co-Transmission Drives Circadian Phase-Advancing Responses to Intrinsically Photosensitive Retinal Ganglion Cell Projections by Suprachiasmatic Nucleus. Front. Neurosci. 2019, 13, 1281. [Google Scholar] [CrossRef]
- Webb, I.C.; Coolen, L.M.; Lehman, M.N. NMDA and PACAP Receptor Signaling Interact to Mediate Retinal-Induced Scn Cellular Rhythmicity in the Absence of Light. PLoS ONE 2013, 8, e76365. [Google Scholar] [CrossRef]
- Irwin, R.P.; Allen, C.N. Calcium Response to Retinohypothalamic Tract Synaptic Transmission in Suprachiasmatic Nucleus Neurons. J. Neurosci. 2007, 27, 11748–11757. [Google Scholar] [CrossRef]
- Chen, D.; Buchanan, G.F.; Ding, J.M.; Hannibal, J.; Gillette, M.U. Pituitary Adenylyl Cyclase-Activating Peptide: A Pivotal Modulator of Glutamatergic Regulation of the Suprachiasmatic Circadian Clock. Proc. Natl. Acad. Sci. USA 1999, 96, 13468–13473. [Google Scholar] [CrossRef] [PubMed]
- Tischkau, S.A.; Mitchell, J.W.; Tyan, S.-H.; Buchanan, G.F.; Gillette, M.U. Ca2+/cAMP Response Element-Binding Protein (CREB)-Dependent Activation of Per1 Is Required for Light-Induced Signaling in the Suprachiasmatic Nucleus Circadian Clock. J. Biol. Chem. 2003, 278, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Roblero, R.; Mercado, C.; Alamilla, J.; Laville, A.; Díaz-Muñoz, M. Ryanodine Receptor Ca2+-Release Channels Are an Output Pathway for the Circadian Clock in the Rat Suprachiasmatic Nuclei. Eur. J. Neurosci. 2007, 26, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Hermanstyne, T.O.; Yang, N.-D.; Granados-Fuentes, D.; Li, X.; Mellor, R.; Jegla, T.; Herzog, E.; Nerbonne, J.M. Kv12-Encoded K+ Channels Drive the Day-Night Switch in the Repetitive Firing Rates of SCN Neurons. bioRxiv 2023, 2023.01.30.526323. [Google Scholar]
- Flourakis, M.; Kula-Eversole, E.; Hutchison, A.L.; Han, T.H.; Aranda, K.; Moose, D.L.; White, K.P.; Dinner, A.R.; Lear, B.C.; Ren, D.; et al. A Conserved Bicycle Model for Circadian Clock Control of Membrane Excitability. Cell 2015, 162, 836–848. [Google Scholar] [CrossRef]
- Fan, J.; Zeng, H.; Olson, D.P.; Huber, K.M.; Gibson, J.R.; Takahashi, J.S. Vasoactive Intestinal Polypeptide (VIP)-Expressing Neurons in the Suprachiasmatic Nucleus Provide Sparse GABAergic Outputs to Local Neurons with Circadian Regulation Occurring Distal to the Opening of Postsynaptic GABAA Ionotropic Receptors. J. Neurosci. 2015, 35, 1905–1920. [Google Scholar] [CrossRef]
- Itri, J.; Colwell, C.S. Regulation of Inhibitory Synaptic Transmission by Vasoactive Intestinal Peptide (VIP) in the Mouse Suprachiasmatic Nucleus. J. Neurophysiol. 2003, 90, 1589–1597. [Google Scholar] [CrossRef]
- Aton, S.J.; Huettner, J.E.; Straume, M.; Herzog, E.D. GABA and Gi/o Differentially Control Circadian Rhythms and Synchrony in Clock Neurons. Proc. Natl. Acad. Sci. USA 2006, 103, 19188–19193. [Google Scholar] [CrossRef]
- Pakhotin, P.; Harmar, A.J.; Verkhratsky, A.; Piggins, H. VIP Receptors Control Excitability of Suprachiasmatic Nuclei Neurones. Pflugers Arch. 2006, 452, 7–15. [Google Scholar] [CrossRef]
- Patton, A.P.; Edwards, M.D.; Smyllie, N.J.; Hamnett, R.; Chesham, J.E.; Brancaccio, M.; Maywood, E.S.; Hastings, M.H. The VIP-VPAC2 Neuropeptidergic Axis Is a Cellular Pacemaking Hub of the Suprachiasmatic Nucleus Circadian Circuit. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef]
- Cheng, M.Y.; Bullock, C.M.; Li, C.; Lee, A.G.; Bermak, J.C.; Belluzzi, J.; Weaver, D.R.; Leslie, F.M.; Zhou, Q.-Y. Prokineticin 2 Transmits the Behavioural Circadian Rhythm of the Suprachiasmatic Nucleus. Nature 2002, 417, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Kraves, S.; Weitz, C.J. A Role for Cardiotrophin-like Cytokine in the Circadian Control of Mammalian Locomotor Activity. Nat. Neurosci. 2006, 9, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Yang, F.C.; Snodgrass, P.; Li, X.; Scammell, T.E.; Davis, F.C.; Weitz, C.J. Regulation of Daily Locomotor Activity and Sleep by Hypothalamic EGF Receptor Signaling. Science 2001, 294, 2511–2515. [Google Scholar] [CrossRef] [PubMed]
- Hermes, M.L.H.J.; Kolaj, M.; Doroshenko, P.; Coderre, E.; Renaud, L.P. Effects of VPAC2 Receptor Activation on Membrane Excitability and GABAergic Transmission in Subparaventricular Zone Neurons Targeted by Suprachiasmatic Nucleus. J. Neurophysiol. 2009, 102, 1834–1842. [Google Scholar] [CrossRef][Green Version]
- Paul, S.; Hanna, L.; Harding, C.; Hayter, E.A.; Walmsley, L.; Bechtold, D.A.; Brown, T.M. Output from VIP Cells of the Mammalian Central Clock Regulates Daily Physiological Rhythms. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Tousson, E.; Meissl, H. Suprachiasmatic Nuclei Grafts Restore the Circadian Rhythm in the Paraventricular Nucleus of the Hypothalamus. J. Neurosci. 2004, 24, 2983–2988. [Google Scholar] [CrossRef]
- Paul, K.N.; Saafir, T.B.; Tosini, G. The Role of Retinal Photoreceptors in the Regulation of Circadian Rhythms. Rev. Endocr. Metab. Disord. 2009, 10, 271–278. [Google Scholar] [CrossRef]
- Morin, L.P.; Allen, C.N. The Circadian Visual System, 2005. Brain Res. Rev. 2006, 51, 1–60. [Google Scholar] [CrossRef]
- Baver, S.B.; Pickard, G.E.; Sollars, P.J.; Pickard, G.E. Two Types of Melanopsin Retinal Ganglion Cell Differentially Innervate the Hypothalamic Suprachiasmatic Nucleus and the Olivary Pretectal Nucleus. Eur. J. Neurosci. 2008, 27, 1763–1770. [Google Scholar] [CrossRef]
- Morin, L.P.; Blanchard, J.H.; Provencio, I. Retinal Ganglion Cell Projections to the Hamster Suprachiasmatic Nucleus, Intergeniculate Leaflet, and Visual Midbrain: Bifurcation and Melanopsin Immunoreactivity. J. Comp. Neurol. 2003, 465, 401–416. [Google Scholar] [CrossRef]
- Duy, P.Q.; Komal, R.; Richardson, M.E.S.; Hahm, K.S.; Fernandez, D.C.; Hattar, S. Light Has Diverse Spatiotemporal Molecular Changes in the Mouse Suprachiasmatic Nucleus. J. Biol. Rhythms 2020, 748730420961214. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.T.; Fahey, B.; Cutler, D.J.; Coogan, A.N.; Piggins, H.D. Aberrant Gating of Photic Input to the Suprachiasmatic Circadian Pacemaker of Mice Lacking the VPAC2 Receptor. J. Neurosci. 2004, 24, 3522–3526. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, D.C.; Chang, Y.-T.; Hattar, S.; Chen, S.-K. Architecture of Retinal Projections to the Central Circadian Pacemaker. Proc. Natl. Acad. Sci. USA 2016, 113, 6047–6052. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, T.; Li, J.Y.; Hayes, N.W.; Chan, J.C.; Okabe, Y.; Belin, S.; Nawabi, H.; Schmidt, T.M. A Noncanonical Inhibitory Circuit Dampens Behavioral Sensitivity to Light. Science 2020, 368, 527–531. [Google Scholar] [CrossRef]
- Wen, S.A.; Ma, D.; Zhao, M.; Xie, L.; Wu, Q.; Gou, L.; Zhu, C.; Fan, Y.; Wang, H.; Yan, J. Spatiotemporal Single-Cell Analysis of Gene Expression in the Mouse Suprachiasmatic Nucleus. Nat. Neurosci. 2020, 23, 456–467. [Google Scholar] [CrossRef]
- Kuhlman, S.J.; Silver, R.; Le Sauter, J.; Bult-Ito, A.; McMahon, D.G. Phase Resetting Light Pulses Induce Per1 and Persistent Spike Activity in a Subpopulation of Biological Clock Neurons. J. Neurosci. 2003, 23, 1441–1450. [Google Scholar] [CrossRef]
- Quintero, J.E.; Kuhlman, S.J.; McMahon, D.G. The Biological Clock Nucleus: A Multiphasic Oscillator Network Regulated by Light. J. Neurosci. 2003, 23, 8070–8076. [Google Scholar] [CrossRef]
- Langel, J.L.; Smale, L.; Esquiva, G.; Hannibal, J. Central Melanopsin Projections in the Diurnal Rodent, Arvicanthis Niloticus. Front. Neuroanat. 2015, 9, 93. [Google Scholar] [CrossRef]
- Schoonderwoerd, R.A.; de Torres Gutiérrez, P.; Blommers, R.; van Beurden, A.W.; Coenen, T.C.J.J.; Klett, N.J.; Michel, S.H.; Meijer, J.H. Inhibitory Responses to Retinohypothalamic Tract Stimulation in the Circadian Clock of the Diurnal Rodent Rhabdomys Pumilio. FASEB J. 2022, 36, e22415. [Google Scholar] [CrossRef]
- Schumann, D.M.; Cooper, H.M.; Hofmeyr, M.D.; Bennett, N.C. Light-Induced Fos Expression in the Suprachiasmatic Nucleus of the Four-Striped Field Mouse, Rhabdomys Pumilio: A Southern African Diurnal Rodent. Brain Res. Bull. 2006, 70, 270–277. [Google Scholar] [CrossRef]
- Saleeba, C.; Dempsey, B.; Le, S.; Goodchild, A.; McMullan, S. A Student’s Guide to Neural Circuit Tracing. Front. Neurosci. 2019, 13, 897. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, G.R.; Antle, M.C. Phenotype and Function of Raphe Projections to the Suprachiasmatic Nucleus. Eur. J. Neurosci. 2010, 31, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Glass, J.D.; Guinn, J.; Kaur, G.; Francl, J.M. On the Intrinsic Regulation of Neuropeptide Y Release in the Mammalian Suprachiasmatic Nucleus Circadian Clock. Eur. J. Neurosci. 2010, 31, 1117–1126. [Google Scholar] [CrossRef]
- Morin, L.P.; Blanchard, J.H. Neuromodulator Content of Hamster Intergeniculate Leaflet Neurons and Their Projection to the Suprachiasmatic Nucleus or Visual Midbrain. J. Comp. Neurol. 2001, 437, 79–90. [Google Scholar] [CrossRef]
- Deurveilher, S.; Semba, K. Indirect Projections from the Suprachiasmatic Nucleus to Major Arousal-Promoting Cell Groups in Rat: Implications for the Circadian Control of Behavioural State. Neuroscience 2005, 130, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Cohen, R.; Kronfeld-Schor, N.; Ramanathan, C.; Baumgras, A.; Smale, L. The Substructure of the Suprachiasmatic Nucleus: Similarities between Nocturnal and Diurnal Spiny Mice. Brain Behav. Evol. 2010, 75, 9–22. [Google Scholar] [CrossRef]
- Goel, N.; Lee, T.M.; Smale, L. Suprachiasmatic Nucleus and Intergeniculate Leaflet in the Diurnal Rodent Octodon Degus: Retinal Projections and Immunocytochemical Characterization. Neuroscience 1999, 92, 1491–1509. [Google Scholar] [CrossRef]
- Smale, L.; Blanchard, J.; Moore, R.Y.; Morin, L.P. Immunocytochemical Characterization of the Suprachiasmatic Nucleus and the Intergeniculate Leaflet in the Diurnal Ground Squirrel, Spermophilus Lateralis. Brain Res. 1991, 563, 77–86. [Google Scholar] [CrossRef]
- Ni, R.-J.; Shu, Y.-M.; Luo, P.-H.; Zhou, J.-N. Whole-Brain Mapping of Afferent Projections to the Suprachiasmatic Nucleus of the Tree Shrew. Tissue Cell 2021, 73, 101620. [Google Scholar] [CrossRef]
- Todd, W.D.; Venner, A.; Anaclet, C.; Broadhurst, R.Y.; De Luca, R.; Bandaru, S.S.; Issokson, L.; Hablitz, L.M.; Cravetchi, O.; Arrigoni, E.; et al. Suprachiasmatic VIP Neurons Are Required for Normal Circadian Rhythmicity and Comprised of Molecularly Distinct Subpopulations. Nat. Commun. 2020, 11, 4410. [Google Scholar] [CrossRef]
- Kahan, A.; Kassraian, P.; Altermatt, M.; Gradinaru, V. Immediate Responses to Ambient Light in the Suprachiasmatic Nucleus Reveal Distinct Activity Profiles and Retinal Connectivity for SCN VIP Neurons. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=4052008 (accessed on 12 February 2023).
- Pickard, G.E. The Afferent Connections of the Suprachiasmatic Nucleus of the Golden Hamster with Emphasis on the Retinohypothalamic Projection. J. Comp. Neurol. 1982, 211, 65–83. [Google Scholar] [CrossRef] [PubMed]
- Moga, M.M.; Moore, R.Y. Organization of Neural Inputs to the Suprachiasmatic Nucleus in the Rat. J. Comp. Neurol. 1997, 389, 508–534. [Google Scholar] [CrossRef]
- Yuan, X.-S.; Wei, H.-H.; Xu, W.; Wang, L.; Qu, W.-M.; Li, R.-X.; Huang, Z.-L. Whole-Brain Monosynaptic Afferent Projections to the Cholecystokinin Neurons of the Suprachiasmatic Nucleus. Front. Neurosci. 2018, 12, 807. [Google Scholar] [CrossRef]
- Hannibal, J.; Hundahl, C.; Fahrenkrug, J.; Rehfeld, J.F.; Friis-Hansen, L. Cholecystokinin (CCK)-Expressing Neurons in the Suprachiasmatic Nucleus: Innervation, Light Responsiveness and Entrainment in CCK-Deficient Mice. Eur. J. Neurosci. 2010, 32, 1006–1017. [Google Scholar] [CrossRef]
- Mistlberger, R.E.; Antle, M.C.; Glass, J.D.; Miller, J.D. Behavioral and Serotonergic Regulation of Circadian Rhythms. Biol. Rhythm Res. 2000, 31, 240–283. [Google Scholar] [CrossRef]
- Webb, I.C.; Antle, M.C.; Mistlberger, R.E. Regulation of Circadian Rhythms in Mammals by Behavioral Arousal. Behav. Neurosci. 2014, 128, 304–325. [Google Scholar] [CrossRef] [PubMed]
- Mrosovsky, N.; Salmon, P.A.; Menaker, M.; Ralph, M.R. Nonphotic Phase Shifting in Hamster Clock Mutants. J. Biol. Rhythms 1992, 7, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Bobrzynska, K.J.; Mrosovsky, N. Phase Shifting by Novelty-Induced Running: Activity Dose-Response Curves at Different Circadian Times. J. Comp. Physiol. A 1998, 182, 251–258. [Google Scholar] [CrossRef]
- Morin, L.P.; Shivers, K.-Y.; Blanchard, J.H.; Muscat, L. Complex Organization of Mouse and Rat Suprachiasmatic Nucleus. Neuroscience 2006, 137, 1285–1297. [Google Scholar] [CrossRef]
- Grippo, R.M.; Tang, Q.; Zhang, Q.; Chadwick, S.R.; Gao, Y.; Altherr, E.B.; Sipe, L.; Purohit, A.M.; Purohit, N.M.; Sunkara, M.D.; et al. Dopamine Signaling in the Suprachiasmatic Nucleus Enables Weight Gain Associated with Hedonic Feeding. Curr. Biol. 2020, 30, 196–208. [Google Scholar] [CrossRef]
- Grippo, R.M.; Purohit, A.M.; Zhang, Q.; Zweifel, L.S.; Güler, A.D. Direct Midbrain Dopamine Input to the Suprachiasmatic Nucleus Accelerates Circadian Entrainment. Curr. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Belle, M. Circadian Tick-Talking Across the Neuroendocrine System and Suprachiasmatic Nuclei Circuits: The Enigmatic Communication Between the Molecular and Electrical Membrane Clocks. J. Neuroendocrinol. 2015, 27, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.P.; Karatsoreos, I.N.; LeSauter, J.; Silver, R. Dose-Dependent Effects of Androgens on the Circadian Timing System and Its Response to Light. Endocrinology 2012, 153, 2344–2352. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, K.M.; Royston, S.E.; Mahoney, M.M. Modulation of Circadian Rhythms through Estrogen Receptor Signaling. Eur. J. Neurosci. 2020, 51, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Model, Z.; Butler, M.P.; LeSauter, J.; Silver, R. Suprachiasmatic Nucleus as the Site of Androgen Action on Circadian Rhythms. Horm. Behav. 2015, 73, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Su, J.D.; Qiu, J.; Zhong, Y.P.; Chen, Y.Z. Expression of Estrogen Receptor -Alpha and -Beta Immunoreactivity in the Cultured Neonatal Suprachiasmatic Nucleus: With Special Attention to GABAergic Neurons. Neuroreport 2001, 12, 1955–1959. [Google Scholar] [CrossRef] [PubMed]
- Clancy, A.N.; Whitman, C.; Michael, R.P.; Albers, H.E. Distribution of Androgen Receptor-like Immunoreactivity in the Brains of Intact and Castrated Male Hamsters. Brain Res. Bull. 1994, 33, 325–332. [Google Scholar] [CrossRef]
- Karatsoreos, I.N.; Wang, A.; Sasanian, J.; Silver, R. A Role for Androgens in Regulating Circadian Behavior and the Suprachiasmatic Nucleus. Endocrinology 2007, 148, 5487–5495. [Google Scholar] [CrossRef]
- Iwahana, E.; Karatsoreos, I.; Shibata, S.; Silver, R. Gonadectomy Reveals Sex Differences in Circadian Rhythms and Suprachiasmatic Nucleus Androgen Receptors in Mice. Horm. Behav. 2008, 53, 422–430. [Google Scholar] [CrossRef]
- Cao, Q.; Gery, S.; Dashti, A.; Yin, D.; Zhou, Y.; Gu, J.; Koeffler, H.P. A Role for the Clock Gene per1 in Prostate Cancer. Cancer Res. 2009, 69, 7619–7625. [Google Scholar] [CrossRef]
- Hashimoto, A.; Fujiki, S.; Nakamura, W.; Nakamura, T.J. Effects of Testosterone on Circadian Rhythmicity in Old Mice. J. Physiol. Sci. 2019, 69, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Vida, B.; Hrabovszky, E.; Kalamatianos, T.; Coen, C.W.; Liposits, Z.; Kalló, I. Oestrogen Receptor Alpha and Beta Immunoreactive Cells in the Suprachiasmatic Nucleus of Mice: Distribution, Sex Differences and Regulation by Gonadal Hormones. J. Neuroendocrinol. 2008, 20, 1270–1277. [Google Scholar] [CrossRef] [PubMed]
- Morin, L.P.; Fitzgerald, K.M.; Zucker, I. Estradiol Shortens the Period of Hamster Circadian Rhythms. Science 1977, 196, 305–307. [Google Scholar] [CrossRef] [PubMed]
- Royston, S.E.; Yasui, N.; Kondilis, A.G.; Lord, S.V.; Katzenellenbogen, J.A.; Mahoney, M.M. ESR1 and ESR2 Differentially Regulate Daily and Circadian Activity Rhythms in Female Mice. Endocrinology 2014, 155, 2613–2623. [Google Scholar] [CrossRef] [PubMed]
- Albers, H.E. Gonadal Hormones Organize and Modulate the Circadian System of the Rat. Am. J. Physiol. 1981, 241, R62–R66. [Google Scholar] [CrossRef]
- Kopp, C.; Ressel, V.; Wigger, E.; Tobler, I. Influence of Estrus Cycle and Ageing on Activity Patterns in Two Inbred Mouse Strains. Behav. Brain Res. 2006, 167, 165–174. [Google Scholar] [CrossRef]
- Xiao, L.; Chang, A.K.; Zang, M.-X.; Bi, H.; Li, S.; Wang, M.; Xing, X.; Wu, H. Induction of the CLOCK Gene by E2-ERα Signaling Promotes the Proliferation of Breast Cancer Cells. PLoS ONE 2014, 9, e95878. [Google Scholar] [CrossRef]
- Gery, S.; Virk, R.K.; Chumakov, K.; Yu, A.; Koeffler, H.P. The Clock Gene Per2 Links the Circadian System to the Estrogen Receptor. Oncogene 2007, 26, 7916–7920. [Google Scholar] [CrossRef]
- Nakamura, T.J.; Sellick, M.T.; Menaker, M.; Block, G.D. Estrogen Directly Modulates Circadian Rhythms of PER2 Expression in the Uterus. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1025–E1031. [Google Scholar] [CrossRef]
- Fatehi, M.; Fatehi-Hassanabad, Z. Effects of 17β-Estradiol on Neuronal Cell Excitability and Neurotransmission in the Suprachiasmatic Nucleus of Rat. Neuropsychopharmacology 2007, 33, 1354–1364. [Google Scholar] [CrossRef]
- Jones, J.R.; Tackenberg, M.C.; McMahon, D.G. Manipulating Circadian Clock Neuron Firing Rate Resets Molecular Circadian Rhythms and Behavior. Nat. Neurosci. 2015, 18, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Murphy, Z.C.; Pezuk, P.; Menaker, M.; Sellix, M.T. Effects of Ovarian Hormones on Internal Circadian Organization in Rats. Biol. Reprod. 2013, 89, 35. [Google Scholar] [CrossRef] [PubMed]
- Kruijver, F.P.M.; Swaab, D.F. Sex Hormone Receptors Are Present in the Human Suprachiasmatic Nucleus. Neuroendocrinology 2002, 75, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S.; Menaker, M. Interaction of Estradiol and Progesterone: Effects on Circadian Locomotor Rhythm of Female Golden Hamsters. Am. J. Physiol. 1980, 239, R497–R504. [Google Scholar] [CrossRef] [PubMed]
- Balsalobre, A.; Brown, S.A.; Marcacci, L.; Tronche, F.; Kellendonk, C.; Reichardt, H.M.; Schütz, G.; Schibler, U. Resetting of Circadian Time in Peripheral Tissues by Glucocorticoid Signaling. Science 2000, 289, 2344–2347. [Google Scholar] [CrossRef]
- Koyanagi, S.; Okazawa, S.; Kuramoto, Y.; Ushijima, K.; Shimeno, H.; Soeda, S.; Okamura, H.; Ohdo, S. Chronic Treatment with Prednisolone Represses the Circadian Oscillation of Clock Gene Expression in Mouse Peripheral Tissues. Mol. Endocrinol. 2006, 20, 573–583. [Google Scholar] [CrossRef]
- Reddy, A.B.; Maywood, E.S.; Karp, N.A.; King, V.M.; Inoue, Y.; Gonzalez, F.J.; Lilley, K.S.; Kyriacou, C.P.; Hastings, M.H. Glucocorticoid Signaling Synchronizes the Liver Circadian Transcriptome. Hepatology 2007, 45, 1478–1488. [Google Scholar] [CrossRef]
- Segall, L.A.; Perrin, J.S.; Walker, C.-D.; Stewart, J.; Amir, S. Glucocorticoid Rhythms Control the Rhythm of Expression of the Clock Protein, Period2, in Oval Nucleus of the Bed Nucleus of the Stria Terminalis and Central Nucleus of the Amygdala in Rats. Neuroscience 2006, 140, 753–757. [Google Scholar] [CrossRef]
- Rosenfeld, P.; van Eekelen, J.A.; Levine, S.; de Kloet, E.R. Ontogeny of Corticosteroid Receptors in the Brain. Cell. Mol. Neurobiol. 1993, 13, 295–319. [Google Scholar] [CrossRef]
- Čečmanová, V.; Houdek, P.; Šuchmanová, K.; Sládek, M.; Sumová, A. Development and Entrainment of the Fetal Clock in the Suprachiasmatic Nuclei: The Role of Glucocorticoids. J. Biol. Rhythms 2019, 34, 307–322. [Google Scholar] [CrossRef]
- Su, Y.; van der Spek, R.; Foppen, E.; Kwakkel, J.; Fliers, E.; Kalsbeek, A. Effects of Adrenalectomy on Daily Gene Expression Rhythms in the Rat Suprachiasmatic and Paraventricular Hypothalamic Nuclei and in White Adipose Tissue. Chronobiol. Int. 2015, 32, 211–224. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, M.P.; Merkus, P.; Romeijn, S.G.; Verhoef, J.C.; Merkus, F.W.H.M. Uptake of Melatonin into the Cerebrospinal Fluid after Nasal and Intravenous Delivery: Studies in Rats and Comparison with a Human Study. Pharm. Res. 2004, 21, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Klosen, P.; Lapmanee, S.; Schuster, C.; Guardiola, B.; Hicks, D.; Pevet, P.; Felder-Schmittbuhl, M.P. MT1 and MT2 Melatonin Receptors Are Expressed in Nonoverlapping Neuronal Populations. J. Pineal Res. 2019, 67, e12575. [Google Scholar] [CrossRef]
- McArthur, A.J.; Hunt, A.E.; Gillette, M.U. Melatonin Action and Signal Transduction in the Rat Suprachiasmatic Circadian Clock: Activation of Protein Kinase C at Dusk and Dawn. Endocrinology 1997, 138, 627–634. [Google Scholar] [CrossRef]
- Pfeffer, M.; Rauch, A.; Korf, H.-W.; von Gall, C. The Endogenous Melatonin (MT) Signal Facilitates Reentrainment of the Circadian System to Light-Induced Phase Advances by Acting upon MT2 Receptors. Chronobiol. Int. 2012, 29, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Kandalepas, P.C.; Mitchell, J.W.; Gillette, M.U. Melatonin Signal Transduction Pathways Require E-Box-Mediated Transcription of Per1 and Per2 to Reset the SCN Clock at Dusk. PLoS ONE 2016, 11, e0157824. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Weaver, D.R.; Jin, X.; Shearman, L.P.; Pieschl, R.L.; Gribkoff, V.K.; Reppert, S.M. Molecular Dissection of Two Distinct Actions of Melatonin on the Suprachiasmatic Circadian Clock. Neuron 1997, 19, 91–102. [Google Scholar] [CrossRef]
- Hunt, A.E.; Al-Ghoul, W.M.; Gillette, M.U.; Dubocovich, M.L. Activation of MT2 Melatonin Receptors in Rat Suprachiasmatic Nucleus Phase Advances the Circadian Clock. Am. J. Physiol.-Cell Physiol. 2001, 280, C110–C118. [Google Scholar] [CrossRef]
- Dubocovich, M.L.; Hudson, R.L.; Sumaya, I.C.; Masana, M.I.; Manna, E. Effect of MT1 Melatonin Receptor Deletion on Melatonin-Mediated Phase Shift of Circadian Rhythms in the C57BL/6 Mouse. J. Pineal Res. 2005, 39, 113–120. [Google Scholar] [CrossRef]
- McArthur, A.J.; Gillette, M.U.; Prosser, R.A. Melatonin Directly Resets the Rat Suprachiasmatic Circadian Clock in Vitro. Brain Res. 1991, 565, 158–161. [Google Scholar] [CrossRef]
- Rawashdeh, O.; Hudson, R.L.; Stepien, I.; Dubocovich, M.L. Circadian Periods of Sensitivity for Ramelteon on the Onset of Running-Wheel Activity and the Peak of Suprachiasmatic Nucleus Neuronal Firing Rhythms in C3H/HeN Mice. Chronobiol. Int. 2011, 28, 31–38. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cassone, V.M.; Chesworth, M.J.; Armstrong, S.M. Entrainment of Rat Circadian Rhythms by Daily Injection of Melatonin Depends upon the Hypothalamic Suprachiasmatic Nuclei. Physiol. Behav. 1986, 36, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Silveyra, E.; Jin, N.; Ribelayga, C.P. A Congenic Line of the C57BL/6J Mouse Strain That Is Proficient in Melatonin Synthesis. J. Pineal Res. 2018, 65, e12509. [Google Scholar] [CrossRef] [PubMed]
- Adamah-Biassi, E.B.; Hudson, R.L.; Dubocovich, M.L. Genetic Deletion of MT1 Melatonin Receptors Alters Spontaneous Behavioral Rhythms in Male and Female C57BL/6 Mice. Horm. Behav. 2014, 66, 619–627. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pfeffer, M.; Korf, H.-W.; Wicht, H. The Role of the Melatoninergic System in Light-Entrained Behavior of Mice. Int. J. Mol. Sci. 2017, 18, 530. [Google Scholar] [CrossRef]
- Reppert, S.M.; Schwartz, W.J. Maternal Suprachiasmatic Nuclei Are Necessary for Maternal Coordination of the Developing Circadian System. J. Neurosci. 1986, 6, 2724–2729. [Google Scholar] [CrossRef]
- Bates, K.; Herzog, E.D. Maternal-Fetal Circadian Communication During Pregnancy. Front. Endocrinol. 2020, 11, 198. [Google Scholar] [CrossRef]
- Bellavía, S.L.; Carpentieri, A.R.; Vaqué, A.M.; Macchione, A.F.; Vermouth, N.T. Pup Circadian Rhythm Entrainment--Effect of Maternal Ganglionectomy or Pinealectomy. Physiol. Behav. 2006, 89, 342–349. [Google Scholar] [CrossRef]
- Naitoh, N.; Watanabe, Y.; Matsumura, K.; Murai, I.; Kobayashi, K.; Imai-Matsumura, K.; Ohtuka, H.; Takagi, K.; Miyake, Y.; Satoh, K.; et al. Alteration by Maternal Pinealectomy of Fetal and Neonatal Melatonin and Dopamine D1 Receptor Binding in the Suprachiasmatic Nuclei. Biochem. Biophys. Res. Commun. 1998, 253, 850–854. [Google Scholar] [CrossRef]
- Yellon, S.M.; Longo, L.D. Effect of Maternal Pinealectomy and Reverse Photoperiod on the Circadian Melatonin Rhythm in the Sheep and Fetus during the Last Trimester of Pregnancy. Biol. Reprod. 1988, 39, 1093–1099. [Google Scholar] [CrossRef]
- Reppert, S.M.; Schwartz, W.J. Maternal Endocrine Extirpations Do Not Abolish Maternal Coordination of the Fetal Circadian Clock. Endocrinology 1986, 119, 1763–1767. [Google Scholar] [CrossRef]
- Viswanathan, N. Maternal Entrainment in the Circadian Activity Rhythm of Laboratory Mouse (C57BL/6J). Physiol. Behav. 1999, 68, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Watts, A.G.; Swanson, L.W.; Sanchez-Watts, G. Efferent Projections of the Suprachiasmatic Nucleus: I. Studies Using Anterograde Transport of Phaseolus Vulgaris Leucoagglutinin in the Rat. J. Comp. Neurol. 1987, 258, 204–229. [Google Scholar] [PubMed]
- Leak, R.K.; Moore, R.Y. Topographic Organization of Suprachiasmatic Nucleus Projection Neurons. J. Comp. Neurol. 2001, 433, 312–334. [Google Scholar] [CrossRef] [PubMed]
- Kalsbeek, A.; Teclemariam-Mesbah, R.; Pévet, P. Efferent Projections of the Suprachiasmatic Nucleus in the Golden Hamster (Mesocricetus Auratus). J. Comp. Neurol. 1993, 332, 293–314. [Google Scholar] [CrossRef]
- Watts, A.G.; Swanson, L.W. Efferent Projections of the Suprachiasmatic Nucleus: II. Studies Using Retrograde Transport of Fluorescent Dyes and Simultaneous Peptide Immunohistochemistry in the Rat. J. Comp. Neurol. 1987, 258, 230–252. [Google Scholar] [CrossRef]
- Zhang, C.; Truong, K.K.; Zhou, Q.-Y. Efferent Projections of Prokineticin 2 Expressing Neurons in the Mouse Suprachiasmatic Nucleus. PLoS ONE 2009, 4, e7151. [Google Scholar] [CrossRef]
- Sunkin, S.M.; Ng, L.; Lau, C.; Dolbeare, T.; Gilbert, T.L.; Thompson, C.L.; Hawrylycz, M.; Dang, C. Allen Brain Atlas: An Integrated Spatio-Temporal Portal for Exploring the Central Nervous System. Nucleic Acids Res. 2013, 41, D996–D1008. [Google Scholar] [CrossRef]
- François, M.; Delgado, I.C.; Lafond, A.; Lewis, E.M.; Kuromaru, M.; Hassouna, R.; Deng, S.; Thaker, V.V.; Dölen, G.; Zeltser, L.M. Amygdala AVPR1A Mediates Susceptibility to Chronic Social Isolation in Females. bioRxiv 2023, 2023.02.15.528679. [Google Scholar]
- Romanov, R.A.; Zeisel, A.; Bakker, J.; Girach, F.; Hellysaz, A.; Tomer, R.; Alpár, A.; Mulder, J.; Clotman, F.; Keimpema, E.; et al. Molecular Interrogation of Hypothalamic Organization Reveals Distinct Dopamine Neuronal Subtypes. Nat. Neurosci. 2017, 20, 176–188. [Google Scholar] [CrossRef]
- Porcu, A.; Nilsson, A.; Booreddy, S.; Barnes, S.A.; Welsh, D.K.; Dulcis, D. Seasonal Changes in Day Length Induce Multisynaptic Neurotransmitter Switching to Regulate Hypothalamic Network Activity and Behavior. Sci. Adv. 2022, 8, eabn9867. [Google Scholar] [CrossRef] [PubMed]
- Van Der Beek, E.M.; Horvath, T.L.; Wiegant, V.M.; Van Den Hurk, R.; Buijs, R.M. Evidence for a Direct Neuronal Pathway from the Suprachiasmatic Nucleus to the Gonadotropin-Releasing Hormone System: Combined Tracing and Light and Electron Microscopic Immunocytochemical Studies. J. Comp. Neurol. 1997, 384, 569–579. [Google Scholar] [CrossRef]
- Christian, C.A.; Moenter, S.M. Vasoactive Intestinal Polypeptide Can Excite Gonadotropin-Releasing Hormone Neurons in a Manner Dependent on Estradiol and Gated by Time of Day. Endocrinology 2008, 149, 3130–3136. [Google Scholar] [CrossRef] [PubMed]
- Russo, K.A.; La, J.L.; Stephens, S.B.Z.; Poling, M.C.; Padgaonkar, N.A.; Jennings, K.J.; Piekarski, D.J.; Kauffman, A.S.; Kriegsfeld, L.J. Circadian Control of the Female Reproductive Axis Through Gated Responsiveness of the RFRP-3 System to VIP Signaling. Endocrinology 2015, 156, 2608–2618. [Google Scholar] [CrossRef] [PubMed]
- Vida, B.; Deli, L.; Hrabovszky, E.; Kalamatianos, T.; Caraty, A.; Coen, C.W.; Liposits, Z.; Kalló, I. Evidence for Suprachiasmatic Vasopressin Neurones Innervating Kisspeptin Neurones in the Rostral Periventricular Area of the Mouse Brain: Regulation by Oestrogen. J. Neuroendocrinol. 2010, 22, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Piet, R.; Fraissenon, A.; Boehm, U.; Herbison, A.E. Estrogen Permits Vasopressin Signaling in Preoptic Kisspeptin Neurons in the Female Mouse. J. Neurosci. 2015, 35, 6881–6892. [Google Scholar] [CrossRef]
- Kalló, I.; Kalamatianos, T.; Wiltshire, N.; Shen, S.; Sheward, W.J.; Harmar, A.J.; Coen, C.W. Transgenic Approach Reveals Expression of the VPAC2 Receptor in Phenotypically Defined Neurons in the Mouse Suprachiasmatic Nucleus and in Its Efferent Target Sites. Eur. J. Neurosci. 2004, 19, 2201–2211. [Google Scholar] [CrossRef]
- Williams, W.P., 3rd; Jarjisian, S.G.; Mikkelsen, J.D.; Kriegsfeld, L.J. Circadian Control of Kisspeptin and a Gated GnRH Response Mediate the Preovulatory Luteinizing Hormone Surge. Endocrinology 2011, 152, 595–606. [Google Scholar] [CrossRef]
- Guzmán-Ruiz, M.A.; Ramirez-Corona, A.; Guerrero-Vargas, N.N.; Sabath, E.; Ramirez-Plascencia, O.D.; Fuentes-Romero, R.; León-Mercado, L.A.; Basualdo Sigales, M.; Escobar, C.; Buijs, R.M. Role of the Suprachiasmatic and Arcuate Nuclei in Diurnal Temperature Regulation in the Rat. J. Neurosci. 2015, 35, 15419–15429. [Google Scholar] [CrossRef]
- Méndez-Hernández, R.; Escobar, C.; Buijs, R.M. Suprachiasmatic Nucleus-Arcuate Nucleus Axis: Interaction Between Time and Metabolism Essential for Health. Obesity 2020, 28 (Suppl. S1), S10–S17. [Google Scholar] [CrossRef]
- Buijs, R.M.; Guzmán Ruiz, M.A.; Hernández, R.M.; Cortés, B.R. The Suprachiasmatic Nucleus; a Responsive Clock Regulating Homeostasis by Daily Changing the Setpoints of Physiological Parameters. Auton. Neurosci. 2019. [Google Scholar] [CrossRef]
- Kalsbeek, A.; La Fleur, S.; Van Heijningen, C.; Buijs, R.M. Suprachiasmatic GABAergic Inputs to the Paraventricular Nucleus Control Plasma Glucose Concentrations in the Rat via Sympathetic Innervation of the Liver. J. Neurosci. 2004, 24, 7604–7613. [Google Scholar] [CrossRef] [PubMed]
- Perreau-Lenz, S.; Kalsbeek, A.; Pévet, P.; Buijs, R.M. Glutamatergic Clock Output Stimulates Melatonin Synthesis at Night. Eur. J. Neurosci. 2004, 19, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Kalsbeek, A.; van Heerikhuize, J.J.; Wortel, J.; Buijs, R.M. A Diurnal Rhythm of Stimulatory Input to the Hypothalamo-Pituitary-Adrenal System as Revealed by Timed Intrahypothalamic Administration of the Vasopressin V1 Antagonist. J. Neurosci. 1996, 16, 5555–5565. [Google Scholar] [CrossRef] [PubMed]
- Teclemariam-Mesbah, R.; Kalsbeek, A.; Pevet, P.; Buijs, R.M. Direct Vasoactive Intestinal Polypeptide-Containing Projection from the Suprachiasmatic Nucleus to Spinal Projecting Hypothalamic Paraventricular Neurons. Brain Res. 1997, 748, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Perreau-Lenz, S.; Kalsbeek, A.; Garidou, M.-L.; Wortel, J.; Van Der Vliet, J.; Van Heijningen, C.; Simonneaux, V.; Pévet, P.; Buijs, R.M. Suprachiasmatic Control of Melatonin Synthesis in Rats: Inhibitory and Stimulatory Mechanisms. Eur. J. Neurosci. 2003, 17, 221–228. [Google Scholar] [CrossRef]
- Kalsbeek, A.; van der Spek, R.; Lei, J.; Endert, E.; Buijs, R.M.; Fliers, E. Circadian Rhythms in the Hypothalamo–pituitary–adrenal (HPA) Axis. Mol. Cell. Endocrinol. 2012, 349, 20–29. [Google Scholar] [CrossRef]
- Ulrich-Lai, Y.M.; Arnhold, M.M.; Engeland, W.C. Adrenal Splanchnic Innervation Contributes to the Diurnal Rhythm of Plasma Corticosterone in Rats by Modulating Adrenal Sensitivity to ACTH. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1128–R1135. [Google Scholar] [CrossRef]
- Jasper, M.S.; Engeland, W.C. Splanchnic Neural Activity Modulates Ultradian and Circadian Rhythms in Adrenocortical Secretion in Awake Rats. Neuroendocrinology 1994, 59, 97–109. [Google Scholar] [CrossRef]
- Kalsbeek, A.; Buijs, R.M.; van Heerikhuize, J.J.; Arts, M.; van der Woude, T.P. Vasopressin-Containing Neurons of the Suprachiasmatic Nuclei Inhibit Corticosterone Release. Brain Res. 1992, 580, 62–67. [Google Scholar] [CrossRef]
- Kalsbeek, A.; Fliers, E.; Hofman, M.A.; Swaab, D.F.; Buijs, R.M. Vasopressin and the Output of the Hypothalamic Biological Clock. J. Neuroendocrinol. 2010, 22, 362–372. [Google Scholar] [CrossRef]
- Jones, J.R.; Chaturvedi, S.; Granados-Fuentes, D.; Herzog, E.D. Circadian Neurons in the Paraventricular Nucleus Entrain and Sustain Daily Rhythms in Glucocorticoids. Nat. Commun. 2021, 12, 5763. [Google Scholar] [CrossRef] [PubMed]
- Mazuski, C.; Chen, S.P.; Herzog, E.D. Different Roles for VIP Neurons in the Neonatal and Adult Suprachiasmatic Nucleus. J. Biol. Rhythms 2020, 748730420932073. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.N.; Coderre, E.; Renaud, L.P. Glutamate and GABA Mediate Suprachiasmatic Nucleus Inputs to Spinal-Projecting Paraventricular Neurons. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R1283–R1289. [Google Scholar] [CrossRef] [PubMed]
- Bartness, T.J.; Song, C.K.; Demas, G.E. SCN Efferents to Peripheral Tissues: Implications for Biological Rhythms. J. Biol. Rhythms 2001, 16, 196–204. [Google Scholar] [CrossRef]
- Amir, S.; Shizgal, P.; Rompré, P.P. Glutamate Injection into the Suprachiasmatic Nucleus Stimulates Brown Fat Thermogenesis in the Rat. Brain Res. 1989, 498, 140–144. [Google Scholar] [CrossRef]
- Borbély, A.A.; Daan, S.; Wirz-Justice, A.; Deboer, T. The Two-Process Model of Sleep Regulation: A Reappraisal. J. Sleep Res. 2016, 25, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Borbély, A.A. A Two Process Model of Sleep Regulation. Hum. Neurobiol. 1982, 1, 195–204. [Google Scholar] [PubMed]
- Lu, J.; Zhang, Y.H.; Chou, T.C.; Gaus, S.E.; Elmquist, J.K.; Shiromani, P.; Saper, C.B. Contrasting Effects of Ibotenate Lesions of the Paraventricular Nucleus and Subparaventricular Zone on Sleep-Wake Cycle and Temperature Regulation. J. Neurosci. 2001, 21, 4864–4874. [Google Scholar] [CrossRef]
- Chou, T.C.; Scammell, T.E.; Gooley, J.J.; Gaus, S.E.; Saper, C.B.; Lu, J. Critical Role of Dorsomedial Hypothalamic Nucleus in a Wide Range of Behavioral Circadian Rhythms. J. Neurosci. 2003, 23, 10691–10702. [Google Scholar] [CrossRef] [PubMed]
- Vujovic, N.; Gooley, J.J.; Jhou, T.C.; Saper, C.B. Projections from the Subparaventricular Zone Define Four Channels of Output from the Circadian Timing System. J. Comp. Neurol. 2015, 523, 2714–2737. [Google Scholar] [CrossRef] [PubMed]
- Scammell, T.E.; Arrigoni, E.; Lipton, J.O. Neural Circuitry of Wakefulness and Sleep. Neuron 2017, 93, 747–765. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Scammell, T.E.; Lu, J. Hypothalamic Regulation of Sleep and Circadian Rhythms. Nature 2005, 437, 1257–1263. [Google Scholar] [CrossRef]
- Kroeger, D.; Ferrari, L.L.; Petit, G.; Mahoney, C.E.; Fuller, P.M.; Arrigoni, E.; Scammell, T.E. Cholinergic, Glutamatergic, and GABAergic Neurons of the Pedunculopontine Tegmental Nucleus Have Distinct Effects on Sleep/Wake Behavior in Mice. J. Neurosci. 2017, 37, 1352–1366. [Google Scholar] [CrossRef]
- Hallanger, A.E.; Levey, A.I.; Lee, H.J.; Rye, D.B.; Wainer, B.H. The Origins of Cholinergic and Other Subcortical Afferents to the Thalamus in the Rat. J. Comp. Neurol. 1987, 262, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.E.; Yizhar, O.; Chikahisa, S.; Nguyen, H.; Adamantidis, A.; Nishino, S.; Deisseroth, K.; de Lecea, L. Tuning Arousal with Optogenetic Modulation of Locus Coeruleus Neurons. Nat. Neurosci. 2010, 13, 1526–1533. [Google Scholar] [CrossRef] [PubMed]
- Landry, G.J.; Simon, M.M.; Webb, I.C.; Mistlberger, R.E. Persistence of a Behavioral Food-Anticipatory Circadian Rhythm Following Dorsomedial Hypothalamic Ablation in Rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1527–R1534. [Google Scholar] [CrossRef]
- Landry, G.J.; Kent, B.A.; Patton, D.F.; Jaholkowski, M.; Marchant, E.G.; Mistlberger, R.E. Evidence for Time-of-Day Dependent Effect of Neurotoxic Dorsomedial Hypothalamic Lesions on Food Anticipatory Circadian Rhythms in Rats. PLoS ONE 2011, 6, e24187. [Google Scholar] [CrossRef]
- Tahara, Y.; Hirao, A.; Moriya, T.; Kudo, T.; Shibata, S. Effects of Medial Hypothalamic Lesions on Feeding-Induced Entrainment of Locomotor Activity and Liver Per2 Expression in Per2::luc Mice. J. Biol. Rhythms 2010, 25, 9–18. [Google Scholar] [CrossRef]
- Faber, C.L.; Deem, J.D.; Phan, B.A.; Doan, T.P.; Ogimoto, K.; Mirzadeh, Z.; Schwartz, M.W.; Morton, G.J. Leptin Receptor Neurons in the Dorsomedial Hypothalamus Regulate Diurnal Patterns of Feeding, Locomotion, and Metabolism. eLife 2021, 10. [Google Scholar] [CrossRef]
- Li, L.; Zhang, M.-Q.; Sun, X.; Liu, W.-Y.; Huang, Z.-L.; Wang, Y.-Q. Role of Dorsomedial Hypothalamus GABAergic Neurons in Sleep-Wake States in Response to Changes in Ambient Temperature in Mice. Int. J. Mol. Sci. 2022, 23, 1270. [Google Scholar] [CrossRef] [PubMed]
- Bell, B.J.; Liu, Q.; Kim, D.W.; Lee, S.S.; Liu, Q.; Blum, I.D.; Wang, A.A.; Bedont, J.L.; Chang, A.J.; Issa, H.; et al. A Clock-Driven Neural Network Critical for Arousal. bioRxiv 2020, 2020.03.12.989921. [Google Scholar]
- Zhou, Q.; Liu, C.; Chen, T.; Liu, Y.; Cao, R.; Ni, X.; Yang, W.Z.; Shen, Q.; Sun, H.; Shen, W.L. Cooling-Activated Dorsomedial Hypothalamic BDNF Neurons Control Cold Defense in Mice. J. Neurochem. 2022. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.-W. Dorsomedial Hypothalamic Prodynorphin Neuron Is Crucial for Expression of Food Anticipation in Mice. Ph.D. Thesis, University of California San Francisco, San Francisco, CA, USA, 2019. Available online: https://escholarship.org/uc/item/87m4w3bx (accessed on 13 February 2023).
- Buijs, F.N.; Guzmán-Ruiz, M.; León-Mercado, L.; Basualdo, M.C.; Escobar, C.; Kalsbeek, A.; Buijs, R.M. Suprachiasmatic Nucleus Interaction with the Arcuate Nucleus; Essential for Organizing Physiological Rhythms. eNeuro 2017, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Coppari, R.; Ichinose, M.; Lee, C.E.; Pullen, A.E.; Kenny, C.D.; McGovern, R.A.; Tang, V.; Liu, S.M.; Ludwig, T.; Chua, S.C., Jr.; et al. The Hypothalamic Arcuate Nucleus: A Key Site for Mediating Leptin’s Effects on Glucose Homeostasis and Locomotor Activity. Cell Metab. 2005, 1, 63–72. [Google Scholar] [CrossRef]
- Padilla, S.L.; Perez, J.G.; Ben-Hamo, M.; Johnson, C.W.; Sanchez, R.E.A.; Bussi, I.L.; Palmiter, R.D.; de la Iglesia, H.O. Kisspeptin Neurons in the Arcuate Nucleus of the Hypothalamus Orchestrate Circadian Rhythms and Metabolism. Curr. Biol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Moriya, T.; Aida, R.; Kudo, T.; Akiyama, M.; Doi, M.; Hayasaka, N.; Nakahata, N.; Mistlberger, R.; Okamura, H.; Shibata, S. The Dorsomedial Hypothalamic Nucleus Is Not Necessary for Food-Anticipatory Circadian Rhythms of Behavior, Temperature or Clock Gene Expression in Mice. Eur. J. Neurosci. 2009, 29, 1447–1460. [Google Scholar] [CrossRef]
- Landry, G.J.; Yamakawa, G.R.; Webb, I.C.; Mear, R.J.; Mistlberger, R.E. The Dorsomedial Hypothalamic Nucleus Is Not Necessary for the Expression of Circadian Food-Anticipatory Activity in Rats. J. Biol. Rhythms 2007, 22, 467–478. [Google Scholar] [CrossRef]
- Acosta-Galvan, G.; Yi, C.-X.; van der Vliet, J.; Jhamandas, J.H.; Panula, P.; Angeles-Castellanos, M.; Del Carmen Basualdo, M.; Escobar, C.; Buijs, R.M. Interaction between Hypothalamic Dorsomedial Nucleus and the Suprachiasmatic Nucleus Determines Intensity of Food Anticipatory Behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 5813–5818. [Google Scholar] [CrossRef]
- Bernardis, L.L. Disruption of Diurnal Feeding and Weight Gain Cycles in Weanling Rats by Ventromedial and Dorsomedial Hypothalamic Lesions. Physiol. Behav. 1973, 10, 855–861. [Google Scholar] [CrossRef]
- Liao, G.-Y.; Kinney, C.E.; An, J.J.; Xu, B. TrkB-Expressing Neurons in the Dorsomedial Hypothalamus Are Necessary and Sufficient to Suppress Homeostatic Feeding. Proc. Natl. Acad. Sci. USA 2019, 116, 3256–3261. [Google Scholar] [CrossRef] [PubMed]
- Garfield, A.S.; Shah, B.P.; Burgess, C.R.; Li, M.M.; Li, C.; Steger, J.S.; Madara, J.C.; Campbell, J.N.; Kroeger, D.; Scammell, T.E.; et al. Dynamic GABAergic Afferent Modulation of AgRP Neurons. Nat. Neurosci. 2016, 19, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Rau, A.R.; Hentges, S.T. GABAergic Inputs to POMC Neurons Originating from the Dorsomedial Hypothalamus Are Regulated by Energy State. J. Neurosci. 2019, 39, 6449–6459. [Google Scholar] [CrossRef]
- Li, A.-J.; Wiater, M.F.; Oostrom, M.T.; Smith, B.R.; Wang, Q.; Dinh, T.T.; Roberts, B.L.; Jansen, H.T.; Ritter, S. Leptin-Sensitive Neurons in the Arcuate Nuclei Contribute to Endogenous Feeding Rhythms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R1313–R1326. [Google Scholar] [CrossRef]
- Wiater, M.F.; Mukherjee, S.; Li, A.-J.; Dinh, T.T.; Rooney, E.M.; Simasko, S.M.; Ritter, S. Circadian Integration of Sleep-Wake and Feeding Requires NPY Receptor-Expressing Neurons in the Mediobasal Hypothalamus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1569–R1583. [Google Scholar] [CrossRef]
- Santoso, P.; Nakata, M.; Ueta, Y.; Yada, T. Suprachiasmatic Vasopressin to Paraventricular Oxytocin Neurocircuit in the Hypothalamus Relays Light Reception to Inhibit Feeding Behavior. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E478–E488. [Google Scholar] [CrossRef] [PubMed]
- Gizowski, C.; Zaelzer, C.; Bourque, C.W. Clock-Driven Vasopressin Neurotransmission Mediates Anticipatory Thirst prior to Sleep. Nature 2016, 537, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Todd, W.D.; Fenselau, H.; Wang, J.L.; Zhang, R.; Machado, N.L.; Venner, A.; Broadhurst, R.Y.; Kaur, S.; Lynagh, T.; Olson, D.P.; et al. A Hypothalamic Circuit for the Circadian Control of Aggression. Nat. Neurosci. 2018, 21, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Luo, A.H.; Aston-Jones, G. Circuit Projection from Suprachiasmatic Nucleus to Ventral Tegmental Area: A Novel Circadian Output Pathway. Eur. J. Neurosci. 2009, 29, 748–760. [Google Scholar] [CrossRef]
- Legoratti-Sánchez, M.O.; Guevara-Guzmán, R.; Solano-Flores, L.P. Electrophysiological Evidences of a Bidirectional Communication between the Locus Coeruleus and the Suprachiasmatic Nucleus. Brain Res. Bull. 1989, 23, 283–288. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Chen, S.; Zhu, Y.; Oshinsky, M.L. A Neural Circuit for Circadian Regulation of Arousal. Nat. Neurosci. 2001, 4, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Conway-Campbell, B.L.; Sarabdjitsingh, R.A.; McKenna, M.A.; Pooley, J.R.; Kershaw, Y.M.; Meijer, O.C.; de Kloet, E.R.; Lightman, S.L. Glucocorticoid Ultradian Rhythmicity Directs Cyclical Gene Pulsing of the Clock Gene Period 1 in Rat Hippocampus. J. Neuroendocrinol. 2010, 22, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Reddy, T.E.; Gertz, J.; Crawford, G.E.; Garabedian, M.J.; Myers, R.M. The Hypersensitive Glucocorticoid Response Specifically Regulates Period 1 and Expression of Circadian Genes. Mol. Cell. Biol. 2012, 32, 3756–3767. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Nakahata, Y.; Tanaka, M.; Yoshida, M.; Soma, H.; Shinohara, K.; Yasuda, A.; Mamine, T.; Takumi, T. Acute Physical Stress Elevates Mouse Period1 mRNA Expression in Mouse Peripheral Tissues via a Glucocorticoid-Responsive Element*. J. Biol. Chem. 2005, 280, 42036–42043. [Google Scholar] [CrossRef]
- Yan, L.; Smale, L.; Nunez, A.A. Circadian and Photic Modulation of Daily Rhythms in Diurnal Mammals. Eur. J. Neurosci. 2020, 51, 551–566. [Google Scholar] [CrossRef]
- Kalsbeek, A.; Verhagen, L.A.W.; Schalij, I.; Foppen, E.; Saboureau, M.; Bothorel, B.; Buijs, R.M.; Pévet, P. Opposite Actions of Hypothalamic Vasopressin on Circadian Corticosterone Rhythm in Nocturnal versus Diurnal Species. Eur. J. Neurosci. 2008, 27, 818–827. [Google Scholar] [CrossRef]
- Torres-Farfan, C.; Valenzuela, F.J.; Ebensperger, R.; Méndez, N.; Campino, C.; Richter, H.G.; Valenzuela, G.J.; Serón-Ferré, M. Circadian Cortisol Secretion and Circadian Adrenal Responses to ACTH Are Maintained in Dexamethasone Suppressed Capuchin Monkeys (Cebus Apella). Am. J. Primatol. 2008, 70, 93–100. [Google Scholar] [CrossRef]
- Castillo, M.R.; Hochstetler, K.J.; Greene, D.M.; Firmin, S.I.; Tavernier, R.J.; Raap, D.K.; Bult-Ito, A. Circadian Rhythm of Core Body Temperature in Two Laboratory Mouse Lines. Physiol. Behav. 2005, 86, 538–545. [Google Scholar] [CrossRef]
- Buhr, E.D.; Yoo, S.-H.; Takahashi, J.S. Temperature as a Universal Resetting Cue for Mammalian Circadian Oscillators. Science 2010, 330, 379–385. [Google Scholar] [CrossRef]
- Brown, S.A.; Zumbrunn, G.; Fleury-Olela, F.; Preitner, N.; Schibler, U. Rhythms of Mammalian Body Temperature Can Sustain Peripheral Circadian Clocks. Curr. Biol. 2002, 12, 1574–1583. [Google Scholar] [CrossRef]
- Tamaru, T.; Hattori, M.; Honda, K.; Benjamin, I.; Ozawa, T.; Takamatsu, K. Synchronization of Circadian Per2 Rhythms and HSF1-BMAL1:CLOCK Interaction in Mouse Fibroblasts after Short-Term Heat Shock Pulse. PLoS ONE 2011, 6, e24521. [Google Scholar] [CrossRef] [PubMed]
- Chappuis, S.; Ripperger, J.A.; Schnell, A.; Rando, G.; Jud, C.; Wahli, W.; Albrecht, U. Role of the Circadian Clock Gene Per2 in Adaptation to Cold Temperature. Mol. Metab 2013, 2, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Kim, J.Y. Mammalian Circadian Networks Mediated by the Suprachiasmatic Nucleus. FEBS J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Deurveilher, S.; Semba, K. Indirect Projections from the Suprachiasmatic Nucleus to the Median Preoptic Nucleus in Rat. Brain Res. 2003, 987, 100–106. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Meaney, M.J.; Amir, S. The Effects of Prostaglandin E2 Injected into the Paraventricular Nucleus of the Hypothalamus on Brown Adipose Tissue Thermogenesis in Spontaneously Hypertensive Rats. Brain Res. 1993, 613, 285–287. [Google Scholar] [CrossRef]
- Ralph, M.R.; Foster, R.G.; Davis, F.C.; Menaker, M. Transplanted Suprachiasmatic Nucleus Determines Circadian Period. Science 1990, 247, 975–978. [Google Scholar] [CrossRef]
- Meyer-Bernstein, E.L.; Jetton, A.E.; Matsumoto, S.I.; Markuns, J.F.; Lehman, M.N.; Bittman, E.L. Effects of Suprachiasmatic Transplants on Circadian Rhythms of Neuroendocrine Function in Golden Hamsters. Endocrinology 1999, 140, 207–218. [Google Scholar] [CrossRef]
- Silver, R.; LeSauter, J.; Tresco, P.A.; Lehman, M.N. A Diffusible Coupling Signal from the Transplanted Suprachiasmatic Nucleus Controlling Circadian Locomotor Rhythms. Nature 1996, 382, 810–813. [Google Scholar] [CrossRef]
- Kramer, A.; Yang, F.-C.; Kraves, S.; Weitz, C.J. A Screen for Secreted Factors of the Suprachiasmatic Nucleus. In Methods in Enzymology; Young, M.W., Ed.; Academic Press: Cambridge, MA, USA, 2005; Volume 393, pp. 645–663. [Google Scholar]
- Yao, Y.; Taub, A.B.; LeSauter, J.; Silver, R. Identification of the Suprachiasmatic Nucleus Venous Portal System in the Mammalian Brain. Nat. Commun. 2021, 12, 5643. [Google Scholar] [CrossRef]
- Maywood, E.S.; Chesham, J.E.; O’Brien, J.A.; Hastings, M.H. A Diversity of Paracrine Signals Sustains Molecular Circadian Cycling in Suprachiasmatic Nucleus Circuits. Proc. Natl. Acad. Sci. USA 2011, 108, 14306–14311. [Google Scholar] [CrossRef]
- Schwartz, W.J.; Coleman, R.J.; Reppert, S.M. A Daily Vasopressin Rhythm in Rat Cerebrospinal Fluid. Brain Res. 1983, 263, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, W.J.; Reppert, S.M. Neural Regulation of the Circadian Vasopressin Rhythm in Cerebrospinal Fluid: A Pre-Eminent Role for the Suprachiasmatic Nuclei. J. Neurosci. 1985, 5, 2771–2778. [Google Scholar] [CrossRef] [PubMed]
- Jolkkonen, J.; Tuomisto, L.; van Wimersma Greidanus, T.B.; Riekkinen, P.J. Vasopressin Levels in the Cerebrospinal Fluid of Rats with Lesions of the Paraventricular and Suprachiasmatic Nuclei. Neurosci. Lett. 1988, 86, 184–188. [Google Scholar] [CrossRef]
- Kagerbauer, S.M.; Debus, J.M.; Martin, J.; Gempt, J.; Jungwirth, B.; Hapfelmeier, A.; Podtschaske, A.H. Absence of a Diurnal Rhythm of Oxytocin and Arginine-Vasopressin in Human Cerebrospinal Fluid, Blood and Saliva. Neuropeptides 2019, 78, 101977. [Google Scholar] [CrossRef] [PubMed]
- Reppert, S.M.; Schwartz, W.J.; Artman, H.G.; Fisher, D.A. Comparison of the Temporal Profiles of Vasopressin and Oxytocin in the Cerebrospinal Fluid of the Cat, Monkey and Rat. Brain Res. 1983, 261, 341–345. [Google Scholar] [CrossRef]
- Gilbert, J.; Davis, F.C. Behavioral Effects of Systemic Transforming Growth Factor-Alpha in Syrian Hamsters. Behav. Brain Res. 2009, 198, 440–448. [Google Scholar] [CrossRef][Green Version]
- Paul, J.R.; Davis, J.A.; Goode, L.K.; Becker, B.K.; Fusilier, A.; Meador-Woodruff, A.; Gamble, K.L. Circadian Regulation of Membrane Physiology in Neural Oscillators Throughout the Brain. Eur. J. Neurosci. 2019. [Google Scholar] [CrossRef]
- Guilding, C.; Piggins, H.D. Challenging the Omnipotence of the Suprachiasmatic Timekeeper: Are Circadian Oscillators Present throughout the Mammalian Brain? Eur. J. Neurosci. 2007, 25, 3195–3216. [Google Scholar] [CrossRef]
- Tonsfeldt, K.J.; Schoeller, E.L.; Brusman, L.E.; Cui, L.J.; Lee, J.; Mellon, P.L. The Contribution of the Circadian Gene Bmal1 to Female Fertility and the Generation of the Preovulatory Luteinizing Hormone Surge. J. Endocr. Soc. 2019. [Google Scholar] [CrossRef]
- Yu, X.; Zecharia, A.; Zhang, Z.; Yang, Q.; Yustos, R.; Jager, P.; Vyssotski, A.L.; Maywood, E.S.; Chesham, J.E.; Ma, Y.; et al. Circadian Factor BMAL1 in Histaminergic Neurons Regulates Sleep Architecture. Curr. Biol. 2014, 24, 2838–2844. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starnes, A.N.; Jones, J.R. Inputs and Outputs of the Mammalian Circadian Clock. Biology 2023, 12, 508. https://doi.org/10.3390/biology12040508
Starnes AN, Jones JR. Inputs and Outputs of the Mammalian Circadian Clock. Biology. 2023; 12(4):508. https://doi.org/10.3390/biology12040508
Chicago/Turabian StyleStarnes, Ashley N., and Jeff R. Jones. 2023. "Inputs and Outputs of the Mammalian Circadian Clock" Biology 12, no. 4: 508. https://doi.org/10.3390/biology12040508
APA StyleStarnes, A. N., & Jones, J. R. (2023). Inputs and Outputs of the Mammalian Circadian Clock. Biology, 12(4), 508. https://doi.org/10.3390/biology12040508