
Ellenberg Indicator Values Disclose Complex Environmental Filtering Processes in Plant Communities along an Elevational Gradient



Abstract
Simple Summary
Abstract
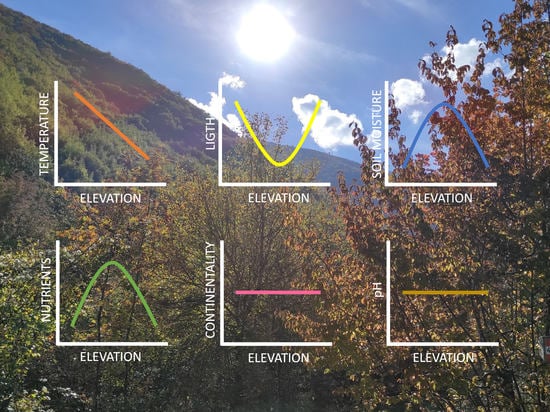
1. Introduction
- (1)
- The EIVs for temperature should decrease with increasing elevation, following the decrease of temperature with increasing elevation (for the temperate zone summer, there is a drop of about 0.6 °C for every 100 m above sea level [56]). Thus, thermophilous (warm-adapted) species (i.e., plants with high EIVs for temperature), which should dominate low-elevation communities, are expected to be replaced by species with progressively lower EIVs (from mesophilous species, adapted to intermediate conditions, to cryophilous species, i.e., cold-adapted species).
- (2)
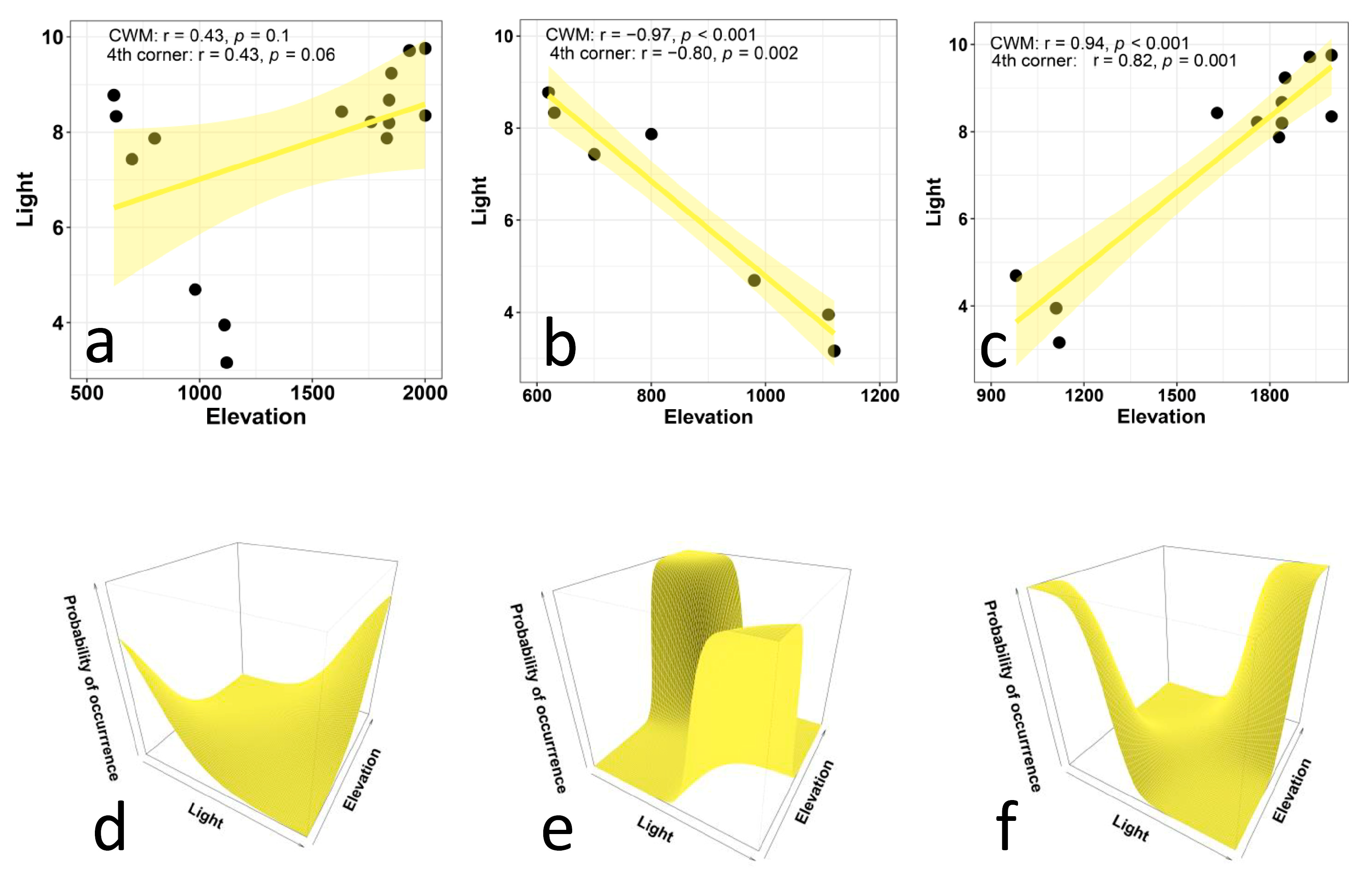
- The EIVs for light should increase with elevation, because light intensity (solar radiation) tends to increase with elevation. Lower air density and particulate matter at higher altitudes translate into greater solar radiation [56]. Additionally, with increasing elevation, vegetation becomes sparse and reduced to few herbaceous species [59]. This means that the shadow provided by trees is progressively reduced and eventually lacking. Therefore, sciophilous species (i.e., shade-loving plants) are expected to be replaced by progressively more heliophilous species (i.e., species adapted to higher levels of direct sunlight).
- (3)
- The EIVs for moisture should increase with elevation, because, at least in the temperate zone, precipitation tends to increase with elevation, which should translate into a higher soil moisture [56].
- (4)
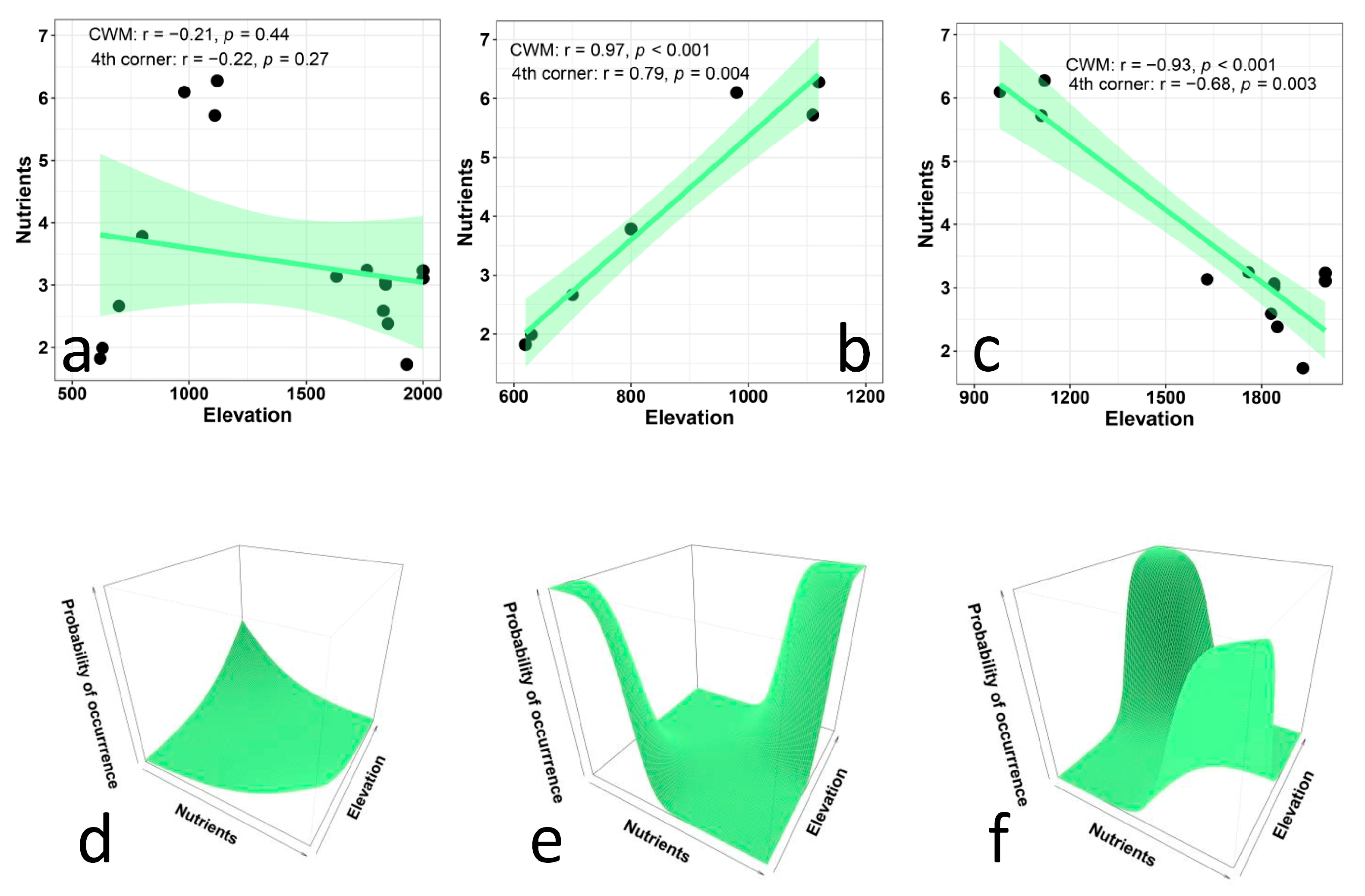
- The EIVs for nutrients should decrease with elevation because soils become less fertile at higher elevations. With an increasing elevation, soil decomposition becomes slower, and since higher slopes tend to become progressively steeper, rain and melting snow carry away more and more soil, making soil thinner and less fertile [56,59]. Thus, species that need a high concentration of soil nutrients are expected to be progressively replaced by those able to survive in soils with low levels of phosphorous, nitrogen, and organic matter.
- (5)
- The EIVs for soil reaction (pH) should increase with elevation because of decreasing values of soil pH. Soil pH tends to decrease with elevation due to the slow decomposition of organic matter (which releases acids) and higher precipitation, which increases the leaching of basic cations [64,65,66,67,68].
- (6)
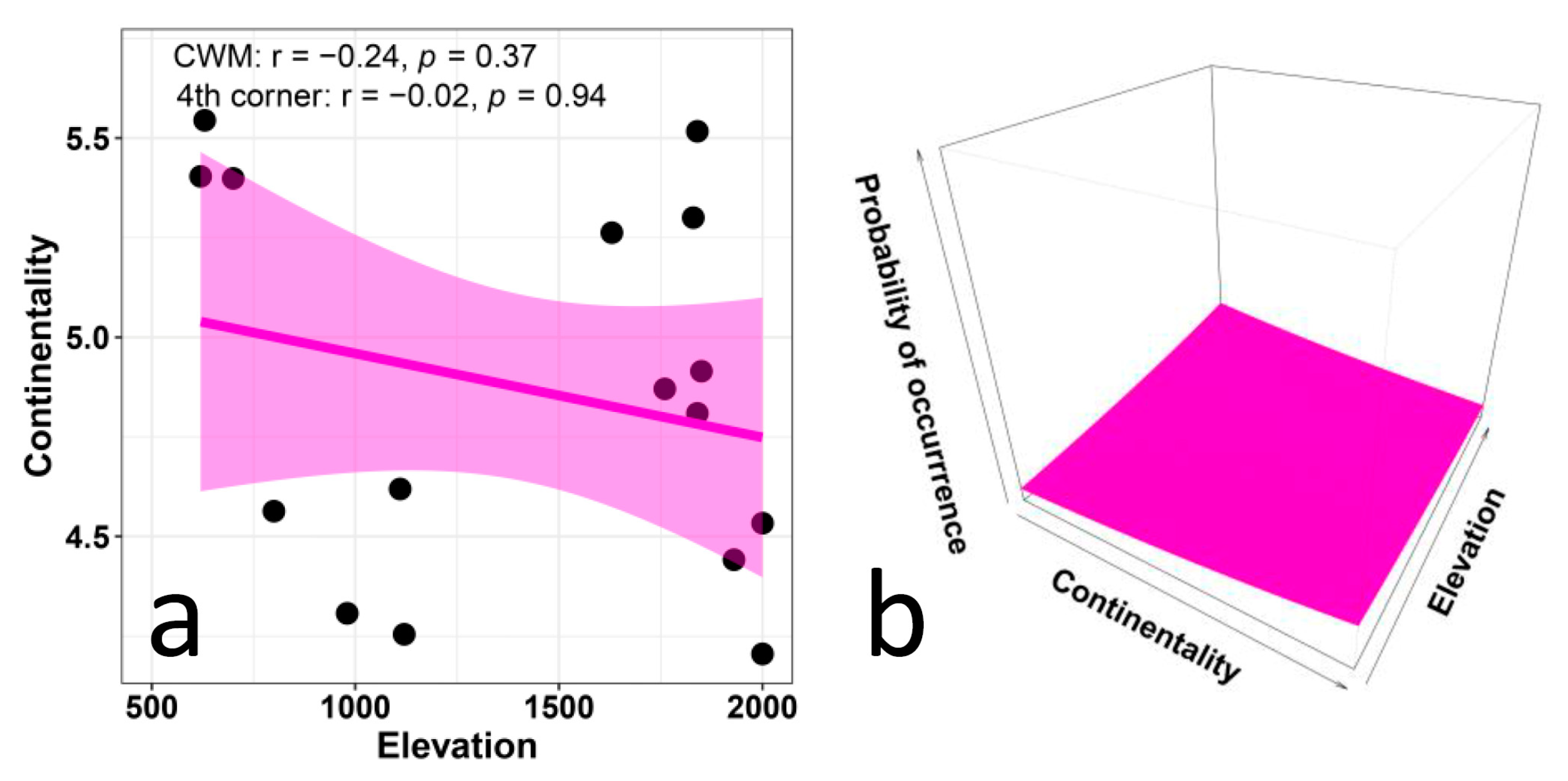
- The EIVs for continentality are not expected to show any distinct variation with elevation, since they tend to not exhibit recognizable patterns of spatial variation and dependence on environmental variables [48,69,70,71]. The concept of continentality integrates thermic and hygric gradients and may reflect geographical proximity to the ocean, as well latitudinal and altitudinal gradients, since the ecological importance of temperature increases toward higher latitudes and altitudes, while the importance of humidity increases towards lower latitudes and altitudes [69]. However, the EIVs for continentality rarely provide meaningful results and were used less frequently than any other EIVs [69]. In particular, studies using the EIVs on a large scale typically did not consider continentality, and its use in small-scale studies only provided barely interpretable results [69,70,71]. Given the very small scale of our study, we do not expect any meaningful variation of continentality values with elevation.
2. Materials and Methods
2.1. Study Area and Data Collection
2.2. Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, T.M.; Smith, R.L. Elements of Ecology, 9th ed.; Pearson Benjamin Cummings Pub Co.: San Francisco, CA, USA, 2014; pp. 1–704. [Google Scholar]
- Keddy, P.A. Plant Ecology. Origins, Processes, Consequences, 2nd ed.; Cambridge University Press: Cambridge, UK, 2017; pp. 1–624. [Google Scholar]
- Schulze, E.D.; Beck, E.; Buchmann, N.; Clemens, S.; Müller-Hohenstein, K.; Scherer-Lorenzen, M. Plant Ecology, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–926. [Google Scholar]
- ter Braak, C.J.F.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003; pp. 1–269. [Google Scholar]
- Cristóbal, E.; Ayuso, S.V.; Justel, A.; Toro, M. Robust optima and tolerance ranges of biological indicators: A new method to identify sentinels of global warming. Ecol. Res. 2014, 29, 55–68. [Google Scholar] [CrossRef]
- Keddy, P.A.; Laughlin, D.C. A Framework for Community Ecology: Species Pools, Filters, and Traits; Cambridge University Press: Cambridge, UK, 2022; pp. 1–370. [Google Scholar]
- Giller, P.S. Community Structure and the Niche; Chapman and Hall: London, UK, 1984; pp. 1–176. [Google Scholar]
- Putman, R.J. Community Ecology; Chapman & Hall: London, UK, 1994; pp. 1–178. [Google Scholar]
- Prinzing, A.; Durka, W.; Klotz, S.; Brandl, R. Geographic variability of ecological niches of plant species: Are competition and stress relevant? Ecography 2002, 25, 721–729. [Google Scholar] [CrossRef]
- Bruno, J.F.; Stachowicz, J.J.; Mark, D.; Bertness, M.D. Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Callaway, R.M. Positive Interactions and Interdependence in Plant Communities; Springer: Dordrecht, The Netherlands, 2007; pp. 1–415. [Google Scholar]
- Brooker, R.W.; Maestre, F.T.; Callaway, R.M.; Lortie, C.L.; Cavieres, L.A.; Kunstler, G.; Liancourt, P.; Tielbörger, K.; Travis, J.M.J.; Anthelme, F.; et al. Facilitation in plant communities: The past, the present, and the future. J. Ecol. 2008, 96, 18–34. [Google Scholar] [CrossRef]
- Smart, S.M.; Andrew Scott, W.; Whitaker, J.; Hill, M.O.; Roy, D.B.; Nigel Critchley, C.; Marini, L.; Evans, C.; Emmett, B.A.; Rowe, C.; et al. Empirical realized niche models for British higher and lower plants—Development and preliminary testing. J. Veg. Sci. 2010, 21, 643–656. [Google Scholar]
- Szymura, T.H.; Szymura, M.; Macioł, A. Bioindication with Ellenberg’s indicator values: A comparison with measured parameters in Central European oak forests. Ecol. Indic. 2014, 46, 495–503. [Google Scholar] [CrossRef]
- Tölgyesi, C.; Bátori, Z.; Erdős, L. Using statistical tests on relative ecological indicator values to compare vegetation units—Different approaches and weighting methods. Ecol. Indic. 2014, 36, 441–446. [Google Scholar] [CrossRef]
- Ellenberg, H. Landwirtschaftliche Pflanzensoziologie II. Wiesen und Weiden und ihre Standörtliche Bewertung; Ulmer: Stuttgart, Germany, 1952; pp. 1–143. [Google Scholar]
- Ellenberg, H. Zeigerwerte der Gefäßpflanzen Mitteleuropas; Verlag Erich Goltze KG: Göttingen, Germany, 1974; pp. 1–97. [Google Scholar]
- Ellenberg, H. Zeigerwerte der Gefäßpflanzen Mitteleuropas, 2nd ed.; Verlag Erich Goltze KG: Göttingen, Germany, 1979; pp. 1–122. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa; Verlag Erich Goltze KG: Göttingen, Germany, 1991; pp. 1–248. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa. 2. Verbesserte und Erweiterte Auflage; Verlag Erich Goltze KG: Göttingen, Germany, 1992; pp. 1–258. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa. 3. Durchgesehene Auflage; Verlag Erich Goltze KG: Göttingen, Germany, 2001; pp. 1–262. [Google Scholar]
- Zarzycki, K.; Trzcińska-Tacik, H.; Różański, W.; Szeląg, Z.; Wołek, J.; Korzeniak, U. Ecological Indicator Values of Vascular Plants of Poland. Ekologiczne Liczby Wskaźnikowe Roślin Naczyniowych Polski; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2002; pp. 1–183. [Google Scholar]
- Borhidi, A. Social behaviour types, the naturalness and relative ecological indicator values of the higher plants in theungariann flora. Acta Bot. Hungar. 1995, 39, 97–181. [Google Scholar]
- Pignatti, S.; Bianco, P.M.; Fanelli, G.; Guarino, R.; Petersen, L.; Tescarollo, P. Reliability and Effectiveness of Ellenberg’s Indices in Checking Flora and Vegetation Changes Induced by Climatic Variations, In Fingerprints of Climate Changes: Adapted Behaviour and Shifting Species Ranges; Walter, G.R., Burga, C.A., Edwards, P.J., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2001; pp. 281–304. [Google Scholar]
- Pignatti, S.; Menegoni, P.; Pietrosanti, S. Valori di biondicazione delle piante vascolari della flora d’Italia. Braun-Blanquetia 2005, 39, 1–97. [Google Scholar]
- Böhling, N.; Greuter, W.; Raus, T. Indicator values for vascular plants in the Southern Aegean (Greece). Braun-Blanquetia 2002, 32, 1–109. [Google Scholar]
- Gégout, J.C.; Krizova, E. Comparison of indicator values of forest understory plant species in Western Carpathians (Slovakia) and Vosges Mountains (France). For. Ecol. Manag. 2003, 182, 1–11. [Google Scholar]
- Hill, M.O.; Mountford, J.O.; Roy, D.B.; Bunce, R.G.H. Ellenberg’s Indicator Values for British Plants; Institute of Terrestrial Ecology: Huntington, UK, 1999; pp. 1–46. [Google Scholar]
- Hill, M.O.; Roy, D.B.; Owen Mountford, J.; Bunce, R.G.H. Extending Ellenber’s Indicator Values to a New Area: An Algorithmic Approach. J. Appl. Ecol. 2000, 37, 3–15. [Google Scholar] [CrossRef]
- Lawesson, J.E.; Fosaa, A.M.; Olsen, E. Calibration of Ellenberg indicator values for Faroe islands. Appl. Veg. Sci. 2003, 6, 53–62. [Google Scholar] [CrossRef]
- Kosić, I.V.; Juračak, J.; Łuczaj, Ł. Using Ellenberg-Pignatti values to estimate habitat preferences of wild food and medicinal plants: An example from northeastern Istria (Croatia). J. Ethnobiol. Ethnomedicine 2017, 13, 31. [Google Scholar] [CrossRef] [PubMed]
- Hedwall, P.-O.; Brunet, J.; Diekmann, M. With Ellenberg indicator values towards the north: Does the indicative power decrease with distance from Central Europe? J. Biogeogr. 2019, 46, 1041–1053. [Google Scholar] [CrossRef]
- Chytrý, M.; Tichý, L.; Dřevojan, P.; Sádlo, J.; Zelený, D. Ellenberg-type indicator values for the Czech flora. Preslia 2018, 90, 83–103. [Google Scholar] [CrossRef]
- Hill, M.O.; Carey, P.D. Prediction of yield in the Rothamsted Park Grass Experiment by Ellenberg indicator values. J. Veg. Sci. 1997, 8, 579–586. [Google Scholar] [CrossRef]
- Van Dobben, H.F.; ter Braak, C.J.F.; Dirkse, G.M. Undergrowth as a biomonitor for deposition of nitrogen and acidity in pine forest. For. Ecol. Manag. 1999, 114, 83–95. [Google Scholar] [CrossRef]
- Bergès, L.; Gégout, J.C.; Franc, A. Can understory vegetation accurately predict site index? A comparative study using floristic and abiotic indices in sessile oak (Quercus petraea Liebl.) stands in northern France. Ann. For. Sci. 2006, 63, 31–42. [Google Scholar] [CrossRef]
- Wagner, M.; Kahmen, A.; Schlumprecht, H.; Audorff, V.; Perner, J.; Buchmann, N.; Weisser, W.W. Prediction of herbage yield in grassland: How well do Ellenberg N-values perform? Appl. Veg. Sci. 2007, 10, 15–24. [Google Scholar] [CrossRef]
- Axmanová, I.; Tichý, L.; Fajmonová, Z.; Hájková, P.; Hettenbergerová, E.; Li, C.F.; Merunková, K.; Najzchlebová, M.; Otýpková, Z.; Vymazalová, M.; et al. Estimation of herbaceous biomass from species composition and cover. Appl. Veg. Sci. 2012, 15, 580–589. [Google Scholar] [CrossRef]
- Häring, T.; Reger, B.; Ewald, J.; Hothorn, T.; Schröder, B. Predicting Ellenberg’s soil moisture indicator value in the Bavarian Alps using additive georegression. Appl. Veg. Sci. 2012, 16, 110–121. [Google Scholar] [CrossRef]
- Hawkes, J.C.; Pyatt, D.G.; White, I.M.S. Using Ellenberg Indicator Values to assess soil quality in British forests from ground vegetation: A pilot study. J. App. Ecol. 1997, 34, 375–387. [Google Scholar] [CrossRef]
- Ertsen, A.C.D.; Alkemade, J.R.M.; Wassen, M.J. Calibrating Ellenberg indicator values for moisture, acidity, nutrient availability and salinity in the Netherlands. Plant Ecol. 1998, 135, 113–124. [Google Scholar] [CrossRef]
- Diekmann, M. Species indicator values as an important tool in applied plant ecology—A review. Basic Appl. Ecol. 2003, 4, 493–506. [Google Scholar] [CrossRef]
- Wamelink, G.W.; Goedhart, P.W.; Van Dobben, H.F.; Berendse, F. Plant species as predictors of soil pH: Replacing expert judgement with measurements. J. Veg. Sci. 2005, 16, 461–470. [Google Scholar] [CrossRef]
- Bartelheimer, M.; Poschlod, P. Functional characterizations of Ellenberg indicator values—A review on ecophysiological determinants. Funct. Ecol. 2016, 30, 506–516. [Google Scholar] [CrossRef]
- Persson, S. Ecological Indicator Values as an Aid in the Interpretation of Ordination Diagrams. J. Ecol. 1981, 69, 71–84. [Google Scholar] [CrossRef]
- Major, J.; Rejmanek, M. Amelanchier alnifolia vegetation in eastern Idaho, USA and its environmental relationships. Vegetatio 1992, 98, 141–156. [Google Scholar] [CrossRef]
- Thompson, K.; Hodgson, J.G.; Grime, J.P.; Rorison, I.H.; Band, S.R.; Spencer, R.E. Ellenberg numbers revisited. Phytocoenologia 1993, 23, 277–289. [Google Scholar] [CrossRef]
- Marcenò, C.; Guarino, R.A. A test on Ellenberg indicator values in the Mediterranean evergreen woods (Quercetea ilicis). Rend. Fis. Acc. Lincei 2015, 26, 345–356. [Google Scholar] [CrossRef]
- Schaffers, A.P.; Sýkora, K.V. Reliability of Ellenberg indicator values for moisture, nitrogen and soil reaction: A comparison with field measurements. J. Veg. Sci. 2000, 11, 225–244. [Google Scholar] [CrossRef]
- Wamelink, G.W.W.; Joosten, V.; van Dobben, H.H.; Berendse, F. Validity of Ellenberg indicator values judged from physico-chemical field measurements. J. Veg. Sci. 2002, 13, 269–278. [Google Scholar] [CrossRef]
- Chytrý, M.; Tichý, L.; Roleček, J. Local and regional patterns of species richness in Central European vegetation types along the pH/calcium gradient. Folia Geobot. 2002, 38, 429–442. [Google Scholar] [CrossRef]
- Sørensen, M.M.; Tybirk, K. Vegetation analysis along a successional gradient from heath to oak forest. Nord. J. Bot. 2000, 20, 537–546. [Google Scholar] [CrossRef]
- Lososová, Z.; Chytrý, M.; Cimalová, S.; Kropáč, Z.; Otýpková, Z.; Pyšek, P.; Tichý, L. Weed vegetation of arable land in Central Europe: Gradients of diversity and species composition. J. Veg. Sci. 2004, 15, 415–422. [Google Scholar] [CrossRef]
- Fraaije, R.G.A.; ter Braak, C.J.F.; Verduyn, B.; Breeman, L.B.S.; Verhoeven, J.T.A.; Soons, M.B. Early plant recruitment stages set the template for the development of vegetation patterns along a hydrological gradient. Funct. Ecol. 2015, 29, 971–980. [Google Scholar] [CrossRef]
- Kutbay, H.; Surmen, B. Ellenberg ecological indicator values, tolerance values, species niche models for soil nutrient availability, salinity, and pH in coastal dune vegetation along a landward gradient (Euxine, Turkey). Turk. J. Bot. 2022, 46, 346–360. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems, 2nd ed.; Springer: Berlin, Germany, 2003; pp. 1–344. [Google Scholar]
- Sundqvist, M.K.; Sanders, N.J.; Wardle, D.A. Community and ecosystem responses to elevational gradients: Processes, mechanisms, and insights for global change. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 261–280. [Google Scholar] [CrossRef]
- Fattorini, S.; Di Biase, L.; Chiarucci, A. Recognizing and interpreting vegetational belts: New wine in the old bottles of a von Humboldt’s legacy. J. Biogeogr. 2019, 46, 1643–1651. [Google Scholar] [CrossRef]
- Fattorini, S.; Mantoni, C.; Di Biase, L.; Pace, L. Mountain biodiversity and sustainable development. In Encyclopedia of the UN Sustainable Development Goals. Life on Land; Leal Filho, W., Azul, A., Brandli, L., Özuyar, P., Wall, T., Eds.; Springer: Cham, Switzerland, 2020; pp. 1–31. [Google Scholar]
- Fattorini, S.; Mantoni, C.; Di Biase, L.; Strona, G.; Pace, L.; Biondi, M. Elevational patterns of generic diversity in the tenebrionid beetles (Coleoptera Tenebrionidae) of Latium (Central Italy). Diversity 2020, 12, 47. [Google Scholar] [CrossRef]
- Moradi, H.; Fattorini, S.; Oldeland, J. Influence of elevation on the species–area relationship. J. Biogeogr. 2020, 47, 2029–2041. [Google Scholar] [CrossRef]
- Bruelheide, H.; Dengler, J.; Purschke, O.; Lenoir, J.; Jiménez-Alfaro, B.; Hennekens, S.M.; Botta-Dukát, Z.; Chytrý, M.; Field, R.; Jansen, F.; et al. Global trait-environment relationships of plant communities. Nat. Ecol. Evol. 2018, 2, 1906–1917. [Google Scholar] [CrossRef]
- Boonman, C.C.F.; Santini, L.; Robroek, B.J.M.; Hoeks, S.; Kelderman, S.; Dengler, J.; Bergamini, A.; Biurrun, I.; Carranza, M.L.; Cerabolini, B.E.L.; et al. Plant functional and taxonomic diversity in European grasslands along climatic gradients. J. Veg. Sci. 2021, 32, e13027. [Google Scholar] [CrossRef]
- Hillman, P. The Basics of Biogeography; Hodder & Stoughton: London, UK, 1985; pp. 1–48. [Google Scholar]
- Jobbagy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Smith, J.L.; Halvorson, J.J.; Bolton, H., Jr. Soil properties and microbial activity across a 500 m elevation gradient in a semi-arid environment. Soil Biol. Biochem. 2002, 34, 1749–1757. [Google Scholar] [CrossRef]
- Northcott, M.L.; Gooseff, M.N.; Barrett, J.E.; Zeglin, L.H.; Takacs-Vesbach, C.D.; Humphrey, J. Hydrologic characteristics of lake and stream-side riparian margins in the McMurdo Dry Valleys, Antarctica. Hydrol. Process 2009, 23, 1255–1267. [Google Scholar] [CrossRef]
- Jeyakumar, S.P.; Dash, B.; Singh, A.K.; Suyal, D.C.; Soni, R. Nutrient cycling at higher altitudes. In Microbiological Advancements for Higher Altitude Agro-Ecosystems & Sustainability; Goel, R., Soni, R., Suyal, D.C., Eds.; Springer Nature: Singapore, Singapore, 2020; pp. 293–305. [Google Scholar]
- Berg, C.; Welk, E.; Jäger, E.J. Revising Ellenberg’s indicator values for continentality based on global vascular plant species distribution. Appl. Veg. Sci. 2017, 20, 482–493. [Google Scholar] [CrossRef]
- Fischer, H.S.; Michler, B.; Ewald, J. Environmental, spatial and structural components in the composition of mountain forest in the Bavarian alps. Folia Geobot. 2014, 49, 361–384. [Google Scholar] [CrossRef]
- Koch, M.; Jurasinski, G. Four decades of vegetation development in a percolation mire complex following intensive drainage and abandonment. Plant Ecol. Diver. 2014, 8, 49–60. [Google Scholar] [CrossRef]
- Pirone, G. Aspetti della vegetazione della riserva naturale guidata monte Genzana e Alto Gizio. In Aree protette in Abruzzo. Contributi alla Conoscenza Naturalistica ed Ambientale; Burri, E., Ed.; Carsa Edizioni: Pescara, Italy, 1998; pp. 120–139. [Google Scholar]
- Piano di Gestione del Sito di Interesse Comunitario IT 7110100 Monte Genzana. Available online: http://www.riservagenzana.it/pdf/Pdg_Monte_Genzana.pdf. (accessed on 1 December 2022).
- Di Biase, L.; Pace, L.; Mantoni, C.; Fattorini, S. Variations in plant richness, biogeographical composition, and life forms along an elevational gradient in a Mediterranean mountain. Plants 2021, 10, 2090. [Google Scholar] [CrossRef] [PubMed]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora dʹItalia, 2nd ed.; Edagricole-New Business Media: Bologna, Italy, 2017–2019; Volume 1–4. [Google Scholar]
- Prodromo della Vegetazione d’Italia. Available online: https://www.prodromo-vegetazione-italia.org/introduzione (accessed on 15 December 2022).
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde, 3rd ed.; Springer: Vienna, Austria, 1964; p. 631. [Google Scholar]
- Hennekens, S.M.; Schaminée, J.H.J. Turboveg, a Comprehensive Data Base Management System for Vegetation Data. J. Veg. Sci. 2001, 12, 589–591. [Google Scholar] [CrossRef]
- Pätsch, R.; Jašková, A.; Chytrý, M.; Kucherov, I.B.; Schaminée, J.H.J.; Bergmeier, E.; Janssen, J.A.M. Making them visible and usable—Vegetation-plot observations from Fennoscandia based on historical species-quantity scales. Appl. Veg. Sci. 2019, 22, 465–473. [Google Scholar] [CrossRef]
- Tichý, L.; Hennekens, S.M.; Novák, P.; Rodwell, J.S.; Schaminée, J.H.J.; Chytrý, M. Optimal transformation of species cover for vegetation classification. Appl. Veg. Sci. 2020, 23, 471–722. [Google Scholar] [CrossRef]
- Guarino, R.; Domina, G.; Pignatti, S. Ellenberg’s Indicator values for the Flora of Italy—First update: Pteridophyta, Gymnospermae and Monocotyledoneae. Flora Mediterr. 2012, 22, 197–209. [Google Scholar] [CrossRef]
- Garnier, E.; Cortez, J.; Neill, C.; Toussaint, J.-P.; Billès, G.; Navas, M.-L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Ricotta, C.; Moretti, M. CWM and Rao’s quadratic diversity: A unified framework for functional ecology. Oecologia 2011, 167, 181–188. [Google Scholar] [CrossRef]
- Di Biase, L.; Fattorini, S.; Cutini, M.; Bricca, A. The role of inter-and intraspecific variations in grassland plant functional traits along an elevational gradient in a Mediterranean mountain area. Plants 2021, 10, 359. [Google Scholar] [CrossRef]
- Chapman, H.; Cordeiro, N.J.; Dutton, P.; Wenny, D.; Kitamura, S.; Kaplin, B.; Melo, F.P.; Lawes, M.J. Seed-dispersal ecology of tropical montane forests. J. Trop. Ecol. 2016, 32, 437–454. [Google Scholar] [CrossRef]
- Funk, J.L.; Larson, J.E.; Ames, G.M.; Butterfield, B.J.; Cavender-Bares, J.; Firn, J.; Laughlin, D.C.; Sutton-Grier, A.E.; Williams, L.; Wright, J. Revisiting the holy grail: Using plant functional traits to understand ecological processes. Biol. Rev. 2017, 92, 1156–1173. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Peres-Neto, P.R.; Dray, S. Simple parametric tests for trait-environment association. J. Veg. Sci. 2018, 29, 801–811. [Google Scholar] [CrossRef]
- Hanif, M.A.; Yu, Q.; Rao, X.; Shen, W. Disentangling the Contributions of Plant Taxonomic and Functional Diversities in Shaping Aboveground Biomass of a Restored Forest Landscape in Southern China. Plants 2019, 8, 612. [Google Scholar] [CrossRef]
- Rolhauser, A.G.; Waller, D.M.; Tucker, C.M. Complex trait–environment relationships underlie the structure of forest plant communities. J. Ecol. 2021, 109, 3794–3806. [Google Scholar] [CrossRef]
- Cheng, X.; Ping, T.; Li, Z.; Tian, W.; Hairong, H.; Epstein, H.E. Effects of environmental factors on plant functional traits across different plant life forms in a temperate forest ecosystem. New For. 2022, 53, 125–142. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Peres-Neto, P.R.; Dray, S. A critical issue in model-based inference for studying trait-based community assembly and a solution. PeerJ 2017, 5, e2885. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.E.D.; Damschen, E.I.; Ives, A.R. Functional traits and community composition: A comparison among community-weighted means, weighted correlations, and multilevel models. Methods Ecol. Evol. 2019, 10, 415–425. [Google Scholar] [CrossRef]
- Dray, S.; Legendre, P. Testing the species traits–environment relationships: The fourth-corner problem revisited. Ecology 2008, 89, 3400–3412. [Google Scholar] [CrossRef] [PubMed]
- Peres-Neto, P.R.; Dray, S.; ter Braak, C.J.F. Linking trait variation to the environment: Critical issues with community-weighted mean correlation resolved by the fourth-corner approach. Ecography 2017, 40, 806–816. [Google Scholar] [CrossRef]
- Brown, A.M.; Warton, D.I.; Andrew, N.R.; Binns, M.; Cassis, G.; Gibb, H. The fourth-corner solution—Using predictive models to understand how species traits interact with the environment. Methods Ecol. Evol. 2014, 5, 344–352. [Google Scholar] [CrossRef]
- Laughlin, D.C.; Gremer, J.R.; Adler, P.B.; Mitchell, R.M.; Moore, M.M. The Net Effect of Functional Traits on Fitness. Trends Ecol. Evol. 2020, 35, 1037–1047. [Google Scholar] [CrossRef] [PubMed]
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: New York, NY, USA, 2006; pp. 1–648. [Google Scholar]
- Laughlin, D.C.; Strahan, R.T.; Adler, P.B.; Moore, M.M. Survival rates indicate that correlations between community weighted mean traits and environments can be unreliable estimates of the adaptive value of traits. Ecol. Lett. 2018, 21, 411–421. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: https://www.R-project.org (accessed on 31 August 2021).
- Laughlin, D. R Code and Data for “A Framework for Community Ecology: Species Pools, Traits, and Filter” by Paul Keddy and Daniel Laughlin. Available online: https://github.com/danielLaughlin/CommunityEcology/blob/master/communityEcology.R (accessed on 20 September 2022).
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R Package Version 1.0-12.1. 2014. Available online: https://cran.r-project.org/web/packages/FD/index.html (accessed on 16 April 2010).
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. ‘Vegan’: Community Ecology Package. R Package Version 2.6–4. Available online: https://CRAN.R-project.org/package=vegan (accessed on 20 September 2022).
- Dray, S.; Dufour, A.-B.; Thioulouse, J. ade4: Analysis of Ecological Data: Exploratory and Euclidean Methods in Environmental Sciences.R Package Version 1.7–20. Available online: https://cran.r-project.org/web/packages/ade4/index.html (accessed on 20 September 2022).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. lme4: Linear Mixed-Effects Models Using Eigen and S4. R Package Version 1.1-31. 2021. Available online: https://cran.r-project.org/web/packages/lme4/ (accessed on 20 September 2022).
- Lüdecke, D.; Makowski, D.; Ben-Shacar, M.S.; Patil, I.; Waggoner, P.; Wiernik, B.M. performance: Assessment of Regression Models Performance. R Package Version 0.10.1. Available online: https://cran.r-project.org/web/packages/performance/index.html (accessed on 20 September 2022).
- Chelli, S.; Marignani, M.; Barni, E.; Petraglia, A.; Puglielli, G.; Wellstein, C.; Acosta, A.T.R.; Bolpagni, R.; Bragazza, L.; Campetella, G.; et al. Plant-environment interactions through a functional traits perspective: A review of Italian studies. Plant Biosyst. 2018, 153, 853–869. [Google Scholar] [CrossRef]
- Boyle, J.R. Forest Soils. In Encyclopedia of Soils in the Environment; Hillel, D., Ed.; Elsevier: Oxford, UK, 2005; pp. 73–79. [Google Scholar]
- Ranger, J. Forest Soils: Characteristics and Sustainability. In Soils as a Key Component of the Critical Zone 1: Functions and Services; Berthelin, J., Valentin, C., Munch, J.C., Eds.; ISTE: London, UK, 2018; Volume 1, pp. 1–320. [Google Scholar]
- Bhandari, J.; Zhang, Y. Effect of altitude and soil properties on biomass and plant richness in the grasslands of Tibet, China, and Manang District, Nepal. Ecosphere 2019, 10, e02915. [Google Scholar] [CrossRef]
- Badía, D.; Ruiz, A.; Girona, A.; Martí, C.; Casanova, J.; Ibarra, P.; Zufiaurre, R. The influence of elevation on soil properties and forest litter in the Siliceous Moncayo Massif, SW Europe. J. Mt. Sci. 2016, 13, 2155–2169. [Google Scholar] [CrossRef]
- Lawesson, J.E. pH optima for Danish forest species compared with Ellenberg reaction values. Folia Geobot. 2003, 38, 403–418. [Google Scholar] [CrossRef]
- Balkovič, J.; Kollár, J.; Šimonovič, V. Experience with using Ellenberg’s R indicator values in Slovakia: Oligotrophic and mesotrophic submontane broad-leaved forests. Biologia 2012, 67, 474–482. [Google Scholar] [CrossRef]
- Carpenter, W.; Goodenough, A. How robust are community-based plant bioindicators? Empirical testing of the relationship between Ellenberg values and direct environmental measures in woodland communities. Comm. Ecol. 2014, 15, 1–11. [Google Scholar] [CrossRef]
- Bricca, A.; Carranza, M.L.; Varricchione, M.; Cutini, M.; Stanisci, A. Exploring Plant Functional Diversity and Redundancy of Mediterranean High-Mountain Habitats in the Apennines. Diversity 2021, 13, 466. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relevé | Elevation (m) | Description | Alliance | Order/Suborder | Class |
|---|---|---|---|---|---|
| 1 | 620 | Garrigue | Cytiso spinescentis-Satureion montanae | Cisto cretici-Ericetalia manipuliflorae | Cisto cretici-Micromerietea julianae |
| 2 | 630 | Garrigue | Cytiso spinescentis-Satureion montanae | Cisto cretici-Ericetalia manipuliflorae | Cisto cretici-Micromerietea julianae |
| 3 | 700 | Garrigue | Cytiso spinescentis-Satureion montanae | Cisto cretici-Ericetalia manipuliflorae | Cisto cretici-Micromerietea julianae |
| 4 | 800 | Xerophilous, steppic, and secondary grassland | Phleo ambigui-Bromion erecti | Phleo ambigui-Brometalia erecti | Festuco valesiacae-Brometea erecti |
| 5 | 980 | Hornbeam forest | Carpinion orientalis | Quercetalia pubescenti-petraeae | Querco roboris-Fagetea sylvaticae |
| 6 | 1110 | Mixed mesophilous forest | Tilio platyphylli-Acerion pseudoplatani | Fagetalia sylvaticae | Querco roboris-Fagetea sylvaticae |
| 7 | 1120 | Beech forest | Geranio versicoloris-Fagion sylvaticae | Fagetalia sylvaticae | Querco roboris-Fagetea sylvaticae |
| 8 | 1630 | Xerophilous, steppic, and secondary grassland | Phleo ambigui-Bromion erecti | Phleo ambigui-Brometalia erecti | Festuco valesiacae-Brometea erecti |
| 9 | 1760 | Meso-hygrophilous grassland | Ranunculo pollinensis-Nardion strictae | Nardetalia strictae | Nardetea strictae |
| 10 | 1830 | Mesophilous, acidophilous, and secondary grassland (pasture) | Ranunculo pollinensis-Nardion strictae | Nardetalia strictae | Nardetea strictae |
| 11 | 1840 | Mesophilous and sub-acidophilous grassland | Ranunculo pollinensis-Nardion strictae | Nardetalia strictae | Nardetea strictae |
| 12 | 1840 | Mesophilous, neutral-subacidophilous, and pioneer grassland | Ranunculo pollinensis-Nardion strictae | Nardetalia strictae | Nardetea strictae |
| 13 | 1850 | Mesophilous, neutral-subacidophilous, and pioneer grassland | Ranunculo pollinensis-Nardion strictae | Nardetalia strictae | Nardetea strictae |
| 14 | 1930 | Scree | Linario-Festucion dimorphae | Thlaspietalia stylosi | Thlaspietea rotundifolii |
| 15 | 2000 | Xerophilous, basophilous, pioneer, and enduring grassland | Seslerion apenninae | Seslerienalia apenninae | Festuco-Seslerietea |
| 16 | 2000 | Mesophilous and sub-acidophilous grassland | Ranunculo pollinensis-Nardion strictae | Nardetalia strictae | Nardetea strictae |
| 16 | 2000 | Mesophilous and sub-acidophilous grassland | Ranunculo pollinensis-Nardion strictae | Nardetalia strictae | Nardetea strictae |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Biase, L.; Tsafack, N.; Pace, L.; Fattorini, S. Ellenberg Indicator Values Disclose Complex Environmental Filtering Processes in Plant Communities along an Elevational Gradient. Biology 2023, 12, 161. https://doi.org/10.3390/biology12020161
Di Biase L, Tsafack N, Pace L, Fattorini S. Ellenberg Indicator Values Disclose Complex Environmental Filtering Processes in Plant Communities along an Elevational Gradient. Biology. 2023; 12(2):161. https://doi.org/10.3390/biology12020161
Chicago/Turabian StyleDi Biase, Letizia, Noelline Tsafack, Loretta Pace, and Simone Fattorini. 2023. "Ellenberg Indicator Values Disclose Complex Environmental Filtering Processes in Plant Communities along an Elevational Gradient" Biology 12, no. 2: 161. https://doi.org/10.3390/biology12020161
APA StyleDi Biase, L., Tsafack, N., Pace, L., & Fattorini, S. (2023). Ellenberg Indicator Values Disclose Complex Environmental Filtering Processes in Plant Communities along an Elevational Gradient. Biology, 12(2), 161. https://doi.org/10.3390/biology12020161