

Elevated CO2 and Water Stress in Combination in Plants: Brothers in Arms or Partners in Crime?

 , and
, and
Abstract
:Simple Summary
Abstract
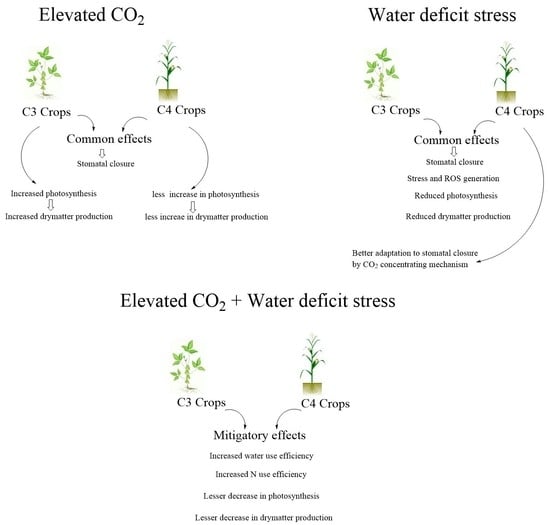
1. Introduction
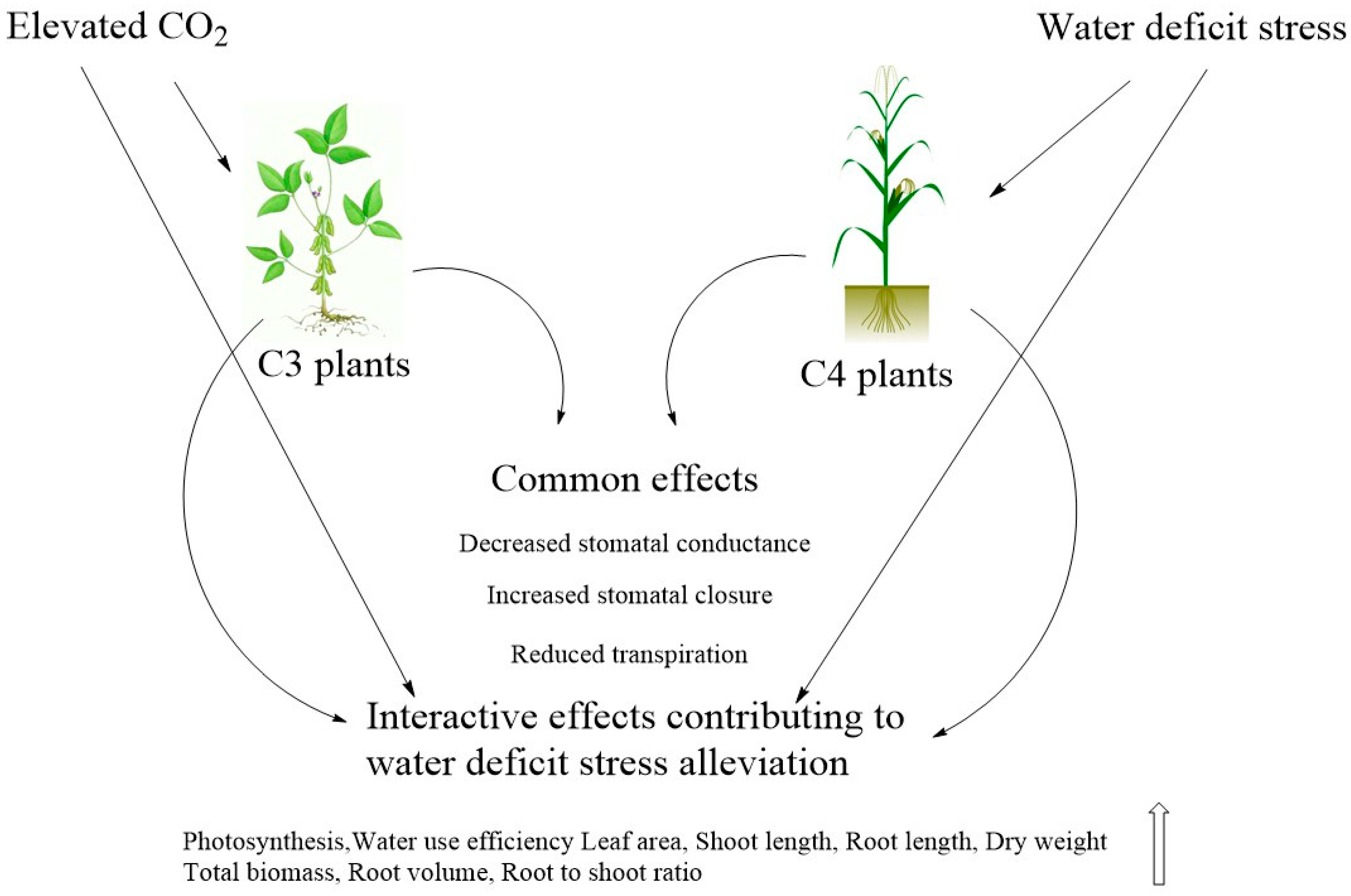
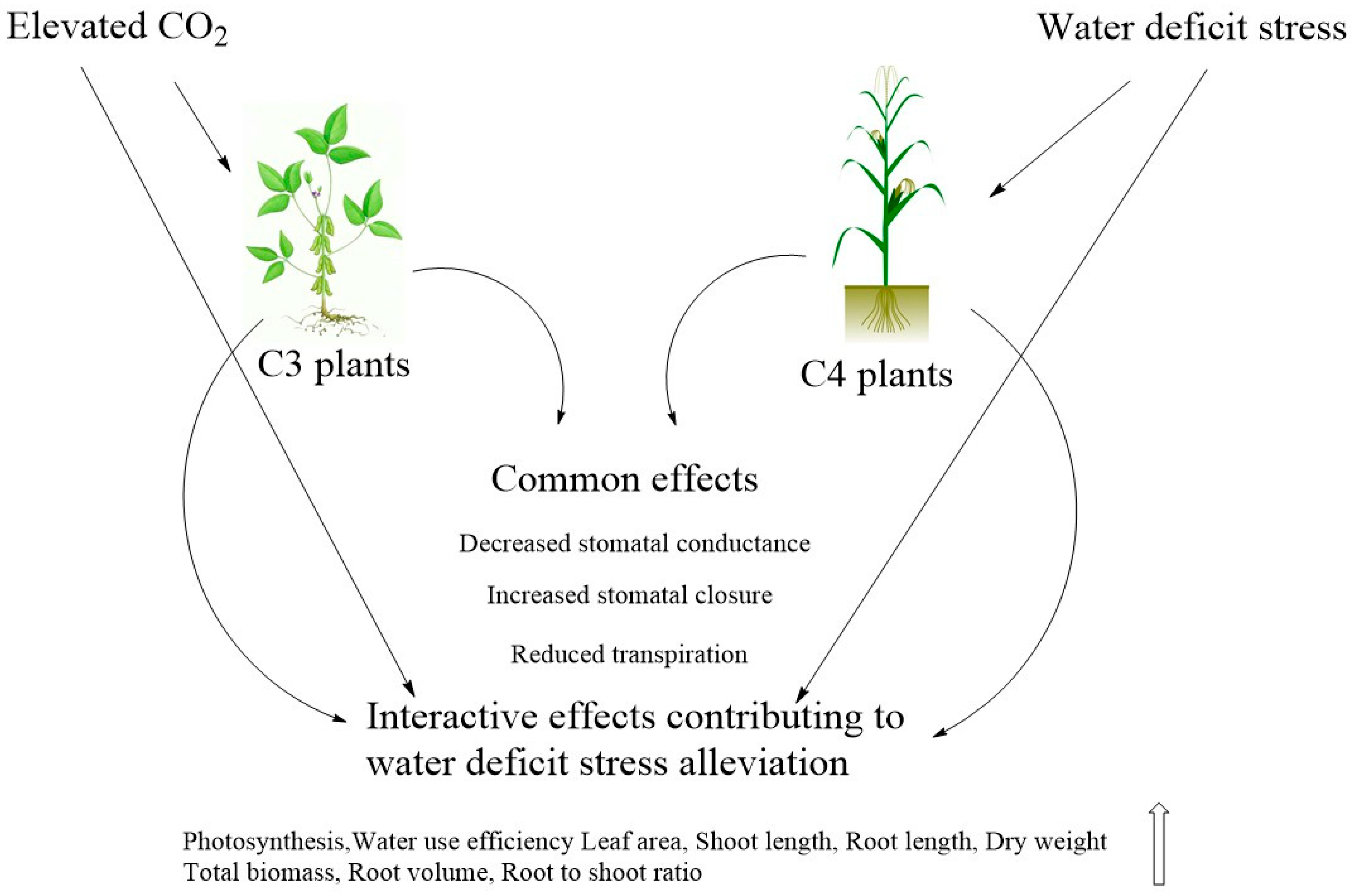
2. Water Relations, Transpiration and Stomatal Conductance
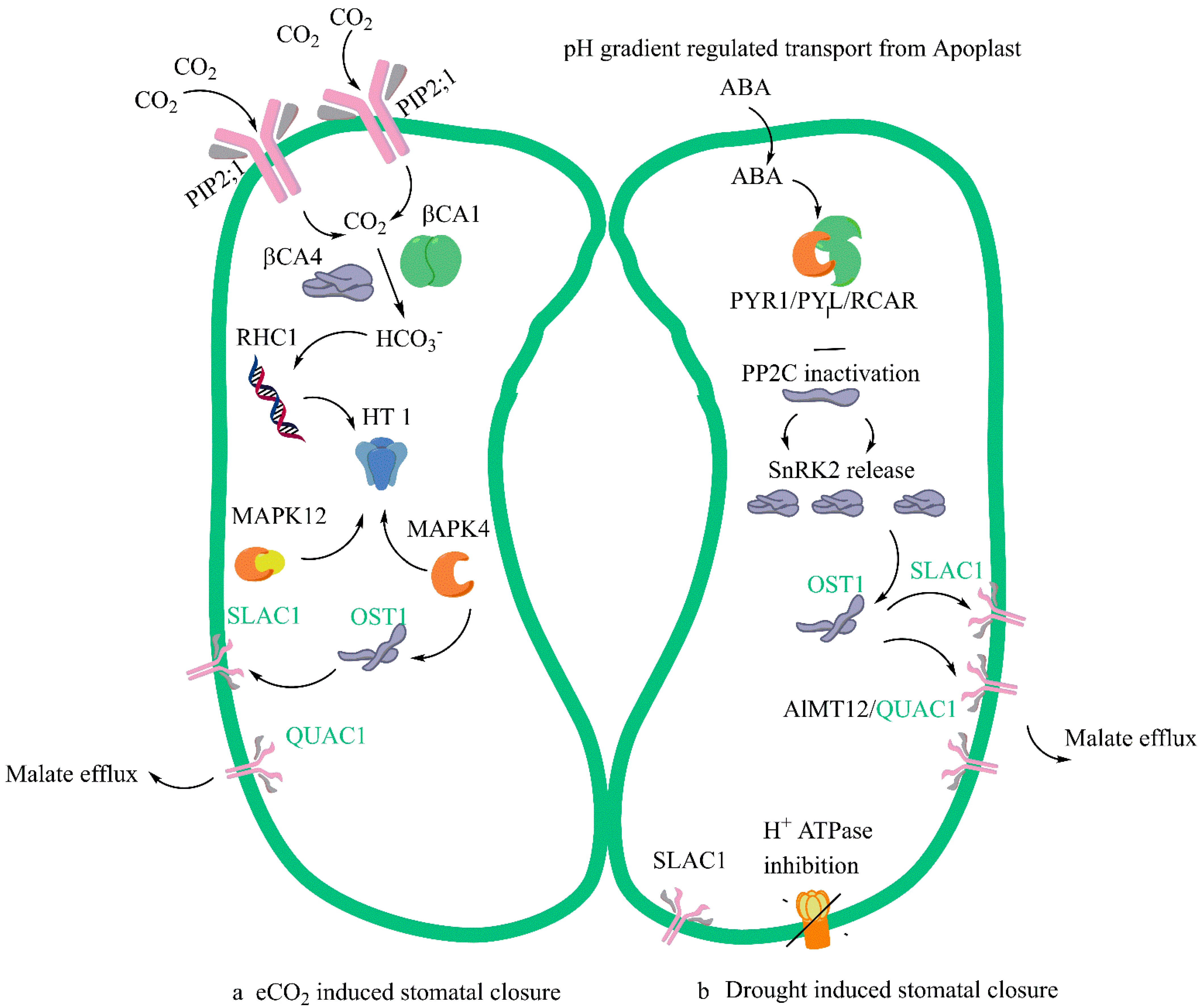
2.1. Stomatal Dynamics
2.2. The ABA Conundrum
2.3. Water Relations
3. Dry Matter Production
3.1. Photosynthesis and Growth
3.2. Malate Maelstrom
3.3. Nutrients in a Nutshell

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant/Crop/Tree | CO2 Concentration | Water Stress Imposition | Effect | Reference |
|---|---|---|---|---|
| Poplar (Populus spp.) | 700 ± 50 µmol mol−1 | Soil drying cycle by withholding water | Reduced Gas exchange, decreased leaf conductance, increased photosynthesis, increased transpiration efficiency | [12] |
| Wheat (Triticum aestivum) | 400 μmol mol−1, 790 μmol mol−1 | Progressive restriction of water from 10 percent to 60 percent pot capacity | Reduced plant biomass, Stomatal conductance and carbon isotope signature indicated water saving strategy. PEPC expression increased | [72] |
| Tabernaemontana divaricata | 1000 μmol mol−1, 700 μmol mol−1 | 70 percent of field capacity (FC) for 4 days and 30 percent of FC for 4 days | Increase in stomatal conductance (gs), plant height (PH) and plant girth (PG) | [82] |
| Maize (Zea mays) | 550 μmol mol−1 | Half water in water stress treatment compared to control | 37 percent reduction in whole plant transpiration | [43] |
| Napier grass (Pennisetum purpureum Schumach × Pennisetum glaucum (L.) R. Br) and hydric common reed grass (Phragmites australis (Cav.) Trin. Ex Steud) | 563 ± 6.7 μmol mol−1 541 ± 6.9 μmol mol−1 601 ± 9.1 μmol mol−1 | Withdrawing irrigation | Increase in Photosynthesis, reduced leaf water potential and increase in transpiration | [44] |
| Maize (Zea mays) | 700 μmol mol−1, 900 μmol mol−1, and 1200 μmol mol−1 | deficit irrigation | Decreases in stomatal conductance and reduced transpiration rate | [45] |
| Soyabean (Glycine max) | Ambient + 200 μmol mol−1 | 35–45 percent of RWC | Elevated CO2 enhanced the resistance to drought by improving the capacity of photosynthesis and WUE in soybean leaves | [46] |
| Pinus halepensis (Aleppo pine) | 867 ± 157 μmol mol−1 | 10 Percent Relative Substrate Water Content | Under drought, the effect of CO2 on WUE was pronounced, with intercellular CO2 being increased near stomatal closure | [83] |
| Lemon (Citrus limon) | 650 and 850 μmol mol−1 | leaf water potential of −3.5 MPa | Stomatal downregulation at elevated CO2 reduced water-use but not photosynthesis. | [84] |
| Soybean (Glycine max) | 800 μmol mol−1 | water deficit was applied by randomly moving plants out of the hydroponic solution exposing the roots to ambient- or elevated-air | Responses of soybean roots to short-term water deficit are buffered by Elevated CO2 | [47] |
| Cassava (Manihot esculenta Crantz) | 750 μmol mol−1 | Stopping irrigation for 7 days | Elevated CO2 reduced the negative effect of drought on transpiration, water use efficiency, all growth measures and harvest index. | [85] |
| Faba bean (Vicia faba L.) | 550 μmol mol−1 | Water was withheld until 30 percent FC | Elevated CO2-induced stimulation of nodulation and nodule density helped maintain N2 fixation under drought | [86] |
| Andiroba (Carapa surinamensis) | 700 μmol mol−1 | 50 percent field capacity | Whole-plant water-use efficiency (WUE) improved under combination treatments | [87] |
| Hymenaea stigonocarpa Mart. ex Hayne, Solanum lycocarpum A. St.-Hil. and Tabebuia aurea (Silva Manso) Benth. and Hook. f. ex S. Moore | 700 μmol mol−1 | Water stress was introduced three times during the experiment by halting irrigation 1 month before the fourth (360 days old), fifth (450 days old) and sixth (540 days old) morphophysiological surveys | Water stress decreased biomass production under high CO2 | [88] |
| Grapevines (Vitis labrusca) | 800 μmol mol−1 | Stopping irrigation | Elevated CO2 delayed drought effects on both net photosynthetic rate and Rubisco activity for four days, by reducing stomatal conductance, transpiration, and stomatal density | [89] |
| Brassica napus | 800 μmol mol−1 | Withholding water for 7 days | Elevated CO2 diminished the adverse effect by improved water relations | [52] |
| Maize (Zea mays) | 550, 700, and 900 μmol mol−1 | Deficit irrigation | Photosynthetic rate in elevated CO2 concentrations was higher under Deficit irrigation than under regular irrigation. | [90] |
| Acrocomia aculeata | 700 μmol mol−1 | Water withholding | Higher Rubisco carboxylation rate (Vc max) and electron transport rate (J max) contributed to recovery from drought | [56] |
| Cucumber (Cucumis sativus L.) | 800 ± 20 μmol mol−1 | ψw = −0.05 MPa and ψw = −0.15 with PEG 6000 | Higher photosynthetic performance and increased grana thickness under moderate drought stress, increased palisade cells length and chloroplasts number per palisade cell under severe drought stress. | [91] |
4. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alexandratos, N.; Bruinsma, J. World Agriculture Towards 2030/2050: The 2012 Revision; FAO: Rome, Italy, 2012. [Google Scholar]
- United Nations. World Population Prospects: The 2012 Revision, Key Findings and Advance Tables; UN: Geneva, Switzerland, 2012. [Google Scholar]
- IPCC (Intergovernmental Panel on Climate Change). Global Warming of 1.5 °C. In An IPCC Special Report on the Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; IPCC: Geneva, Switzerland, 2018. [Google Scholar]
- Friedlingstein, P.; Jones, M.; O’sullivan, M.; Andrew, R.; Hauck, J.; Peters, G.; Peters, W.; Pongratz, J.; Sitch, S.; Le Quéré, C.; et al. Global carbon budget 2019. Earth Syst. Sci. Data 2019, 11, 1783–1838. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2007: Impacts, Adaptation and Vulnerability; IPCC Secretariat: Geneva, Switzerland, 2007. [Google Scholar]
- Shanker, A.K.; Maheswari, M.; Yadav, S.K.; Desai, S.; Bhanu, D.; Attal, N.B.; Venkateswarlu, B. Drought stress responses in crops. Funct. Integr. Genom. 2014, 14, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Kimball, B.A.; LaMorte, R.L.; Seay, R.S.; Pinter, P.J., Jr.; Rokey, R.R.; Hunsaker, D.J.; Dugas, W.A.; Heuer, M.L.; Mauney, J.R.; Hendrey, G.R.; et al. Effects of free-air CO2 enrichment on energy balance and evapotranspiration of cotton. Agric. For. Meteorol. 1994, 70, 259–278. [Google Scholar] [CrossRef]
- Pinter, P.J., Jr.; Kimball, B.A.; Garcia, R.L.; Wall, G.W.; Hunsaker, D.J.; LaMorte, R.L. Free-air CO2 enrichment: Responses of cotton and wheat crops. In Carbon Dioxide and Terrestrial Ecosystems; Koch, G.W., Mooney, H.A., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 215–264. [Google Scholar]
- De Luis, I.; Irigoyen, J.J.; Sánchez-Díaz, M. Elevated CO2 enhances plant growth in droughted N2-fixing alfalfa without improving water status. Physiol. Plant. 1999, 107, 84–89. [Google Scholar] [CrossRef]
- Clark, H.; Newton, P.C.D.; Barker, D.J. Physiological and morphological responses to elevated CO2 and a soil moisture deficit of temperate pasture species growing in an established plant community. J. Exp. Bot. 1999, 50, 233–242. [Google Scholar] [CrossRef]
- Johnson, J.D.; Tognetti, R.; Paris, P. Water relations and gas exchange in poplar and willow under water stress and elevated atmospheric CO2. Physiol. Plant. 2002, 115, 93–100. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Jia, B.; Zhou, G. Elevated-CO2 response of stomata and its dependence on environmental factors. Front. Plant Sci. 2016, 7, 657. [Google Scholar] [CrossRef]
- Zhang, X.; Pu, P.; Tang, Y.; Zhang, L.; Lv, J. C4 photosynthetic enzymes play a key role in wheat spike bracts primary carbon metabolism response under water deficit. Plant Physiol. Biochem. 2019, 142, 163–172. [Google Scholar] [CrossRef]
- Habermann, E.; Dias de Oliveira, E.A.; Contin, D.R.; San Martin, J.A.; Curtarelli, L.; Gonzalez-Meler, M.A.; Martinez, C.A. Stomatal development and conductance of a tropical forage legume are regulated by elevated [CO2] under moderate warming. Front. Plant Sci. 2019, 10, 609. [Google Scholar] [CrossRef]
- Caldera, H.I.U.; De Costa, W.J.M.; Woodward, F.I.; Lake, J.A.; Ranwala, S.M. Effects of elevated carbon dioxide on stomatal characteristics and carbon isotope ratio of Arabidopsis thaliana ecotypes originating from an altitudinal gradient. Physiol. Plant. 2017, 159, 74–92. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Abdelhakim, L.O.A.; Fang, L.; Peng, X.; Liu, J.; Liu, F. Elevated CO2 effect on the response of stomatal control and water use efficiency in amaranth and maize plants to progressive drought stress. Agric. Water Manag. 2022, 266, 107609. [Google Scholar] [CrossRef]
- Marek, S.; Tomaszewski, D.; Żytkowiak, R.; Jasińska, A.; Zadworny, M.; Boratyńska, K.; Dering, M.; Danusevičius, D.; Oleksyn, J.; Wyka, T.P. Stomatal density in Pinus sylvestris as an indicator of temperature rather than CO2: Evidence from a pan-European transect. Plant Cell Environ. 2022, 45, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Israel, W.K.; Watson-Lazowski, A.; Chen, Z.H.; Ghannoum, O. High intrinsic water use efficiency is underpinned by high stomatal aperture and guard cell potassium flux in C3 and C4 grasses grown at glacial CO2 and low light. J. Exp. Bot. 2022, 73, 1546–1565. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Ding, R.; Yang, X.; Du, T. How are leaf carbon-and water-related traits coordinated acclimation to elevated CO2 by its anatomy: A case study in tomato. Environ. Exp. Bot. 2022, 199, 104898. [Google Scholar] [CrossRef]
- Lim, C.W.; Baek, W.; Jung, J.; Kim, J.H.; Lee, S.C. Function of ABA in stomatal defense against biotic and drought stresses. Int. J. Mol. Sci. 2015, 16, 15251–15270. [Google Scholar] [CrossRef]
- Bharath, P.; Gahir, S.; Raghavendra, A.S. Abscisic acid-induced stomatal closure: An important component of plant defense against abiotic and biotic stress. Front. Plant Sci. 2021, 12, 324. [Google Scholar] [CrossRef]
- Li, S.; Fang, L.; Hegelund, J.N.; Liu, F. Elevated CO2 modulates plant hydraulic conductance through regulation of PIPs under progressive soil drying in tomato plants. Front. Plant Sci. 2021, 12, 916. [Google Scholar] [CrossRef]
- Ma, X.; Bai, L. Elevated CO2 and reactive oxygen species in stomatal closure. Plants 2021, 10, 410. [Google Scholar] [CrossRef]
- Murata, Y.; Mori, I.C.; Munemasa, S. Diverse stomatal signaling and the signal integration mechanism. Annu. Rev. Plant Biol. 2015, 66, 369–392. [Google Scholar] [CrossRef]
- Assmann, S.M.; Jegla, T. Guard cell sensory systems: Recent insights on stomatal responses to light, abscisic acid, and CO2. Curr. Opin. Plant Biol. 2016, 33, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Negi, J.; Matsuda, O.; Nagasawa, T.; Oba, Y.; Takahashi, H.; Kawai-Yamada, M.; Uchimiya, H.; Hashimoto, M.; Iba, K. CO2 regulator SLAC1 and its homologues are essential for anion homeostasis in plant cells. Nature 2008, 452, 483–486. [Google Scholar] [CrossRef] [PubMed]
- Monda, K.; Mabuchi, A.; Negi, J.; Iba, K. Cuticle permeability is an important parameter for the trade-off strategy between drought tolerance and CO2 uptake in land plants. Plant Signal. Behav. 2021, 16, 1908692. [Google Scholar] [CrossRef] [PubMed]
- Dubeaux, G.; Hsu, P.K.; Ceciliato, P.H.; Swink, K.J.; Rappel, W.J.; Schroeder, J.I. Deep dive into CO2-dependent molecular mechanisms driving stomatal responses in plants. Plant Physiol. 2021, 187, 2032–2042. [Google Scholar] [CrossRef]
- Jalakas, P.; Nuhkat, M.; Vahisalu, T.; Merilo, E.; Brosché, M.; Kollist, H. Combined action of guard cell plasma membrane rapid-and slow-type anion channels in stomatal regulation. Plant Physiol. 2021, 187, 21262133. [Google Scholar] [CrossRef]
- Zhang, J.; De-oliveira-Ceciliato, P.; Takahashi, Y.; Schulze, S.; Dubeaux, G.; Hauser, F.; Azoulay-Shemer, T.; Tõldsepp, K.; Kollist, H.; Rappel, W.J.; et al. Insights into the molecular mechanisms of CO2-mediated regulation of stomatal movements. Curr. Biol. 2018, 28, 1356–1363. [Google Scholar] [CrossRef]
- Driesen, E.; Van den Ende, W.; De Proft, M.; Saeys, W. Influence of environmental factors light, CO2, temperature, and relative humidity on stomatal opening and development: A review. Agronomy 2020, 10, 1975. [Google Scholar] [CrossRef]
- Ren, Z.; Suolang, B.; Fujiwara, T.; Yang, D.; Saijo, Y.; Kinoshita, T.; Wang, Y. Promotion and upregulation of a plasma membrane proton-atpase strategy: Principles and applications. Front. Plant Sci. 2021, 12, 749337. [Google Scholar] [CrossRef]
- Nobori, T. Closing the gap: A plasma membrane H+-ATPase regulates stomatal closure. Plant Cell. 2022, 34, 2582–2583. [Google Scholar] [CrossRef]
- Schulze, S.; Dubeaux, G.; Ceciliato, P.H.; Munemasa, S.; Nuhkat, M.; Yarmolinsky, D.; Aguilar, J.; Diaz, R.; Azoulay-Shemer, T.; Steinhorst, L.; et al. A role for calcium-dependent protein kinases in differential CO2-and ABA-controlled stomatal closing and low CO2-induced stomatal opening in Arabidopsis. New Phytol. 2021, 229, 2765–2779. [Google Scholar] [CrossRef]
- Jing, X.; Cai, C.; Fan, S.; Wang, L.; Zeng, X. Spatial and temporal calcium signaling and its physiological effects in Moso Bamboo under drought stress. Forests 2019, 10, 224. [Google Scholar] [CrossRef]
- Kudla, J.; Becker, D.; Grill, E.; Hedrich, R.; Hippler, M.; Kummer, U.; Parniske, M.; Romeis, T.; Schumacher, K. Advances and current challenges in calcium signaling. New Phytol. 2018, 218, 414–431. [Google Scholar] [CrossRef]
- Richards, M.; Xu, W.; Mallozzi, A.; Errera, R.M.; Supan, J. Production of calcium-binding proteins in Crassostrea virginica in response to increased environmental CO2 concentration. Front. Mar. Sci. 2018, 5, 203. [Google Scholar] [CrossRef]
- Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Merilo, E.; Laanemets, K.; Waadt, R.; Pater, D.; Kollist, H.; Schroeder, J.I. Abscisic acid-independent stomatal CO2 signal transduction pathway and convergence of CO2 and ABA signaling downstream of OST1 kinase. Proc. Natl. Acad. Sci. USA 2018, 115, 9971–9980. [Google Scholar] [CrossRef] [PubMed]
- Engineer, C.B.; Hashimoto-Sugimoto, M.; Negi, J.; Israelsson-Nordström, M.; Azoulay-Shemer, T.; Rappel, W.J.; Iba, K.; Schroeder, J.I. CO2 sensing and CO2 regulation of stomatal conductance: Advances and open questions. Trends Plant Sci. 2016, 21, 16–30. [Google Scholar] [CrossRef]
- Kellner, J.; Houska, T.; Manderscheid, R.; Weigel, H.J.; Breuer, L.; Kraft, P. Response of maize biomass and soil water fluxes on elevated CO2 and drought—From field experiments to process-based simulations. Glob. Chang. Biol. 2019, 25, 2947–2957. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.Z.; VanLoocke, A.; Siebers, M.H.; Ruiz-Vera, U.M.; Cody Markelz, R.J.; Leakey, A.D.; Ort, D.R.; Bernacchi, C.J. Future carbon dioxide concentration decreases canopy evapotranspiration and soil water depletion by field-grown maize. Glob. Chang. Biol. 2013, 19, 1572–1584. [Google Scholar] [CrossRef]
- Manderscheid, R.; Erbs, M.; Burkart, S.; Wittich, K.P.; Lopmeier, F.J.; Weigel, H.J. Effects of free-air carbon dioxide enrichment on sap flow and canopy microclimate of maize grown under different water supply. J. Agron. Crop Sci. 2016, 202, 255–268. [Google Scholar] [CrossRef]
- Mwendia, S.W.; Yunusa, I.A.M.; Sindel, B.M.; Whalley, R.D.B.; Bruhl, J.J. Osmotic adjustment, stomata morphology and function show contrasting responses to water stress in mesic and hydric grasses under elevated CO2 concentration. Photosynthetica 2019, 57, 121–131. [Google Scholar] [CrossRef]
- Liu, J.; Kang, S.; Davies, W.J.; Ding, R. Elevated [CO2] alleviates the impacts of water deficit on xylem anatomy and hydraulic properties of maize stems. Plant Cell Environ. 2020, 43, 563–578. [Google Scholar] [CrossRef]
- Wang, A.; Lam, S.K.; Hao, X.; Li, F.Y.; Zong, Y.; Wang, H.; Li, P. Elevated CO2 reduces the adverse effects of drought stress on a high-yielding soybean (Glycine max (L.) Merr.) cultivar by increasing water use efficiency. Plant Physiol. Biochem. 2018, 132, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Bencke-Malato, M.; De Souza, A.P.; Ribeiro-Alves, M.; Schmitz, J.F.; Buckeridge, M.S.; Alves-Ferreira, M. Short-term responses of soybean roots to individual and combinatorial effects of elevated [CO2] and water deficit. Plant Sci. 2019, 280, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Andresen, L.C.; Yuan, N.; Seibert, R.; Moser, G.; Kammann, C.I.; Luterbacher, J.; Erbs, M.; Muller, C. Biomass responses in a temperate European grassland through 17 years of elevated CO2. Glob. Chang. Biol. 2018, 24, 3875–3885. [Google Scholar] [CrossRef] [PubMed]
- Weigel, H.J.; Manderscheid, R. Crop growth responses to free air CO2 enrichment and nitrogen fertilization: Rotating barley, ryegrass, sugar beet and wheat. Eur. J. Agron. 2012, 43, 97–107. [Google Scholar] [CrossRef]
- Leakey, A.D.; Uribelarrea, M.; Ainsworth, E.A.; Naidu, S.L.; Rogers, A.; Ort, D.R.; Long, S.P. Photosynthesis, productivity, and yield of maize are not affected by open-air elevation of CO2 concentration in the absence of drought. Plant Physiol. 2006, 140, 779–790. [Google Scholar] [CrossRef]
- Manderscheid, R.; Erbs, M.; Weigel, H.J. Interactive effects of free-air CO2 enrichment and drought stress on maize growth. Eur. J. Agron. 2014, 52, 11–21. [Google Scholar] [CrossRef]
- Diksaityte, A.; Virsile, A.; Zaltauskaite, J.; Januskaitiene, I.; Juozapaitiene, G. Growth and photosynthetic responses in Brassica napus differ during stress and recovery periods when exposed to combined heat, drought and elevated CO2. Plant Physiol. Biochem. 2019, 142, 59–72. [Google Scholar] [CrossRef]
- South, P.F.; Cavanagh, A.P.; Lopez-Calcagno, P.E.; Raines, C.A.; Ort, D.R. Optimizing photorespiration for improved crop productivity. J. Integr. Plant Biol. 2018, 60, 1217–1230. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef]
- Poorter, H.; Knopf, O.; Wright, I.J.; Temme, A.A.; Hogewoning, S.W.; Graf, A.; Cernusak, L.A.; Pons, T.L. A meta-analysis of responses of C3 plants to atmospheric CO2: Dose–response curves for 85 traits ranging from the molecular to the whole-plant level. New Phytol. 2022, 233, 1560–1596. [Google Scholar] [CrossRef]
- Rosa, B.L.; Souza, J.P.; Pereira, E.G. Increased atmospheric CO2 changes the photosynthetic responses of Acrocomia aculeata (Arecaceae) to drought. Acta Bot. Bras. 2019, 33, 486–497. [Google Scholar] [CrossRef]
- Avila, R.T.; de Almeida, W.L.; Costa, L.C.; Machado, K.L.; Barbosa, M.L.; de Souza, R.P.; Martino, P.B.; Juárez, M.A.; Marçal, D.M.; Martins, S.C.; et al. Elevated air [CO2] improves photosynthetic performance and alters biomass accumulation and partitioning in drought-stressed coffee plants. Environ. Exp. Bot. 2020, 177, 104137. [Google Scholar] [CrossRef]
- Li, B.; Feng, Y.; Zong, Y.; Zhang, D.; Hao, X.; Li, P. Elevated CO2-induced changes in photosynthesis, antioxidant enzymes and signal transduction enzyme of soybean under drought stress. Plant Physiol. Biochem. 2020, 154, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Ewert, F. Modelling plant responses to elevated CO2: How important is leaf area index? Ann. Bot. 2004, 93, 619–627. [Google Scholar] [CrossRef]
- Lemordant, L.; Gentine, P.; Swann, A.S.; Cook, B.I.; Scheff, J. Critical impact of vegetation physiology on the continental hydrologic cycle in response to increasing CO2. Proc. Natl. Acad. Sci. USA 2018, 115, 4093–4098. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Liu, J.; Leung, F. A framework to quantify impacts of elevated CO2 concentration, global warming and leaf area changes on seasonal variations of water resources on a river basin scale. J. Hydrol. 2019, 570, 508–522. [Google Scholar] [CrossRef]
- Jena, U.R.; Bhattacharya, S.; Swain, D.K.; Maiti, M.K. Differential effect of elevated carbon dioxide on sucrose transport and accumulation in developing grains of three rice cultivars. Plant Gene 2021, 28, 100337. [Google Scholar] [CrossRef]
- Rao, X.; Dixon, R.A. The differences between NAD-ME and NADP-ME subtypes of C4 photosynthesis: More than decarboxylating enzymes. Front. Plant Sci. 2016, 7, 1525. [Google Scholar] [CrossRef]
- Maier, A.; Zell, M.B.; Maurino, V.G. Malate decarboxylases: Evolution and roles of NAD (P)-ME isoforms in species performing C4 and C3 photosynthesis. J. Exp. Bot. 2011, 62, 3061–3069. [Google Scholar] [CrossRef]
- Tronconi, M.A.; Andreo, C.S.; Drincovich, M.F. Chimeric structure of plant malic enzyme family: Different evolutionary scenarios for NAD-and NADP-dependent isoforms. Front. Plant Sci. 2018, 9, 565. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, M.; Busch, F.A.; Khoshravesh, R.; Covshoff, S. Understanding C4 evolution and function. Front. Plant Sci. 2021, 12, 774818. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Wang, Y.; Xia, C.; Zhang, Y.; Zhang, H. Chloroplastic SaNADP-ME4 of C3–C4 woody desert species Salsola laricifolia confers drought and salt stress resistance to Arabidopsis. Plants 2021, 10, 1827. [Google Scholar] [CrossRef] [PubMed]
- Begara-Morales, J.C.; Sánchez-Calvo, B.; Gómez-Rodríguez, M.V.; Chaki, M.; Valderrama, R.; Mata-Pérez, C.; López-Jaramillo, J.; Corpas, F.J.; Barroso, J.B. Short-term low temperature induces nitro-oxidative stress that deregulates the NADP-malic enzyme function by tyrosine nitration in Arabidopsis thaliana. Antioxidants 2019, 8, 448. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, B.; Ding, H.; Zhang, J.; Li, S. The role of NADP-malic enzyme in plants under stress. Plant Sci. 2019, 281, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Correia, P.M.; da Silva, A.B.; Vaz, M.; Carmo-Silva, E.; da Silva, J.M. Efficient regulation of CO2 assimilation enables greater resilience to high temperature and drought in Maize. Front. Plant Sci. 2021, 12, 675546. [Google Scholar] [CrossRef]
- Sun, X.; Han, G.; Meng, Z.; Lin, L.; Sui, N. Roles of malic enzymes in plant development and stress responses. Plant Signal. Behav. 2019, 14, 1644596. [Google Scholar] [CrossRef]
- Medina, S.; Vicente, R.; Amador, A.; Araus, J.L. Interactive effects of elevated [CO2] and water stress on physiological traits and gene expression during vegetative growth in four durum wheat genotypes. Front. Plant Sci. 2016, 7, 1738. [Google Scholar] [CrossRef]
- Curtis, P.S.; Drake, B.G.; Whigham, D.F. Nitrogen and carbon dynamics in C3 and C4 estuarine marsh plants grown under elevated CO2 in situ. Oecologia 1989, 78, 297–301. [Google Scholar] [CrossRef]
- Delucia, E.H.; Sasek, T.W.; Strain, B.R. Photosynthetic inhibition after long-term exposure to elevated levels of atmospheric carbon dioxide. Photosynth. Res. 1985, 7, 175–184. [Google Scholar] [CrossRef]
- Jin, J.; Armstrong, R.; Tang, C. Impact of elevated CO2 on grain nutrient concentration varies with crops and soils—A long-term FACE study. Sci. Total Environ. 2019, 651, 2641–2647. [Google Scholar] [CrossRef]
- Houshmandfar, A.; Fitzgerald, G.J.; O’Leary, G.; Tausz-Posch, S.; Fletcher, A.; Tausz, M. The relationship between transpiration and nutrient uptake in wheat changes under elevated atmospheric CO2. Physiol. Plant. 2018, 163, 516–529. [Google Scholar] [CrossRef]
- Adavi, S.B.; Sathee, L. Elevated CO2 alters tissue balance of nitrogen metabolism and downregulates nitrogen assimilation and signalling gene expression in wheat seedlings receiving high nitrate supply. Protoplasma 2021, 258, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Adavi, S.B.; Sathee, L. Elevated CO2 differentially regulates root nitrate transporter kinetics in a genotype and nitrate dose-dependent manner. Plant Sci. 2021, 305, 110807. [Google Scholar] [CrossRef] [PubMed]
- Umnajkitikorn, K.; Sade, N.; Rubio Wilhelmi, M.D.M.; Gilbert, M.E.; Blumwald, E. Silencing of OsCV (chloroplast vesiculation) maintained photorespiration and N assimilation in rice plants grown under elevated CO2. Plant Cell Environ. 2020, 43, 920–933. [Google Scholar] [CrossRef]
- Jin, J.; Lauricella, D.; Armstrong, R.; Sale, P.; Tang, C. Phosphorus application and elevated CO2 enhance drought tolerance in field pea grown in a phosphorus-deficient vertisol. Ann. Bot. 2015, 116, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.; Yilmaz, O.; Ozturk, L. Potassium deficiency impedes elevated carbon dioxide-induced biomass enhancement in well watered or drought-stressed bread wheat. J. Plant Nutr. Soil Sci. 2017, 180, 474–481. [Google Scholar] [CrossRef]
- Thruppoyil, S.B.; Ksiksi, T. Time-dependent stomatal conductance and growth responses of Tabernaemontana divaricata to short-term elevated CO2 and water stress at higher than optimal growing temperature. Curr. Plant Biol. 2020, 22, 100127. [Google Scholar] [CrossRef]
- Birami, B.; Nägele, T.; Gattmann, M.; Preisler, Y.; Gast, A.; Arneth, A.; Ruehr, N.K. Hot drought reduces the effects of elevated CO2 on tree water-use efficiency and carbon metabolism. New Phytol. 2020, 226, 1607–1621. [Google Scholar] [CrossRef] [PubMed]
- Paudel, I.; Halpern, M.; Wagner, Y.; Raveh, E.; Yermiyahu, U.; Hoch, G.; Klein, T. Elevated CO2 compensates for drought effects in lemon saplings via stomatal downregulation, increased soil moisture, and increased wood carbon storage. Environ. Exp. Bot. 2018, 148, 117–127. [Google Scholar] [CrossRef]
- Cruz, J.L.; LeCain, D.R.; Alves, A.A.; Coelho Filho, M.A.; Coelho, E.F. Elevated CO2 reduces whole transpiration and substantially improves root production of cassava grown under water deficit. Arch. Agron. Soil Sci. 2018, 64, 1623–1634. [Google Scholar] [CrossRef]
- Parvin, S.; Uddin, S.; Tausz-Posch, S.; Fitzgerald, G.; Armstrong, R.; Tausz, M. Elevated CO2 improves yield and N2 fixation but not grain N concentration of faba bean (Vicia faba L.) subjected to terminal drought. Environ. Exp. Bot. 2019, 165, 161–173. [Google Scholar] [CrossRef]
- De Oliveira, M.F.; Marenco, R.A. Gas exchange, biomass allocation and water-use efficiency in response to elevated CO2 and drought in andiroba (Carapa surinamensis, Meliaceae). iForest-Biogeosci. For. 2019, 12, 61–68. [Google Scholar] [CrossRef]
- Souza, J.P.; Melo, N.M.; Pereira, E.G.; Halfeld, A.D.; Gomes, I.N.; Prado, C.H.B. Responses of woody Cerrado species to rising atmospheric CO2 concentration and water stress: Gains and losses. Funct. Plant Biol. 2016, 43, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J.R.; Patterson, A.E.; Rodrigues, W.P.; Campostrini, E.; Griffin, K.L. Photosynthetic acclimation to elevated CO2 combined with partial rootzone drying results in improved water use efficiency, drought tolerance and leaf carbon balance of grapevines (Vitis labrusca). Environ. Exp. Bot. 2017, 134, 82–95. [Google Scholar] [CrossRef]
- Li, X.; Kang, S.; Zhang, X.; Li, F.; Lu, H. Deficit irrigation provokes more pronounced responses of maize photosynthesis and water productivity to elevated CO2. Agric. Water Manag. 2018, 195, 71–83. [Google Scholar] [CrossRef]
- Liu, B.B.; Li, M.; Li, Q.M.; Cui, Q.Q.; Zhang, W.D.; Ai, X.Z.; Bi, H.G. Combined effects of elevated CO2 concentration and drought stress on photosynthetic performance and leaf structure of cucumber (Cucumis sativus L.) seedlings. Photosynthetica 2018, 56, 942–952. [Google Scholar] [CrossRef]

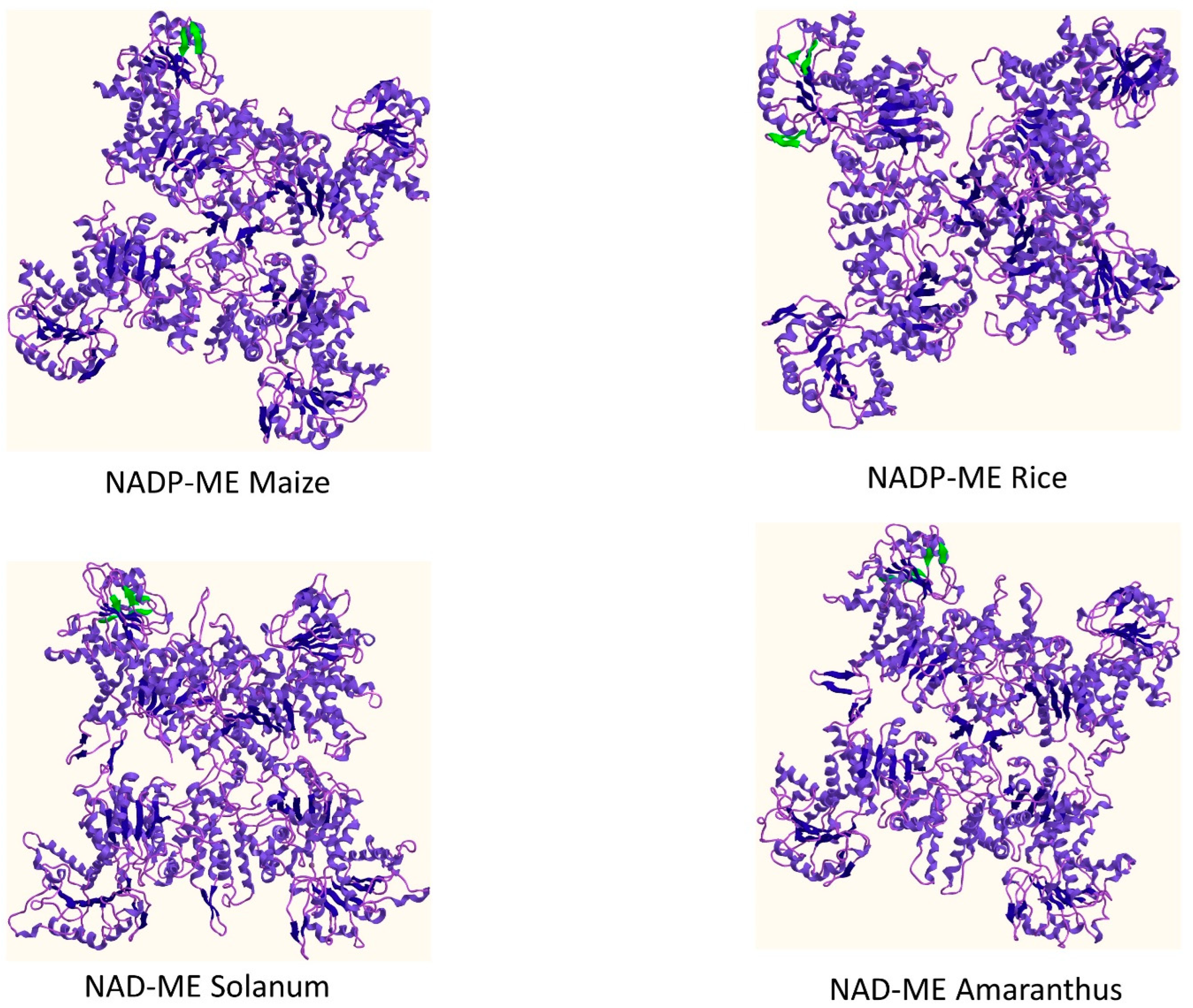
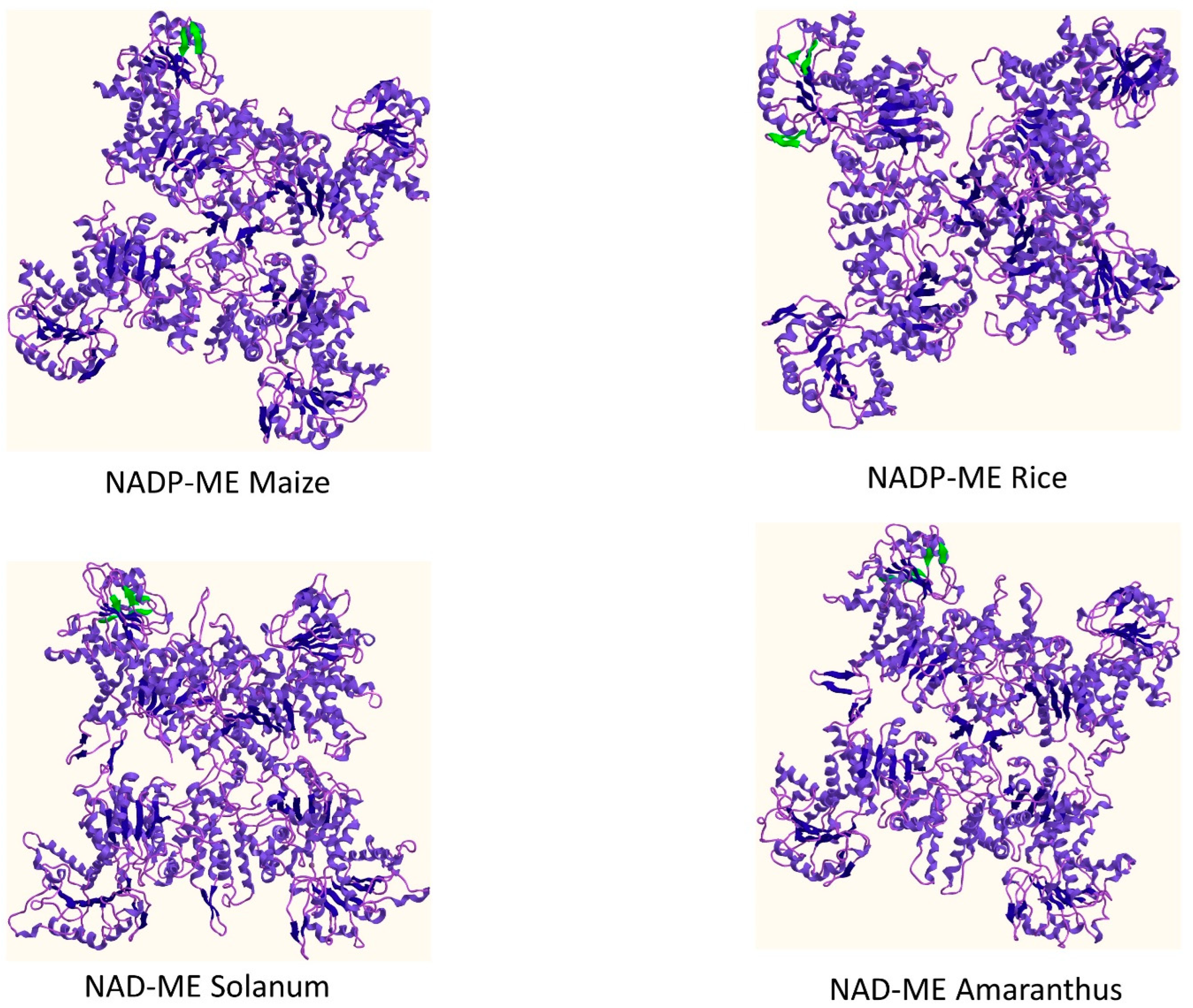
| Enzyme | Sheets | Beta Alpha Beta Units | Beta Hairpins | Beta Bulge | Strands | Helices | Helix-Helix Interaction | Beta Turns | Gamma Turns |
|---|---|---|---|---|---|---|---|---|---|
| NADP-ME Maize | 4 | 5 | 1 | 0 | 14 | 33 | 47 | 36 | 3 |
| NADP-ME Rice | 4 | 5 | 2 | 0 | 15 | 33 | 44 | 33 | 4 |
| NAD-ME Solanum | 3 | 5 | 2 | 1 | 14 | 35 | 44 | 47 | 6 |
| NAD-ME Amaranthus | 4 | 5 | 2 | 1 | 14 | 36 | 45 | 39 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shanker, A.K.; Gunnapaneni, D.; Bhanu, D.; Vanaja, M.; Lakshmi, N.J.; Yadav, S.K.; Prabhakar, M.; Singh, V.K. Elevated CO2 and Water Stress in Combination in Plants: Brothers in Arms or Partners in Crime? Biology 2022, 11, 1330. https://doi.org/10.3390/biology11091330
Shanker AK, Gunnapaneni D, Bhanu D, Vanaja M, Lakshmi NJ, Yadav SK, Prabhakar M, Singh VK. Elevated CO2 and Water Stress in Combination in Plants: Brothers in Arms or Partners in Crime? Biology. 2022; 11(9):1330. https://doi.org/10.3390/biology11091330
Chicago/Turabian StyleShanker, Arun Kumar, Deepika Gunnapaneni, Divya Bhanu, Maddi Vanaja, Narayana Jyothi Lakshmi, Sushil Kumar Yadav, Mathyam Prabhakar, and Vinod Kumar Singh. 2022. "Elevated CO2 and Water Stress in Combination in Plants: Brothers in Arms or Partners in Crime?" Biology 11, no. 9: 1330. https://doi.org/10.3390/biology11091330
APA StyleShanker, A. K., Gunnapaneni, D., Bhanu, D., Vanaja, M., Lakshmi, N. J., Yadav, S. K., Prabhakar, M., & Singh, V. K. (2022). Elevated CO2 and Water Stress in Combination in Plants: Brothers in Arms or Partners in Crime? Biology, 11(9), 1330. https://doi.org/10.3390/biology11091330