
Genome-Based Analysis of Verticillium Polyketide Synthase Gene Clusters


 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Full Genome Sequences
2.2. Prediction, Annotation, and Identification of Polyketide Synthase Genes and Gene Clusters
2.3. Prediction of Verticillium PKS Domains
2.4. PKS Cluster Comparison Analysis of Verticillium and Other Fungi
2.5. Phylogenetic Analysis
3. Results
3.1. Prediction of Verticillium PKS Domains
3.2. Comparison of Verticillium PKS Cluster with Those of Other Fungi
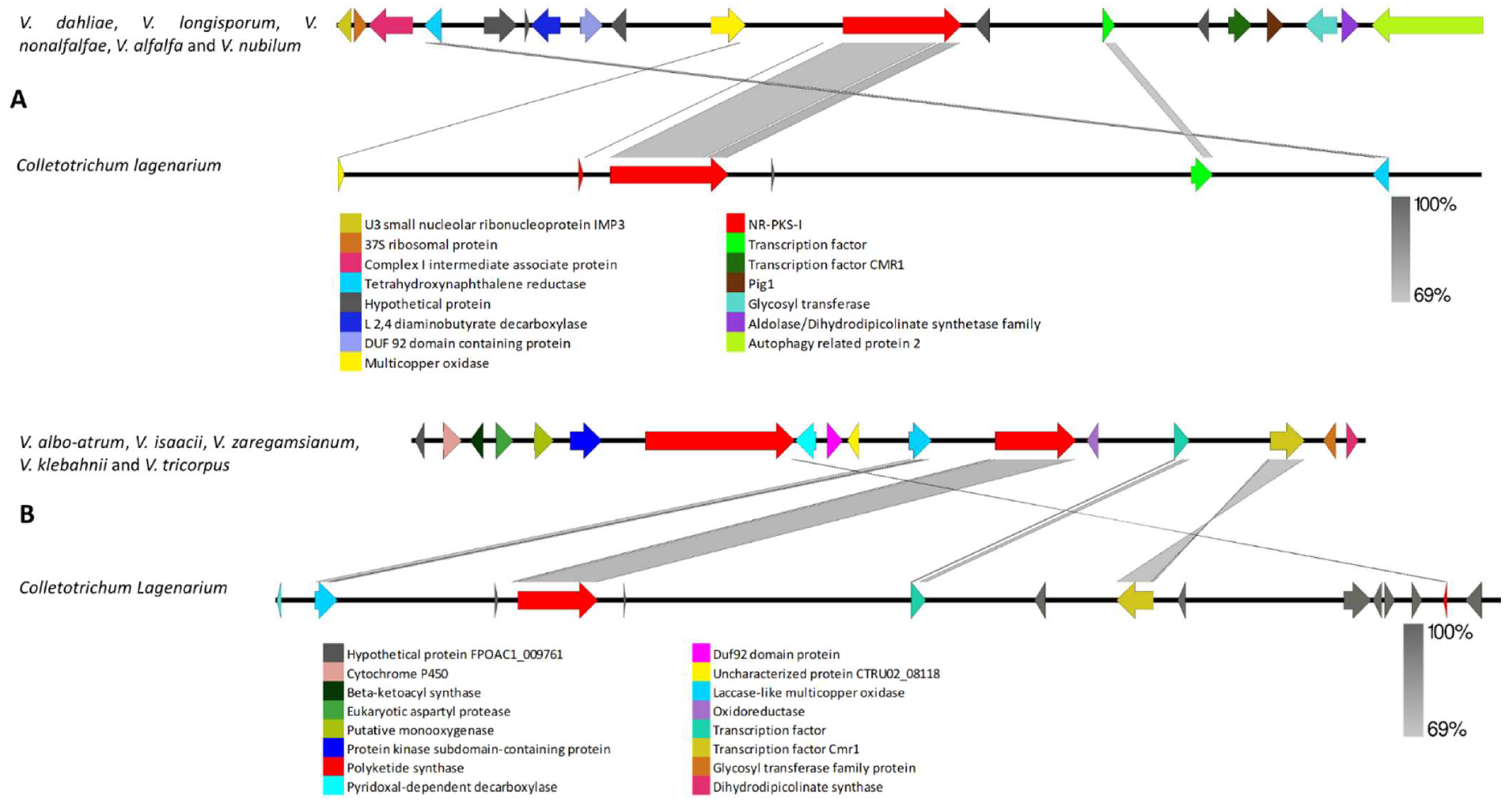
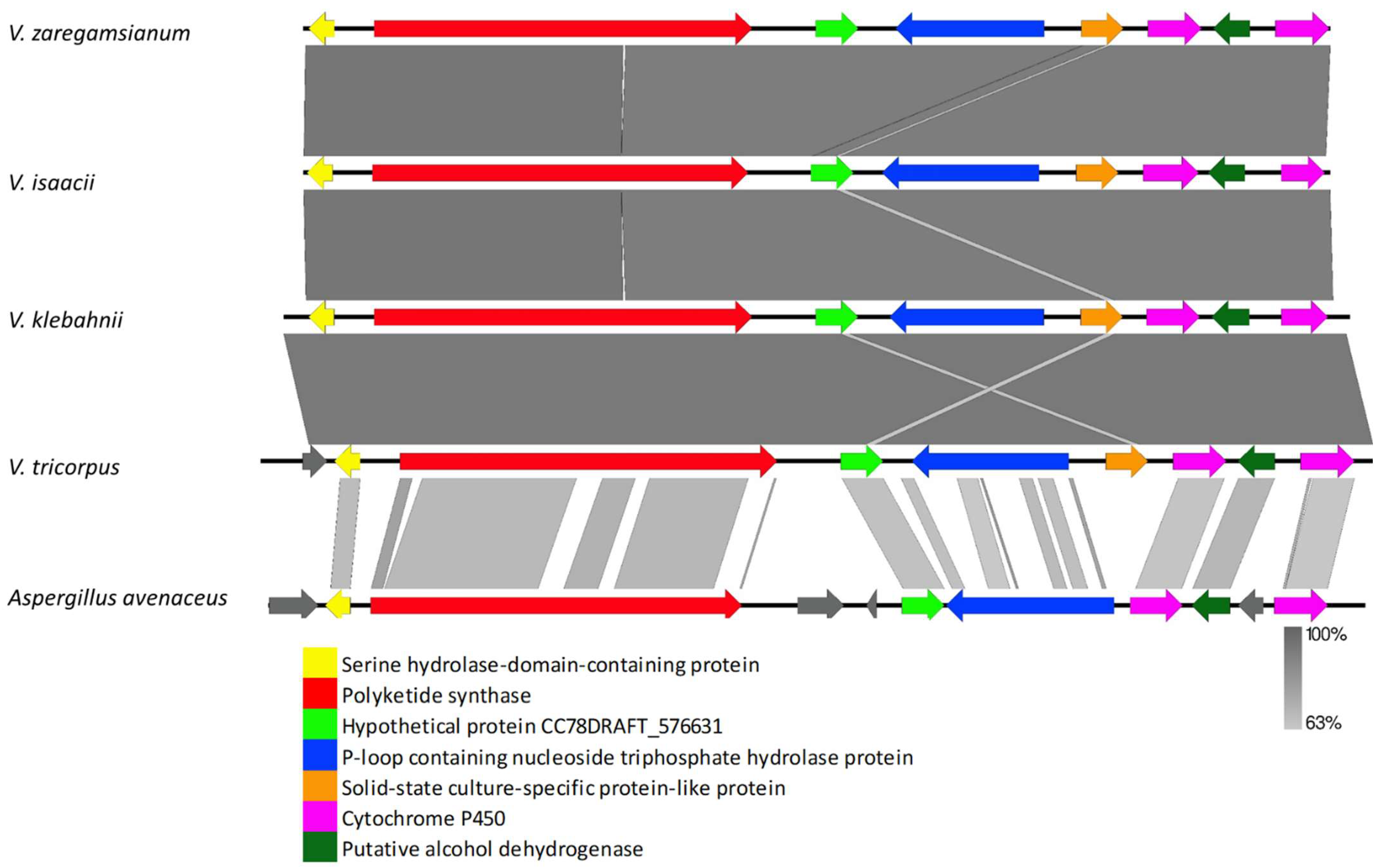
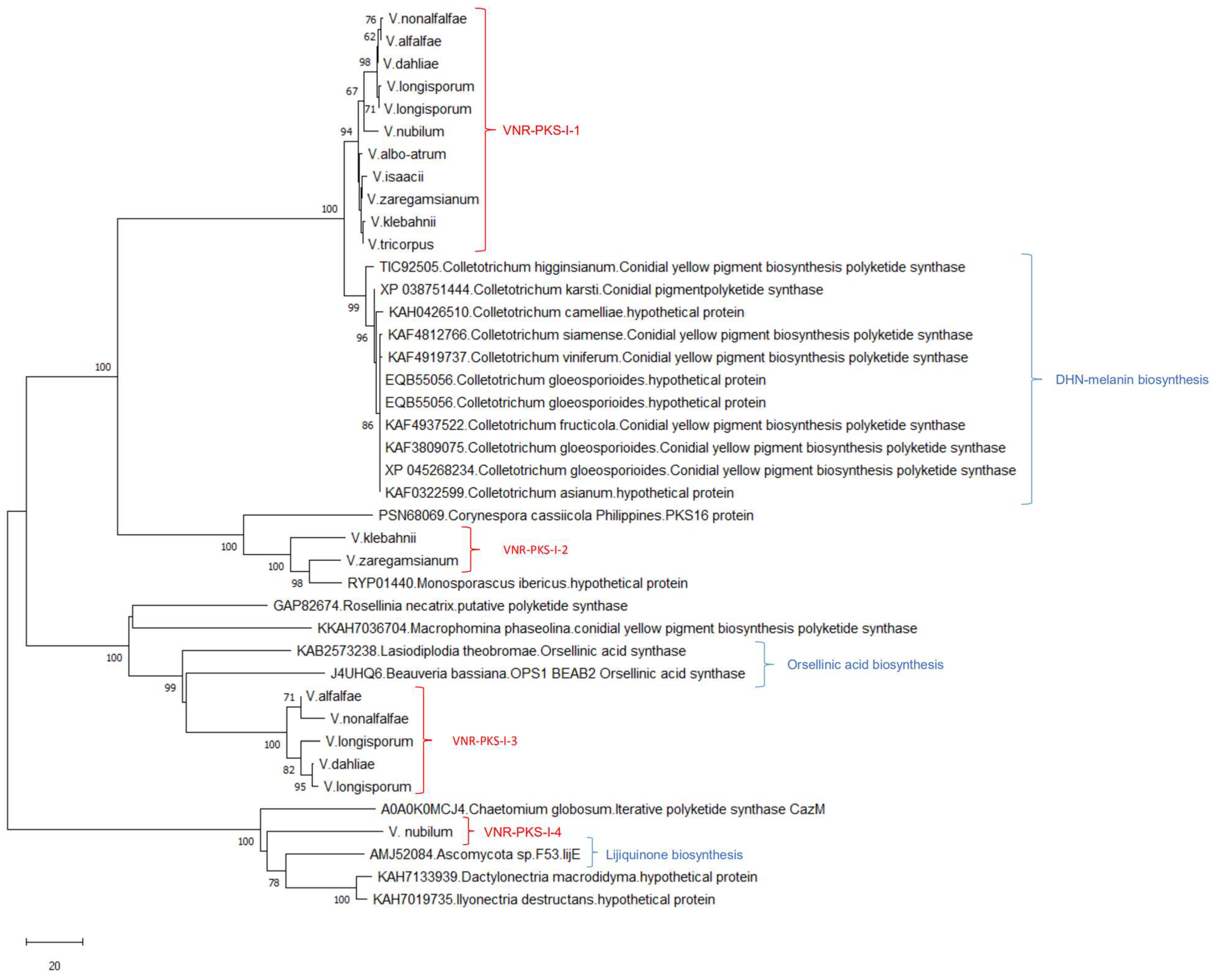
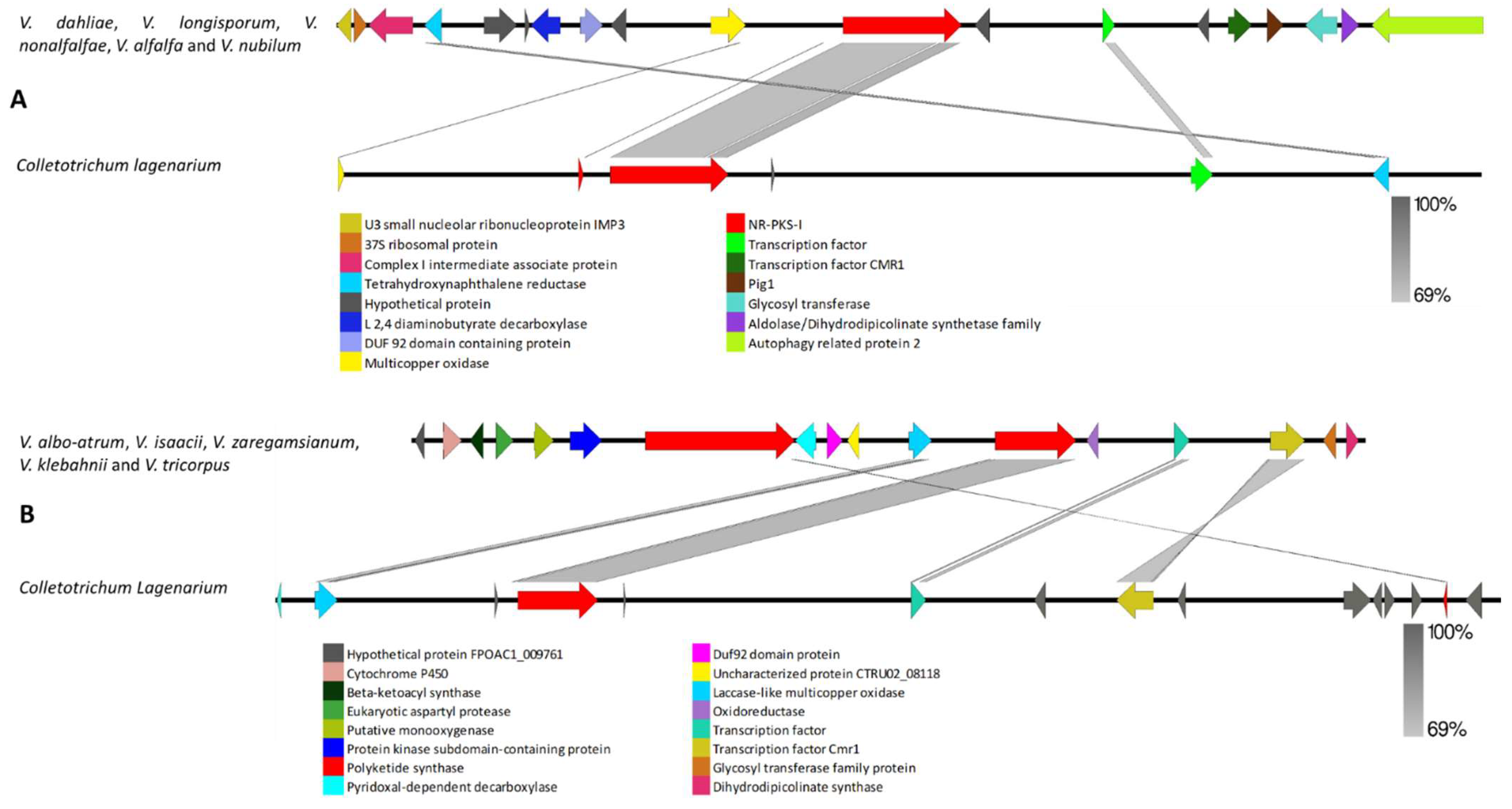
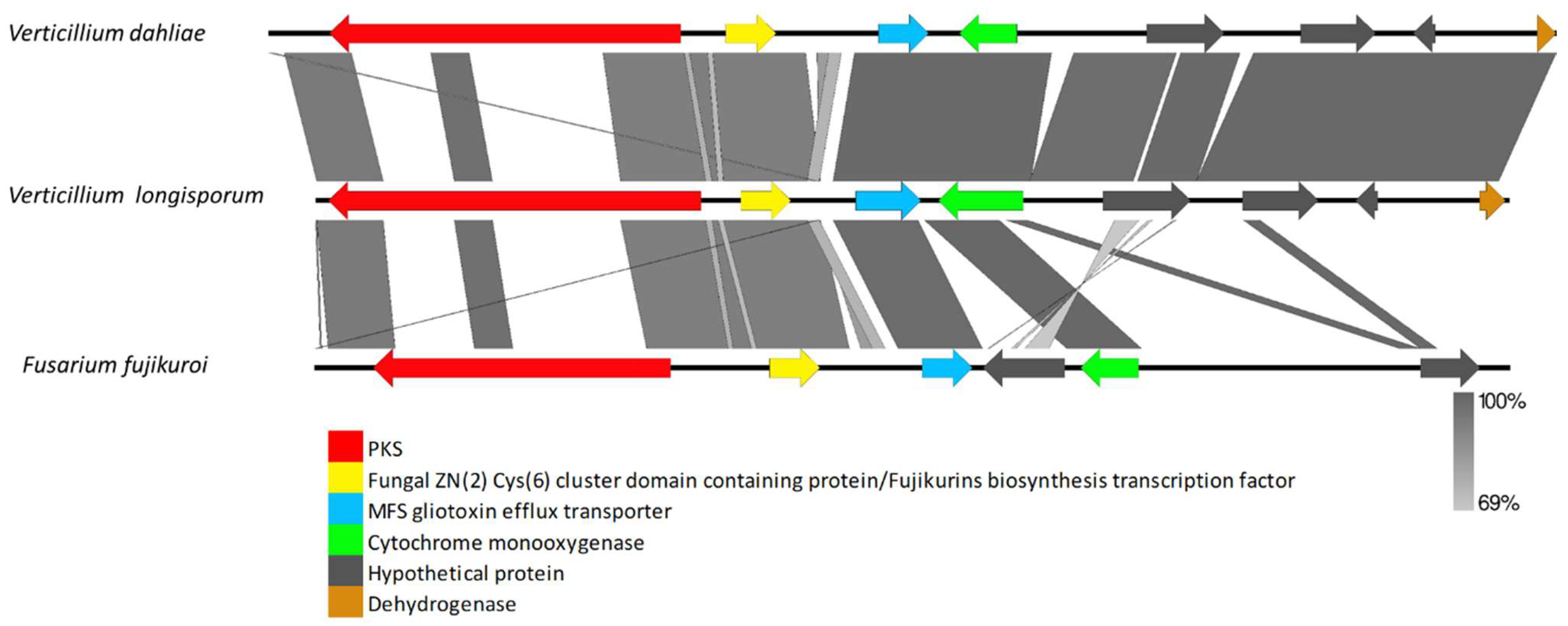
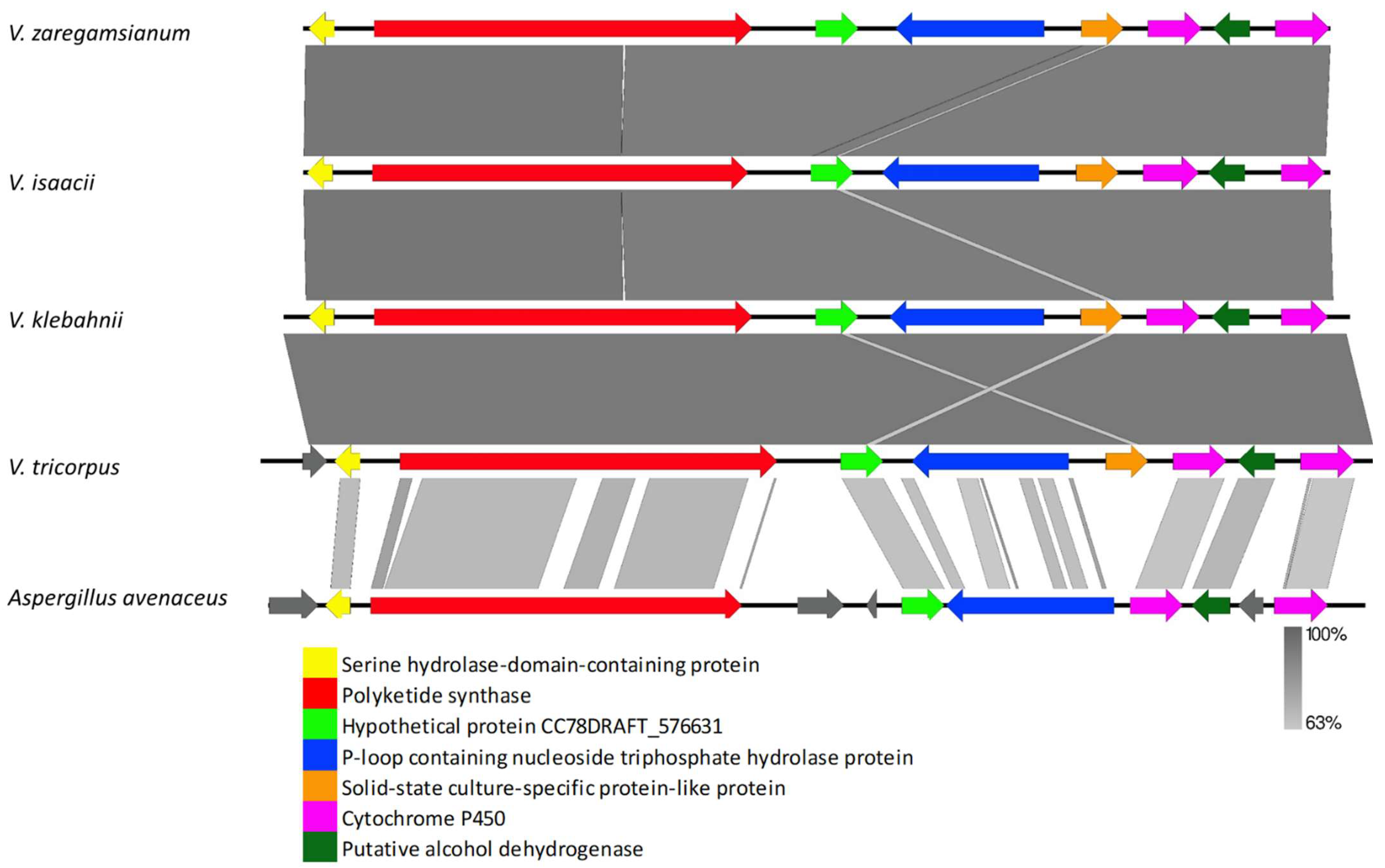
3.2.1. Clusters Common among All Examined Verticillium Genomes
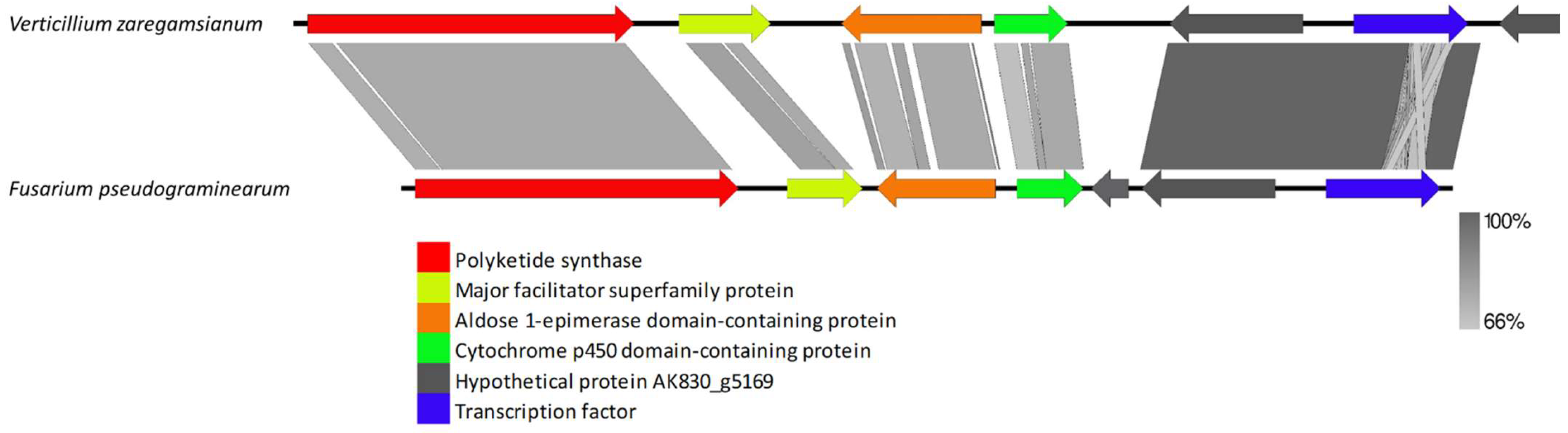
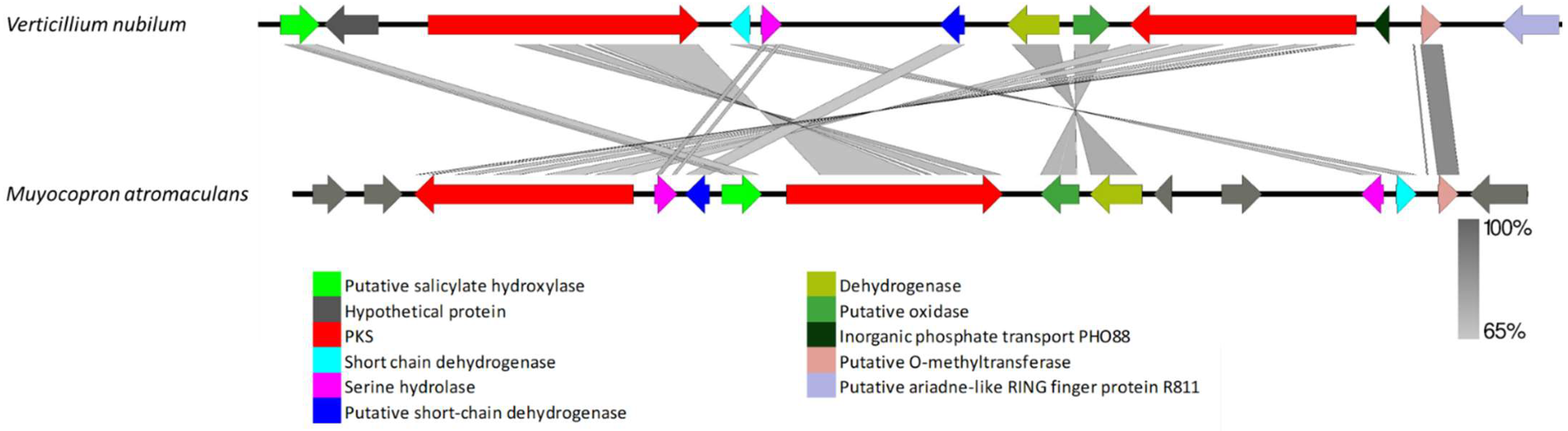
3.2.2. Clusters Missing in a Few Genomes but Present in Other Species
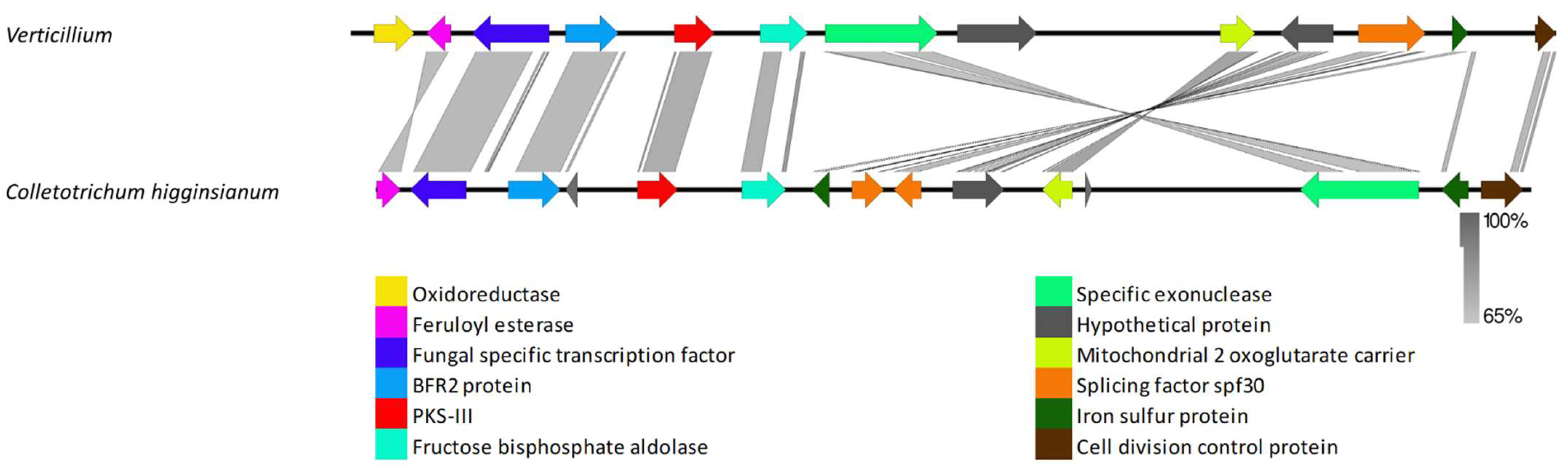
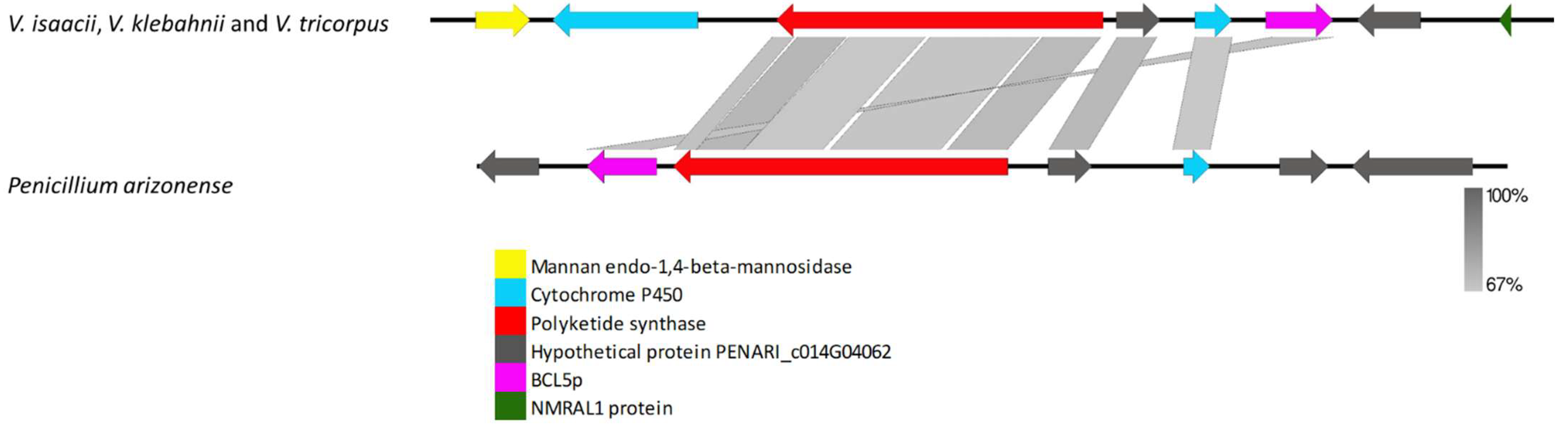
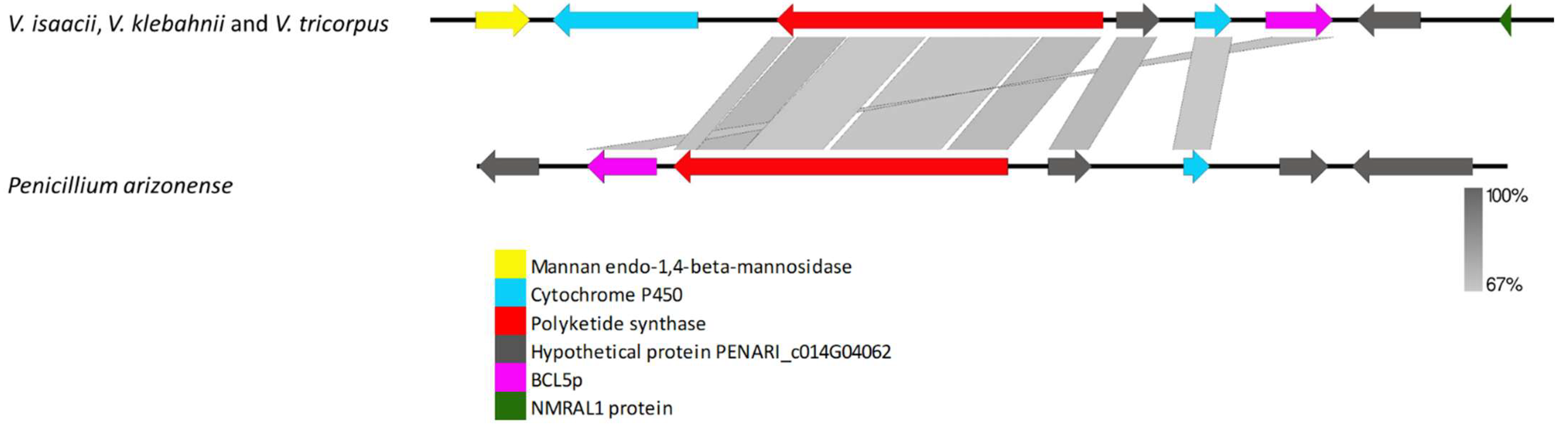
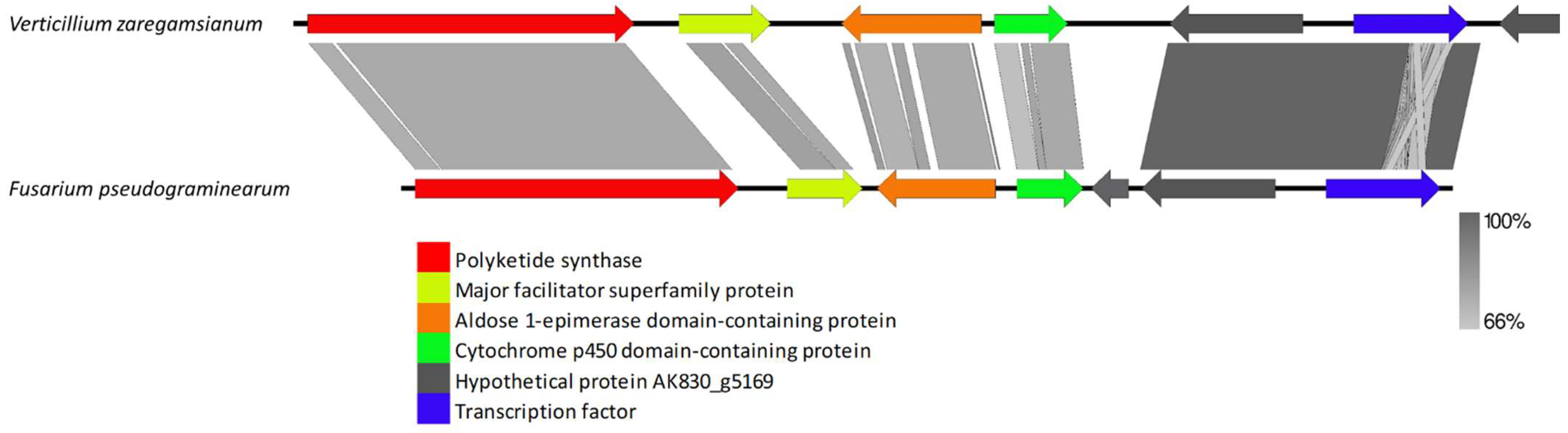
3.2.3. Species-Specific Clusters
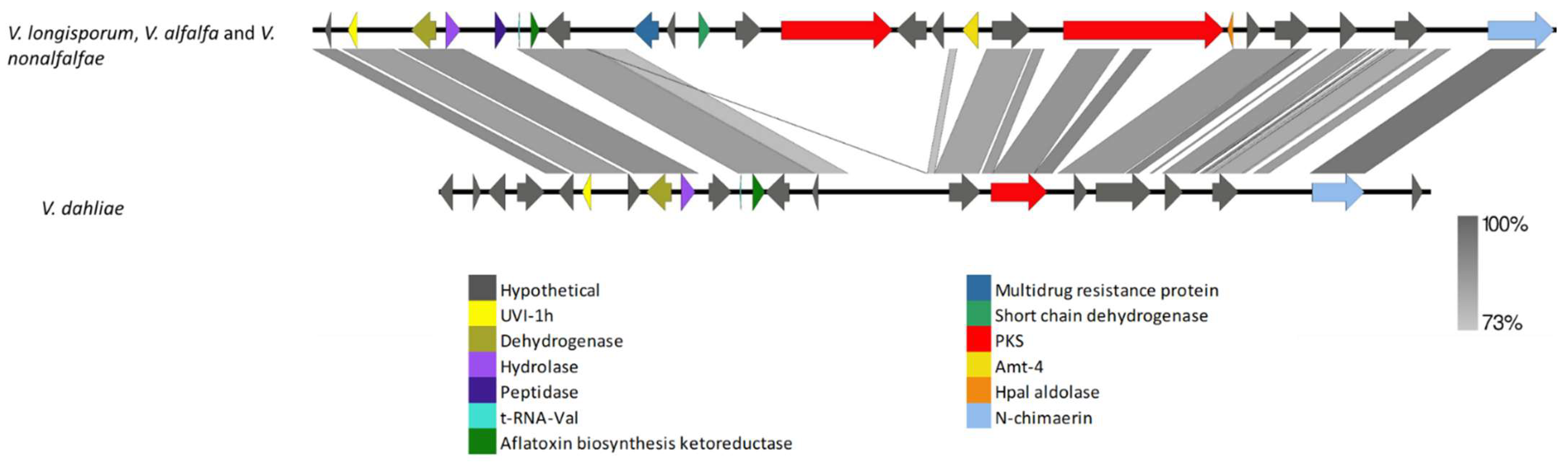
3.3. Missing PKS Gene Clusters in Closely Related Species
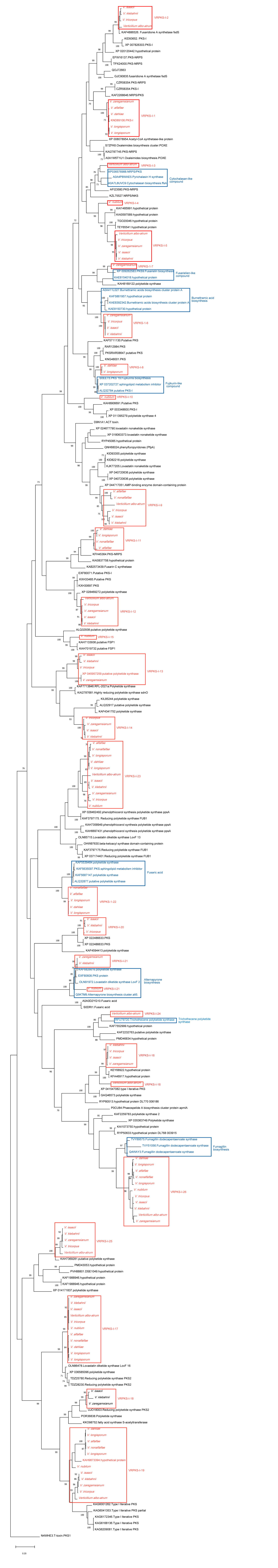
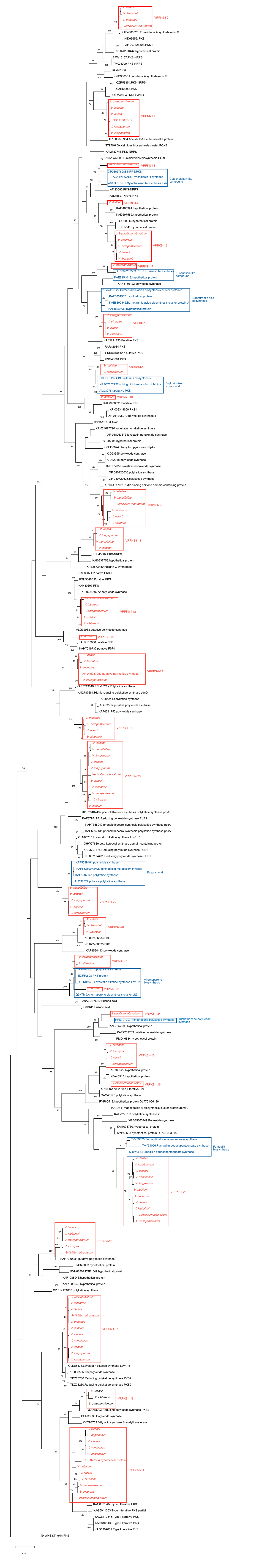
3.4. Phylogenetic Analysis
4. Discussion
4.1. PKS Identification
4.2. Clusters Common among All Examined Verticillium Genomes
4.3. Species-Specific Clusters
4.4. Clusters Missing in a Few Genomes but Present in Others
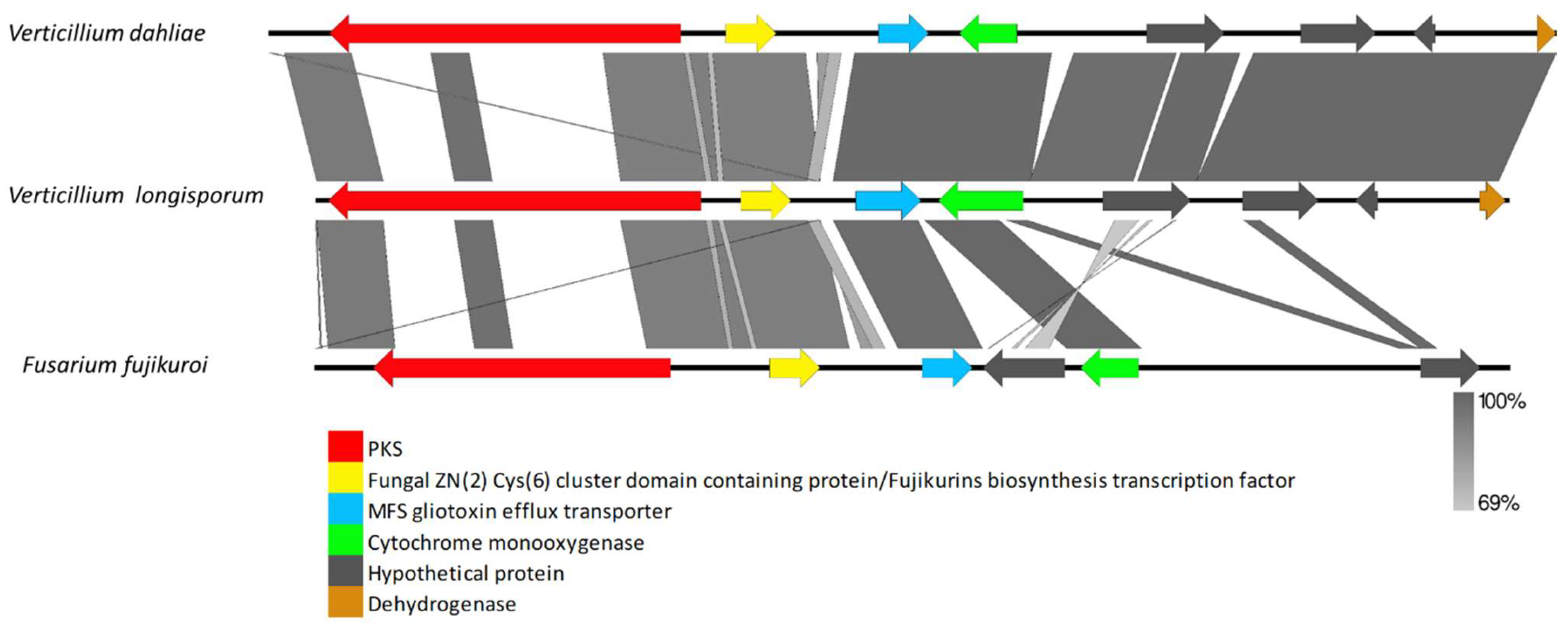
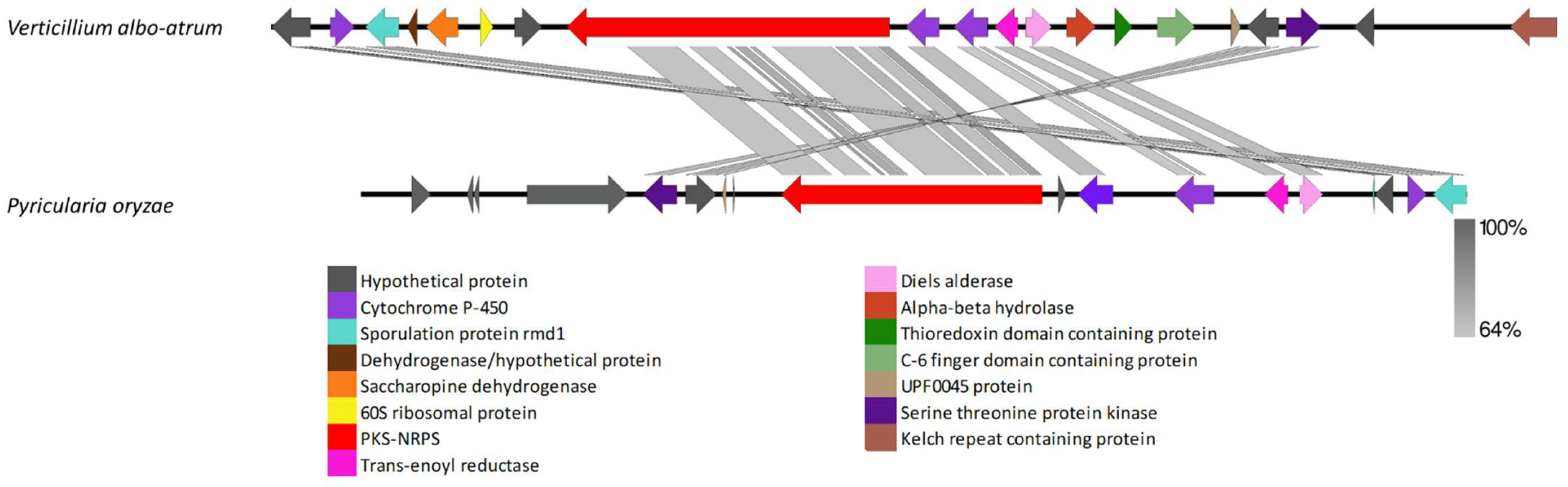
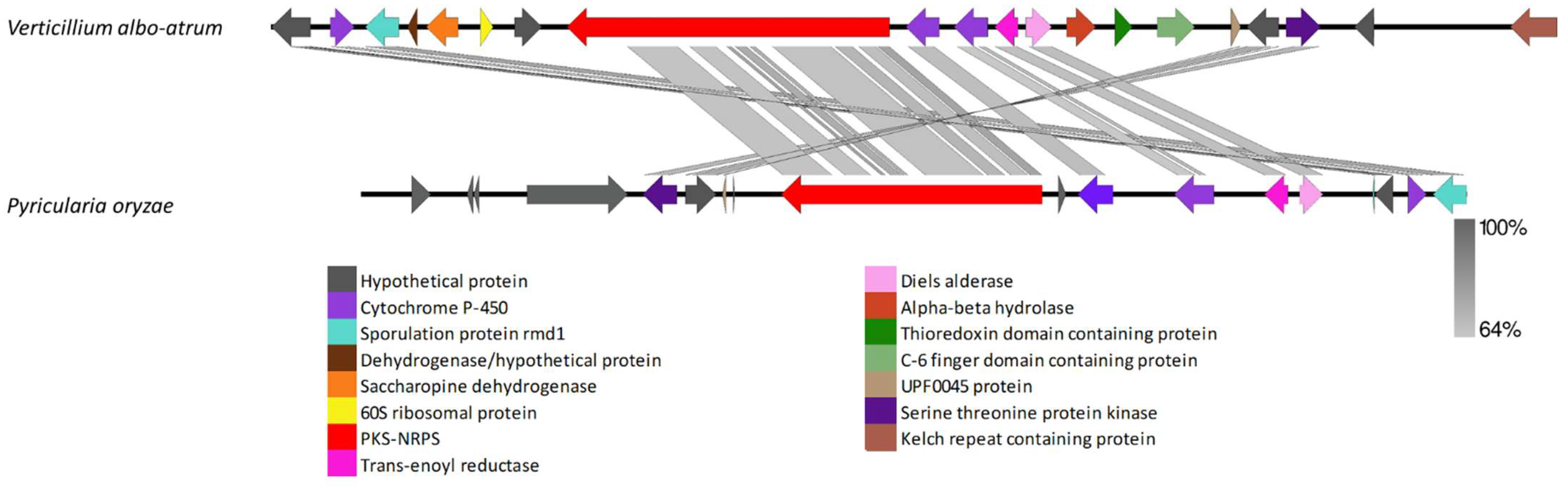
4.5. Clusters Found in Distantly Related Fungi
4.6. Comparison of the Number of PKS Genes in Pathogenic and Non-Pathogenic Verticillium Strains
4.7. Missing PKS Gene Clusters in Closely Related Species
4.8. PKS-NRPS Cluster Comparison among Different Verticillium Strains
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pegg, G.F.; Brady, B.L. Verticillium Wilts; CABI: Wallingford, UK, 2002. [Google Scholar]
- Klosterman, S.J.; Atallah, Z.K.; Vallad, G.E.; Subbarao, K.V. Diversity, pathogenicity, and management of Verticillium species. Annu. Rev. Phytopathol. 2009, 47, 39–62. [Google Scholar] [CrossRef] [PubMed]
- Barbara, D.J.; Clewes, E. Plant pathogenic Verticillium species: How many of them are there? Mol. Plant Pathol. 2003, 4, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Rowe, R.C.; Powelson, M.L. Potato early dying: Management challenges in a changing production environment. Plant Dis. 2002, 86, 1184–1193. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S. Longevity of the Verticillium wilt fungus in the laboratory and field. Phytopathology 1955, 45, 180–181. [Google Scholar]
- Song, R.; Li, J.; Xie, C.; Jian, W.; Yang, X. An overview of the molecular genetics of plant resistance to the Verticillium wilt pathogen Verticillium dahliae. Int. J. Mol. Sci. 2020, 21, 1120. [Google Scholar] [CrossRef]
- Wu, L.; Du, G.; Bao, R.; Li, Z.; Gong, Y.; Liu, F. De novo assembly and discovery of genes involved in the response of Solanum sisymbriifolium to Verticillium dahlia. Physiol. Mol. Biol. Plants 2019, 25, 1009–1027. [Google Scholar] [CrossRef]
- Luo, X.; Xie, C.; Dong, J.; Yang, X.; Sui, A. Interactions between Verticillium dahliae and its host: Vegetative growth, pathogenicity, plant immunity. Appl. Microbiol. Biotechnol. 2014, 98, 6921–6932. [Google Scholar] [CrossRef]
- Depotter, J.R.; Shi-Kunne, X.; Missonnier, H.; Liu, T.; Faino, L.; van den Berg, G.C.; Wood, T.A.; Zhang, B.; Jacques, A.; Seidl, M.F. Dynamic virulence-related regions of the plant pathogenic fungus Verticillium dahliae display enhanced sequence conservation. Mol. Ecol. 2019, 28, 3482–3495. [Google Scholar] [CrossRef]
- Zhang, D.-D.; Wang, X.-Y.; Chen, J.-Y.; Kong, Z.-Q.; Gui, Y.-J.; Li, N.-Y.; Bao, Y.-M.; Dai, X.-F. Identification and characterization of a pathogenicity-related gene VdCYP1 from Verticillium dahliae. Sci. Rep. 2016, 6, 27979. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, X.; Fang, Y.; Anchieta, A.; Goldman, P.H.; Hernandez, G.; Klosterman, S.J. Transcription factor VdCmr1 is required for pigment production, protection from UV irradiation, and regulates expression of melanin biosynthetic genes in Verticillium dahliae. Microbiology 2018, 164, 685. [Google Scholar] [CrossRef]
- Zhu, X.; Sayari, M.; Daayf, F. Role of Exopolygalacturonase-Related Genes in Potato-Verticillium dahliae Interaction. Pathogens 2021, 10, 642. [Google Scholar] [CrossRef]
- Zhu, X.; Soliman, A.; Islam, M.R.; Adam, L.R.; Daayf, F. Verticillium dahliae’s isochorismatase hydrolase is a virulence factor that contributes to interference with potato’s salicylate and jasmonate defense signaling. Front. Plant Sci. 2017, 8, 399. [Google Scholar] [CrossRef]
- Zhu, X.; Sayari, M.; Islam, M.R.; Daayf, F. NOXA Is Important for Verticillium dahliae’s Penetration Ability and Virulence. J. Fungi 2021, 7, 814. [Google Scholar] [CrossRef]
- Vining, L.C. Functions of secondary metabolites. Annu. Rev. Microbiol. 1990, 44, 395–427. [Google Scholar] [CrossRef]
- Mosunova, O.; Navarro-Muñoz, J.C.; Collemare, J. The biosynthesis of fungal secondary metabolites: From fundamentals to biotechnological applications. Ref. Modul. Life Sci. 2021, 2, 458–476. [Google Scholar]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef]
- Awad, N.E.; Kassem, H.A.; Hamed, M.A.; El-Feky, A.M.; Elnaggar, M.A.; Mahmoud, K.; Ali, M.A. Isolation and characterization of the bioactive metabolites from the soil derived fungus Trichoderma viride. Mycology 2018, 9, 70–80. [Google Scholar] [CrossRef]
- Cole, R.J.; Schweikert, M.A.; Jarvis, B.B. Handbook of Secondary Fungal Metabolites; Gulf Professional Publishing: Houston, TX, USA, 2003; Volume 3. [Google Scholar]
- Scharf, D.H.; Heinekamp, T.; Brakhage, A.A. Human and plant fungal pathogens: The role of secondary metabolites. PLoS Pathog. 2014, 10, e1003859. [Google Scholar] [CrossRef]
- Shi-Kunne, X.; Jové, R.d.P.; Depotter, J.R.; Ebert, M.K.; Seidl, M.F.; Thomma, B.P. In silico prediction and characterisation of secondary metabolite clusters in the plant pathogenic fungus Verticillium dahliae. FEMS Microbiol. Lett. 2019, 366, fnz081. [Google Scholar] [CrossRef]
- Brown, D.W.; Proctor, R.H. Insights into natural products biosynthesis from analysis of 490 polyketide synthases from Fusarium. Fungal Genet. Biol. 2016, 89, 37–51. [Google Scholar] [CrossRef]
- Collemare, J.; Billard, A.; Böhnert, H.U.; Lebrun, M.-H. Biosynthesis of secondary metabolites in the rice blast fungus Magnaporthe grisea: The role of hybrid PKS-NRPS in pathogenicity. Mycol. Res. 2008, 112, 207–215. [Google Scholar] [CrossRef]
- Keller, N.P. Translating biosynthetic gene clusters into fungal armor and weaponry. Nat. Chem. Biol. 2015, 11, 671–677. [Google Scholar] [CrossRef]
- Dias, D.A.; Urban, S.; Roessner, U. A historical overview of natural products in drug discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef]
- Boruta, T. Uncovering the repertoire of fungal secondary metabolites: From Fleming’s laboratory to the International Space Station. Bioengineered 2018, 9, 12–16. [Google Scholar] [CrossRef]
- Lobanovska, M.; Pilla, G. Focus: Drug development: Penicillin’s discovery and antibiotic resistance: Lessons for the future? Yale J. Biol. Med. 2017, 90, 135. [Google Scholar]
- Medema, M.H.; Fischbach, M.A. Computational approaches to natural product discovery. Nat. Chem. Biol. 2015, 11, 639–648. [Google Scholar] [CrossRef]
- Wiemann, P.; Keller, N.P. Strategies for mining fungal natural products. J. Ind. Microbiol. Biotechnol. 2014, 41, 301–313. [Google Scholar] [CrossRef]
- Ballester, A.-R.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lázaro, C.; Carmona, L.; Wisniewski, M.; Droby, S.; González-Candelas, L.; Gabaldón, T. Genome, transcriptome, and functional analyses of Penicillium expansum provide new insights into secondary metabolism and pathogenicity. Mol. Plant-Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef]
- Hoffmeister, D.; Keller, N.P. Natural products of filamentous fungi: Enzymes, genes, and their regulation. Nat. Prod. Rep. 2007, 24, 393–416. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, J.; Yan, J.; Guo, M.; Xu, L.; Hou, L.; Zou, Q. Comparative genome analysis of plant ascomycete fungal pathogens with different lifestyles reveals distinctive virulence strategies. BMC Genom. 2022, 23, 34. [Google Scholar] [CrossRef]
- Graham-Taylor, C.; Kamphuis, L.G.; Derbyshire, M.C. A detailed in silico analysis of secondary metabolite biosynthesis clusters in the genome of the broad host range plant pathogenic fungus Sclerotinia sclerotiorum. BMC Genom. 2020, 21, 7. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Noar, R.D.; Daub, M.E. Bioinformatics prediction of polyketide synthase gene clusters from Mycosphaerella fijiensis. PLoS ONE 2016, 11, e0158471. [Google Scholar] [CrossRef] [PubMed]
- Skellam, E. Biosynthesis of fungal polyketides by collaborating and trans-acting enzymes. Nat. Prod. Rep. 2022, 39, 754–783. [Google Scholar] [CrossRef]
- Cox, R.J.; Simpson, T.J. Fungal type I polyketide synthases. Methods Enzymol. 2009, 459, 49–78. [Google Scholar]
- Khaldi, N.; Seifuddin, F.T.; Turner, G.; Haft, D.; Nierman, W.C.; Wolfe, K.H.; Fedorova, N.D. SMURF: Genomic mapping of fungal secondary metabolite clusters. Fungal Genet. Biol. 2010, 47, 736–741. [Google Scholar] [CrossRef]
- Wiemann, P.; Willmann, A.; Straeten, M.; Kleigrewe, K.; Beyer, M.; Humpf, H.U.; Tudzynski, B. Biosynthesis of the red pigment bikaverin in Fusarium fujikuroi: Genes, their function and regulation. Mol. Microbiol. 2009, 72, 931–946. [Google Scholar] [CrossRef]
- Yu, J.; Bhatnagar, D.; Cleveland, T.E. Completed sequence of aflatoxin pathway gene cluster in Aspergillus parasiticus. FEBS Lett. 2004, 564, 126–130. [Google Scholar] [CrossRef]
- Cairns, T.; Meyer, V. In silico prediction and characterization of secondary metabolite biosynthetic gene clusters in the wheat pathogen Zymoseptoria tritici. BMC Genom. 2017, 18, 631. [Google Scholar] [CrossRef]
- Klosterman, S.J.; Subbarao, K.V.; Kang, S.; Veronese, P.; Gold, S.E.; Thomma, B.P.; Chen, Z.; Henrissat, B.; Lee, Y.H.; Park, J.; et al. Comparative genomics yields insights into niche adaptation of plant vascular wilt pathogens. PLoS Pathog. 2011, 7, e1002137. [Google Scholar] [CrossRef]
- Shi-Kunne, X.; Faino, L.; van den Berg, G.C.M.; Thomma, B.; Seidl, M.F. Evolution within the fungal genus Verticillium is characterized by chromosomal rearrangement and gene loss. Environ. Microbiol. 2018, 20, 1362–1373. [Google Scholar] [CrossRef]
- Kasson, M.T.; Kasson, L.R.; Wickert, K.L.; Davis, D.D.; Stajich, J.E. Genome sequence of a lethal vascular wilt fungus, Verticillium nonalfalfae, a biological control used against the invasive Ailanthus altissima. Microbiol. Resour. Announc. 2019, 8, e01619-18. [Google Scholar] [CrossRef]
- Harting, R.; Starke, J.; Kusch, H.; Pöggeler, S.; Maurus, I.; Schlüter, R.; Landesfeind, M.; Bulla, I.; Nowrousian, M.; de Jonge, R. A 20-kb lineage-specific genomic region tames virulence in pathogenic amphidiploid Verticillium longisporum. Mol. Plant Pathol. 2021, 22, 939–953. [Google Scholar] [CrossRef]
- Seidl, M.F.; Faino, L.; Shi-Kunne, X.; van den Berg, G.C.; Bolton, M.D.; Thomma, B.P. The genome of the saprophytic fungus Verticillium tricorpus reveals a complex effector repertoire resembling that of its pathogenic relatives. Mol. Plant-Microbe Interact. 2015, 28, 362–373. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; Van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Stanke, M.; Morgenstern, B. AUGUSTUS: A web server for gene prediction in eukaryotes that allows user-defined constraints. Nucleic Acids Res. 2005, 33, W465–W467. [Google Scholar] [CrossRef]
- Zdobnov, E.M.; Apweiler, R. InterProScan—An integration platform for the signature-recognition methods in InterPro. Bioinformatics 2001, 17, 847–848. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2010, 39, D225–D229. [Google Scholar] [CrossRef]
- Bachmann, B.O.; Ravel, J. Methods for in silico prediction of microbial polyketide and nonribosomal peptide biosynthetic pathways from DNA sequence data. Methods Enzymol. 2009, 458, 181–217. [Google Scholar]
- Cox, R.J. Polyketides, proteins and genes in fungi: Programmed nano-machines begin to reveal their secrets. Org. Biomol. Chem. 2007, 5, 2010–2026. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Abascal, F.; Zardoya, R.; Posada, D. ProtTest: Selection of best-fit models of protein evolution. Bioinformatics 2005, 21, 2104–2105. [Google Scholar] [CrossRef]
- Armaleo, D.; Sun, X.; Culberson, C. Insights from the first putative biosynthetic gene cluster for a lichen depside and depsidone. Mycologia 2011, 103, 741–754. [Google Scholar] [CrossRef]
- Feng, P.; Shang, Y.; Cen, K.; Wang, C. Fungal biosynthesis of the bibenzoquinone oosporein to evade insect immunity. Proc. Natl. Acad. Sci. USA 2015, 112, 11365–11370. [Google Scholar] [CrossRef]
- Kudo, F.; Matsuura, Y.; Hayashi, T.; Fukushima, M.; Eguchi, T. Genome mining of the sordarin biosynthetic gene cluster from Sordaria araneosa Cain ATCC 36386: Characterization of cycloaraneosene synthase and GDP-6-deoxyaltrose transferase. J. Antibiot. 2016, 69, 541–548. [Google Scholar] [CrossRef]
- Inderbitzin, P.; Asvarak, T.; Turgeon, B.G. Six new genes required for production of T-toxin, a polyketide determinant of high virulence of Cochliobolus heterostrophus to maize. Mol. Plant-Microbe Interact. 2010, 23, 458–472. [Google Scholar] [CrossRef]
- Brown, D.W.; Lee, S.-H.; Kim, L.-H.; Ryu, J.-G.; Lee, S.; Seo, Y.; Kim, Y.H.; Busman, M.; Yun, S.-H.; Proctor, R.H. Identification of a 12-gene fusaric acid biosynthetic gene cluster in Fusarium species through comparative and functional genomics. Mol. Plant-Microbe Interact. 2015, 28, 319–332. [Google Scholar] [CrossRef]
- Grum-Grzhimaylo, A.A.; Falkoski, D.L.; van den Heuvel, J.; Valero-Jiménez, C.A.; Min, B.; Choi, I.G.; Lipzen, A.; Daum, C.G.; Aanen, D.K.; Tsang, A. The obligate alkalophilic soda-lake fungus Sodiomyces alkalinus has shifted to a protein diet. Mol. Ecol. 2018, 27, 4808–4819. [Google Scholar] [CrossRef]
- Lin, H.-C.; Chooi, Y.-H.; Dhingra, S.; Xu, W.; Calvo, A.M.; Tang, Y. The fumagillin biosynthetic gene cluster in Aspergillus fumigatus encodes a cryptic terpene cyclase involved in the formation of β-trans-bergamotene. J. Am. Chem. Soc. 2013, 135, 4616–4619. [Google Scholar] [CrossRef] [PubMed]
- Morishita, Y.; Zhang, H.; Taniguchi, T.; Mori, K.; Asai, T. The discovery of fungal polyene macrolides via a postgenomic approach reveals a polyketide macrocyclization by trans-acting thioesterase in fungi. Org. Lett. 2019, 21, 4788–4792. [Google Scholar] [CrossRef] [PubMed]
- Hansen, F.T.; Gardiner, D.M.; Lysøe, E.; Fuertes, P.R.; Tudzynski, B.; Wiemann, P.; Sondergaard, T.E.; Giese, H.; Brodersen, D.E.; Sørensen, J.L. An update to polyketide synthase and non-ribosomal synthetase genes and nomenclature in Fusarium. Fungal Genet. Biol. 2015, 75, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Fatema, U.; Broberg, A.; Jensen, D.F.; Karlsson, M.; Dubey, M. Functional analysis of polyketide synthase genes in the biocontrol fungus Clonostachys rosea. Sci. Rep. 2018, 8, 15009. [Google Scholar] [CrossRef]
- Brown, D.W.; Butchko, R.A.; Baker, S.E.; Proctor, R.H. Phylogenomic and functional domain analysis of polyketide synthases in Fusarium. Fungal Biol. 2012, 116, 318–331. [Google Scholar] [CrossRef]
- Wiemann, P.; Sieber, C.M.; Von Bargen, K.W.; Studt, L.; Niehaus, E.-M.; Espino, J.J.; Huss, K.; Michielse, C.B.; Albermann, S.; Wagner, D. Deciphering the cryptic genome: Genome-wide analyses of the rice pathogen Fusarium fujikuroi reveal complex regulation of secondary metabolism and novel metabolites. PLoS Pathog. 2013, 9, e1003475. [Google Scholar] [CrossRef]
- Throckmorton, K.; Wiemann, P.; Keller, N.P. Evolution of chemical diversity in a group of non-reduced polyketide gene clusters: Using phylogenetics to inform the search for novel fungal natural products. Toxins 2015, 7, 3572–3607. [Google Scholar] [CrossRef]
- Campbell, M.A.; Rokas, A.; Slot, J.C. Horizontal transfer and death of a fungal secondary metabolic gene cluster. Genome Biol. Evol. 2012, 4, 289–293. [Google Scholar] [CrossRef]
- Reynolds, H.T.; Slot, J.C.; Divon, H.H.; Lysøe, E.; Proctor, R.H.; Brown, D.W. Differential retention of gene functions in a secondary metabolite cluster. Mol. Biol. Evol. 2017, 34, 2002–2015. [Google Scholar] [CrossRef]
- Lind, A.L.; Wisecaver, J.H.; Lameiras, C.; Wiemann, P.; Palmer, J.M.; Keller, N.P.; Rodrigues, F.; Goldman, G.H.; Rokas, A. Drivers of genetic diversity in secondary metabolic gene clusters within a fungal species. PLoS Biol. 2017, 15, e2003583. [Google Scholar] [CrossRef]
- Tsuji, G.; Kenmochi, Y.; Takano, Y.; Sweigard, J.; Farrall, L.; Furusawa, I.; Horino, O.; Kubo, Y. Novel fungal transcriptional activators, Cmr1p of Colletotrichum lagenarium and Pig1p of Magnaporthe grisea, contain Cys2His2 zinc finger and Zn (II) 2Cys6 binuclear cluster DNA-binding motifs and regulate transcription of melanin biosynthesis genes in a developmentally specific manner. Mol. Microbiol. 2000, 38, 940–954. [Google Scholar]
- Li, H.; Wang, D.; Zhang, D.-D.; Geng, Q.; Li, J.-J.; Sheng, R.-C.; Xue, H.-S.; Zhu, H.; Kong, Z.-Q.; Dai, X.-F. A polyketide synthase from Verticillium dahliae modulates melanin biosynthesis and hyphal growth to promote virulence. BMC Biol. 2022, 20, 125. [Google Scholar] [CrossRef]
- Navarro-Muñoz, J.C.; Collemare, J. Evolutionary histories of type III polyketide synthases in fungi. Front. Microbiol. 2020, 3018. [Google Scholar] [CrossRef]
- Seshime, Y.; Juvvadi, P.R.; Fujii, I.; Kitamoto, K. Discovery of a novel superfamily of type III polyketide synthases in Aspergillus oryzae. Biochem. Biophys. Res. Commun. 2005, 331, 253–260. [Google Scholar] [CrossRef]
- Shimizu, Y.; Ogata, H.; Goto, S. Type III polyketide synthases: Functional classification and phylogenomics. ChemBioChem 2017, 18, 50–65. [Google Scholar] [CrossRef]
- Yan, H.; Sun, L.; Huang, J.; Qiu, Y.; Xu, F.; Yan, R.; Zhu, D.; Wang, W.; Zhan, J. Identification and heterologous reconstitution of a 5-alk (en) ylresorcinol synthase from endophytic fungus Shiraia sp. Slf14. J. Microbiol. 2018, 56, 805–812. [Google Scholar] [CrossRef]
- Sayari, M.; Steenkamp, E.T.; van der Nest, M.A.; Wingfield, B.D. Diversity and evolution of polyketide biosynthesis gene clusters in the Ceratocystidaceae. Fungal Biol. 2018, 122, 856–866. [Google Scholar] [CrossRef]
- Hashimoto, M.; Nonaka, T.; Fujii, I. Fungal type III polyketide synthases. Nat. Prod. Rep. 2014, 31, 1306–1317. [Google Scholar] [CrossRef]
- Skellam, E. The biosynthesis of cytochalasans. Nat. Prod. Rep. 2017, 34, 1252–1263. [Google Scholar] [CrossRef]
- Heard, S.C.; Wu, G.; Winter, J.M. Discovery and characterization of a cytochalasan biosynthetic cluster from the marine-derived fungus Aspergillus flavipes CNL-338. J. Antibiot. 2020, 73, 803–807. [Google Scholar] [CrossRef]
- Qiao, K.; Chooi, Y.-H.; Tang, Y. Identification and engineering of the cytochalasin gene cluster from Aspergillus clavatus NRRL 1. Metab. Eng. 2011, 13, 723–732. [Google Scholar] [CrossRef]
- Wang, C.; Hantke, V.; Cox, R.J.; Skellam, E. Targeted gene inactivations expose silent cytochalasans in Magnaporthe grisea NI980. Org. Lett. 2019, 21, 4163–4167. [Google Scholar] [CrossRef]
- Cain, J.W.; Miller, K.I.; Kalaitzis, J.A.; Chau, R.; Neilan, B.A. Genome mining of a fungal endophyte of Taxus yunnanensis (Chinese yew) leads to the discovery of a novel azaphilone polyketide, lijiquinone. Microb. Biotechnol. 2020, 13, 1415–1427. [Google Scholar] [CrossRef]
- Sato, M.; Winter, J.M.; Kishimoto, S.; Noguchi, H.; Tang, Y.; Watanabe, K. Combinatorial generation of chemical diversity by redox enzymes in chaetoviridin biosynthesis. Org. Lett. 2016, 18, 1446–1449. [Google Scholar] [CrossRef]
- Sieber, C.M.; Lee, W.; Wong, P.; Münsterkötter, M.; Mewes, H.-W.; Schmeitzl, C.; Varga, E.; Berthiller, F.; Adam, G.; Güldener, U. The Fusarium graminearum genome reveals more secondary metabolite gene clusters and hints of horizontal gene transfer. PLoS ONE 2014, 9, e110311. [Google Scholar]
- Droce, A.; Saei, W.; Jørgensen, S.H.; Wimmer, R.; Giese, H.; Wollenberg, R.D.; Sondergaard, T.E.; Sørensen, J.L. Functional analysis of the fusarielin biosynthetic gene cluster. Molecules 2016, 21, 1710. [Google Scholar] [CrossRef]
- Von Bargen, K.W.; Niehaus, E.-M.; Krug, I.; Bergander, K.; Würthwein, E.-U.; Tudzynski, B.; Humpf, H.-U. Isolation and structure elucidation of fujikurins A–D: Products of the PKS19 gene cluster in Fusarium fujikuroi. J. Nat. Prod. 2015, 78, 1809–1815. [Google Scholar] [CrossRef]
- Sbaraini, N.; Andreis, F.C.; Thompson, C.E.; Guedes, R.L.; Junges, Â.; Campos, T.; Staats, C.C.; Vainstein, M.H.; Ribeiro de Vasconcelos, A.T.; Schrank, A. Genome-Wide Analysis of Secondary Metabolite Gene Clusters in O phiostoma ulmi and Ophiostoma novo-ulmi Reveals a Fujikurin-Like Gene Cluster with a Putative Role in Infection. Front. Microbiol. 2017, 8, 1063. [Google Scholar] [CrossRef] [PubMed]
- Kakule, T.B.; Sardar, D.; Lin, Z.; Schmidt, E.W. Two related pyrrolidinedione synthetase loci in Fusarium heterosporum ATCC 74349 produce divergent metabolites. ACS Chem. Biol. 2013, 8, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Dander, J.E.; Sato, C.; Hung, Y.-S.; Gao, S.-S.; Tang, M.-C.; Hang, L.; Winter, J.M.; Garg, N.K.; Watanabe, K. Collaborative biosynthesis of maleimide-and succinimide-containing natural products by fungal polyketide megasynthases. J. Am. Chem. Soc. 2017, 139, 5317–5320. [Google Scholar] [CrossRef] [PubMed]
- Grijseels, S.; Nielsen, J.C.; Randelovic, M.; Nielsen, J.; Nielsen, K.F.; Workman, M.; Frisvad, J.C. Penicillium arizonense, a new, genome sequenced fungal species, reveals a high chemical diversity in secreted metabolites. Sci. Rep. 2016, 6, 35112. [Google Scholar] [CrossRef]
- Li, H.; Gilchrist, C.L.; Lacey, H.J.; Crombie, A.; Vuong, D.; Pitt, J.I.; Lacey, E.; Chooi, Y.-H.; Piggott, A.M. Discovery and heterologous biosynthesis of the burnettramic acids: Rare PKS-NRPS-derived bolaamphiphilic pyrrolizidinediones from an Australian fungus, Aspergillus burnettii. Org. Lett. 2019, 21, 1287–1291. [Google Scholar] [CrossRef]
- Fujii, I.; Yoshida, N.; Shimomaki, S.; Oikawa, H.; Ebizuka, Y. An iterative type I polyketide synthase PKSN catalyzes synthesis of the decaketide alternapyrone with regio-specific octa-methylation. Chem. Biol. 2005, 12, 1301–1309. [Google Scholar] [CrossRef]
- Chen, J.Y.; Liu, C.; Gui, Y.J.; Si, K.W.; Zhang, D.D.; Wang, J.; Short, D.P.; Huang, J.Q.; Li, N.Y.; Liang, Y. Comparative genomics reveals cotton-specific virulence factors in flexible genomic regions in Verticillium dahliae and evidence of horizontal gene transfer from Fusarium. New Phytol. 2018, 217, 756–770. [Google Scholar] [CrossRef]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Li, J.; Cornelissen, B.; Rep, M. Host-specificity factors in plant pathogenic fungi. Fungal Genet. Biol. 2020, 144, 103447. [Google Scholar] [CrossRef]
- De Jonge, R.; Peter van Esse, H.; Maruthachalam, K.; Bolton, M.D.; Santhanam, P.; Saber, M.K.; Zhang, Z.; Usami, T.; Lievens, B.; Subbarao, K.V. Tomato immune receptor Ve1 recognizes effector of multiple fungal pathogens uncovered by genome and RNA sequencing. Proc. Natl. Acad. Sci. USA 2012, 109, 5110–5115. [Google Scholar] [CrossRef]
- Flot, J.-F.; Hespeels, B.; Li, X.; Noel, B.; Arkhipova, I.; Danchin, E.G.; Hejnol, A.; Henrissat, B.; Koszul, R.; Aury, J.-M. Genomic evidence for ameiotic evolution in the bdelloid rotifer Adineta vaga. Nature 2013, 500, 453–457. [Google Scholar] [CrossRef]
- Han, M.V.; Thomas, G.W.; Lugo-Martinez, J.; Hahn, M.W. Estimating gene gain and loss rates in the presence of error in genome assembly and annotation using CAFE 3. Mol. Biol. Evol. 2013, 30, 1987–1997. [Google Scholar] [CrossRef]
- Olson, M.V. When less is more: Gene loss as an engine of evolutionary change. Am. J. Hum. Genet. 1999, 64, 18–23. [Google Scholar] [CrossRef]
- Albalat, R.; Cañestro, C. Evolution by gene loss. Nat. Rev. Genet. 2016, 17, 379–391. [Google Scholar] [CrossRef]
- Brown, D.W.; Villani, A.; Susca, A.; Moretti, A.; Hao, G.; Kim, H.-S.; Proctor, R.H.; McCormick, S.P. Gain and loss of a transcription factor that regulates late trichothecene biosynthetic pathway genes in Fusarium. Fungal Genet. Biol. 2020, 136, 103317. [Google Scholar] [CrossRef]
- De Jonge, R.; Bolton, M.D.; Kombrink, A.; van den Berg, G.C.; Yadeta, K.A.; Thomma, B.P. Extensive chromosomal reshuffling drives evolution of virulence in an asexual pathogen. Genome Res. 2013, 23, 1271–1282. [Google Scholar] [CrossRef]
- Faino, L.; Seidl, M.F.; Shi-Kunne, X.; Pauper, M.; van den Berg, G.C.; Wittenberg, A.H.; Thomma, B.P. Transposons passively and actively contribute to evolution of the two-speed genome of a fungal pathogen. Genome Res. 2016, 26, 1091–1100. [Google Scholar] [CrossRef]
- Boettger, D.; Hertweck, C. Molecular diversity sculpted by fungal PKS–NRPS hybrids. ChemBioChem 2013, 14, 28–42. [Google Scholar] [CrossRef]
- Xiong, D.; Wang, Y.; Tian, C. A novel gene from a secondary metabolism gene cluster is required for microsclerotia formation and virulence in Verticillium dahliae. Phytopathol. Res. 2019, 1, 31. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isolate Number | GenBank Accession Number | References |
|---|---|---|---|
| V. dahliae | VdLs.17 | ABJE00000000 | [42] |
| V. albo-atrum | PD747 | NMXJ00000000 | [43] |
| V. isaacii | PD618 | NMXN00000000 | [43] |
| V.alfalfae | VaMs.102 | ABPE00000000 | [42] |
| V. nonalfalfae | VnAa140/PSU140/NRRL 66861 | RBVV00000000 | [44] |
| V. nubilum | PD621 | NMXI00000000 | [43] |
| V. zaregamsianum | PD739 | NMXM00000000 | [43] |
| V. longisporum | VL1 | JAETXT000000000 | [45] |
| V. klebahnii | PD401 | NMXL00000000 | [43] |
| V. tricorpus | MUCL 9792 | JPET00000000 | [46] |
| Verticillium NR-PKS-I Used as a Query | Size | Domain Organization * | Information Regarding the Top BLASTp Hit | |||
|---|---|---|---|---|---|---|
| Accession Number | Species | Size (aa) | Domain Organization * | |||
| VNR-PKS-I-1 | ||||||
| All Verticillium genomes | 2164–2189 | KS-AT-ACP-ACP-TE | XP_038751444 | Colletotrichum karsti | 2179 | KS-AT-ACP-ACP-TE |
| VNR-PKS-I-2 | ||||||
| V. zaregamsianum and V. klebahnii | 2124–2126 | KS-AT-ACP-ACP-TE | RYP01440 | Monosporascus ibericus | 2108 | KS-AT-DH-ACP-ACP-TE |
| VNR-PKS-I-3 | ||||||
| V. alfalfa, V. nonalfalfae and V. longisporum | 1092–1911 | KS-AT-ACP-ACP-TE | XP_028463040 | Sodiomyces alkalinus | 1919 | KS-AT-ACP-ACP-TE |
| V. longisporum | 1259 | KS-AT-ACP-ACP-TE-MFS | KND89106 | Tolypocladium ophioglossoides | 2147 | KS-AT-ACP-TE |
| V. dahliae | 2158 | KS-AT-ACP-ACP-TE-MFS | XP_028463040 | Sodiomyces alkalinus | 1919 | KS-AT-ACP-ACP-TE |
| VNR-PKS-I-4 | ||||||
| V. nubilum | 2578 | KS-AT-ACP-MT-NAD binding 4 | KAH7133939 | Dactylonectria macrodidyma | 2280 | KS-AT-ACP-MT |
| Verticillium R-PKS-I Used as a Query | Size | Domain Organization * | Information Regarding the Top BLASTp Hit | |||
|---|---|---|---|---|---|---|
| Accession | Name | Size (aa) | Domain Organization * | |||
| VRPKS-I-1 (PKS-NRPS) | ||||||
| V. dahliae, V. zaregamsianum, V. alfalfae and V. longisporum | 3750–4123 | KS-AT-DH-ER-KR-C-A-ACP | KAF2874833 | Massariosphaeria phaeospora | 3929 | KS-AT-DH-MET-ER-KR-C-A-ACP |
| VRPKS-I-2 (PKS-NRPS) | ||||||
| V. isaacii, V. klebahnii, V. tricorpus and V. albo-atrum | 3800–4429 | KS-AT-DH-ER-KR-ACP-C-A-ACP | KAF4886526 | Colletotrichum fructicola | 3989 | KS-AT-DH-ER-KR-ACP-C-A-ACP |
| VRPKS-I-3 (PKS-NRPS) | ||||||
| V. albo-atrum | 4677 | KS-AT-DH-ER-KR-C-A-ACP-NN * | XP_036576988 | Colletotrichum truncatum | 4127 | KS-AT-DH-ER-KR-C-A-ACP |
| VRPKS-I-4 (PKS-NRPS) | ||||||
| V. nubilum | 3987 | KS-AT-DH-MET-ER-KR- C-A-ACP | KAI1485991 | Biscogniauxia mediterranea | 3976 | KS-AT-DH-MET-ER-KR-C-A-ACP |
| VRPKS-I-5 (PKS-NRPS) | ||||||
| V. albo-atrum, V. zaregamsianum, V. isaacii and V. klebahnii | 3912–4017 | KS-AT-DH-MET-ER-KR-C-A-ACP | TGO20046 | Botrytis tulipae | 4020 | KS-AT-DH-ER-KR-C-A-ACP |
| V. tricorpus | 4131 | KS-AT-DH-MET-ER-KR-C-A-ACP-NAD_binding_4 | KAI1485991 | Biscogniauxia mediterranea | 3976 | KS-AT-DH-MET-ER-KR-C-A-ACP |
| VRPKS-I-6 (PKS-NRPS) | ||||||
| V. tricorpus, V. zaregamsianum, V. isaacii and V. klebahnii | 4028 | KS-AT-DH-ER-KR-ACP-C-A-ACP | KAE8150730 | Aspergillus avenaceus | 4038 | KS-AT-DH-MET-ER-KR-ACP-C-A-ACP |
| VRPKS-I-7 | ||||||
| V. zaregamsianum | 2690 | KS-AT-DH-MET-ER-KR | KAH8169122 | Sarocladium implicatum | 2675 | KS-AT-DH-MET-ER-KR |
| VRPKS-I-8 | ||||||
| V. dahliae and V. longisporum | 2506–2574 | KS-AT-DH-ER-KR | ALQ32784 | Fusarium babinda | 2460 | KS-AT-DH-ER-KR |
| VRPKS-I-9 (PKS-NRPS) | ||||||
| V. alfalfae, V. nonalfalfae, V. albo-atrum, V. tricorpus, V. isaacii and V. klebahnii | 3644–4064 | KS-AT-DH-ER-KR-ACP-C-A-ACP | XP_044717051 | Hirsutella rhossilie | 3845 | KS-AT-DH-ER-KR-ACP-C-A-ACP |
| VRPKS-I-10 | ||||||
| V. nubilum | 1703 | KS-AT-KR-ACP | KAH8906691 | Coniochaeta sp. | 2576 | KS-AT-DH-ER-KR |
| VRPKS-I-11(PKS-NRPS) | ||||||
| V. alfalfae | 3529 | KS-AT-DH-ER-KR-C-A-ACP | KAB2573439 | Lasiodiplodia theobromae | 3721 | KS-AT-DH-ER-KR-ACP-C-A-ACP |
| V. dahliae | 1113 | KS-AT-DH | KAF4511962 | Ophiocordyceps sinensis | 1029 | KS-AT-DH |
| V. nonalfalfae | 3858 | KS-AT-DH-ER-KR-ACP-C-A-ACP | KFH45364 | Acremonium chrysogenum | 3926 | KS-AT-DH-ER-KR-ACP-C-A-ACP |
| V. longisporum contig | 2672 | KS-AT-DH-ER-KR-ACP-C | KFH45364 | Acremonium chrysogenum | 3926 | KS-AT-DH-ER-KR-ACP-C-A-ACP |
| VRPKS-I-12 (PKS-NRPS) | ||||||
| V. isaacii, V. klebahnii and V. tricorpus | 3943–3982 | KS-AT-DH-MET-ER-KR-ACP- C-A-ACP-NAD_binding_4 | XP_028469272 | Sodiomyces alkalinus | 3890 | KS-AT-DH-MET-ER-KR-ACP- C-A-ACP- |
| V. zaregamsianum and V. albo-atrum | 3891–3972 | KS-AT-DH-MET-ER-KR-ACP-C-A-ACP | XP_028469272 | Sodiomyces alkalinus | 3890 | KS-AT-DH-MET-ER-KR-ACP-C-A-ACP |
| VRPKS-I-13 | ||||||
| V. isaacii and V. klebahnii | 3080–3128 | KS-AT-DH-ER-KR-ACP | KIL85244 | Fusarium avenaceum | 3197 | KS-AT-DH-MET-ER-KR-ACP |
| V. tricorpus and V. zaregamsianum | 3176–3209 | KS-AT-DH-MET-ER-KR-ACP | KIL85244 | Fusarium avenaceum | 3197 | KS-AT-DH-MET-ER-KR-ACP |
| VRPKS-I-14 | ||||||
| V. zaregamsianum, V. tricorpus, V. isaacii and V. klebahnii | 2898–3161 | KS-AT-DH-MET-ER-KR-ACP | XP_045957259 | Truncatella angustata | 3173 | KS-AT-DH-MET-ER-KR-ACP |
| VRPKS-I-15 | ||||||
| V. nubilum contig | 2690 | KS-AT-DH-MET-ER-KR | KAH7133936 | Dactylonectria macrodidyma | 2606 | KS-AT-DH-MET-ER-KR-ACP |
| VRPKS-I-16 | ||||||
| V. isaacii, V. albo-atrum, V. zaregamsianum, V. tricorpus, and V. klebahnii | 2575–2718 | KS-AT-DH-MET-ER-KR-ACP | KAH7369281 | Plectosphaerella cucumerina | 2570 | KS-AT-DH-MET-ER-KR-ACP |
| VRPKS-I-17 | ||||||
| V. zaregamsianum, V. albo-atrum, V. isaacii, V. alfalfae, V. nonalfalfae, V. tricorpus and V. klebahnii | 2158–2468 | KS-AT-DH-ER-KR | OLN86478 | Colletotrichum chlorophyti | 2290 | KS-AT-DH-ER-KR |
| V. nubilum, V. dahliae and V. longisporum | 2268–2296 | KS-AT-DH-ER-KR-ACP | OLN86478 | Colletotrichum chlorophyti | 2290 | KS-AT-DH-ER-KR |
| VRPKS-I-18 | ||||||
| V. isaacii | 2256 | KS-AT-DH-ER-KR | POR36838 | Tolypocladium paradoxum | 2322 | KS-AT-DH-ER-KR |
| V. klebahnii and V. zaregamsianum | 2565–2857 | AMP-KS-AT-DH-ER-KR | UJO18003 | Fulvia fulva | 2256 | KS-AT-DH-ER-KR |
| VRPKS-I-19 | ||||||
| V. longisporum, V. dahliae, V. zaregamsianum, Verticillium albo-atrum, V. klebahnii, V. isaacii, V. nonalfalfae, V. tricorpus, and V. longisporum | 2108–2291 | KS-AT-DH-ER-KR-ACP | KAG6264852 | Claviceps purpurea | 2196 | KS-AT-DH-ER-KR |
| V. alfalfae and V. nubilum | 1524–2174 | KS-AT-DH-ER-KR | KAG6041353 | Epichloe festucae | 2081 | KS-AT-DH-ER-KR |
| VRPKS-I-20 | ||||||
| V. isaacii contig | 3430 | KS-AT-DH-ER-KR-ACP-CYP120A1-CYPOR | KAF4594413 | Ophiocordyceps camponoti-floridani | 2547 | KS-AT-DH-MET-ER-KR |
| V. klebahnii contig | 2354 | KS-AT-DH-ER-KR-ACP | XP_022486633 | Penicillium arizonense | 2589 | KS-AT-DH-MET-ER-KR-ACP |
| V. tricorpus contig | 2475 | KS-AT-DH-MET-ER-KR-ACP | XP_022486633 | Penicillium arizonense | 2589 | KS-AT-DH-MET-ER-KR-ACP |
| VRPKS-I-21 | ||||||
| V. zaregamsianum, V. klebahnii and V. nubilum | 2331–2532 | KS-AT-DH-MET-ER-KR | KAF6826616 | Colletotrichum plurivorum | 2541 | KS-AT-DH-MET-ER-KR-ACP |
| VRPKS-I-22 | ||||||
| V. dahliae, V. alfalfae, V. nonalfalfae and V. longisporum | 2554–2635 | KS-AT-DH-MET-ER-KR-ACP | ALQ32877 | Fusarium miscanthi | 2644 | KS-AT-DH-MET-ER-KR-ACP |
| VRPKS-I-23 | ||||||
| V. dahliae, V. longisporum, V. alfalfae, V. nonalfalfae, V. klebahnii, V. zaregamsianum and V. tricorpus | 2291–2402 | KS-AT-DH-ER-KR-ACP | XP_028462492 | Sodiomyces alkalinus | 2401 | KS-AT-DH-ER-KR |
| V. nubilum | 2376 | KS-AT-DH-ER-KR | XP_028462492 | Sodiomyces alkalinus | 2401 | KS-AT-DH-ER-KR |
| V. albo-atrum contig | 2651 | KS-AT-DH-MET-ER-KR | XP_028462492 | Sodiomyces alkalinus | 2401 | KS-AT-DH-ER-KR |
| V. isaacii contig | 2347 | KS-AT-DH-ER-KR | XP_028462492 | Sodiomyces alkalinus | 2401 | KS-AT-DH-ER-KR |
| VRPKS-I-24 | ||||||
| V. albo-atrum contig | 2453 | KS-AT-DH-MET-ER-KR-ACP | KAF7552999 | Cylindrodendrum hubeiense | 2560 | KS-AT-DH-MET-ER-KR |
| VRPKS-I-25 | ||||||
| V. klebahnii, V. isaacii, V. zaregamsianum, V. albo-atrum and V. tricorpus | 2103–2358 | KS-AT-DH-ER-KR-ACP | KFA46917 | Stachybotrys chartarum | 2393 | KS-AT-DH-ER-KR |
| VRPKS-I-26 | ||||||
| V. dahliae, V. longisporum, V. alfalfae, V. nonalfalfae, | 1748–2345 | KS-AT-DH-KR-ACP | RYP50633 | Monosporascus sp. | 2358 | KS-AT-DH-MET-ER-KR-ACP |
| Verticillium albo-atrum | 2326 | KS-AT-DH-KR-TTL | RYP50633 | Monosporascus sp. mg162 | 2425 | KS-AT-DH-MET-ER-KR-ACP |
| V. zaregamsianum | 2349 | KS-AT-DH-ER-KR-ACP | P0CU84 | Apiospora sphaerosperma | 2466 | KS-AT-DH-ER-KR-ACP |
| V. isaacii, V. nubilum, V. klebahnii and V. tricorpus | 2345–2350 | KS-AT-DH-ER-KR | P0CU84 | Apiospora sphaerosperma | 2466 | KS-AT-DH-ER-KR-ACP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayari, M.; Dolatabadian, A.; El-Shetehy, M.; Rehal, P.K.; Daayf, F. Genome-Based Analysis of Verticillium Polyketide Synthase Gene Clusters. Biology 2022, 11, 1252. https://doi.org/10.3390/biology11091252
Sayari M, Dolatabadian A, El-Shetehy M, Rehal PK, Daayf F. Genome-Based Analysis of Verticillium Polyketide Synthase Gene Clusters. Biology. 2022; 11(9):1252. https://doi.org/10.3390/biology11091252
Chicago/Turabian StyleSayari, Mohammad, Aria Dolatabadian, Mohamed El-Shetehy, Pawanpuneet Kaur Rehal, and Fouad Daayf. 2022. "Genome-Based Analysis of Verticillium Polyketide Synthase Gene Clusters" Biology 11, no. 9: 1252. https://doi.org/10.3390/biology11091252
APA StyleSayari, M., Dolatabadian, A., El-Shetehy, M., Rehal, P. K., & Daayf, F. (2022). Genome-Based Analysis of Verticillium Polyketide Synthase Gene Clusters. Biology, 11(9), 1252. https://doi.org/10.3390/biology11091252