

Genome-Based Analysis of the Potential Bioactivity of the Terrestrial Streptomyces vinaceusdrappus Strain AC-40

 , , , ,
, , , , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Purification of the Strain AC-40
2.2. Solvent Extraction of the Strain AC-40 Bioactive Metabolites
2.3. Antimicrobial Activity Screening of the Strain AC-40 Bioactive Metabolites
2.4. Genomic DNA Extraction, and Sequencing of the Strain AC-40
2.5. Genome Assembly, Scaffolding, and Annotation of the Strain AC-40
2.6. Strain AC-40 Typing and Phylogeny
2.7. General Genome Feature and Pathways of the Strain AC-40
2.8. Screening the Genome of the Strain AC-40 for Genes Related to Beneficial Traits
2.9. Pan-Genome and Comparative Genome Analysis
3. Results
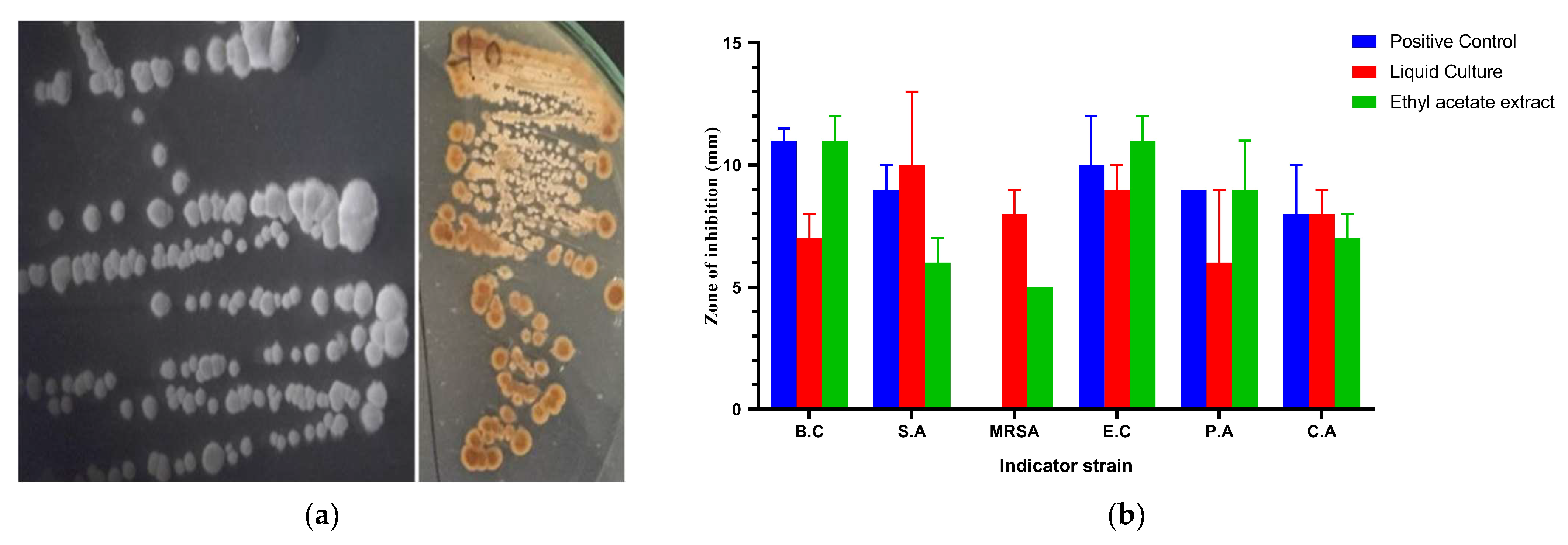
3.1. General Features and Antimicrobial Activity of the Strain AC-40
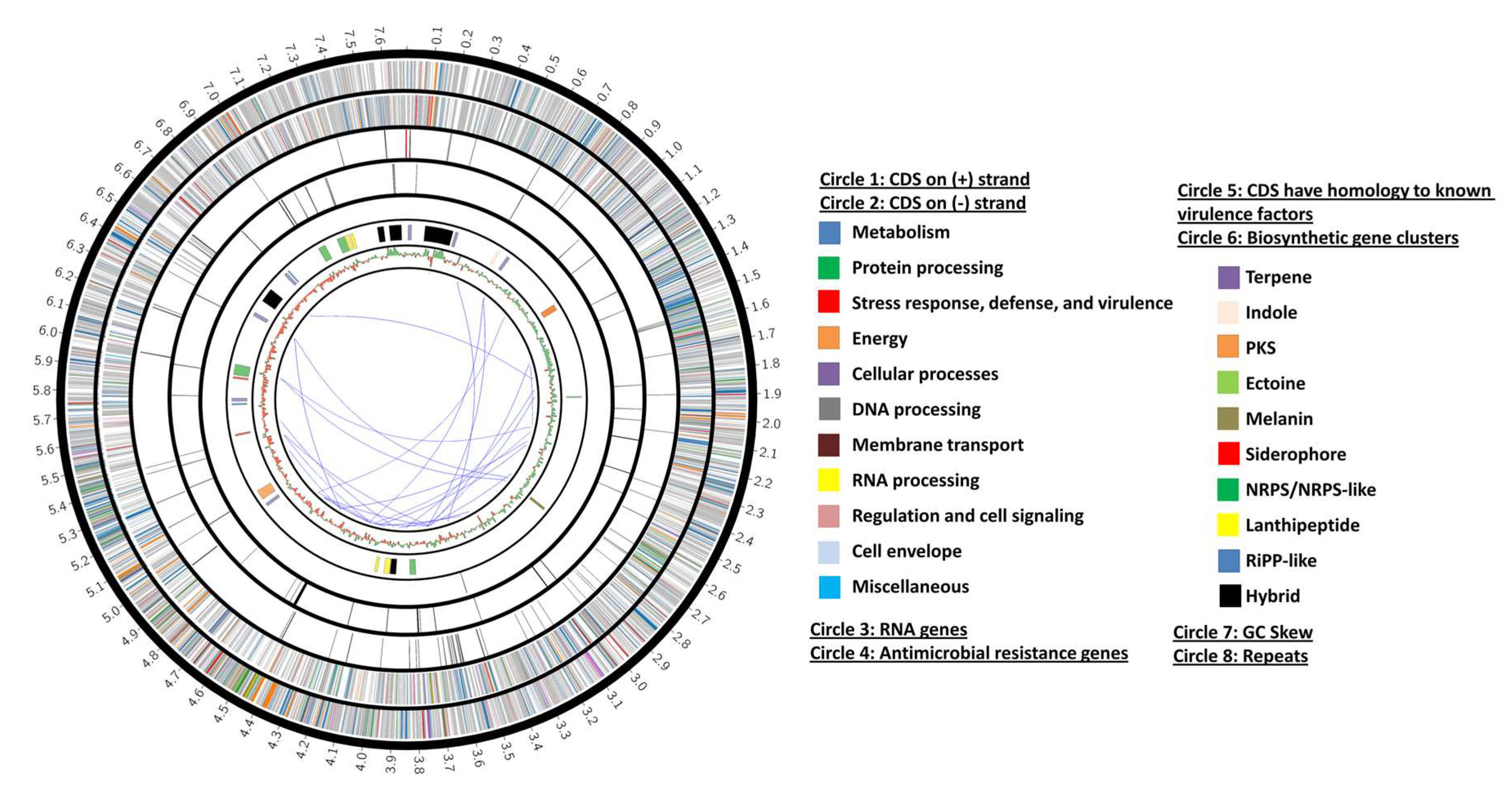
3.2. AC-40 Genome Assembly, Scaffolding, and Annotation
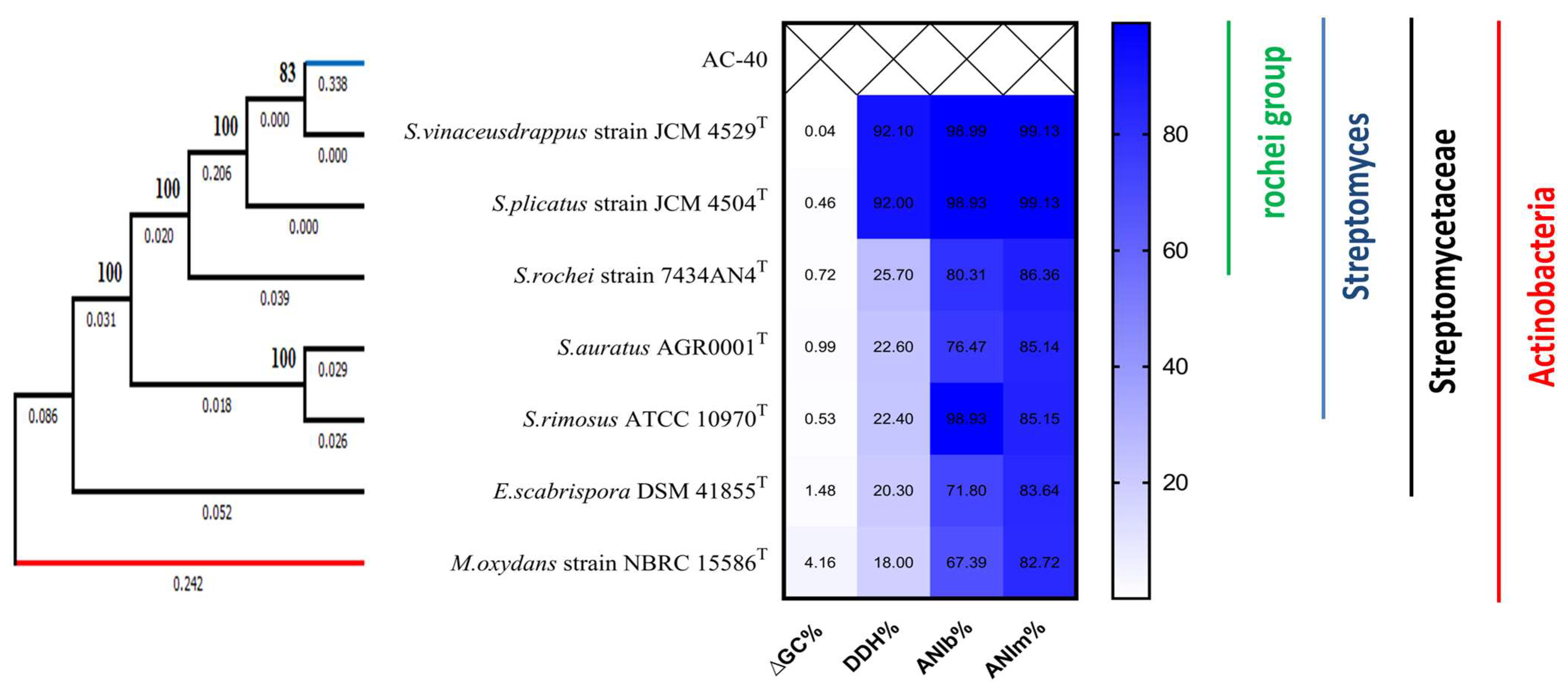
3.3. AC-40 Typing and Phylogeny
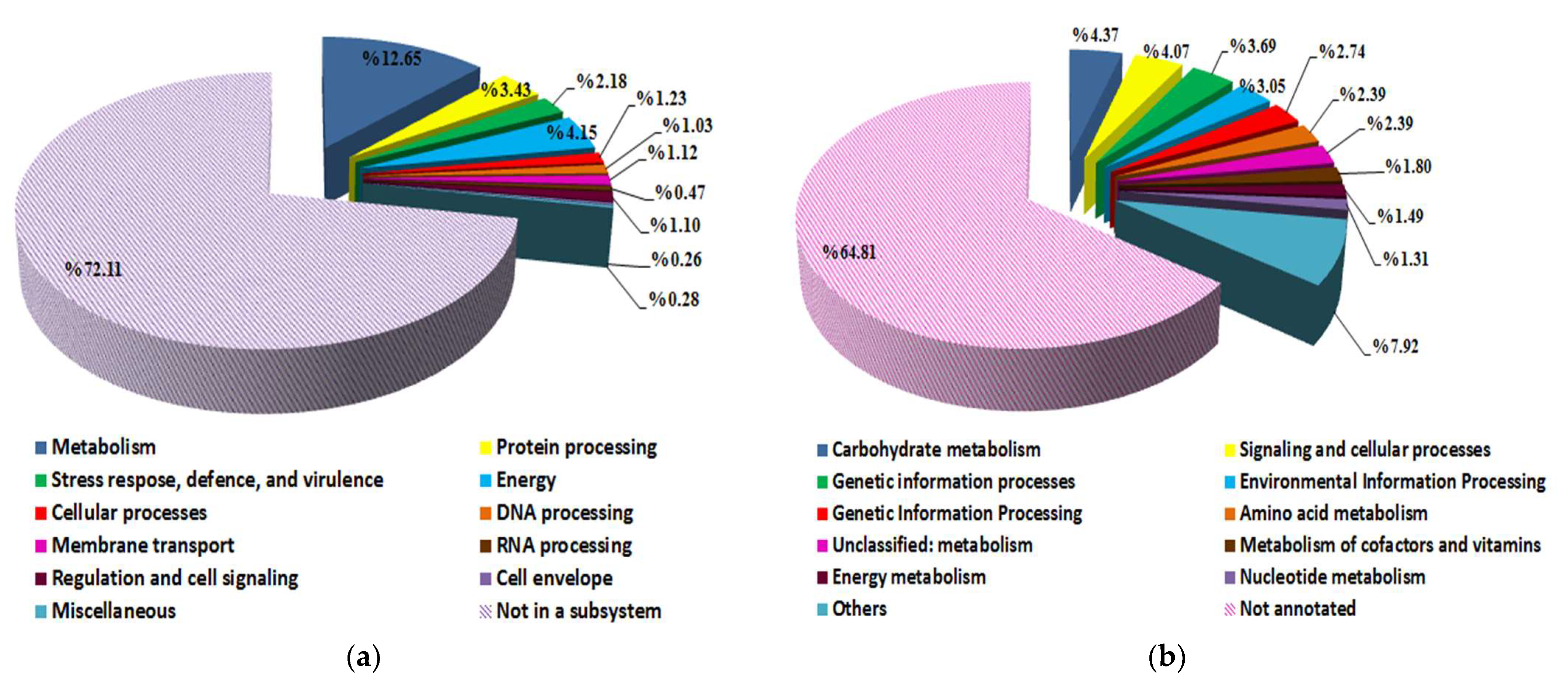
3.4. Streptomyces vinaceusdrappus Strain AC-40 Genome Features and Pathways Reconstruction
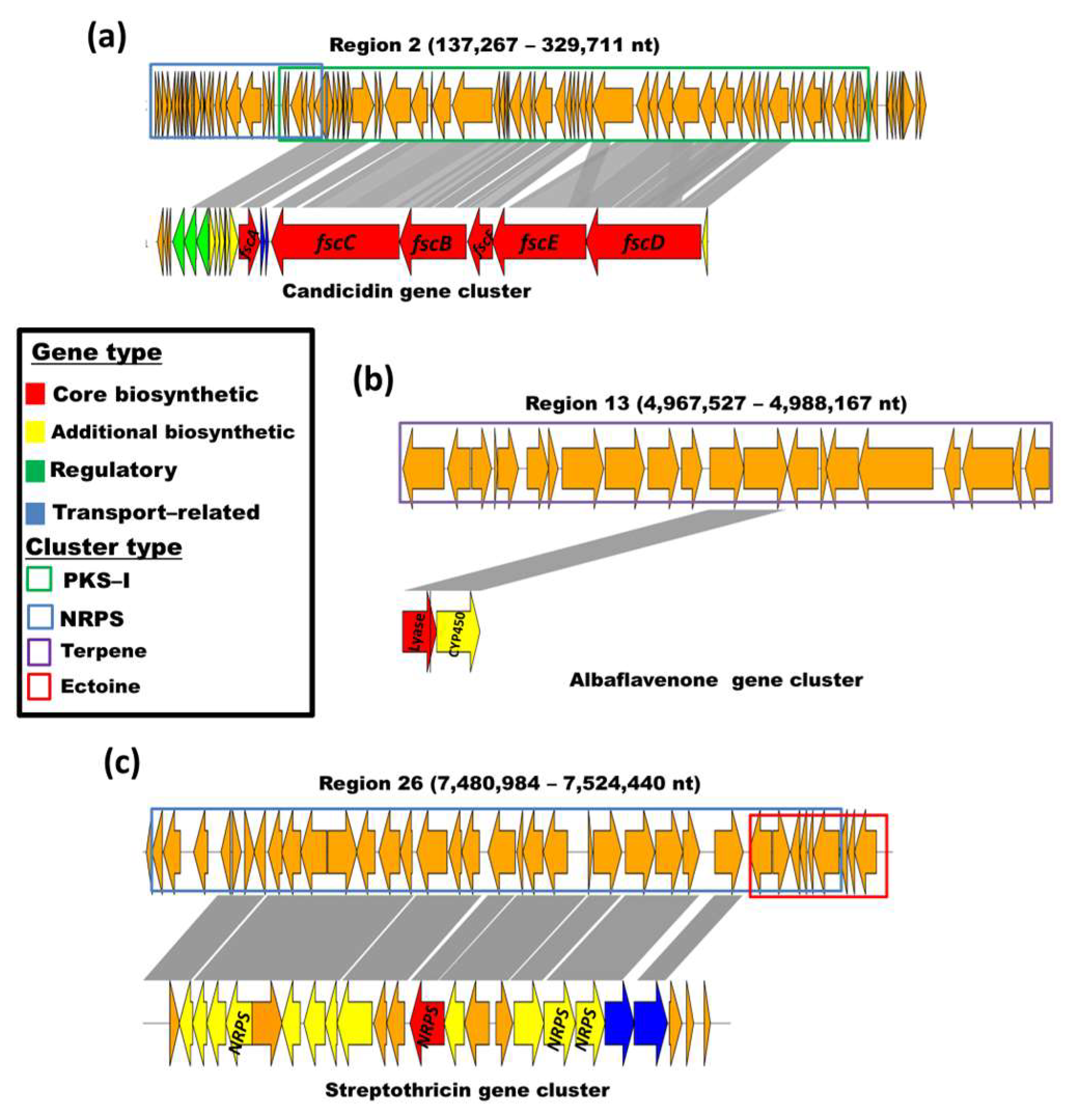
3.5. Streptomyces vinaceusdrappus Strain AC-40 Secondary Metabolites-Related Gene Clusters
3.6. Streptomyces vinaceusdrappus Strain AC-40 Genes Related to Plant Growth Promotion Traits
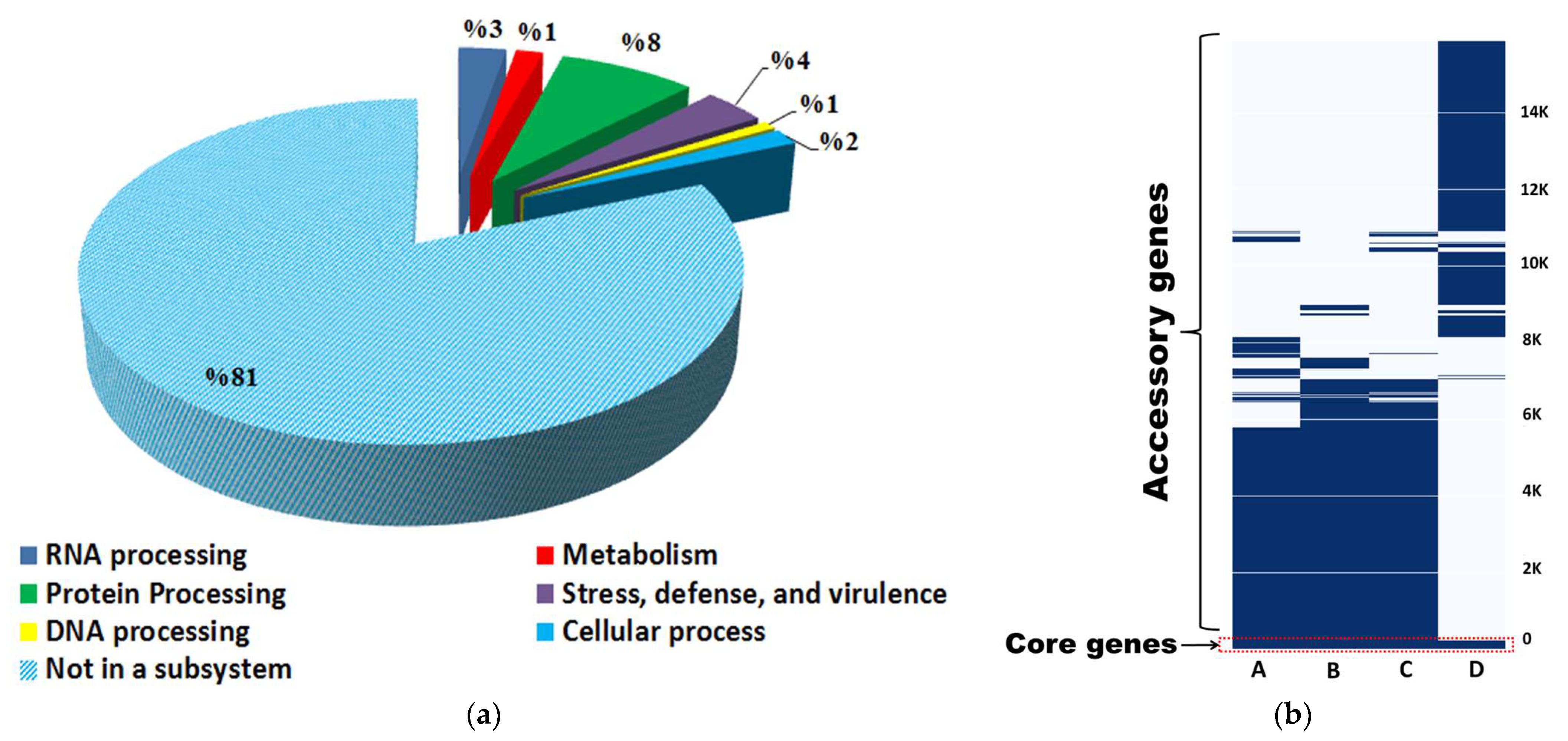
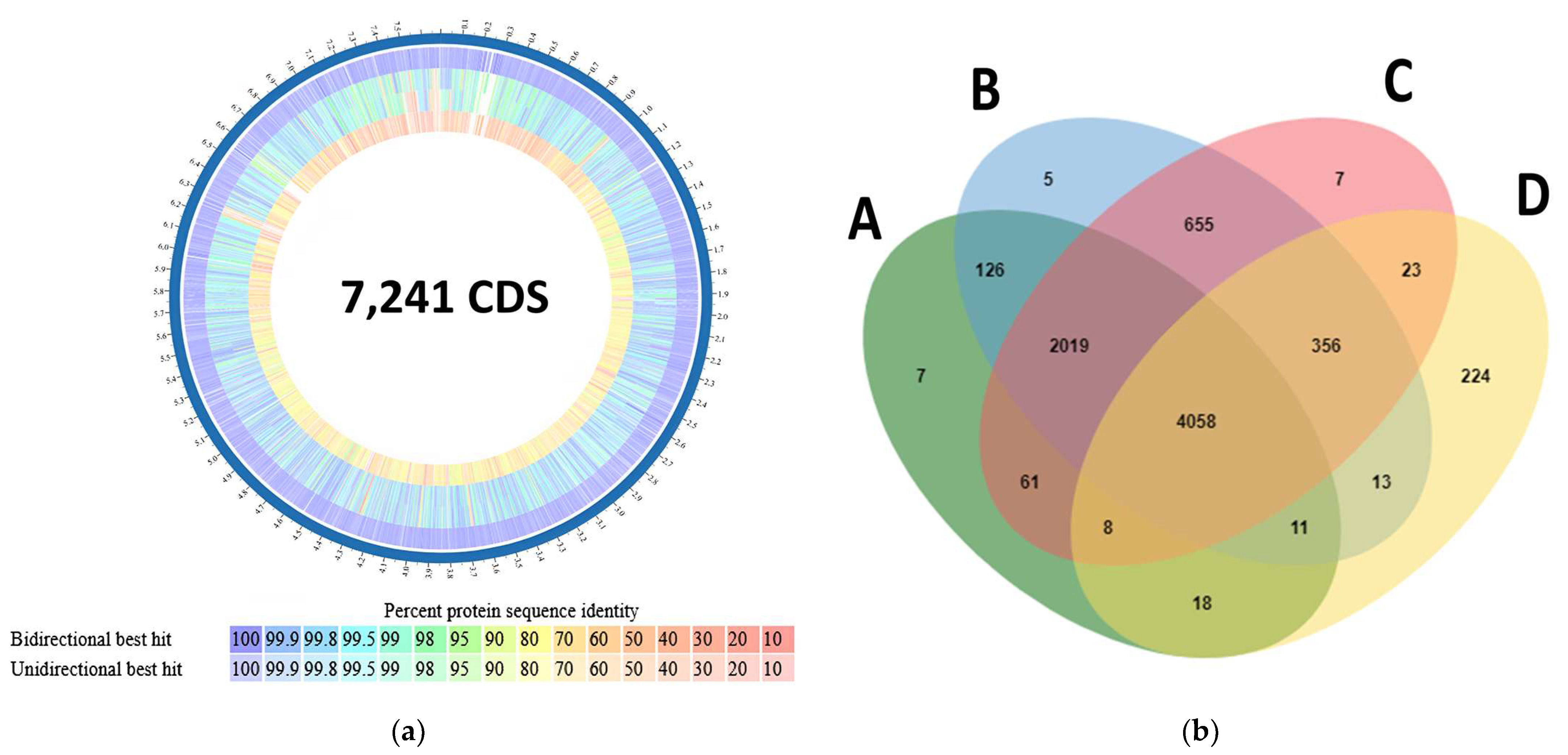
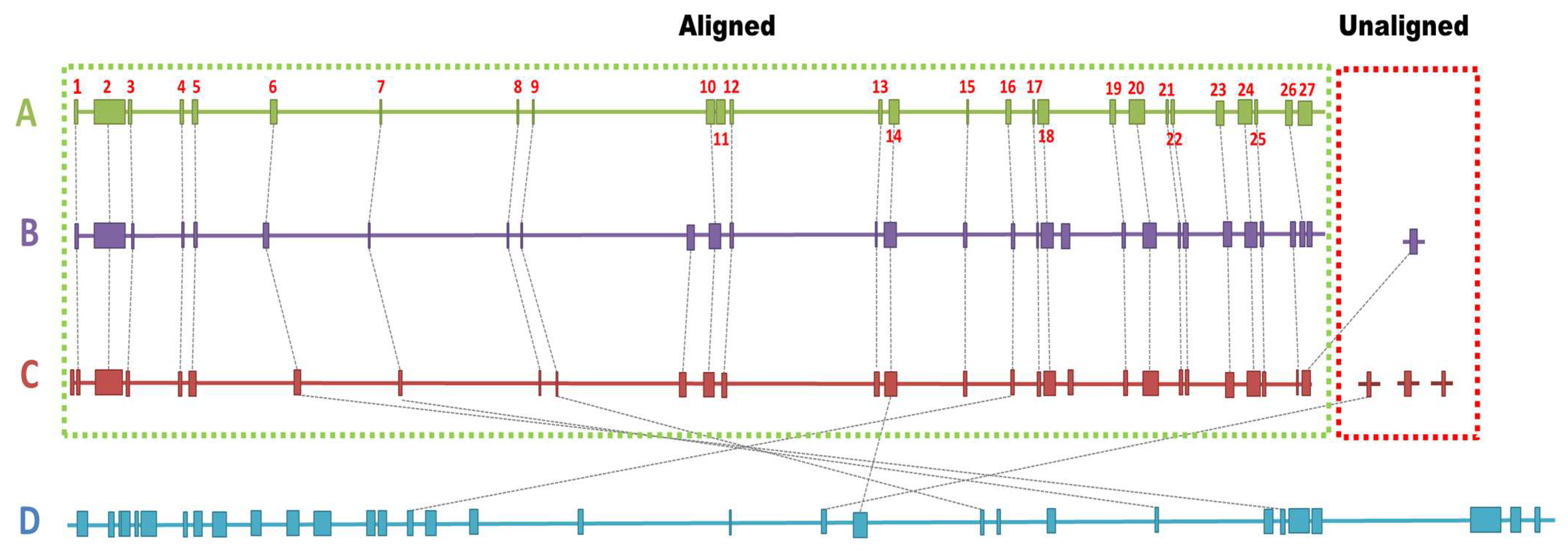
3.7. Pan-Genome and Comparative Genomics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aminov, R.I. A brief history of the antibiotic era: Lessons learned and challenges for the future. Front. Microbiol. 2010, 1, 134. [Google Scholar] [CrossRef]
- Hoffman, P.S. Antibacterial discovery: 21st century challenges. Antibiotics 2020, 9, 213. [Google Scholar] [CrossRef]
- Lee, L.-H.; Goh, B.-H.; Chan, K.-G. Editorial: Actinobacteria: Prolific Producers of Bioactive Metabolites. Front. Microbiol. 2020, 11, 1612. [Google Scholar] [CrossRef]
- Bundale, S.; Singh, J.; Begde, D.; Nashikkar, N.; Upadhyay, A. Rare actinobacteria: A potential source of bioactive polyketides and peptides. World J. Microbiol. Biotechnol. 2019, 35, 92. [Google Scholar] [CrossRef]
- Donald, L.; Pipite, A.; Subramani, R.; Owen, J.; Keyzers, R.A.; Taufa, T. Streptomyces: Still the Biggest Producer of New Natural Secondary Metabolites, a Current Perspective. Microbiol. Res. 2022, 13, 418–465. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef]
- Sedeek, A.M.; Ismail, M.M.; Elsayed, T.R.; Ramadan, M.A. Recent methods for discovering novel bioactive metabolites, specifically antimicrobial agents, from marine-associated micro-organisms. Lett. Appl. Microbiol. 2022, 75, 511–525. [Google Scholar] [CrossRef]
- Jia, F.; Liu, C.; Zhao, J.; Zhang, Y.; Li, L.; Zhou, S.; Shen, Y.; Wang, X.; Xiang, W. Streptomyces vulcanius sp. nov., a novel actinomycete isolated from volcanic sediment. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2015, 107, 15–21. [Google Scholar] [CrossRef]
- Zerouki, C.; Bensalah, F.; Kuittinen, S.; Pappinen, A.; Turunen, O. Whole-genome sequencing of two Streptomyces strains isolated from the sand dunes of Sahara. BMC Genom. 2021, 22, 578. [Google Scholar] [CrossRef]
- Risdian, C.; Mozef, T.; Wink, J. Biosynthesis of polyketides in Streptomyces. Microorganisms 2019, 7, 124. [Google Scholar] [CrossRef]
- Lee, N.; Hwang, S.; Kim, J.; Cho, S.; Palsson, B.; Cho, B.K. Mini review: Genome mining approaches for the identification of secondary metabolite biosynthetic gene clusters in Streptomyces. Comput. Struct. Biotechnol. J. 2020, 18, 1548–1556. [Google Scholar] [CrossRef]
- Quinn, G.A.; Banat, A.M.; Abdelhameed, A.M.; Banat, I.M. Streptomyces from traditional medicine: Sources of new innovations in antibiotic discovery. J. Med. Microbiol. 2020, 69, 1040–1048. [Google Scholar] [CrossRef]
- Supong, K.; Thawai, C.; Choowong, W.; Kittiwongwattana, C.; Thanaboripat, D.; Laosinwattana, C.; Koohakan, P.; Parinthawong, N.; Pittayakhajonwut, P. Antimicrobial compounds from endophytic Streptomyces sp. BCC72023 isolated from rice (Oryza sativa L.). Res. Microbiol. 2016, 167, 290–298. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Giovanardi, D.; Stefani, E. Plant Growth Promoting and Biocontrol Activity of Streptomyces spp. as Endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef]
- Ruangwong, O.U.; Kunasakdakul, K.; Daengsuwan, W.; Wonglom, P.; Pitija, K.; Sunpapao, A. A Streptomyces rhizobacterium with antifungal properties against spadix rot in flamingo flowers. Physiol. Mol. Plant Pathol. 2022, 117, 101784. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Ondov, B.D.; Treangen, T.J.; Melsted, P.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Mash: Fast genome and metagenome distance estimation using MinHash. Genome Biol. 2016, 17, 132. [Google Scholar] [CrossRef]
- Alonge, M.; Soyk, S.; Ramakrishnan, S.; Wang, X.; Goodwin, S.; Sedlazeck, F.J.; Lippman, Z.B.; Schatz, M.C. RaGOO: Fast and accurate reference-guided scaffolding of draft genomes. Genome Biol. 2019, 20, 224. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A web server for whole-genome comparison and annotation of orthologous clusters across multiple species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef] [PubMed]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 871, 1473. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wen, W.; Qin, M.; He, Y.; Xu, D.; Li, L. Biosynthetic Mechanisms of Secondary Metabolites Promoted by the Interaction Between Endophytes and Plant Hosts. Front. Microbiol. 2022, 13, 2584. [Google Scholar] [CrossRef] [PubMed]
- Hamidpour, R.; Hamidpour, S.; Hamidpour, M.; Marshall, V.; Hamidpour, R. Pelargonium graveolens (Rose Geranium)-A Novel Therapeutic Agent for Antibacterial, Antioxidant, Antifungal and Diabetics. Arch. Cancer Res. 2017, 5, 134. [Google Scholar] [CrossRef]
- Arakawa, K.; Kodama, K.; Tatsuno, S.; Ide, S.; Kinashi, H. Analysis of the loading and hydroxylation steps in lankamycin biosynthesis in Streptomyces rochei. Antimicrob. Agents Chemother. 2006, 50, 1946–1952. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhang, H.; Li, S.; Xiao, J.; Zhang, G.; Zhu, Y.; Niu, S.; Ju, J.; Zhang, C. Characterization of the amicetin biosynthesis gene cluster from Streptomyces vinaceusdrappus NRRL 2363 implicates two alternative strategies for amide bond formation. Appl. Environ. Microbiol. 2012, 78, 2393–2401. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, U.; Augustin, J.; Spröer, C.; Gelbrecht, J.; Schumann, P.; Ulrich, A. Taxonomic characterisation of Proteus terrae sp. nov., a N2O-producing, nitrate-ammonifying soil bacterium. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2015, 108, 1457–1468. [Google Scholar] [CrossRef]
- Dooley, F.D.; Nair, S.P.; Ward, P.D. Increased Growth and Germination Success in Plants following Hydrogen Sulfide Administration. PLoS ONE 2013, 8, e62048. [Google Scholar] [CrossRef]
- Heyduk, K. Evolution of Crassulacean acid metabolism in response to the environment: Past, present, and future. Plant Physiol. 2022, 190, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, J.; Feng, G.; Li, Z. New Genomic Insights into “Entotheonella” Symbionts in Theonella swinhoei: Mixotrophy, Anaerobic Adaptation, Resilience, and Interaction. Front. Microbiol. 2016, 7, 1333. [Google Scholar] [CrossRef] [PubMed]
- Sedeek, A.M.; Ismail, M.M.; Elsayed, T.R.; Ramadan, M.A. Evaluation of the Marine Bacterial Population in the Great Bitter Lake, Egypt, as a Source of Antimicrobial Secondary Metabolites. Fermentation 2022, 8, 309. [Google Scholar] [CrossRef]
- Ji, Z.; Wang, M.; Zhang, J.; Wei, S.; Wu, W. Two new members of streptothricin class antibiotics from Streptomyces qinlingensis sp. nov. J. Antibiot. 2007, 60, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Gil, J.A.; Campelo-Diez, A.B. Candicidin biosynthesis in Streptomyces griseus. Appl. Microbiol. Biotechnol. 2003, 60, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Lin, X.; Lei, L.; Lamb, D.C.; Kelly, S.L.; Waterman, M.R.; Cane, D.E. Biosynthesis of the sesquiterpene antibiotic albaflavenone in Streptomyces coelicolor A3(2). J. Biol. Chem. 2008, 283, 8183–8189. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guo, M.; Yang, J.; Chen, J.; Xie, B.; Sun, Z. Potential TSPO Ligand and Photooxidation Quencher Isorenieratene from Arctic Ocean Rhodococcus sp. B7740. Mar. Drugs 2019, 17, 316. [Google Scholar] [CrossRef]
- Bilstein, A.; Heinrich, A.; Rybachuk, A.; Mösges, R. Ectoine in the Treatment of Irritations and Inflammations of the Eye Surface. Biomed Res. Int. 2021, 2021, 8885032. [Google Scholar] [CrossRef]
- Graf, R.; Anzali, S.; Buenger, J.; Pfluecker, F.; Driller, H. The multifunctional role of ectoine as a natural cell protectant. Clin. Dermatol. 2008, 26, 326–333. [Google Scholar] [CrossRef]
- Challis, G.L.; Ravel, J. Coelichelin, a new peptide siderophore encoded by the Streptomyces coelicolor genome: Structure prediction from the sequence of its non-ribosomal peptide synthetase. FEMS Microbiol. Lett. 2000, 187, 111–114. [Google Scholar] [CrossRef]
- Zhao, B.; Moody, S.C.; Hider, R.C.; Lei, L.; Kelly, S.L.; Waterman, M.R.; Lamb, D.C. Structural Analysis of Cytochrome P450 105N1 Involved in the Biosynthesis of the Zincophore, Coelibactin. Int. J. Mol. Sci. 2012, 13, 8500–8513. [Google Scholar] [CrossRef] [PubMed]
- Lagedroste, M.; Reiners, J.; Knospe, C.V.; Smits, S.H.J.; Schmitt, L. A Structural View on the Maturation of Lanthipeptides. Front. Microbiol. 2020, 11, 1183. [Google Scholar] [CrossRef] [PubMed]
- Hegemann, J.D.; Süssmuth, R.D. Matters of class: Coming of age of class III and IV lanthipeptides. RSC Chem. Biol. 2020, 1, 110–127. [Google Scholar] [CrossRef] [PubMed]
- Kodani, S.; Hudson, M.E.; Durrant, M.C.; Buttner, M.J.; Nodwell, J.R.; Willey, J.M. The SapB morphogen is a lantibiotic-like peptide derived from the product of the developmental gene ramS in Streptomyces coelicolor. Proc. Natl. Acad. Sci. USA 2004, 101, 11448–11453. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Start | End | BGC Type | Most Similar Known Metabolite | %Identity |
|---|---|---|---|---|---|
| 1 | 11,426 | 34,005 | Terpene | Isorenieratene | 85% |
| 2 | 137,267 | 329,711 | PKS-I/NRPS | Candicidin | 95% |
| 3 | 347,918 | 368,491 | Terpene | Lysolipin I | 4% |
| 4 | 669,221 | 690,342 | Indole | 5-dimethylallylindole-3-acetonitrile | 100% |
| 5 | 748,998 | 771,393 | Terpene | Carotenoid | 54% |
| 6 | 1,220,910 | 1,262,076 | PKS-III | Flaviolin/1,3,6,8-tetrahydroxynaphthalene | 100% |
| 7 | 1,898,364 | 1,907,075 | Ectoine | Ectoine | 100% |
| 8 | 2,737,184 | 2,747,792 | Melanin | Istamycin | 4% |
| 9 | 2,831,522 | 2,842,444 | Siderophore | Desferrioxamin B and E | 83% |
| 10 | 3,919,974 | 3,960,930 | PKS-like/furan | Methylenomycin A | 9% |
| 11 | 3,962,785 | 4,005,126 | Lanthipeptide-V | Cacaoidin | 7% |
| 12 | 4,052,201 | 4,071,203 | Lanthipeptide-III | Catenulipeptin | 60% |
| 13 | 4,967,527 | 4,988,167 | Terpene | Albaflavenone | 100% |
| 14 | 5,025,751 | 5,094,254 | PKS-II | Spore pigment | 66% |
| 15 | 5,510,542 | 5,520,672 | Siderophore | - | - |
| 16 | 5,755,125 | 5,776,362 | Terpene | Geosmin | 100% |
| 17 | 5,916,847 | 5,928,346 | Siderophore | Grincamycin | 8% |
| 18 | 5,942,866 | 6,016,271 | NRPS | Lipopeptide | 72% |
| 19 | 6,380,102 | 6,406,068 | Terpene | Hopene | 100% |
| 20 | 6,500,253 | 6,601,623 | PKS-I | Vicenistatin | 80% |
| 21 | 6,739,417 | 6,760,448 | Terpene | Versipelostatin | 5% |
| 22 | 6,773,508 | 6,783,723 | Ripp-like | Informatipeptin | 42% |
| 23 | 7,035,002 | 7,091,898 | NRPS | Coelichelin | 100% |
| 24 | 7,186,423 | 7,269,342 | NRPS/Lanthipeptide-I | Coelibactin | 100% |
| 25 | 7,280,111 | 7,302,690 | Lanthipeptide-III | SapB | 100% |
| 26 | 7,480,984 | 7,524,440 | NRPS/Ectoine | Streptothricin | 87% |
| 27 | 7,563,953 | 7,649,675 | PKS-II/Butyrolactone | Fluostatins M-Q | 60% |
| Trait | Genes | Product |
|---|---|---|
| Nitrogen assimilation | nasD, nirD | Nitrite reductase |
| nark | Nitrate/nitrite transporter | |
| Phosphate solubilization | gpm2 | Acid phosphatase |
| phoD | Alkaline phosphatase D | |
| ycdX | Phosphatase | |
| - | Putative phosphatase | |
| Iron sequestration | yfiZ, yfhA | putative siderophore transport system permease protein |
| yusV | putative siderophore transport system ATP-binding protein | |
| Phytohormone synthesis | trpC | Indole-3-glycerol phosphate synthase |
| Biocatalyst | htpX, prtS | Protease |
| snpA | Extracellular small neutral protease | |
| ydeA | Putative protease | |
| lip1 | Lipase 1 | |
| - | Lipase 2 | |
| - | Thermostable monoacylglycerol lipase | |
| chiA | putative bifunctional chitinase/lysozyme | |
| chiC, chiD, chtA | Chitinase | |
| - | Exochitinase 1 | |
| aml | Alpha-amylase | |
| katA, katE | Catalase | |
| bca | Bromoperoxidase-catalase | |
| katG | Catalase-peroxidase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sedeek, A.M.; Salah, I.; Kamel, H.L.; Soltan, M.A.; Nour, E.; Alshammari, A.; Riaz Rajoka, M.S.; Elsayed, T.R. Genome-Based Analysis of the Potential Bioactivity of the Terrestrial Streptomyces vinaceusdrappus Strain AC-40. Biology 2023, 12, 345. https://doi.org/10.3390/biology12030345
Sedeek AM, Salah I, Kamel HL, Soltan MA, Nour E, Alshammari A, Riaz Rajoka MS, Elsayed TR. Genome-Based Analysis of the Potential Bioactivity of the Terrestrial Streptomyces vinaceusdrappus Strain AC-40. Biology. 2023; 12(3):345. https://doi.org/10.3390/biology12030345
Chicago/Turabian StyleSedeek, Abdelrahman M., Israa Salah, Hasnaa L. Kamel, Mohamed A. Soltan, Eman Nour, Abdulrahman Alshammari, Muhammad Shahid Riaz Rajoka, and Tarek R. Elsayed. 2023. "Genome-Based Analysis of the Potential Bioactivity of the Terrestrial Streptomyces vinaceusdrappus Strain AC-40" Biology 12, no. 3: 345. https://doi.org/10.3390/biology12030345
APA StyleSedeek, A. M., Salah, I., Kamel, H. L., Soltan, M. A., Nour, E., Alshammari, A., Riaz Rajoka, M. S., & Elsayed, T. R. (2023). Genome-Based Analysis of the Potential Bioactivity of the Terrestrial Streptomyces vinaceusdrappus Strain AC-40. Biology, 12(3), 345. https://doi.org/10.3390/biology12030345