
Comparative Neuroanatomy of Pediveliger Larvae of Various Bivalves from the Sea of Japan


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Larvae
2.2. Fixation of Larvae
2.3. Immunocytochemical Staining
2.4. Antibodies
2.5. Confocal Microscopy
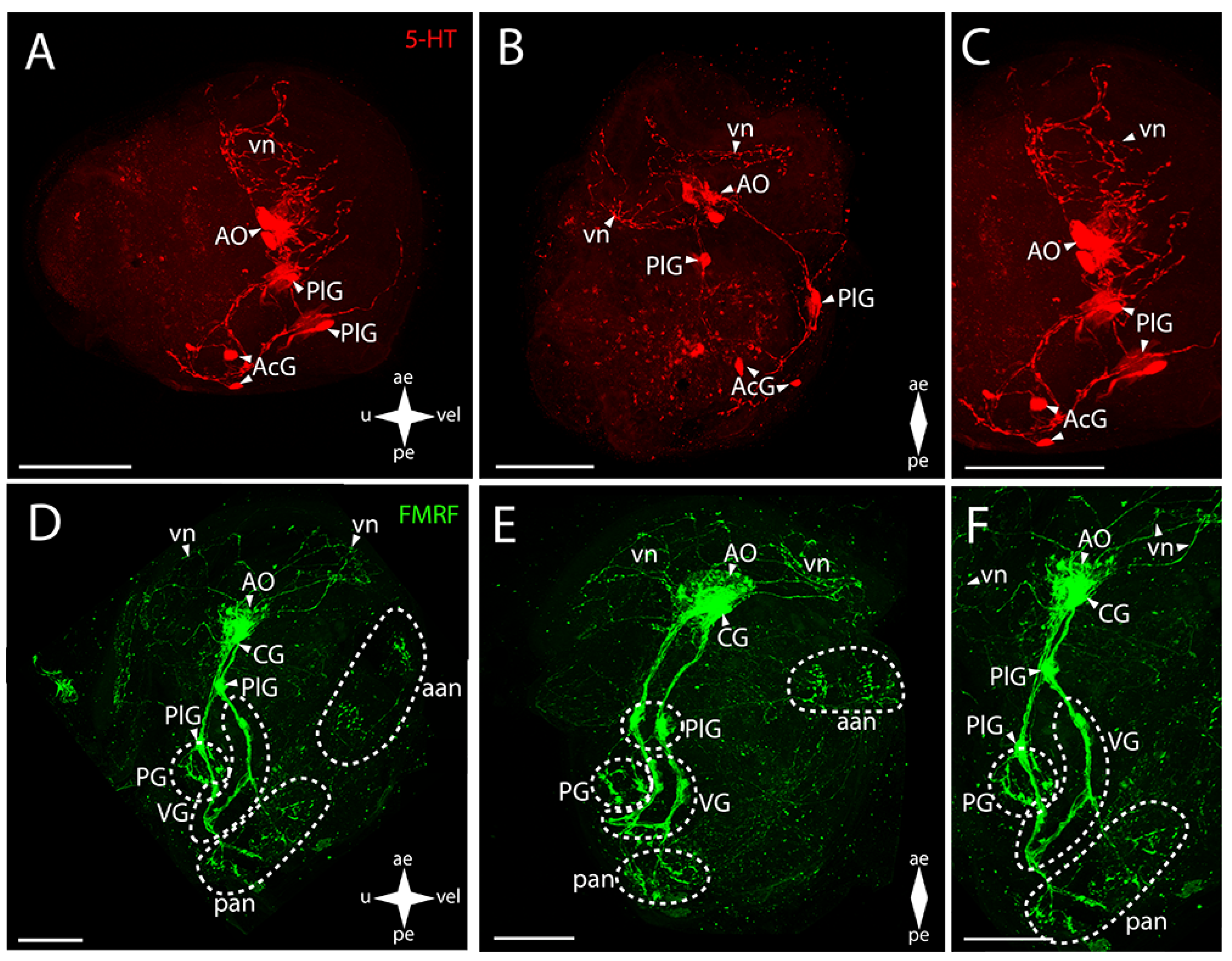
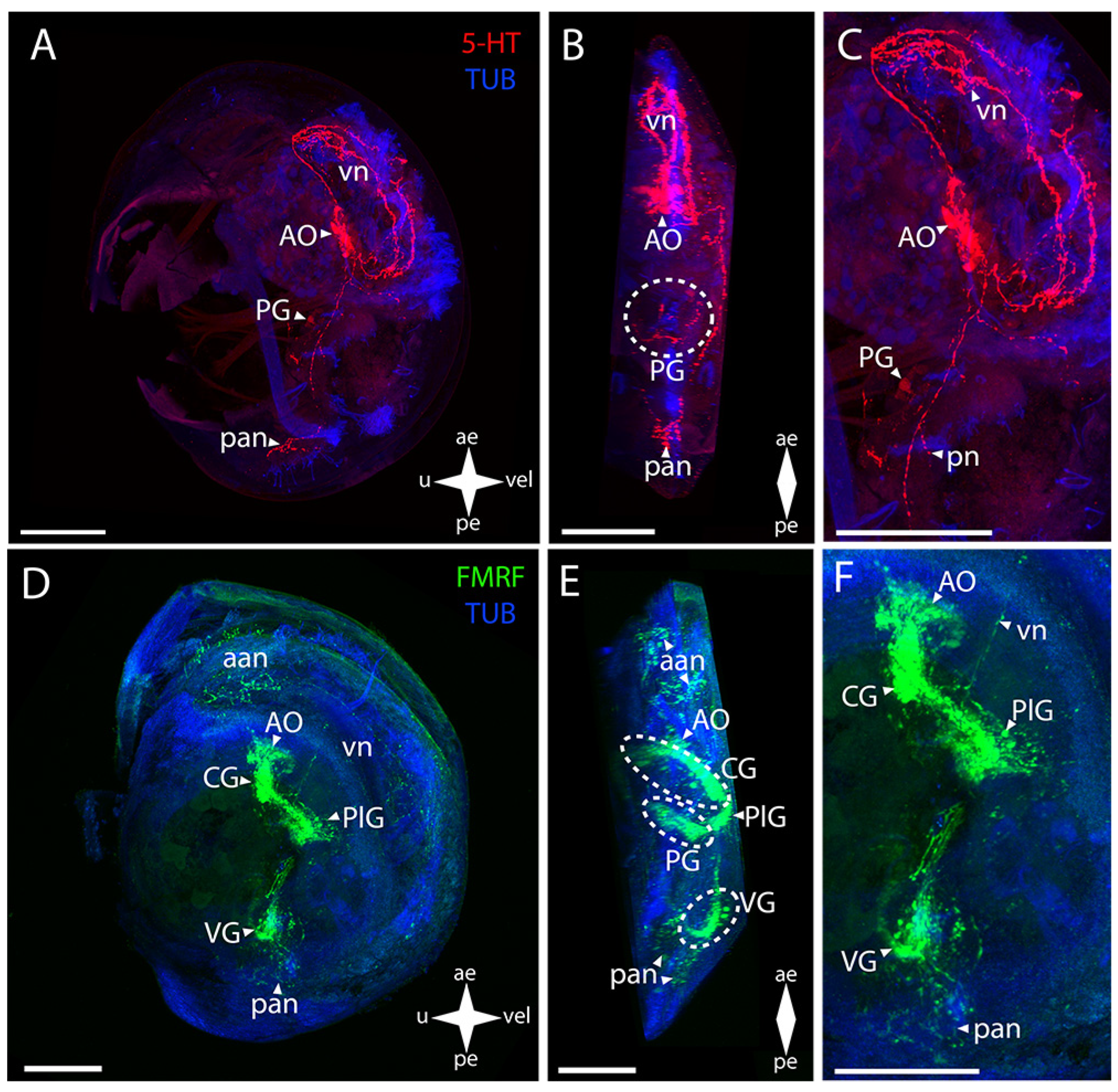
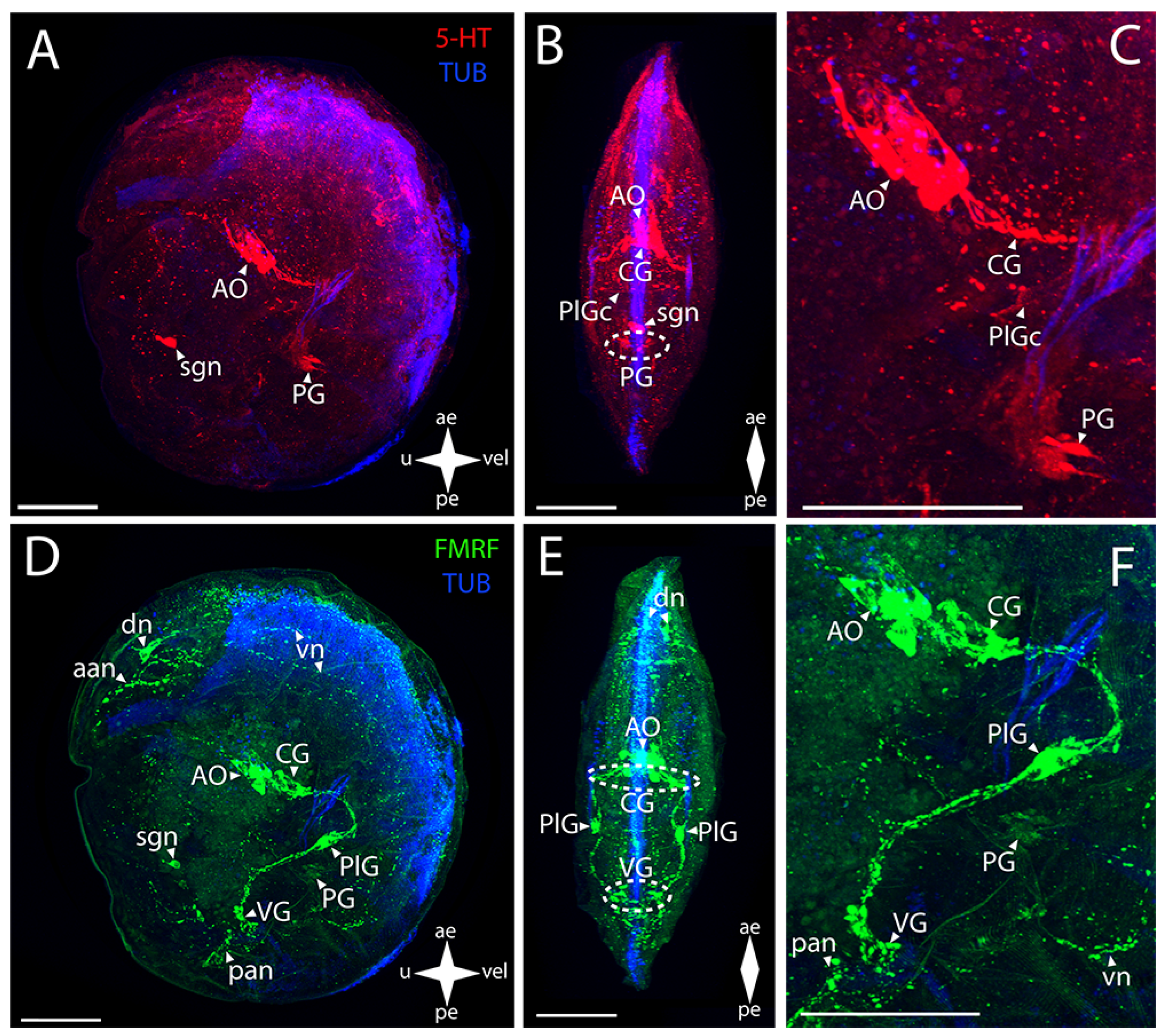
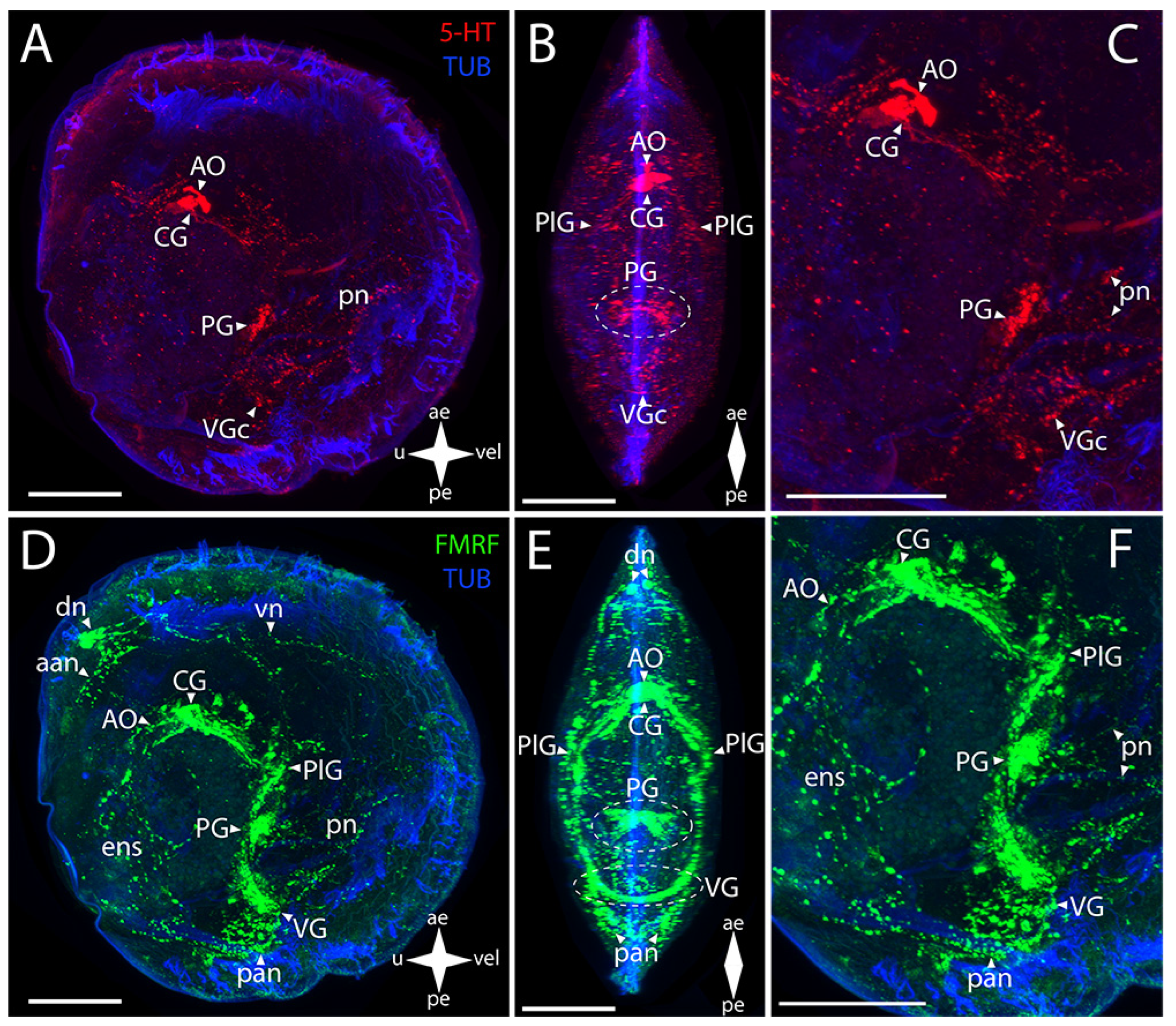
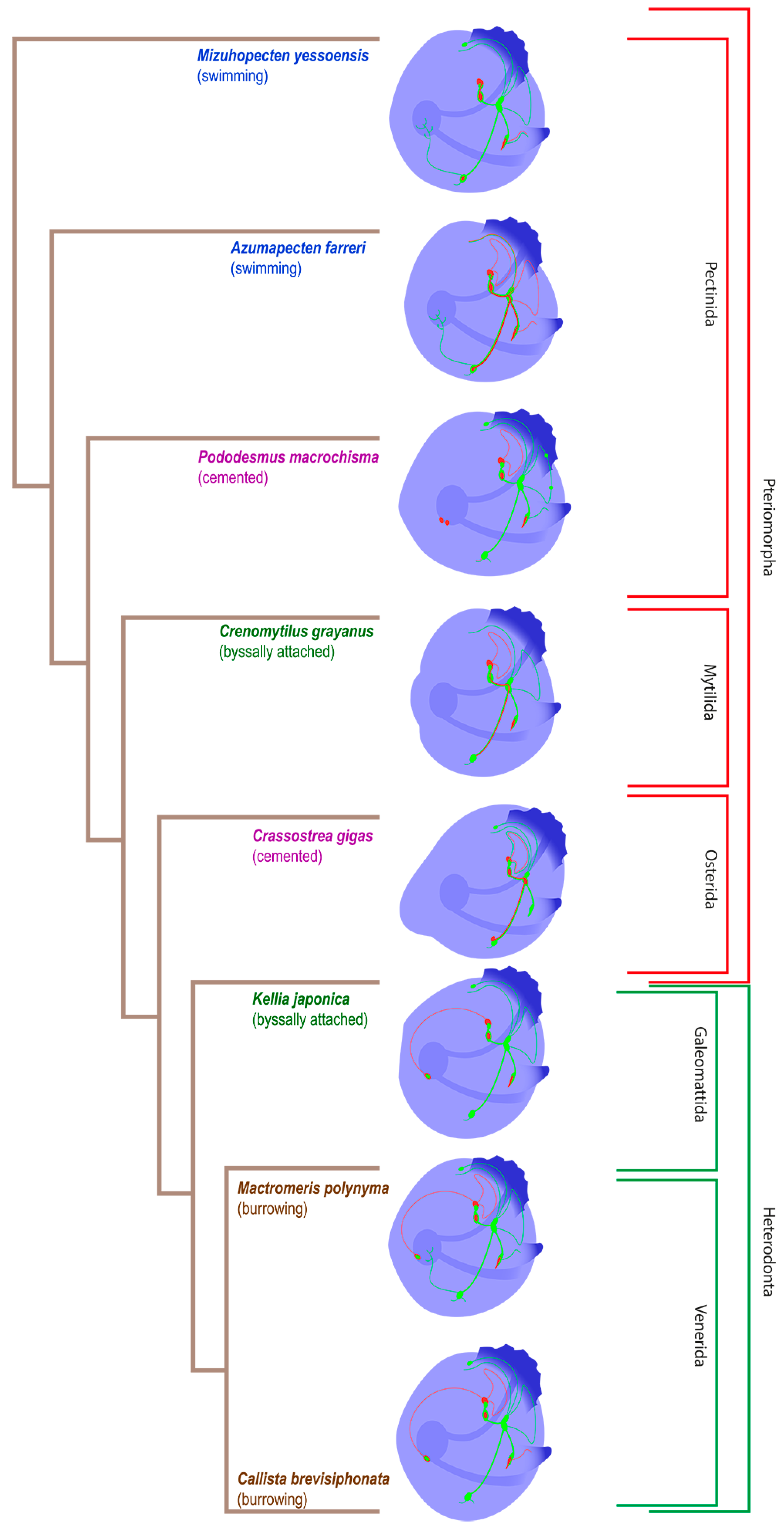
3. Results
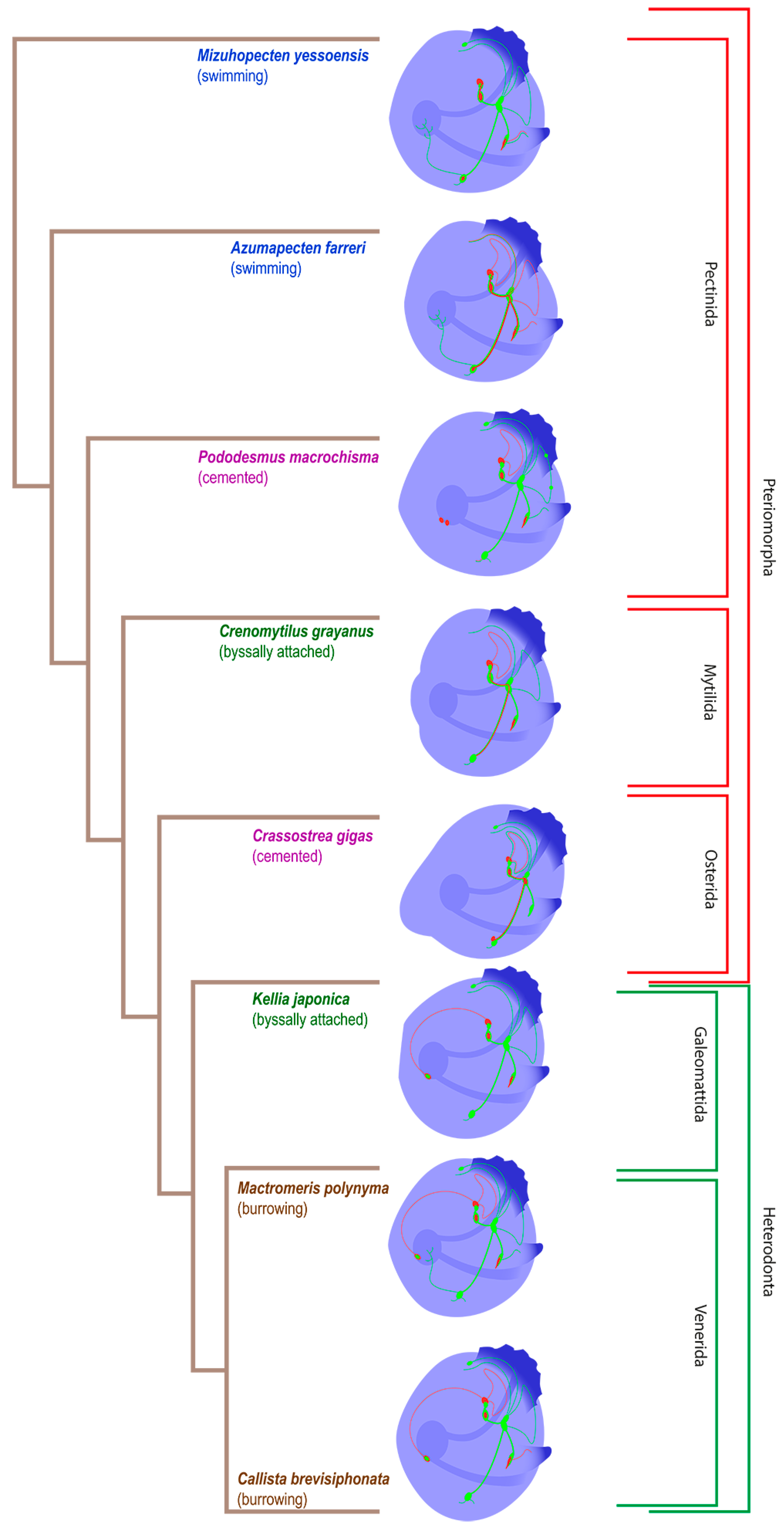
3.1. Species and Criteria
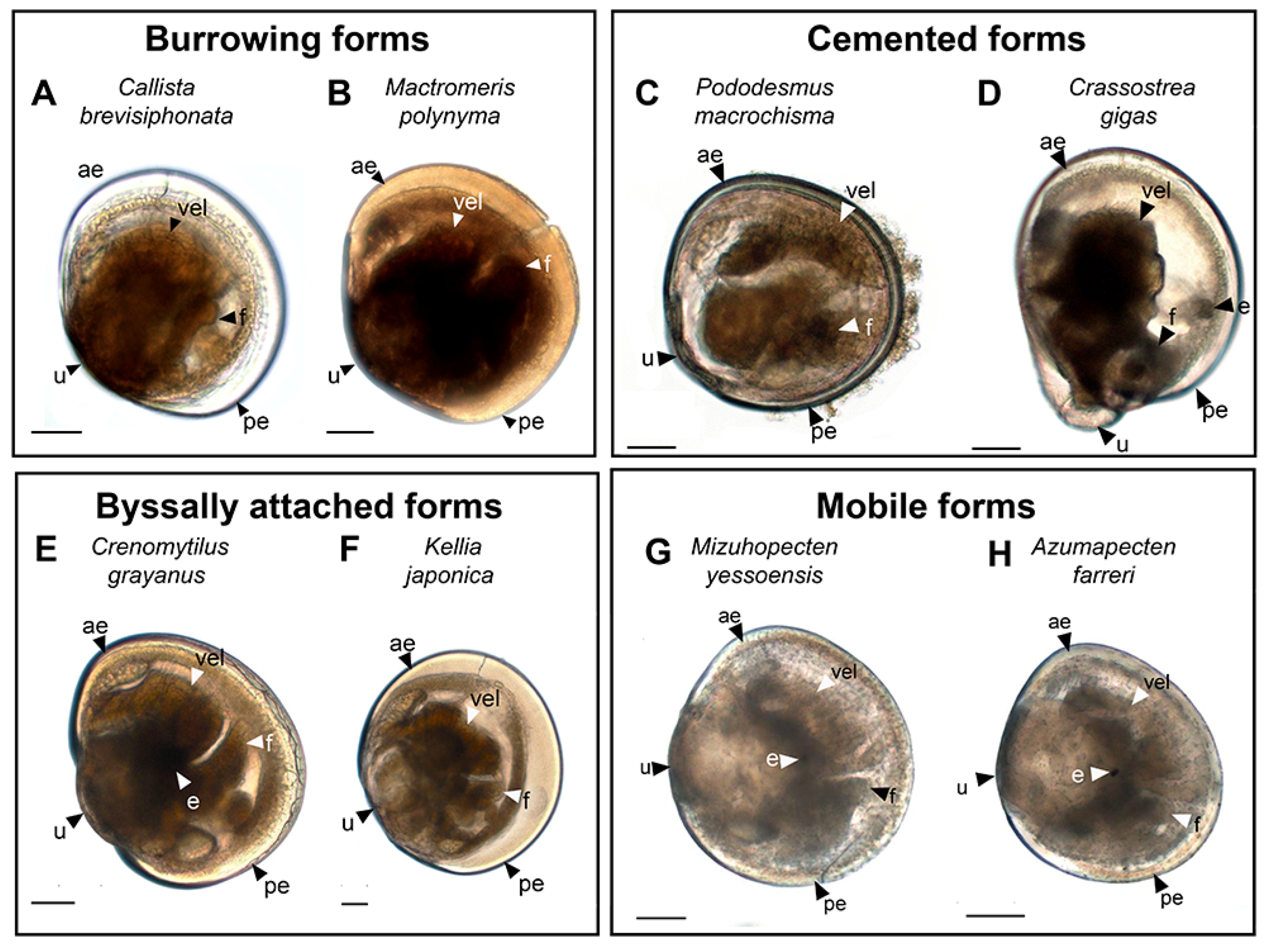
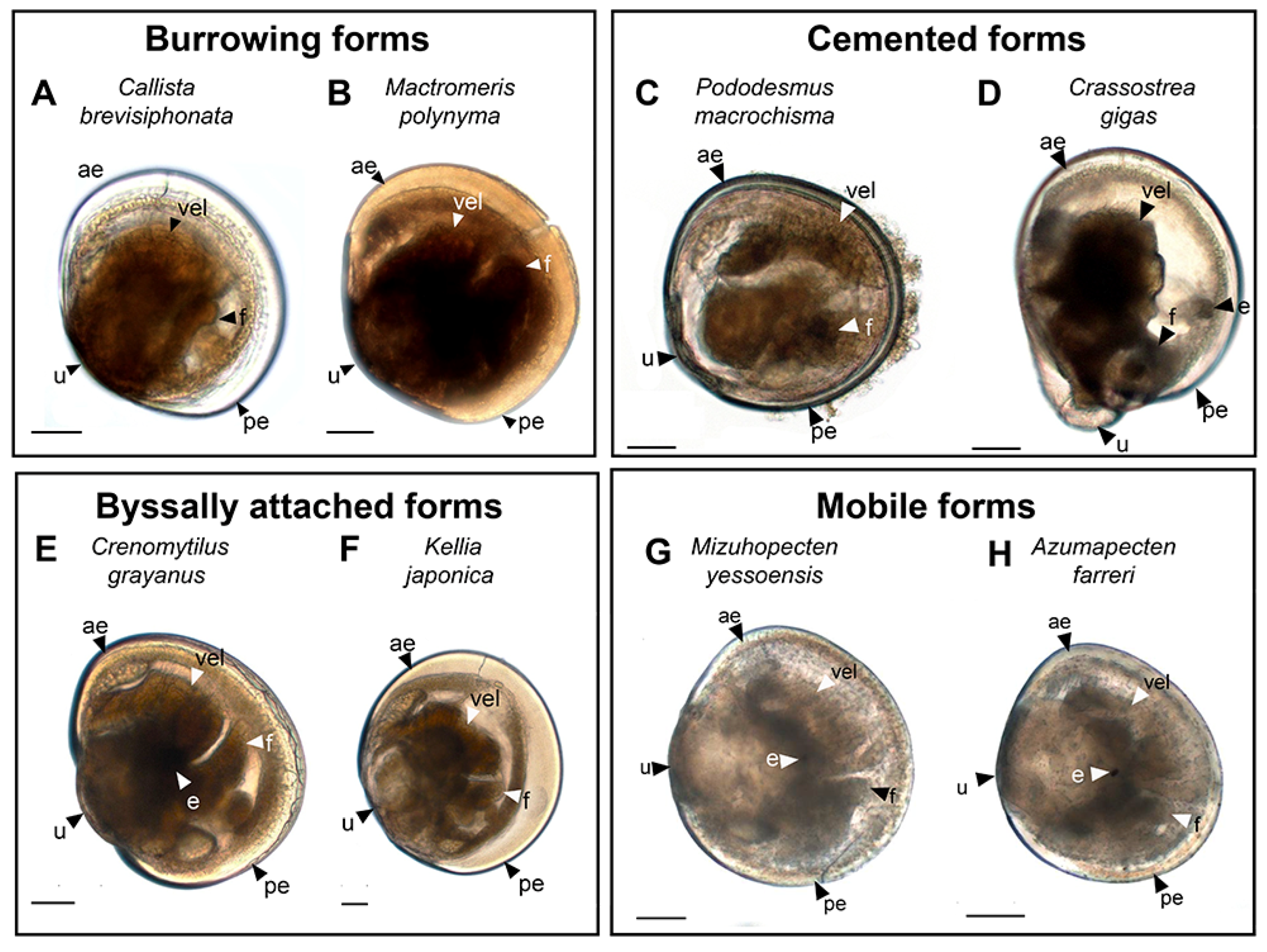
3.2. Burrowing Forms of Bivalve Mollusks
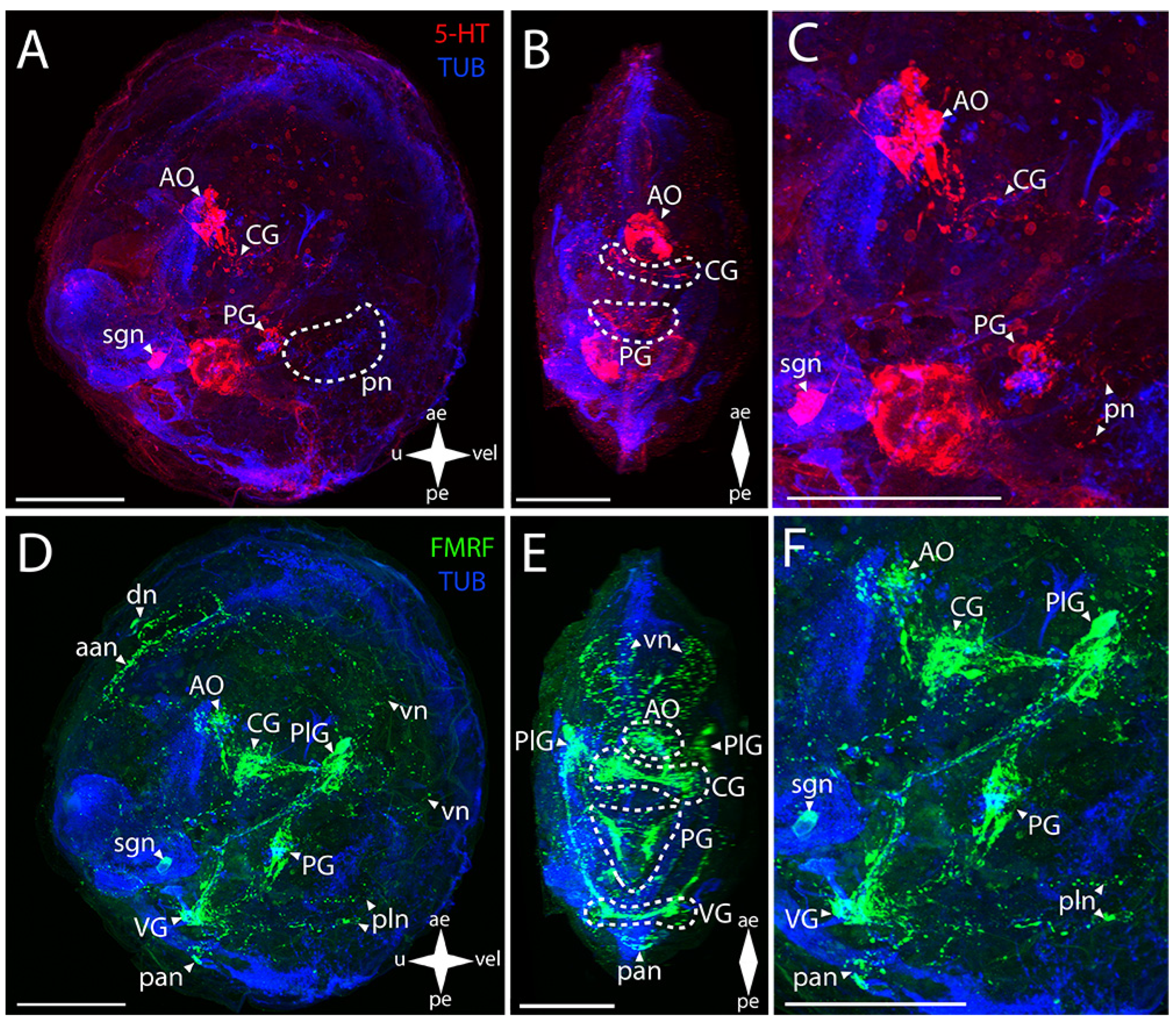
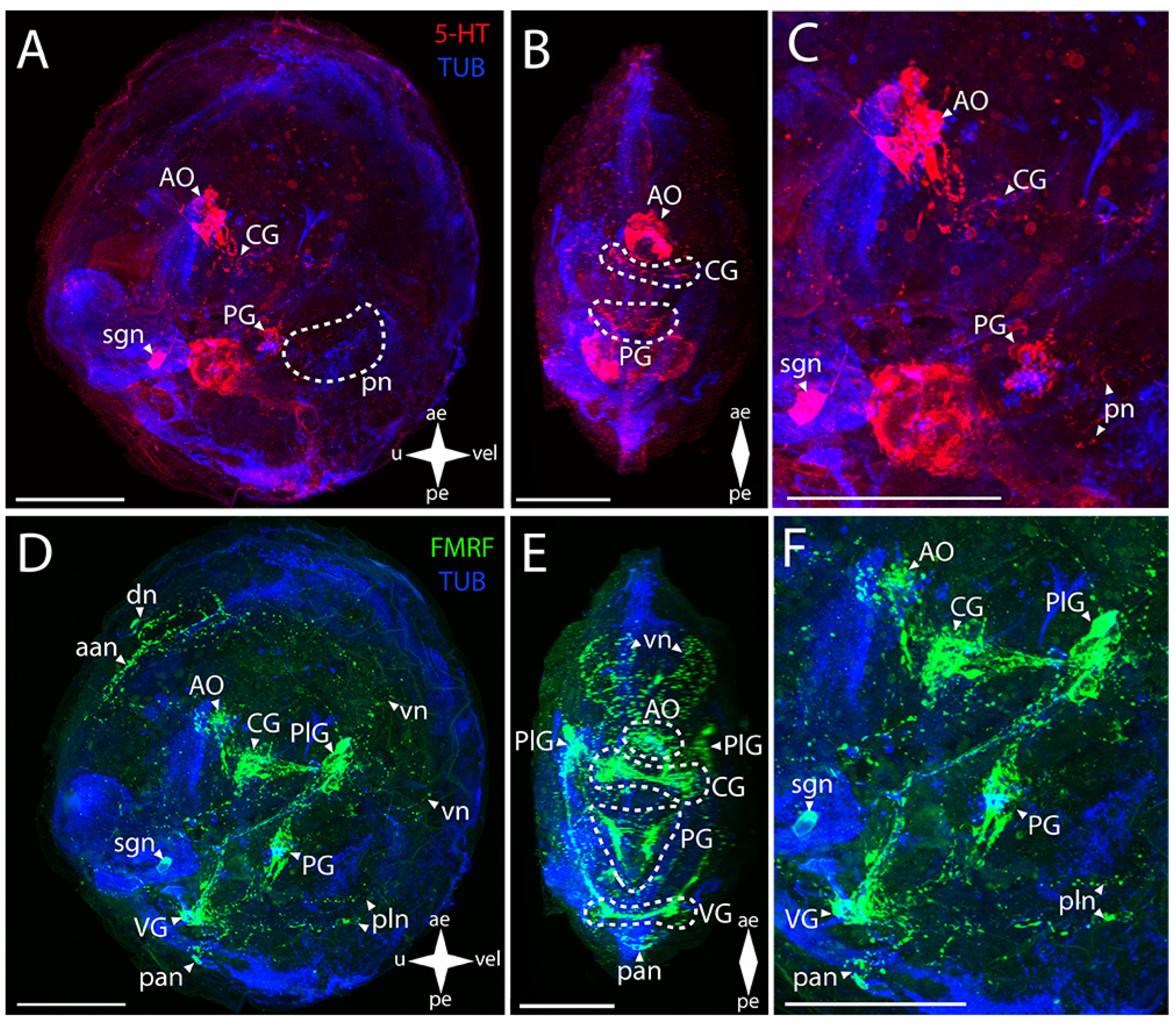
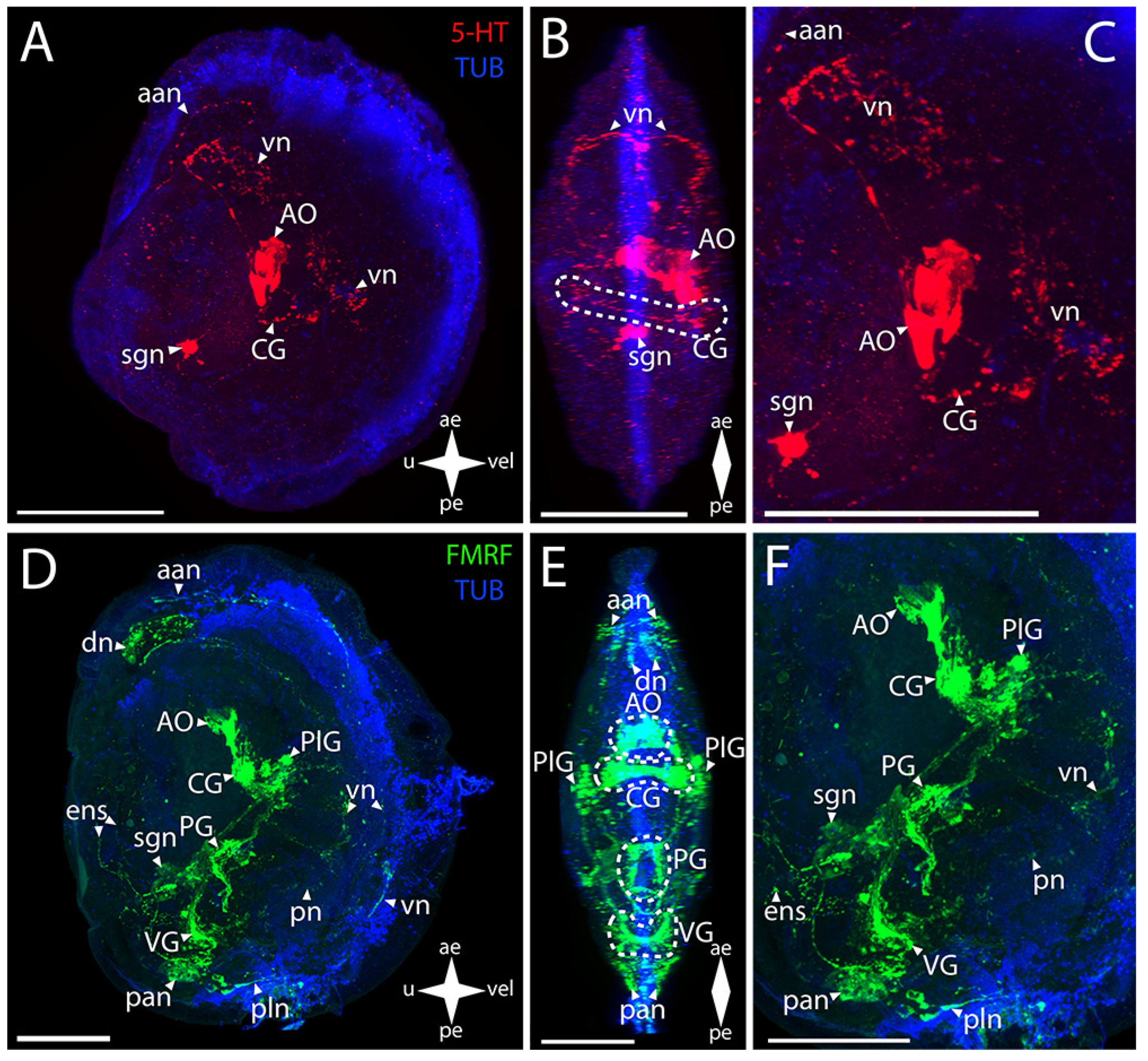
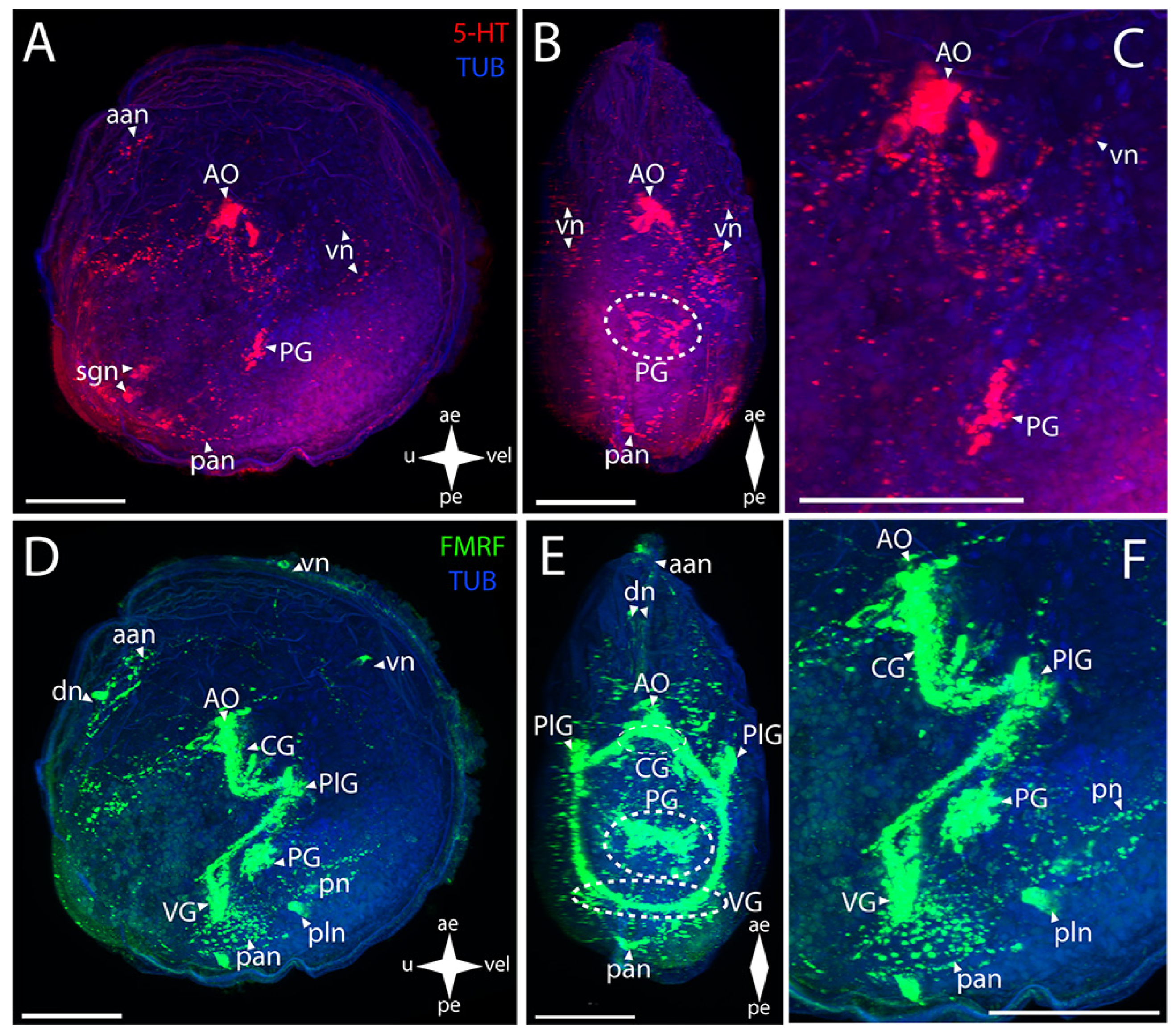
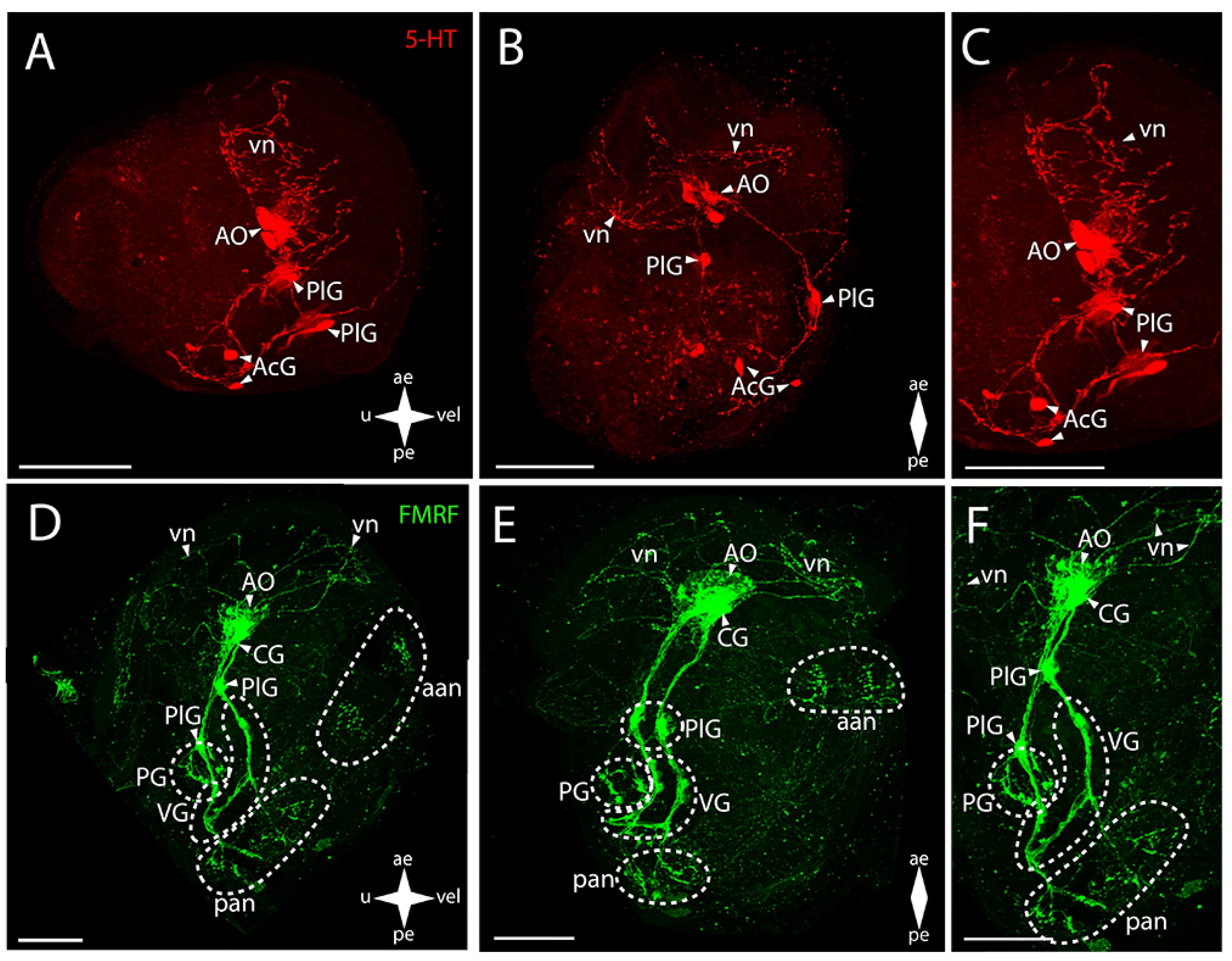
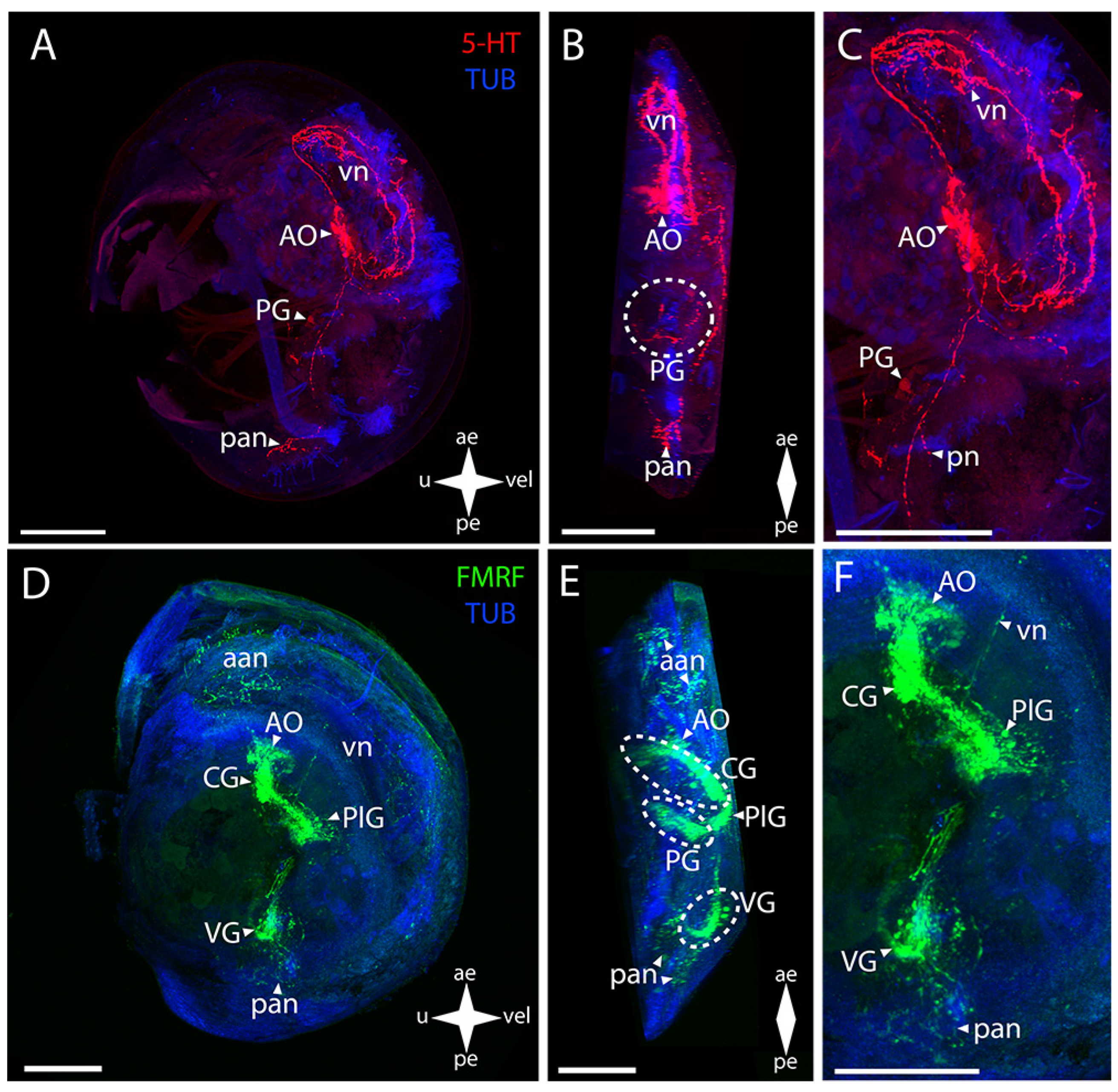
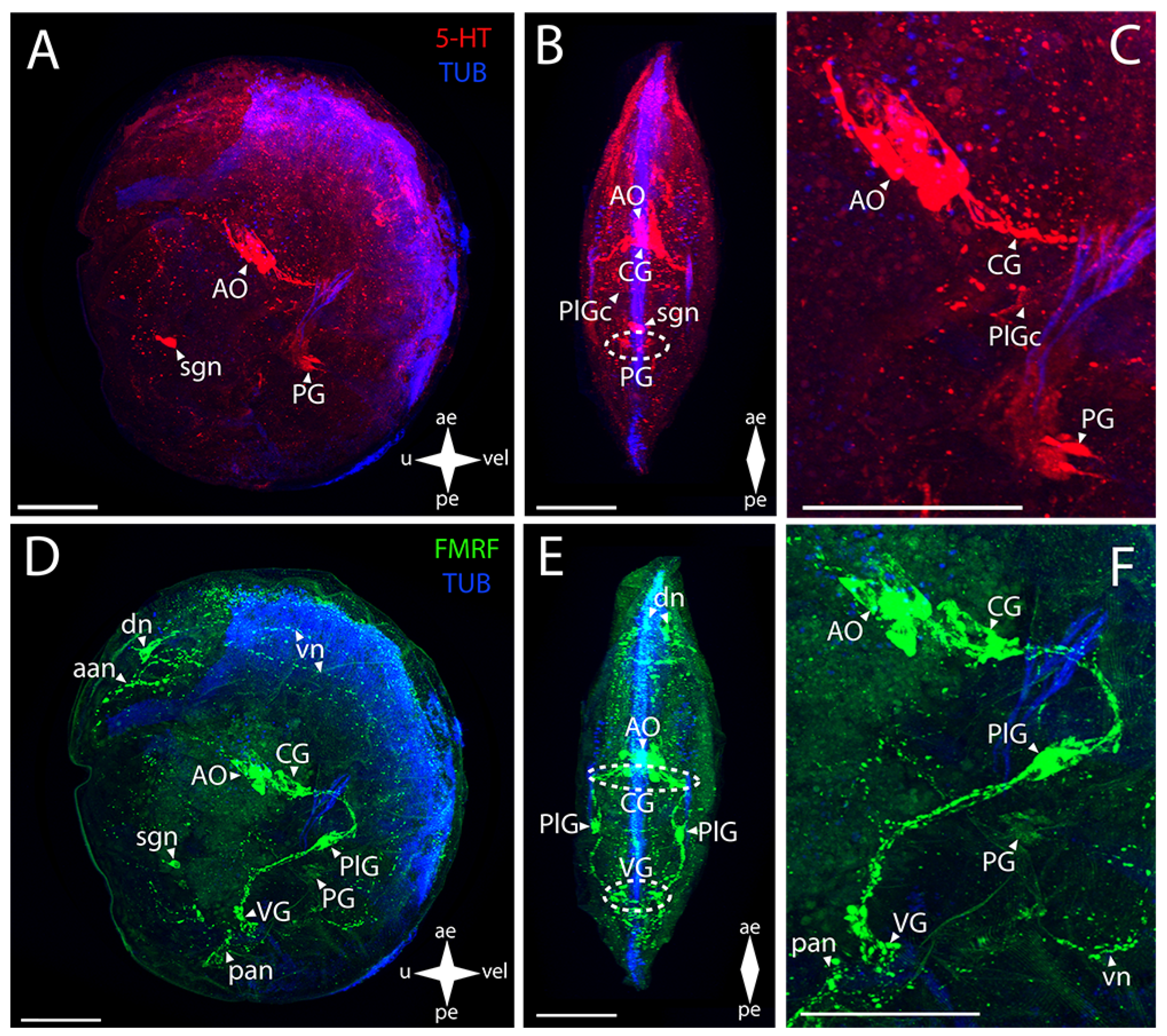
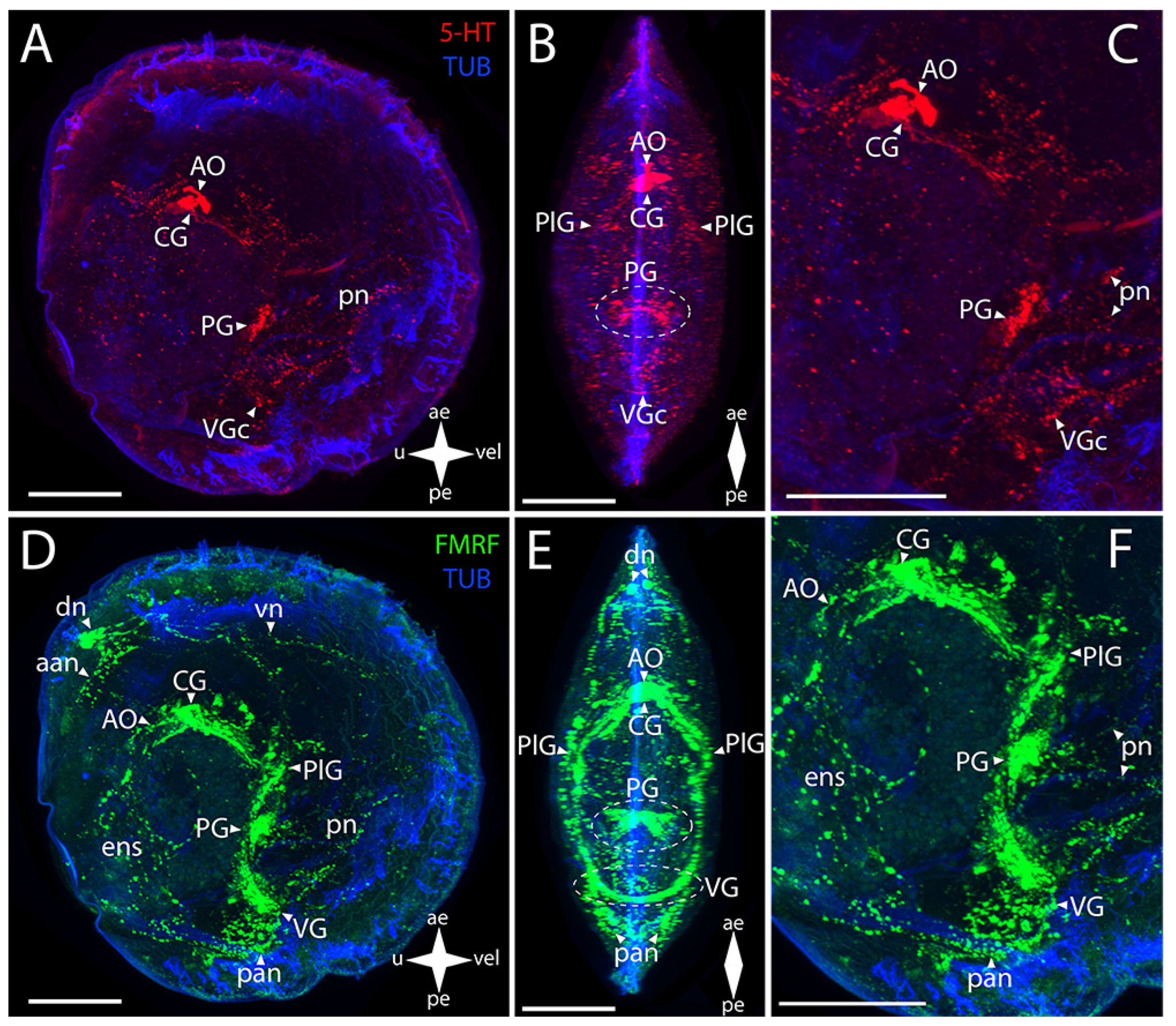
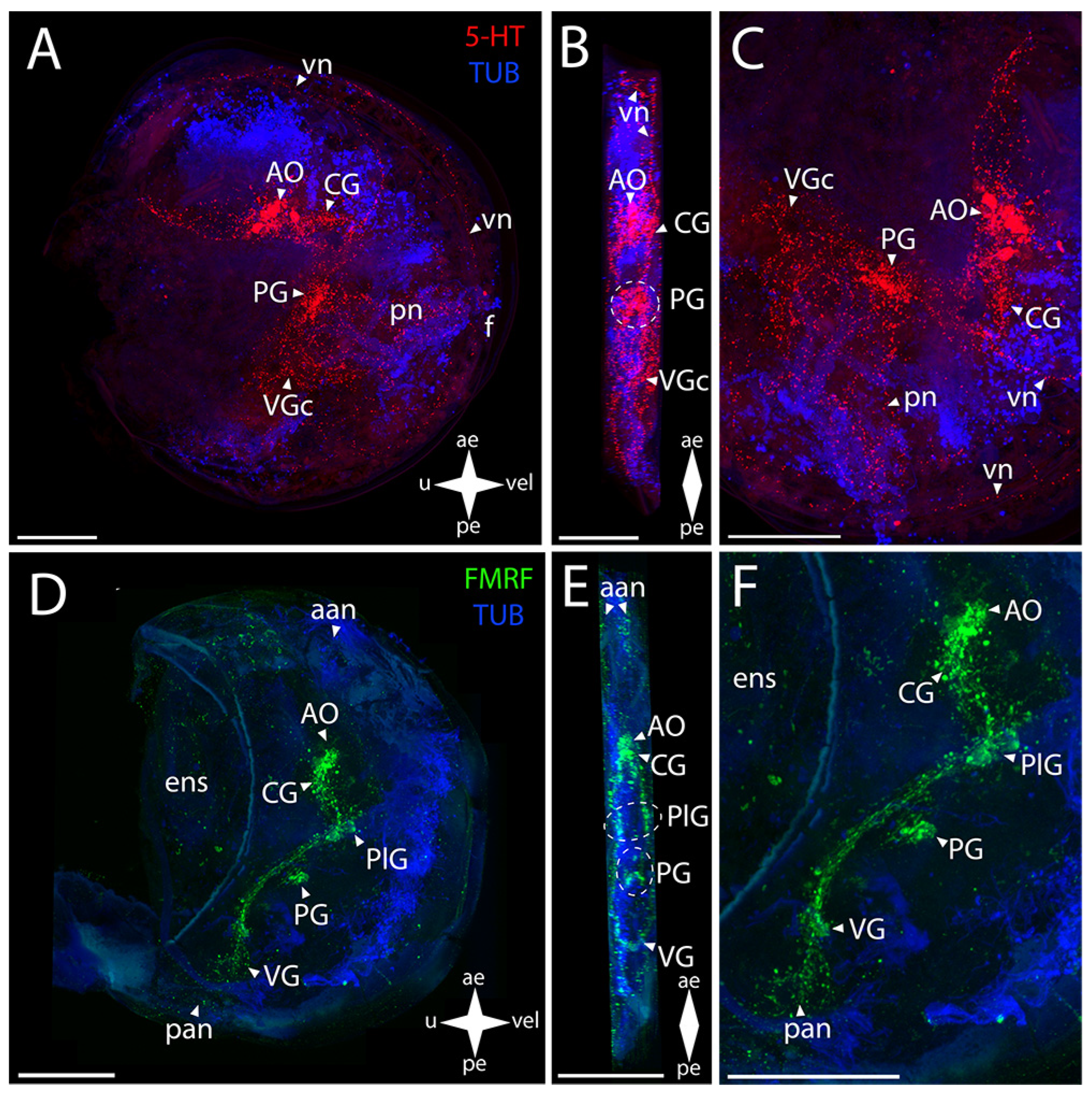
3.2.1. Larval Neuromorphology of Callista brevisiphonata
3.2.2. Larval Neuromorphology of Mactromeris polynyma
3.3. Cemented Forms of Bivalve Mollusks
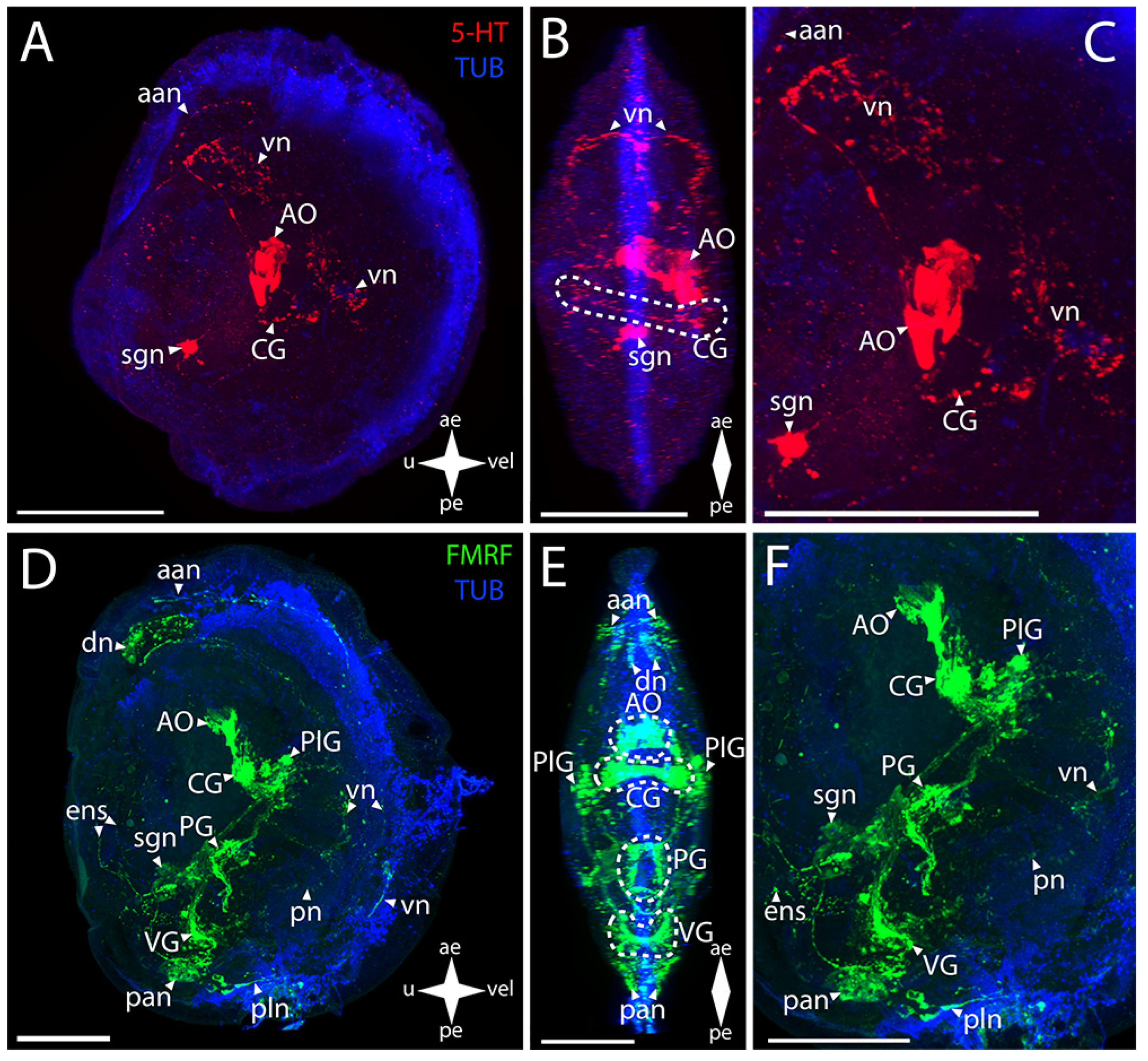
3.3.1. Larval Neuromorphology of Pododesmus macrochisma (Deshayes, 1839), Green Falsejingle
3.3.2. Larval Neuromorphology of Crassostrea gigas (Thunberg, 1793), Pacific Oyster
3.4. Byssally Attached Forms of Bivalve Mollusks
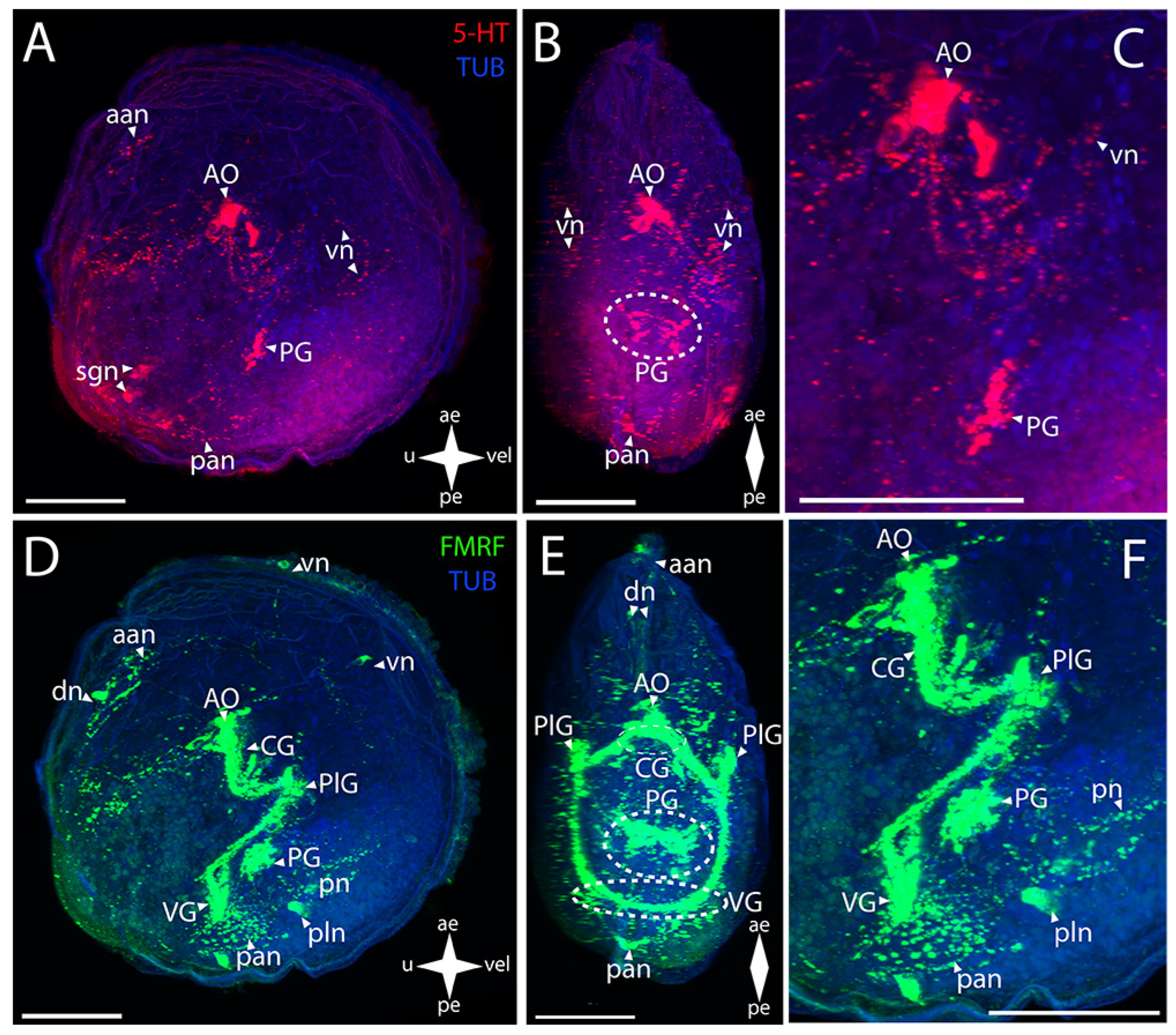
3.4.1. Larval Neuromorphology of Crenomytilus grayanus (Dunker, 1853), Gray’s Mussel
3.4.2. Larval Neuromorphology of Kellia japonica (Pilsbry, 1895), Japanese Kellia
3.5. Mobile Forms of Bivalves
3.5.1. Larval Neuromorphology of Mizuhopecten yessoensis (Jay, 1857), Yesso Scallop
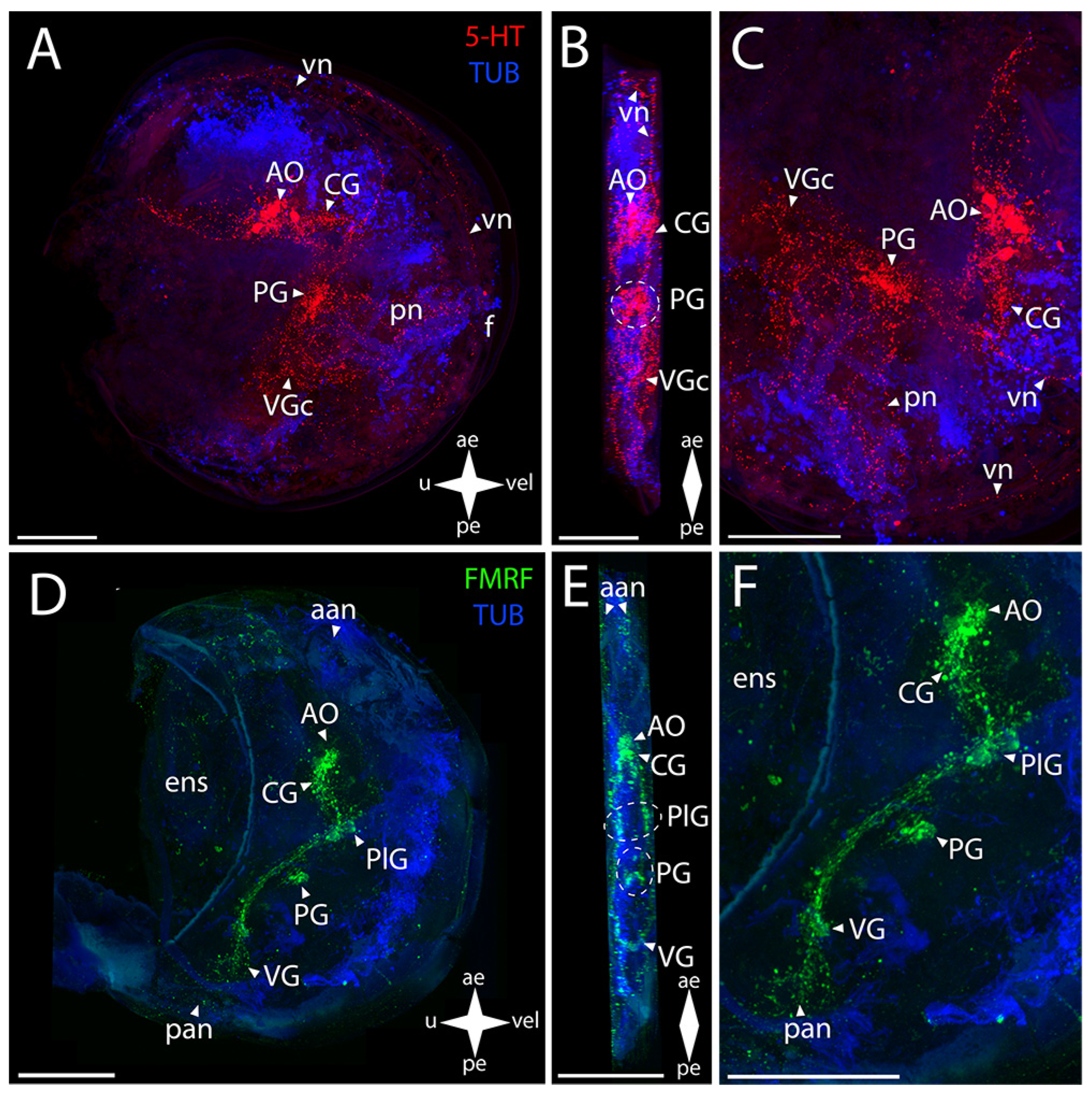
3.5.2. Larval Neuromorphology of Azumapecten farreri (Jones et Preston, 1904), Farrer’s Scallop
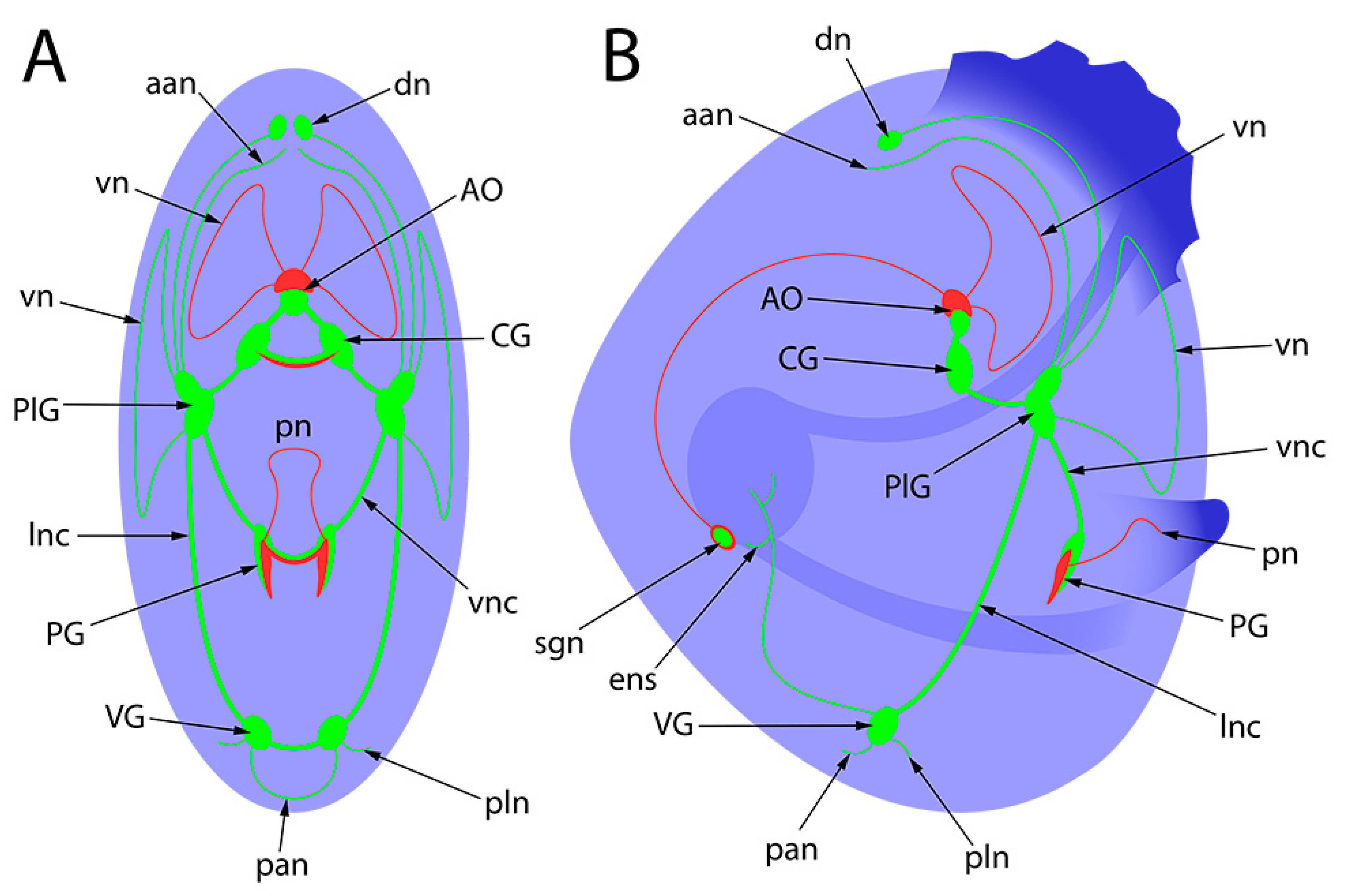
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
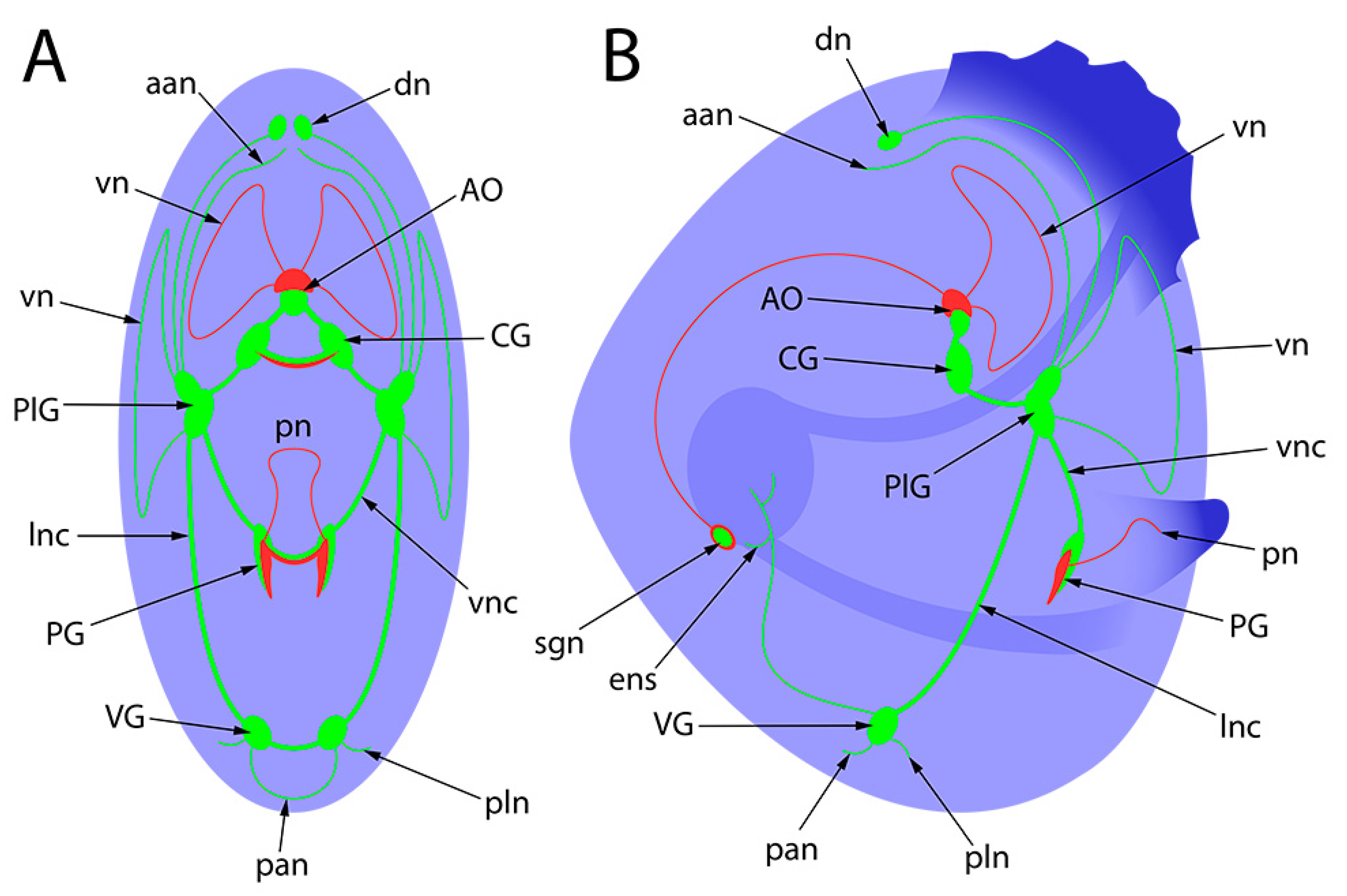
| For bright field figures: | |
| ae | anterior end |
| e | eye |
| f | foot |
| pe | posterior end |
| u | umbro |
| vel | velum |
| For whole-mount confocal figures: | |
| aan | anterior adductor nerves |
| aam | anterior adductor muscle |
| AcG | accessory ganglia |
| AO | apical organ |
| CG | cerebral ganglia |
| dn | dorsal neurons |
| ens | enteric nervous system |
| f | foot |
| pan | posterior adductor nerves |
| PG | pedal ganglia |
| PlG | pleural ganglia |
| pln | pallial nerves |
| pn | pedal nerves |
| sgn | stomatogastric neuron |
| VG | Visceral ganglia |
| VGc | visceral ganglia commissure |
| vn | velum nerves |
References
- Bieler, R.; Mikkelsen, P.M.; Giribet, G. Bivalvia—A discussion of known unknowns. Am. Malacol. Bull. 2013, 31, 123–133. [Google Scholar] [CrossRef]
- Stanley, S.M. Relation of Shell Form to Life Habits of the Bivalvia (Mollusca); The Geological Society of America: Boulder, CO, USA, 1970; ISBN 8137-1125-8. [Google Scholar]
- Slobodskova, V.V.; Leskova, S.E.; Chelomin, V.P. Assessment of the physiological state of cultivated hydrobionts Mizuhopecten yessoensis. N. Sam. Sci. Cent. Rus. Acad. Sci. 2015, 17, 40–44. [Google Scholar]
- Gosling, E. Bivalve Molluscs: Biology, Ecology and Culture; Fishing News Books, Blackwell Publishing Ltd.: Oxford, UK, 2002; ISBN 0-85238-234-0. [Google Scholar]
- Reverol, Y.M.; Delgado, J.G.; de Severeyn, Y.G.; Severeyn, H.J. Embrionary and Larval Development of the Marine Clam Tivela Mactroides (Bivalvia: Veneridae) in Zulia State, Venezuela. Rev. Biol. Trop. 2004, 52, 903–909. [Google Scholar] [PubMed]
- Da Costa, F.; Darriba, S.; Martinez-Patino, D. Embryonic and larval development of Ensis arcuatus (Jeffreys, 1865) (Bivalvia: Pharidae). J. Moll. Stud. 2008, 74, 103–109. [Google Scholar] [CrossRef]
- Voronezhskaya, O.V.; Nezlin, L.P.; Odintsova, N.A.; Plummer, J.T.; Croll, R.P. Neuronal development in larval mussel Mytilus trossulus (Mollusca: Bivalvia). Zoomorphology. 2008, 127, 97–110. [Google Scholar] [CrossRef]
- Dyachuk, V.; Odintsova, N. Development of the larval muscle system in the mussel Mytilus trossulus (Mollusca, Bivalvia). Dev. Grow. Diff. 2009, 51, 69–79. [Google Scholar] [CrossRef]
- Yurchenko, O.V.; Skiteva, O.I.; Voronezhskaya, E.E.; Dyachuk, V.A. Nervous system development in the Pacific oyster, Crassostrea gigas (Mollusca: Bivalvia). Front. Zool. 2018, 15, 10. [Google Scholar] [CrossRef]
- Kniazkina, M.; Dyachuk, V. Neurogenesis of the scallop Azumapecten farreri: From the first larval sensory neurons to the definitive nervous system of juveniles. Front. Zool. 2022, 19, 22. [Google Scholar] [CrossRef]
- Bullock, H.T.; Horridge, G.A. Structure and Function in the Nervous System of Invertebrates; W. H. Freeman and Company Ltd.: San Francisco, CA, USA; London, UK, 1965; Chaprer 28; 1722p. [Google Scholar]
- Kotsyuba, E.; Kalachev, A.; Kameneva, P.; Dyachuk, V. Distribution of molecules related to neurotransmission in the nervous system of the mussel Crenomytilus grayanus. Front. Neuroanat. 2020, 14, 35. [Google Scholar] [CrossRef]
- Wanninger, A. Shaping the things to come: Ontogeny of lophotrochozoan neuromuscular systems and the Tetraneuralia concept. Biol. Bull. 2009, 216, 293–306. [Google Scholar] [CrossRef]
- Yurchenko, O.V.; Savelieva, A.V.; Kolotuchina, N.K.; Voronezhskaya, E.E.; Dyachuk, V.A. Peripheral sensory neurons govern development of the nervous system in bivalve larvae. EvoDevo 2019, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Ivanova-Kazas, O.M. Comparative Embryology of Invertebrates. Trochozoa, Tentaculata, Chaethognatha, Pogonophora; Nauka: Moscow, Russia, 1977; pp. 1–312. [Google Scholar]
- Ellis, I.; Kemph, S.C. Characterization of the central nervous system and various peripheral innervations during larval development of the oyster Crassostrea virginica. Invertebr. Biol. 2011, 130, 236–250. [Google Scholar] [CrossRef]
- Sweeney, D.C. The anatomical distribution of monoamines in a fresh-water bivalve mollusc, Sphaerium sulcatum (L.). Comp. Biochem. Physiol. 1968, 25, 601–613. [Google Scholar] [CrossRef]
- Matsutani, T.; Nomura, T. Serotonin-like immunoreactivity in the central nervous system and gonad of the scallop, Patinopecten yessoensis. Cell Tis. Res. 1986, 244, 515–517. [Google Scholar] [CrossRef]
- Morton, B. The evolution of eyes in the Bivalvia. Ocean. Mar. Biol. Annu. Rev. 2001, 39, 165–205. [Google Scholar]
- Kotsyuba, E.P. Histophysiology of the Central Nervous System of Some Species of Bivalve Mollusks with Different Lifestyles. Ph.D. Thesis, Vladivostok State Medical University, Vladivostok, Russia, 1991. [Google Scholar]
- Kotsyuba, E.; Dyachuk, V. Effect of Air Exposure-Induced Hypoxia on Neurotransmitters and Neurotransmission Enzymes in Ganglia of the Scallop Azumapecten farreri. Int. J. Mol. Sci. 2022, 23, 2027. [Google Scholar] [CrossRef]
- Cragg, S.M.; Nott, J.A. The ultrastructure of the statocysts in the pediveliger larvae of Pecten maximus (L.) (Bivalvia). J. Exp. Mar. Biol. Ecol. 1977, 27, 23–36. [Google Scholar] [CrossRef]
- Kasyanov, V.L.; Kryuchkova, G.A.; Kulikova, V.A.; Medvedeva, L.A. Larvae of Marine Bivalves and Echinoderms; Nauka: Moscow, Russia, 1998; p. 216. [Google Scholar]
- Rakov, V.A. Identification of Bivalve Mollusks of Primorsky Krai; Dalnauka: Vladivostok, Russia, 2006; p. 100. [Google Scholar]
- Evseev, G.A.; Kolotukhina, N.K.; Semenikhina, O.Y. Shell morphogenesis of several venerid bivalves. J. Shellfish Res. 2001, 20, 1279–1284. [Google Scholar]
- Rees, C.B. The identification and classification of lamellibranch larvae. Hull. Bull. Mar. Ecol. 1950, 3, 72–104. [Google Scholar]
- Hayashi, T.; Terai, K. Study on the larvae and young of Japanese surf clam, Spisula (S.) sachalinensis (Schrenk), at Shikuzu, Muroran City. 1. Taxonomy of the Pelecypoda’s veliger larvae in plankton. Sci. Rep. Hokkaido Fish. Exp. Stn. 1964, 2, 7–38. [Google Scholar]
- Kulikova, V.A.; Kolotukhina, N.K. Pelagic larvae of bivalve mollusks of the Sea of Japan. In Methods, Morphology, and Identification; Far Eastern Branch of the Academy of Science of the USSR: Vladivostok, Russia, 1989. [Google Scholar]
- Kolotukhina, N.K.; Kulikova, V.A. The morphology of the larval and juvenile shell of Pododesmus macrochisma (Deshayes, 1839) (Bivalvia: Anomiidae). Rus. J. Mar. Biol. 2016, 42, 102–105. [Google Scholar] [CrossRef]
- Semenikhina, O.Y.; Kolotukhina, N.K.; Evseev, G.A. Morphology of larvae of the family Mytilidae (Bivalvia) from the north-western part of the Sea of Japan. J. Mar. Biol. Assoc. 2008, 88, 331–339. [Google Scholar] [CrossRef]
- Dyachuk, V. Extracellular matrix is required for muscle differentiation in primary cell cultures of larval Mytilus trossulus (Mollusca: Bivalvia). Cytotechnology 2013, 65, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, N.A.; Dyachuk, V.A.; Nezlin, L.P. Muscle and neuronal differentiation in primary cell culture of larval Mytilus trossulus (Mollusca: Bivalvia). Cell Tis. Res. 2010, 339, 625–637. [Google Scholar] [CrossRef]
- Dyachuk, V.A.; Maiorova, M.A.; Odintsova, N.A. Identification of β integrin-like-and fibronectin-like proteins in the bivalve mollusk Mytilus trossulus. Dev. Growth Diff. 2015, 57, 515–528. [Google Scholar] [CrossRef]
- Battonyai, I.; Voronezhskaya, E.E.; Obukhova, A.; Horvath, R.; Nezlin, L.P.; Elekes, K. Neuronal development in the larvae of the invasive biofouler Dreissena polymorpha (Mollusca: Bivalvia), with special attention to sensory elements and swimming behavior. Biol. Bull. 2018, 234, 192–206. [Google Scholar] [CrossRef]
- Pavlicek, A.; Schwaha, T.; Wanninger, A. Towards a ground pattern reconstruction of bivalve nervous systems: Neurogenesis in the zebra mussel Dreissena polymorpha. Org. Divers. Evol. 2018, 18, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Wanninger, A. Evolutionary Developmental Biology of Invertebrates; Springer: Wien, Austria; Heidelberg, Germany; New York, NY, USA, 2015. [Google Scholar]
- Magarlamov, T.Y.; Dyachuk, V.; Chernyshev, A.V. Does the frontal sensory organ in adults of the hoplonemertean Quasitetrastemma stimpsoni originate from the larval apical organ? Front. Zool. 2020, 17, 2. [Google Scholar] [CrossRef]
- Temereva, E.; Rimskaya-Korsakova, N.; Dyachuk, V. Detailed morphology of tentacular apparatus and central nervous system in Owenia borealis (Annelida, Oweniidae). Zool. Let. 2021, 7, 15. [Google Scholar] [CrossRef]
- Richter, S.; Loesel, R.; Purschke, G.; Schmidt-Rhaesa, A.; Scholtz, G.; Stach, T.; Vogt, L.; Wanninger, A.; Brenneis, G.; Döring, C.; et al. Invertebrate neurophylogeny: Suggested terms and definitions for a neuroanatomical glossary. Front. Zool. 2010, 7, 29. [Google Scholar] [CrossRef]
- Kotsyuba, E.P. Localization of NADPH-diaphorase and choline acetyltransferase in the central nervous system of the bivalve mollusk Mactra sulcatoria. Cytology 2007, 49, 48–54. [Google Scholar]
- Welsh, J.H. Excitation of the heart of Venus mercenaria. Arch. Exp. Pathol. Pharmakol. 1953, 219, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Kodirov, S.A. The neuronal control of cardiac functions in Molluscs. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 160, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Twarog, B.M. Responses of a molluscan smooth muscle to acetylcholine and 5-hydroxytryptamine. J. Cell. Comp. Physiol. 1954, 44, 141–163. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.H. Neurohormones of Mollusca. Am. Zool. 1961, 1, 267–272. [Google Scholar] [CrossRef]
- Raineri, M.; Ospovat, M. The initial development of gangliar rudiments in a posterior position in Mytilus galloprovincialis (Mollusca: Bivalvia). J. Mar. Biol. Assoc. U.K. 1994, 74, 73–77. [Google Scholar] [CrossRef]
- Karhunen, T.; Panula, P. Histamine in the nervous system of Macoma balthica (Bivalvia). Agents Actions 1991, 33, 116–118. [Google Scholar] [CrossRef]
- Karhunen, T.; Airaksinen, M.S.; Tuomisto, L.; Panula, P. Neurotransmitters in the nervous system of Macoma balthica (Bivalvia). J. Comp. Neurol. 1993, 334, 477–488. [Google Scholar] [CrossRef]
- Bonar, D.B.; Coon, S.L.; Walch, M.; Weiner, R.M.; Fitt, W. Control of oyster settlement and metamorphosis by endogenous and exogenous chemical cues. Bull. Mar. Sci. 1990, 46, 484–498. [Google Scholar]
- Beiras, R.; Widdows, J. Effect of the neurotransmitters dopamine, serotonin and norepinephrine on the ciliary activity of mussel (Mytilus edulis) larvae. Mar. Biol. 1995, 122, 597–603. [Google Scholar] [CrossRef]
- Coon, S.L.; Bonar, D.B. Pharmacological evidence that α-adrenoceptors mediate metamorphosis of the pacific oyster, Crassostrea gigas. Neuroscience 1987, 23, 1169–1174. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Liu, B.; Tang, B.; Zhang, T.; Xiang, J. Pharmacological and immunocytochemical investigation of the role of catecholamines on larval metamorphosis by β-adrenergic-like receptor in the bivalve Meretrix meretrix. Aquaculture 2006, 258, 611–618. [Google Scholar] [CrossRef]
- Croll, R.P.; Jackson, D.L.; Voronezhskaya, E.E. Catecholamine-containing cells in larval and postlarval bivalve molluscs. Biol. Bull. 1997, 193, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Flyachinskaya, L.P.; Raikova, O.I. A study of the nervous system of the pediveliger bivalve mollusk Mya arenaria. Zool. J. 2014, 93, 489. [Google Scholar]
- Kreiling, J.A.; Jessen-Eller, K.; Miller, J.; Seegal, R.F.; Reinisch, C.L. Early development of the serotonergic and dopaminergic nervous system in Spisula solidissima (surf clam) larvae. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 130, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Kotsyuba, E.P. Effect of elevated temperature and of hypoxia on NO activity in the central nervous system of bivalve molluscs. J. Evol. Biochem. Physiol. 2008, 44, 237–246. [Google Scholar] [CrossRef]
- Malyshev, I.Y.; Manukhina, E.B. Stress, Adaptation and Nitric Oxide. Biokhimiya 1998, 63, 992–1006. [Google Scholar]
- Seilacher, A. Constructional morphology of bivalves: Evolutionary pathways in primary versus secondary soft-bottom dwellers. Palaeontology 1984, 27, 207–237. [Google Scholar]
- González, V.L.; Andrade, S.C.; Bieler, R.; Collins, T.M.; Dunn, C.W.; Mikkelsen, P.M.; Taylor, J.D.; Giribet, G. A phylogenetic backbone for Bivalvia: An RNA-seq approach. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142332. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Immunized Animal | Titer | Manufacturer, Country | No. of Antibody |
|---|---|---|---|---|
| Primary antibodies | ||||
| Antibodies against serotonin (5-hydroxytryptamine, 5-HT) | Goat | 1:500–1:1000 | Immunostar, Hudson, WI, USA | 20079 |
| Antibodies against FMRFamide (cardio-excitatory peptide) | Rabbit | 1:500–1:1000 | Immunostar, Hudson, WI, USA | 20091 |
| Antibodies against acetylated (α-tubulin) | Mouse | 1:500–1:1000 | Santa Cruz Biotechnology, Santa Cruz, CA, USA | sc-23950 |
| Secondary antibodies | ||||
| Antibodies against goat blood (Alexa Fluor 555 donkey anti-goat IgG (H + L)) | Donkey | 1:500–1:1000 | Invitrogen, Eugene, OR, USA | A21432 |
| Antibodies against rabbit blood serum (Alexa Fluor 488 donkey anti-rabbit IgG (H + L)) | Donkey | 1:500–1:1000 | Invitrogen, Eugene, OR, USA | A21206 |
| Antibodies against mouse blood serum (Alexa Fluor Plus 647 donkey anti-mouse IgG) | Donkey | 1:500–1:1000 | Invitrogen, Eugene, OR, USA | A31571 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikishchenko, V.; Kolotukhina, N.; Dyachuk, V. Comparative Neuroanatomy of Pediveliger Larvae of Various Bivalves from the Sea of Japan. Biology 2023, 12, 1341. https://doi.org/10.3390/biology12101341
Nikishchenko V, Kolotukhina N, Dyachuk V. Comparative Neuroanatomy of Pediveliger Larvae of Various Bivalves from the Sea of Japan. Biology. 2023; 12(10):1341. https://doi.org/10.3390/biology12101341
Chicago/Turabian StyleNikishchenko, Viktoriya, Nataliya Kolotukhina, and Vyacheslav Dyachuk. 2023. "Comparative Neuroanatomy of Pediveliger Larvae of Various Bivalves from the Sea of Japan" Biology 12, no. 10: 1341. https://doi.org/10.3390/biology12101341
APA StyleNikishchenko, V., Kolotukhina, N., & Dyachuk, V. (2023). Comparative Neuroanatomy of Pediveliger Larvae of Various Bivalves from the Sea of Japan. Biology, 12(10), 1341. https://doi.org/10.3390/biology12101341