
Evaluating the Phenotypic and Genomic Characterization of Some Egyptian Phages Infecting Shiga Toxin-Producing Escherichia coli O157:H7 for the Prospective Application in Food Bio-Preservation


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain
2.2. Water Samples Collection and Preparation
2.3. Phage Isolation and Purification
2.4. Phenotypic Phage Characterization
2.4.1. Morphological Characterization
2.4.2. Determination of Structural Proteins
2.4.3. Host Range Analysis
2.4.4. Determination of Multiplicity of Infection (MOI)
2.4.5. Determination of Lytic Activity
2.4.6. UV Radiation Stability
2.4.7. Thermal Stability
2.4.8. pH Stability
2.4.9. Bile Salt Stability
2.4.10. Simulated Gastric Fluid (SGF) Stability
2.4.11. Simulated Intestinal Fluid (SIF) Stability
2.5. Genome Sequencing and Analysis
2.6. Phage Genome Annotation
2.7. Comparative Genomics
2.8. Phylogenetic Analysis
2.9. Statistical Analysis
3. Results
3.1. Isolation of Enterohaemorrhagic E. coli O 157:H7 Infecting Phages
3.2. Phage Morphology
3.3. Phage Structural Proteins
3.4. Host Range Assay
3.5. Determination of MOI and Lytic Activity
3.6. Effect of UV Radiation on Phage Stability
3.7. Thermal Stability
3.8. pH Stability
3.9. Phage Stability in Bile Salt
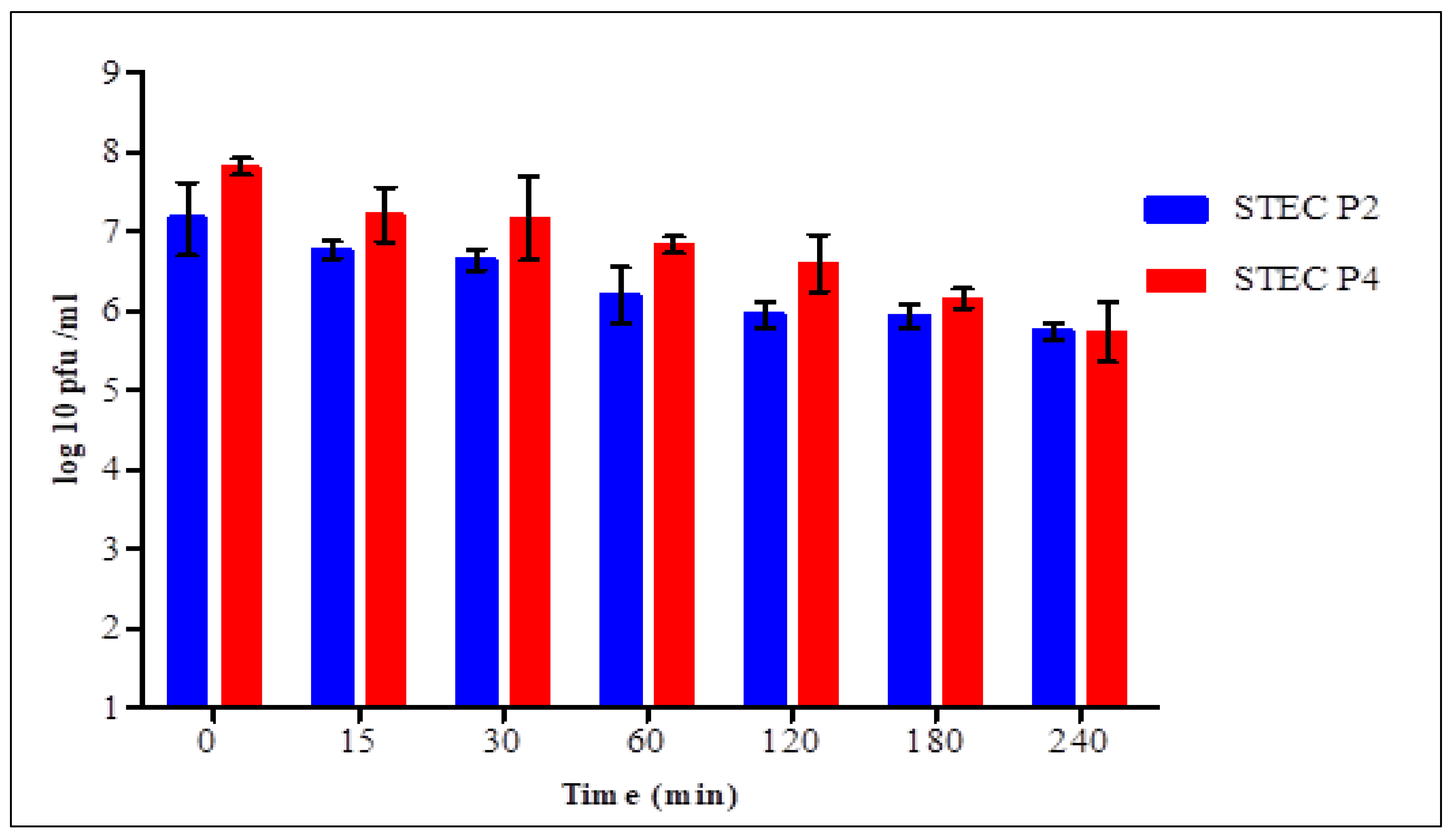
3.10. Phage Stability in Simulated Gastric (SGF) and Intestinal Fluids (SIF)
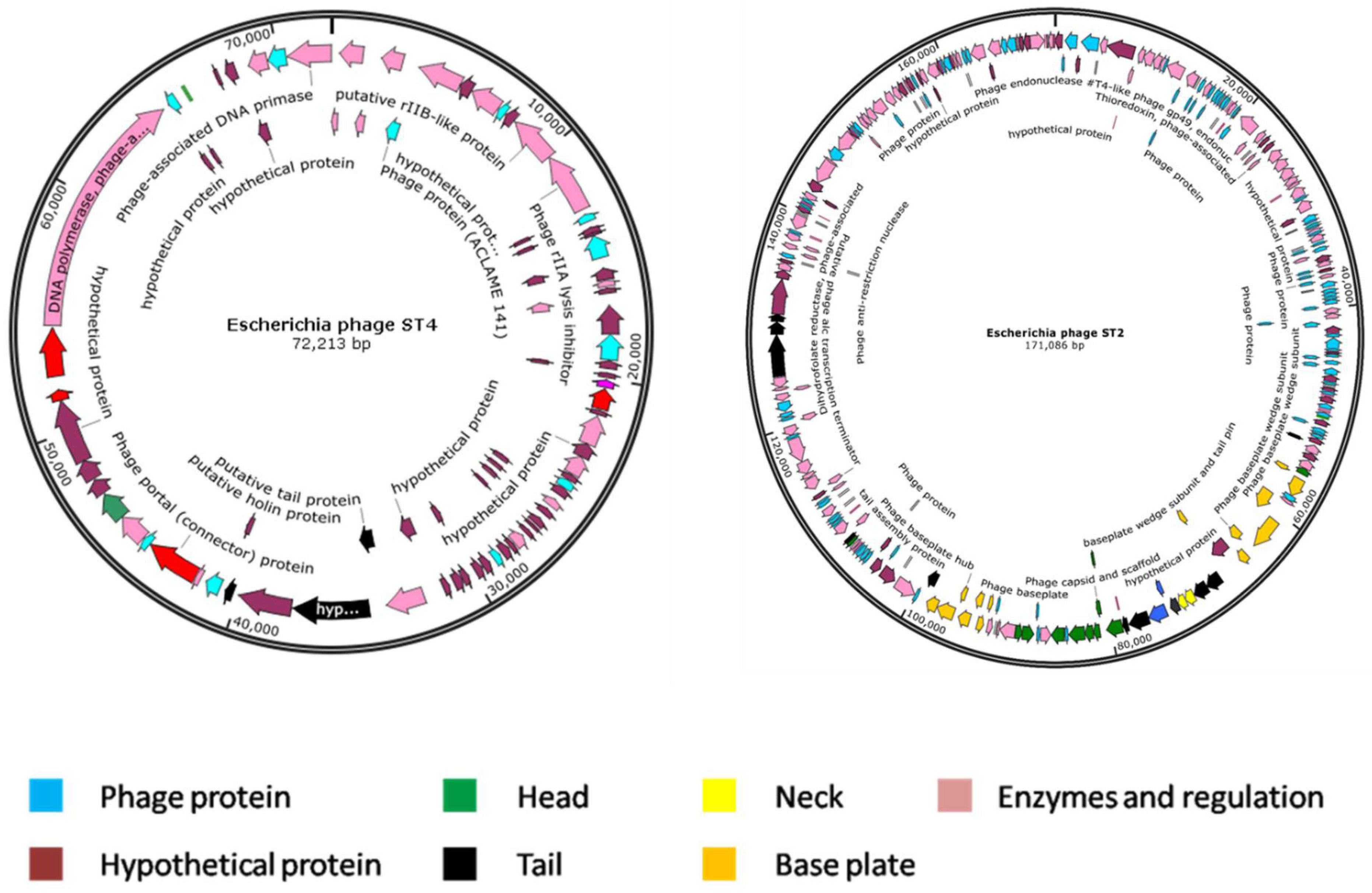
3.11. Genome Characterization
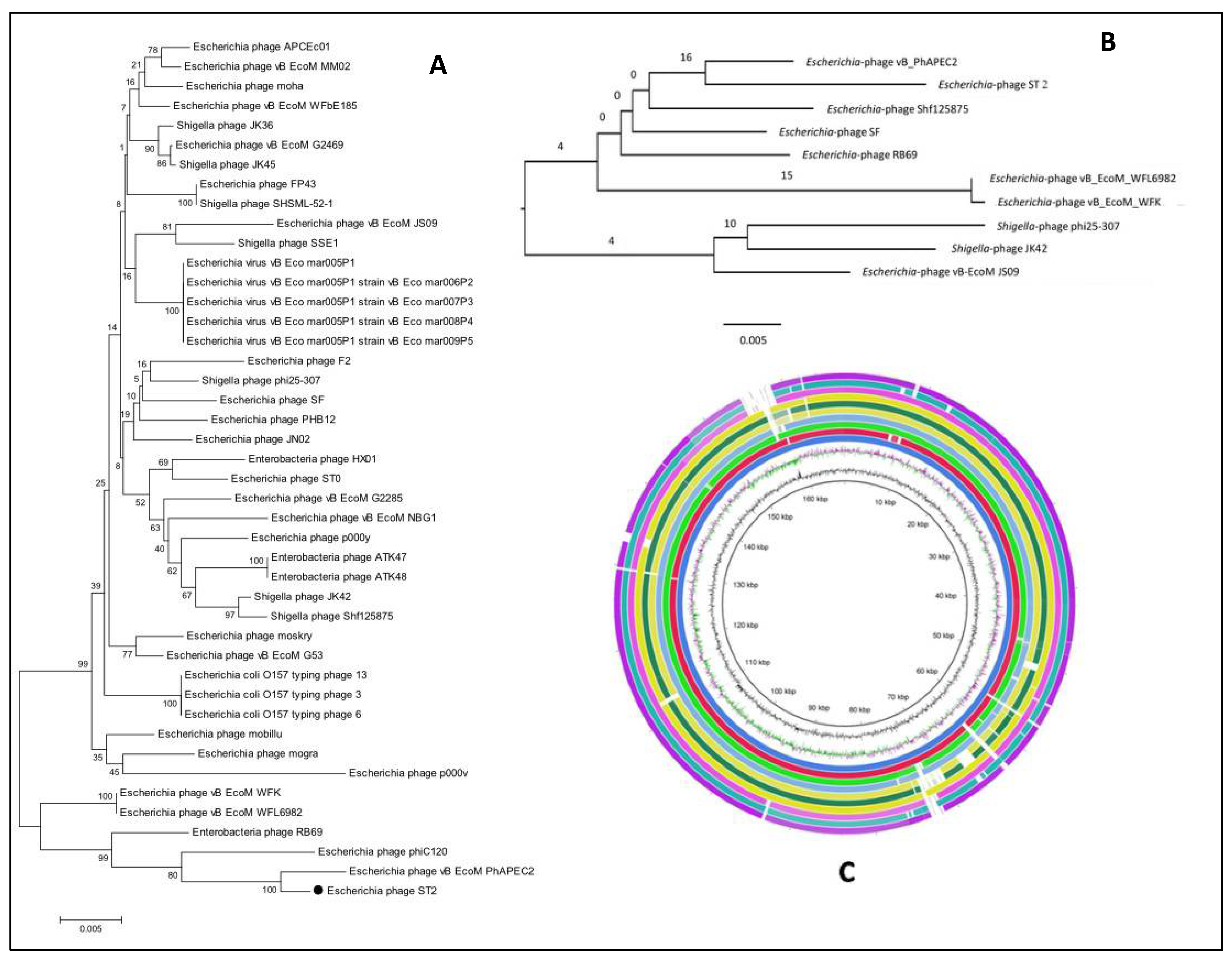
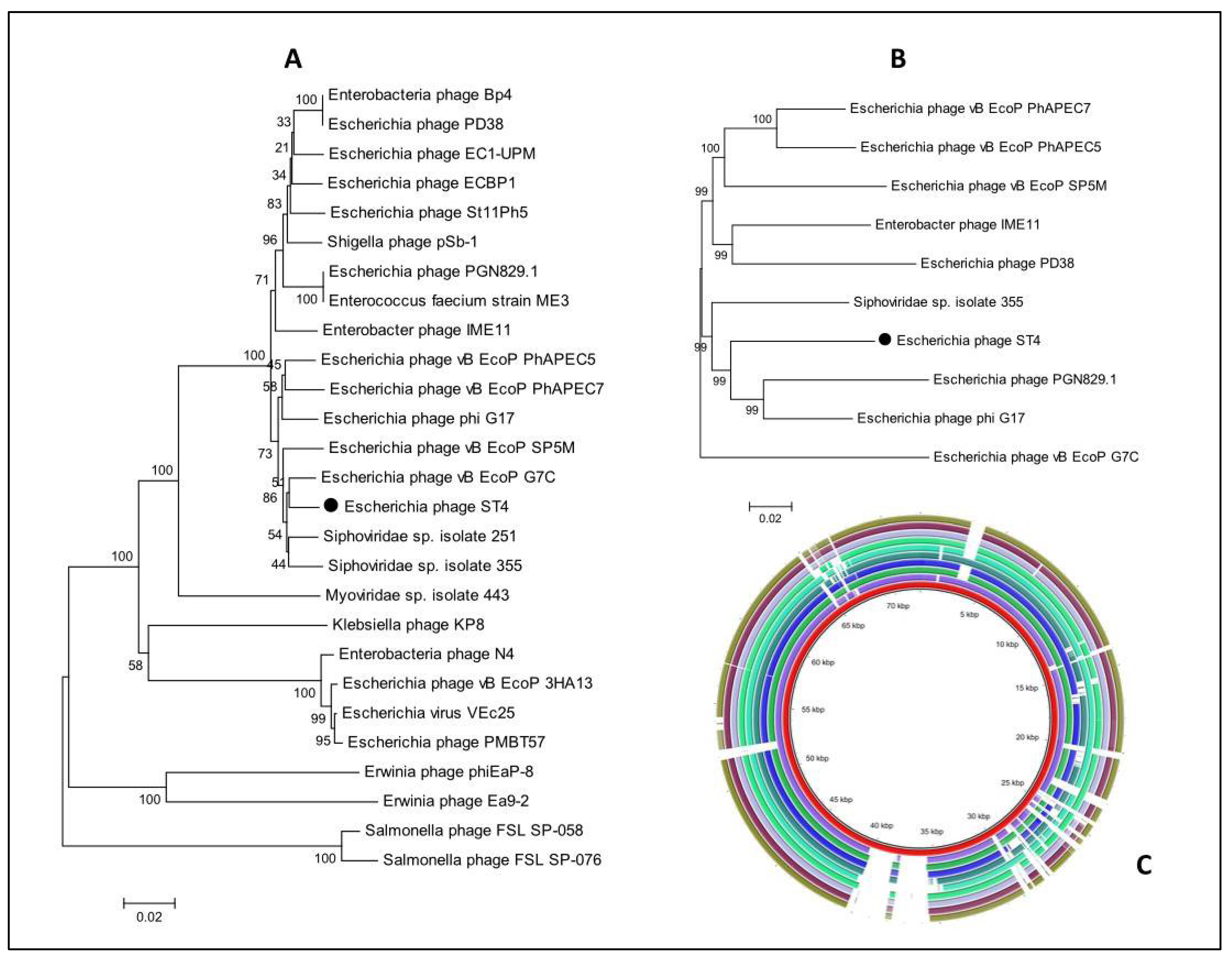
3.12. Comparative Genome Analysis
3.13. Phylogenetic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rabinowitz, R.P.; Donnenberg, M.S. Enteric Infections and Immunity; Paradise, L.J., Bendinelli, M., Friedman, H., Eds.; Springer USA: Boston, MA, USA, 1996; pp. 101–131. [Google Scholar]
- Etcheverria, A.I.; Padola, N.L. Shiga toxin-producing Escherichia coli: Factors involved in virulence and cattle colonization. Virulence 2013, 4, 366–372. [Google Scholar] [CrossRef]
- Rahal, E.A.; Fadlallah, S.M.; Nassar, F.; Kazzi, N.; Matar, G.M. Approaches to treatment of emerging Shiga toxin-producing Escherichia coli infections highlighting the O104:H4 serotype. Front. Cell Infect. Microbiol. 2015, 5, 24. [Google Scholar] [CrossRef] [PubMed]
- Howie, H.; Mukerjee, A.; Cowden, J.; Leith, J.; Reid, T. Investigation of an outbreak of Escherichia coli O157 infection caused by environmental exposure at a scout camp. Epidemiol. Infect. 2003, 131, 1063–1069. [Google Scholar] [CrossRef]
- Caprioli, A.; Morabito, S.; Oswald, E. Enterohaemorrhagic Escherichia coli: Emerging issues on virulence and modes of transmission. Vet. Res. 2005, 36, 289–311. [Google Scholar] [CrossRef]
- Rice, D.H.; Sheng, H.Q.; Wynia, S.A.; Hovde, C.J. Rectoanal mucosal swab culture is more sensitive than fecal culture and distinguishes Escherichia coli O157:H7-colonized cattle and those transiently shedding the same organism. J. Clin. Microbiol. 2003, 41, 4924–4929. [Google Scholar] [CrossRef]
- Álvarez-Fernández, E.; Amaya, C.; Carmen, D.-V.; Rosa, C.; Carlos, A.-C. Antimicrobial resistance in E. coli isolates from conventionally and organically reared poultry: A comparison of agar disc diffusion and Sensi Test Gram-negative methods. Food Control 2013, 30, 227–234. [Google Scholar] [CrossRef]
- Ramirez, K.; Cazarez-Montoya, C.; Lopez-Moreno, H.S.; Campo, N.C.-D. Bacteriophage cocktail for biocontrol of Escherichia coli O157:H7: Stability and potential allergenicity study. PLoS ONE 2018, 13, e0195023. [Google Scholar] [CrossRef] [PubMed]
- Hagens, S.; Loessner, M.J. Bacteriophage for biocontrol of foodborne pathogens: Calculations and considerations. Curr. Pharm. Biotechnol. 2010, 11, 58–68. [Google Scholar] [CrossRef]
- Coffey, B.; Mills, S.; Coffey, A.; McAuliffe, O.; Ross, R.P. Phage and their lysins as biocontrol agents for food safety applications. Annu. Rev. Food Sci. Technol. 2010, 1, 449–468. [Google Scholar] [CrossRef]
- Guenther, S.; Huwyler, D.; Richard, S.; Loessner, M.J. Virulent bacteriophage for efficient biocontrol of Listeria monocytogenes in ready-to-eat foods. Appl. Environ. Microbiol. 2009, 75, 93–100. [Google Scholar] [CrossRef]
- Abuladze, T.; Li, M.; Menetrez, M.Y.; Dean, T.; Senecal, A.; Sulakvelidze, A. Bacteriophages reduce experimental contamination of hard surfaces, tomato, spinach, broccoli, and ground beef by Escherichia coli O157:H7. Appl. Environ. Microbiol. 2008, 74, 6230–6238. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.D.; Parks, A.; Abuladze, T.; Li, M.; Woolston, J.; Magnone, J.; Senecal, A.; Kropinski, A.M.; Sulakvelidze, A. Bacteriophage cocktail significantly reduces Escherichia coli O157:H7 contamination of lettuce and beef, but does not protect against recontamination. Bacteriophage 2012, 2, 178–185. [Google Scholar] [CrossRef]
- Sharma, M.; Patel, J.R.; Conway, W.S.; Ferguson, S.; Sulakvelidze, A. Effectiveness of bacteriophages in reducing Escherichia coli O157:H7 on fresh-cut cantaloupes and lettucet. J. Food Prot. 2009, 72, 1481–1485. [Google Scholar] [CrossRef]
- Ferguson, S.; Roberts, C.; Handy, E.; Sharma, M. Lytic bacteriophages reduce Escherichia coli O157: H7 on fresh cut lettuce introduced through cross-contamination. Bacteriophage 2013, 3, e24323. [Google Scholar] [CrossRef] [PubMed]
- Mahony, J.; McAuliffe, O.; Ross, R.P.; van Sinderen, D. Bacteriophages as biocontrol agents of food pathogens. Curr. Opin. Biotechnol. 2011, 22, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Gaynor, P. GRAS Notice-Exemption Claim for Escherichia coli-specific Phage Preparation (GRAS Notice (GRN) No. 724); F.U.S.F.A.D. Adminsteration, Ed.; Office of Food Additive Safety (HFS-200), Center for Food Safety and Applied Nutrition, Food and Drug Administration 5100 Paint Branch Parkway: College Park, MD, USA, 2017; pp. 20740–23835.
- ISO-TS-13136, ISO/TS 13136:2012; Microbiology of Food and Animal Feed—Real-Time Polymerase Chain Reaction (PCR)-Based Method for the Detection of Food-Borne Pathogens—Horizontal Method for the Detection of Shiga Toxin-Producing Escherichia coli (STEC) and the Determination of O157, O111, O26, O103 and O145 Serogroups, in ICS > 07 > 07.100 > 07.100.30. ISO. International Organization for Standardization: Geneva, Switzerland, 2012; p. 22.
- Mei, G.Y.; Tang, J.; Bach, S.; Kostrzynska, M. Changes in Gene Transcription Induced by Hydrogen Peroxide Treatment of Verotoxin-Producing Escherichia coli O157:H7 and Non-O157 Serotypes on Romaine Lettuce. Front. Microbiol. 2017, 8, 477. [Google Scholar] [CrossRef]
- Adams, M.H. Bacteriophages; Interscience Publishers: New York, NY, USA, 1959. [Google Scholar]
- Ji, X.; Zhang, C.; Fang, Y.; Zhang, Q.; Lin, L.; Tang, B.; Wei, Y. Isolation and characterization of glacier VMY22, a novel lytic cold-active bacteriophage of Bacillus cereus. Virol. Sin. 2015, 30, 52–58. [Google Scholar] [CrossRef]
- Lu, Z.; Breidt, F. Escherichia coli O157:H7 bacteriophage Phi241 isolated from an industrial cucumber fermentation at high acidity and salinity. Front. Microbiol. 2015, 6, 67. [Google Scholar] [CrossRef]
- Kumar, H.; Rangrez, A.Y.; Dayananda, K.M.; Atre, A.N.; Patole, M.S.; Shouche, Y.S. Lactobacillus plantarum (VR1) isolated from an ayurvedic medicine (Kutajarista) ameliorates in vitro cellular damage caused by Aeromonas veronii. BMC Microbiol. 2011, 11, 152. [Google Scholar] [CrossRef]
- Wang, J.; Yadav, V.; Smart, A.L.; Tajiri, S.; Basit, A.W. Toward oral delivery of biopharmaceuticals: An assessment of the gastrointestinal stability of 17 peptide drugs. Mol. Pharm. 2015, 12, 966–973. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. VICTOR: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.O.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical methods, 8th ed.; Iowa State University Press: Ames, IA, USA, 1994. [Google Scholar]
- Tsonos, J.; Oosterik, L.H.; Huruma, N.; Tuntufye, H.N.; Klumpp, J.; Butaye, P.; Greve, H.D.; Hernalsteens, J.-P.; Lavigne, R.; Goddeeris, B.M. A cocktail of in vitro efficient phages is not a guarantee for in vivo therapeutic results against avian colibacillosis. Vet. Microbiol. 2014, 171, 470–479. [Google Scholar] [CrossRef]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Piérard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [PubMed]
- El-Leithy, M.E.-S.; El-Senousy, W.; Abou-Zeid, M.; El-Taweel, G. Detection of Six E. coli O157 virulence genes in water samples using multiplex PCR. Egypt. J. Microbiol. 2012, 47, 171–188. [Google Scholar]
- Selim, S.A.; Ahmed, S.F.; Aziz, M.H.A.; Zakaria, A.M.; Klena, J.D.; Pangallo, D. Prevalence and characterization of Shiga-toxin O157:H7 and Non-O157:H7 enterohemorrhagic Escherichia coli isolated from different sources. Biotechnol. Biotechnol. Equip. 2013, 27, 3834–3842. [Google Scholar] [CrossRef]
- Hamed, O.M.; Sabry, M.A.; Hassanain, N.A.; Hamza, E.; Hegazi, A.G.; Salman, M.B. Occurrence of virulent and antibiotic-resistant Shiga toxin-producing Escherichia coli in some food products and human stool in Egypt. Vet. World 2017, 10, 1233–1240. [Google Scholar] [CrossRef]
- Abd El Gany, S.S.H.; Mahmoud Gad, G.F.; Mousa, S.O.; Ibrahem, R.A. Characterization of verotoxigenic E. coli and enteropathogenic E. coli isolated from infants with diarrhea in combination with antimicrobial resistance pattern in Minia, Egypt. J. Adv. Biomed. Pharm. Sci. 2020, 3, 101–109. [Google Scholar]
- Elmonir, W.; Shalaan, S.; Tahoun, A.; Mahmoud, S.F.; Remela, E.M.A.; Eissa, R.; El-Sharkawy, H.; Shukry, M.; Zahran, R.N. Prevalence, antimicrobial resistance, and genotyping of Shiga toxin-producing Escherichia coli in foods of cattle origin, diarrheic cattle, and diarrheic humans in Egypt. Gut. Pathog. 2021, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Raya, R.L.R.; H’bert, E.M. Isolation of Phage via Induction of Lysogens. In Bacteriophages: Methods and Protocols; Isolation, Characterization, and Interactions; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: Totowa, NJ, USA, 2009; Volume 1, pp. 23–32. [Google Scholar]
- Dubey, K.C.S.; Sao, S.; Gupta, A. Isolation and characterization of virulent coliphages from sewage sample. Int. J. Curr. Microbiol. App. Sci. 2015, 4, 6. [Google Scholar]
- Rodríguez-Rubio, L.; Haarmann, N.; Schwidder, M.; Muniesa, M.; Schmidt, H. Bacteriophages of Shiga toxin-producing Escherichia coli and their contribution to pathogenicity. Pathogens 2021, 10, 404. [Google Scholar] [CrossRef]
- Hudson, J.; Billington, C.; Cornelius, A.; Wilson, T.; On, S.; Premaratne, A.; King, N. Use of a bacteriophage to inactivate Escherichia coli O157:H7 on beef. Food Microbiol. 2013, 36, 14–21. [Google Scholar] [CrossRef]
- Jamal, M.; Hussain, T.; Das, C.R.; Andleeb, S. Isolation and characterization of a Myoviridae MJ1 bacteriophage against multi-drug resistant Escherichia coli 3. Jundishapur. J. Microbiol. 2015, 8, e25917. [Google Scholar] [CrossRef]
- Smith, D.L.; Rooks, D.J.; Fogg, P.C.; Darby, A.C.; Thomson, N.R.; McCarthy, A.J.; E Allison, H. Comparative genomics of Shiga toxin encoding bacteriophages. BMC Genom. 2012, 13, 311. [Google Scholar] [CrossRef]
- Muniesa, M.; Schmidt, H. Shiga toxin-encoding phages: Multifunctional gene ferries. In Pathogenic Escherichia coli: Molecular and Cellular Microbiology; Morabito, S., Ed.; Caister Academic Press: Wymondham, UK, 2014; pp. 57–77. [Google Scholar]
- García-Aljaro, C.; Balleste, E.; Muniesa, M. Beyond the canonical strategies of horizontal gene transfer in prokaryotes. Curr. Opin. Microbiol. 2017, 38, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.; Denyer, S.; Maillard, J.-Y. Rapid and quantitative automated measurement of bacteriophage activity against cystic fibrosis isolates of Pseudomonas aeruginosa. J. Appl. Microbiol. 2011, 110, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Bourdin, G.; Navarro, A.; Sarker, S.A.; Pittet, A.C.; Qadri, F.; Sultana, S.; Cravioto, A.; Talukder, K.A.; Reuteler, G.; Brüssow, H. Coverage of diarrhoea-associated Escherichia coli isolates from different origins with two types of phage cocktails. Microb. Biotechnol. 2014, 7, 165–176. [Google Scholar] [CrossRef]
- Konopacki, M.; Grygorcewicz, B.; Dołęgowska, B.; Kordas, M.; Rakoczy, R. PhageScore: A simple method for comparative evaluation of bacteriophages lytic activity. Biochem. Eng. J. 2020, 161, 107652. [Google Scholar] [CrossRef]
- Tey, B.; Ooi, S.; Yong, K.; Tan, N.; Ling, T.; Tan, W. Production of fusion m13 phage bearing the di-sulphide constrained peptide sequence (C-WSFFSNI-C) that interacts with hepatitis B core antigen. Afr. J. Biotechnol. 2009, 8, 6. [Google Scholar]
- Chandra, M.; Thakur, S.; Narang, D.; Saxena, H.M. Isolation of a bacteriophage against Salmonella Dublin and determination of its physical resistance under varied in vitro conditions. Afr. J. Microbiol. Res. 2011, 15, 2044–2047. [Google Scholar]
- Basdew, I.H.; Mark, D.L. Stress sensitivity assays of bacteriophages associated with Staphylococcus aureus, causal organism of bovine mastitis. Afr. J. Microbiol. Res. 2014, 8, 11. [Google Scholar]
- Ullah, A.; Qamash, T.; Khan, F.A.; Sultan, A.; Ahmad, S.; Abbas, M.; Khattak, M.A.K.; Begum, N.; Din, S.U.; Jamil, J.; et al. Characterization of a coliphage AS1 isolated from sewage effluent in Pakistan. Braz. J. Biol. 2021, 82, e240943. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, J.C.; Peabody, D.S. Stability and assembly in vitro of bacteriophage PP7 virus-like particles. J. Nanobiotechnol. 2007, 5, 10. [Google Scholar] [CrossRef]
- Wünsche, L. Importance of bacteriophages in fermentation processes. Acta Biotechnol. 2004, 9, 395–419. [Google Scholar] [CrossRef]
- Coffey, B.; Rivas, L.; Duffy, G.; Coffey, A.; Ross, R.P.; McAuliffe, O. Assessment of Escherichia coli O157:H7-specific bacteriophages e11/2 and e4/1c in model broth and hide environments. Int. J. Food Microbiol. 2011, 147, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Salmond, G.P.C.; Fineran, P.C. A century of the phage: Past, present and future. Nat. Rev. Microbiol. 2015, 13, 777–786. [Google Scholar] [CrossRef]
- Leptihn, S.; Gottschalk, J.; Kuhn, A. T7 ejectosome assembly: A story unfolds. Bacteriophage 2016, 6, e1128513. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Strelkov, S.V.; Mesyanzhinov, V.V.; Rossmann, M.G. Structure of bacteriophage T4 fibritin: A segmented coiled coil and the role of the C-terminal domain. Structure 1997, 5, 789–798. [Google Scholar] [CrossRef]
- White, R.; Chiba, S.; Pang, T.; Dewey, J.S.; Savva, C.G.; Holzenburg, A.; Pogliano, K.; Young, R. Holin triggering in real time. Proc. Natl. Acad. Sci. USA 2011, 108, 798–803. [Google Scholar] [CrossRef] [PubMed]
- Young, R. Phage lysis: Three steps, three choices, one outcome. J. Microbiol. 2014, 52, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Krieger, I.V.; Kuznetsov, V.; Chang, J.-Y.; Zhang, J.; Moussa, S.H.; Young, R.F.; Sacchettini, J.C. The structural basis of T4 phage lysis control: DNA as the signal for lysis inhibition. J. Mol. Biol. 2020, 432, 4623–4636. [Google Scholar] [CrossRef] [PubMed]
- Parma, D.H.; Snyder, M.; Sobolevski, S.; Nawroz, M.; Brody, E.; Gold, L. The Rex system of bacteriophage lambda: Tolerance and altruistic cell death. Genes. Dev. 1992, 6, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Snyder, L.; Kaufmann, G. T4 phage exclusion mechanisms. In Molecular Biology of Bacteriophage T4; Karam, J.D., Ed.; ASM: Washington, DC, USA, 1994; pp. 391–396. [Google Scholar]
- Paddison, P.; Abedon, S.T.; Dressman, H.K.; Gailbreath, K.; Tracy, J.; Mosser, E.; Neitzel, J.; Guttman, B.; Kutter, E. The roles of the bacteriophage T4 r genes in lysis inhibition and fine-structure genetics: A new perspective. Genetics 1998, 148, 1539–1550. [Google Scholar] [CrossRef]
- Folimonova, S.Y. Superinfection exclusion is an active virus-controlled function that requires a specific viral protein. J. Virol. 2012, 86, 5554–5561. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Ryu, S. Antirepression system associated with the life cycle switch in the temperate podoviridae phage SPC32H. J. Virol. 2013, 87, 11775–11786. [Google Scholar] [CrossRef] [PubMed][Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | Isolated Coliphages | |||
|---|---|---|---|---|
| STEC P1 | STEC P2 | STEC P3 | STEC P4 | |
| Shiga toxin-producing Escherichia coli | ||||
| Escherichia coli O157:H7 ATCC 35150 | + | + | + | + |
| Escherichia coli O157 86.24 | + | + | + | + |
| Escherichia coli O103 87-293 | + | + | + | + |
| Escherichia coli O26 Decaf | − | + | − | − |
| Escherichia coli O145 6940 | − | − | − | + |
| Escherichia coli O111 TB226A | − | − | − | − |
| Escherichia coli O45 4309-6 | − | − | − | − |
| Nontoxigenic Escherichia coli | ||||
| Escherichia coli O157 ATCC 100700 | + | + | + | + |
| Escherichia coli NRRL B-3008 | − | + | + | − |
| Escherichia coli K-12 MG1655: b2748 | − | + | − | − |
| Escherichia coli B-51077 | − | − | − | − |
| Escherichia coli ATCC 35218 | − | − | − | − |
| Escherichia coli ATCC 8739 | − | − | − | − |
| Other G- bacteria | ||||
| Salmonella typhimurium ATCC 14028 | − | − | − | − |
| Enterobacter aerogenes B-14144 | − | − | − | − |
| Shigella sonni ATCC 29930 | − | − | − | − |
| Shigella boydii * | − | + | − | − |
| Klebsiella pneumoniae * | − | − | − | − |
| Klebsiella pneumoniae * | − | − | − | − |
| Klebsiella quasipneumoniae * | − | − | − | − |
| Enterobacter cloacae * | − | − | − | − |
| Enterobacter cloacae * | − | − | − | − |
| Enterobacter asburiae * | − | − | − | − |
| Enterobacter ludwigii * | − | − | − | − |
| Enterobacter hormaechei subsp. Xiangfangensis * | − | − | − | − |
| MOI (pfu/cfu) | Phage Count (Log Average ± S.D.) | |
|---|---|---|
| STEC P2 | STEC P4 | |
| 1 (group 1) | 9.54 ± 0.50 cd | 10.67 ± 0.17 a |
| 0.1 (group 2) | 10.64 ± 0.15 a | 9.94 ± 0.29 bc |
| 0.01 (group 3) | 10.44 ± 0.30 ab | 9.24 ± 0.49 de |
| 0.001 (group 4) | 8.69 ± 0.27 e | 6.82 ± 0.37 f |
| Time (min) | STEC P2 | STEC P4 | ||
|---|---|---|---|---|
| Surviving Virions (Log Average ± S.D.) | Bacteriophage Reduction (%) | Surviving Virions (Log Average ± S.D.) | Bacteriophage Reduction (%) | |
| Zero time | 10.13 ± 0.24 a | 0.0 | 9.15 ± 0.23 b | 0.0 |
| 15 | 6.91 ± 0.76 c | 31.68 | 6.71 ± 0.07 c | 26.67 |
| 30 | 5.40 ± 0.66 d | 46.54 | 5.50 ± 0.36 d | 39.89 |
| 45 | 3.78 ± 0.1.06 ef | 62.57 | 4.44 ± 0.36 e | 51.58 |
| 60 | 1.80 ± 0.1 hi | 82.18 | 3.43 ± 0.68 fg | 62.55 |
| 75 | 1.10 ± 0.1 i | 89.11 | 2.61 ± 0.61 gh | 71.51 |
| Time | STEC P2 | STEC P4 | ||
|---|---|---|---|---|
| Surviving Virions (Log Average ± S.D.) | Bacteriophage Reduction (%) | Surviving Virions (Log Average ± S.D.) | Bacteriophage Reduction (%) | |
| Zero time | 11.65 ± 0.34 a | 0.0 | 10.38 ± 0.53 b | 0.0 |
| 72 °C/15 s | 10.53 ± 0.49 b | 9.6 | 9.62 ± 0.15 bc | 7.32 |
| 63 °C/30 min | 5.30 ± 0.73 d | 53.1 | 8.73 ± 0.67 c | 18.89 |
| 100 °C/10 min | 0.00 | 100 | 0.00 | 100 |
| 100 °C/20 min | 0.00 | 100 | 0.00 | 100 |
| 100 °C/30 min | 0.00 | 100 | 0.00 | 100 |
| pH | STEC P2 | STEC P4 | ||
|---|---|---|---|---|
| Surviving Virions (Log Average ± S.D.) | Bacteriophage Reduction (%) | Surviving Virions (Log Average ± S.D.) | Bacteriophage Reduction (%) | |
| 7 | 9.70 ± 0.1 a | 0.0 | 9.69 ± 0.17 a | 0.0 |
| 1 | 0.00 i | 100 | 7.73 ± 0.34 f | 20.23 |
| 3 | 6.60 ± 0.0 g | 31.96 | 8.21 ± 0.34 de | 15.27 |
| 5 | 8.80 ± 0.2 c | 9.28 | 8.33 ± 0.064 d | 14.04 |
| 9 | 9.45 ± 0.0 ab | 2.58 | 8.03 ± 0.32 e | 17.13 |
| 11 | 9.28 ± 0.1 b | 4.33 | 7.95 ± 0.31 ef | 17.96 |
| 13 | 1.88 ± 0.12 h | 80.62 | 0.00 i | 100 |
| Location on Escherichia Phage ST2 | Size bp | Closest Hits from NCBI | Query Cover | Similarity% | Function |
|---|---|---|---|---|---|
| 16,103–16,372 | 270 | Shigella phage JK42 | 100 | 100 | HP |
| 16,992–17,231 | 240 | Escherichia phage moskry | 100 | 99.58 | HP |
| 33,907–34,113 | 207 | Shigella phage JK42 | 100 | 100 | HP |
| 34,110–34,322 | 213 | Escherichia phage vB EcoM KAW3E185 | 100 | 99.06 | HP |
| 38,256–38,585 | 330 | Escherichia phage moskry | 100 | 99.39 | HP |
| 38,611–38,925 | 315 | Escherichia phage vB-EcoM-NBG1 | 100 | 99.37 | HP |
| 47,138–47,302 | 165 | Shigella phage JK42 | 100 | 99.39 | HP |
| 167,349–167,552 | 204 | Klebsiella phage Patroon | 100 | 89.71 | Holin |
| 167,590–167,976 | 387 | No significant similarity | - | - | Single-stranded DNA-binding protein |
| 167,998–168,645 | 648 | No significant similarity | - | - | Tail fiber protein |
| 168,675–170,066 | 1392 | Yersinia phage phiYe-F10 (Caudovirales) | 99 | 97.77 | Internal virion protein D |
| 170,080–170,235 | 156 | Citrobacter phage SH2(Caudovirales) | 100 | 100 | HP |
| 170,336–170,437 | 102 | Serratia phage SM9-3Y (Podoviridae) | 100 | 100 | HP |
| 170,448–170,903 | 456 | 100 | 99.34 | N-acetylmuramoyl-L-alanine amidase | |
| 170,982–171,086 | 105 | 85 | 96.67 | No significant similarity found |
| Location on Escherichia Phage ST4 | Size bp | Closest Hits from NCBI | Query Cover | Similarity% | Function |
|---|---|---|---|---|---|
| 2–331 | 330 | Escherichia phage vB EcoP G7C | 100 | 98.48 | Phage-associated DNA primase |
| 24,211–24,444 | 234 | No significant similarity | - | - | HP |
| 24,472–24,822 | 351 | Siphoviridae sp. | 98 | 95.36 | Phage protein |
| 25,203–25,445 | 243 | Escherichia phage vB EcoP G7C | 100 | 96.3 | HP |
| 25,753–25,935 | 183 | No significant similarity | - | - | HP |
| 25,932–26,174 | 243 | No significant similarity | - | - | HP |
| 27,946–28,152 | 207 | Enterobacter phage IME11 | 33 | 98.57 | HP |
| 29,561–29,902 | 342 | Siphoviridae sp. isolate 355 | 91 | 95.19 | HP |
| 34,547–37,714 | 3168 | No significant similarity | - | - | HP |
| 37,768–39,996 | 2229 | Escherichia albertii strain 2012EL-1823B | 60 | 85.41 | Peptidase |
| 40,002–40,151 | 150 | No significant similarity | - | - | HP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Sayed, D.; Elsayed, T.; Amin, N.; Al-Shahaby, A.; Goda, H. Evaluating the Phenotypic and Genomic Characterization of Some Egyptian Phages Infecting Shiga Toxin-Producing Escherichia coli O157:H7 for the Prospective Application in Food Bio-Preservation. Biology 2022, 11, 1180. https://doi.org/10.3390/biology11081180
El-Sayed D, Elsayed T, Amin N, Al-Shahaby A, Goda H. Evaluating the Phenotypic and Genomic Characterization of Some Egyptian Phages Infecting Shiga Toxin-Producing Escherichia coli O157:H7 for the Prospective Application in Food Bio-Preservation. Biology. 2022; 11(8):1180. https://doi.org/10.3390/biology11081180
Chicago/Turabian StyleEl-Sayed, Dina, Tarek Elsayed, Nadia Amin, Ahmad Al-Shahaby, and Hanan Goda. 2022. "Evaluating the Phenotypic and Genomic Characterization of Some Egyptian Phages Infecting Shiga Toxin-Producing Escherichia coli O157:H7 for the Prospective Application in Food Bio-Preservation" Biology 11, no. 8: 1180. https://doi.org/10.3390/biology11081180
APA StyleEl-Sayed, D., Elsayed, T., Amin, N., Al-Shahaby, A., & Goda, H. (2022). Evaluating the Phenotypic and Genomic Characterization of Some Egyptian Phages Infecting Shiga Toxin-Producing Escherichia coli O157:H7 for the Prospective Application in Food Bio-Preservation. Biology, 11(8), 1180. https://doi.org/10.3390/biology11081180