
Cryopreservation of Endangered Ornamental Plants and Fruit Crops from Tropical and Subtropical Regions



Abstract
:Simple Summary
Abstract
1. Introduction
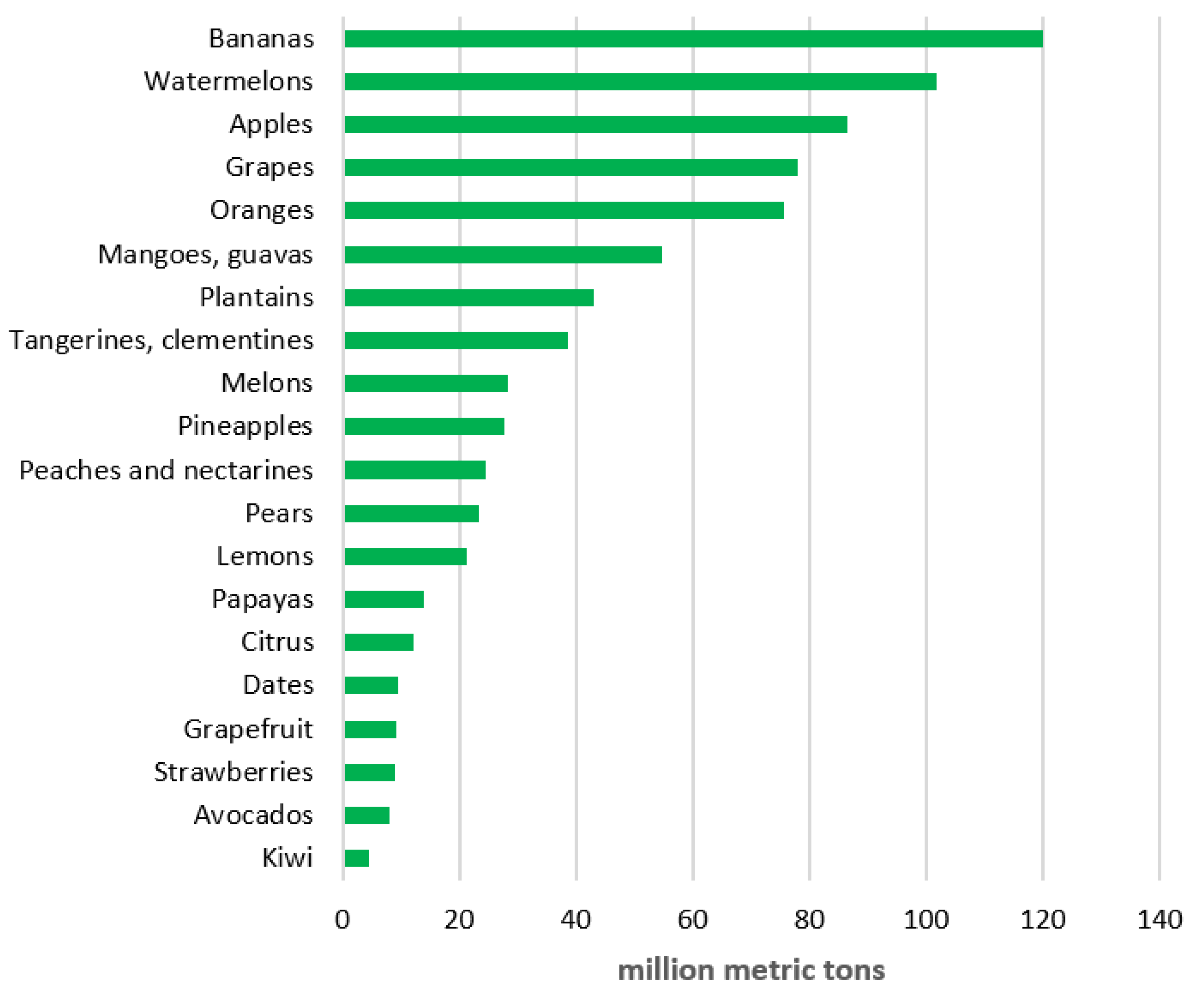
2. Tropical and Subtropical Ornamental and Fruit Plants
3. Importance and General Principles of Cryopreservation
4. Cryopreservation of Endangered Ornamentals and Fruit Crops
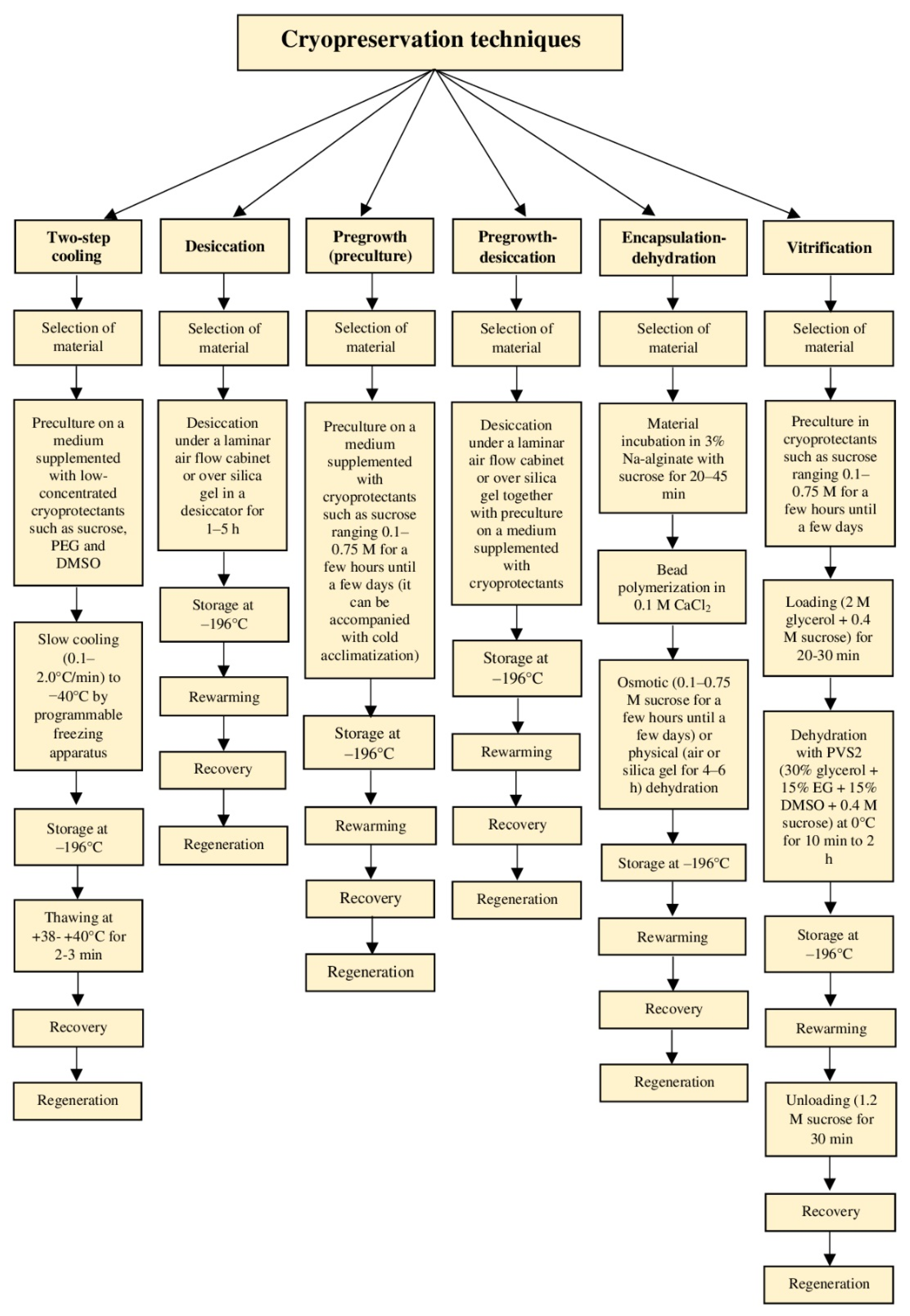
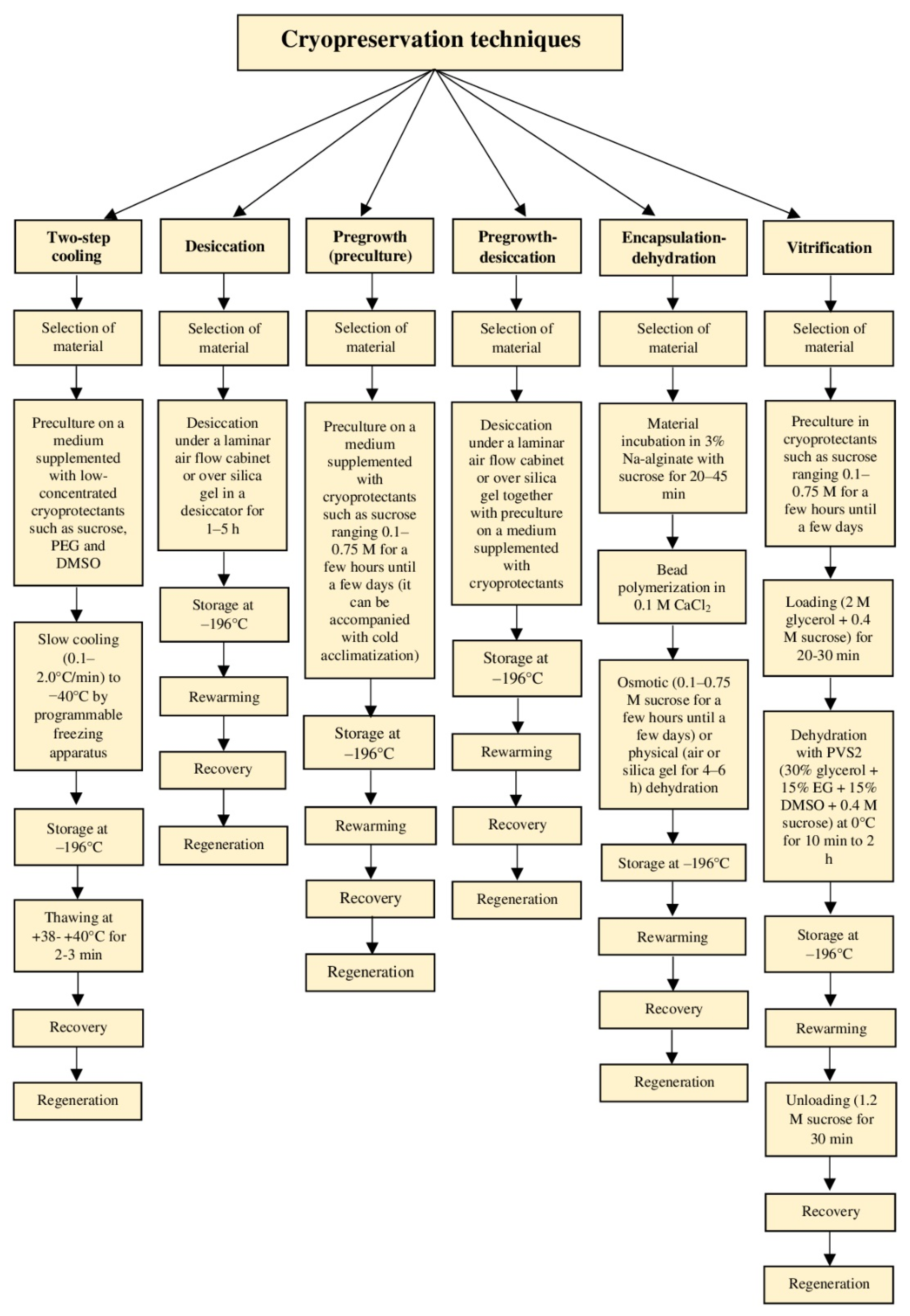
4.1. Conventional Methods (Two-Step Cooling)
4.2. Modern Methods (One-Step Cooling or Vitrification-Based Methods)
4.2.1. Desiccation
4.2.2. Pregrowth or Preculture
4.2.3. Pregrowth-Desiccation
4.2.4. Encapsulation-Dehydration
4.2.5. Vitrification
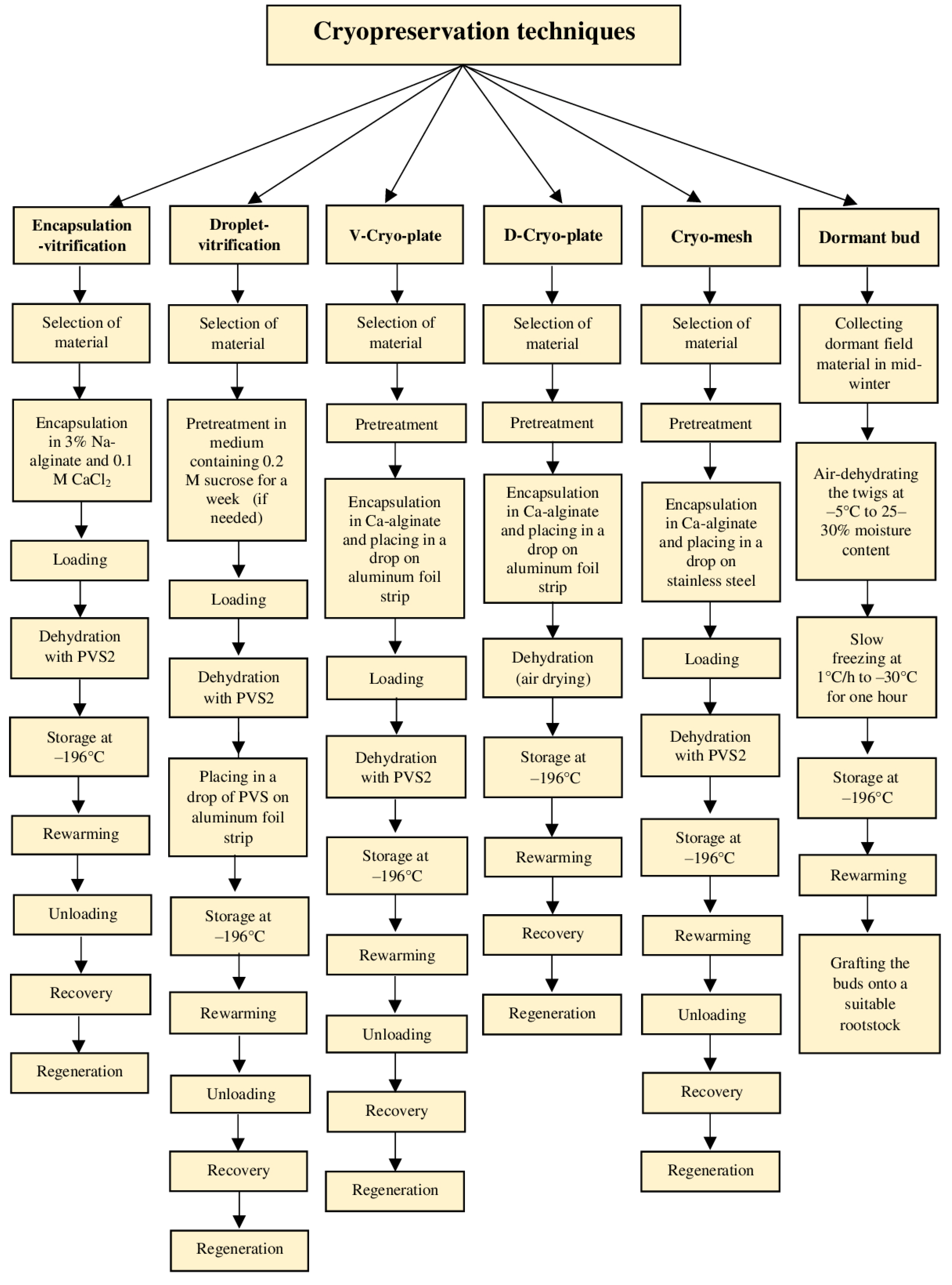
4.2.6. Encapsulation-Vitrification
4.2.7. Droplet-Vitrification
4.2.8. Cryo-Plates and Cryo-Mesh
4.2.9. Dormant Bud Cryopreservation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species and/or Cultivar | Explant Used | Method Applied | Survival (Recovery) [%] | Reference |
|---|---|---|---|---|
| Bletilla striata | Immature seeds | Direct immersion in LN vitrification | 8 81–92 | [105] |
| Bletilla striata | Mature seeds a, Germinating seeds b, Protocormsc | Droplet-vitrification | 93 a 91 b 84 c | [106] |
| Brassidium Shooting Star | PLBs | Droplet-vitrification | 30 | [85] |
| Brassidium Shooting Star | PLBs | Vitrification | No data | [107] |
| Brassidium Shooting Star | PLBs | Vitrification | No data | [108] |
| Buxus hyrcana | Shoot tips | Encapsulation-dehydration | 60.00 | [16] |
| Buxus sempervirens | Shoot tips | Encapsulation-dehydration a Encapsulation-vitrification b | 66.30 a 60.00 b | [109] |
| Cattleya spp. | Seeds | Vitrification | No data | [110] |
| Celisostoma areitinum | Protocorms | Encapsulation-dehydration | 49 | [111] |
| Centaurium rigualii | Nodes | Encapsulation-dehydration | 70 | [112] |
| Cymbidium hookerianum | PLBs | Preculture | 70 | [113] |
| Cyrtopodium hatschbachii | Immature seeds | Encapsulation-dehydration | 64 | [114] |
| Dendrobium cruentum | Protocorms | Vitrification Encapsulation-dehydration | 33 27 | [115] |
| Dendrobium candidum | PLBs | Encapsulation-vitrification | 85–89 | [80] |
| Dendrobium cariniferum | Protocorms | Encapsulation-vitrification | 15 | [115] |
| Dendrobium heterocarpum | Protocorms | Encapsulation-dehydration | 8 | [116] |
| Dendrobium nobile | PLBs | Encapsulation-dehydration | 53 (50) | [58] |
| Dendrobium nobile | PLBs | Encapsulation-vitrification | 78 (76) | [58] |
| Dendrobium Walter Oumae | Shoot tips | Encapsulation-dehydration | 16 (13) | [117] |
| Doritis pulcherrima | Seeds | Vitrification | 62 | [118] |
| Fritillaria imperialis Lubra Maxima | Bulb scale | Encapsulation-dehydration a Encapsulation-vitrification b | 74.30 a 81.6 b | [119] |
| Gentiana cruciata | Proembryogenic masses | Droplet-vitrification | 82 | [120] |
| Gentiana cruciata | Proembryogenic masses | Slow cooling | 2.5–2.7 | [121] |
| Gentiana cruciata | Proembryogenic masses | Vitrification | 86–91 | [121] |
| Gentiana scabra | Axillary buds | Preculture-desiccation | 5–90 | [49] |
| Gentiana sp. | Shoot tips a Axillary buds b | Vitrification | 74 a 78 b | [79] a, [49] b |
| Grammatophyllum speciosum | Protocorms | Droplet-vitrification | 38 | [59] |
| Grammatophyllum speciosum | Protocorms | Encapsulation-dehydration | 24 | [59] |
| Grammatophyllum speciosum | Protocorms | Encapsulation-vitrification | 14 | [59] |
| Lilium japonicum | Apical meristems | Vitrification | 68 | [71] |
| Lilium sp. | Shoot tips | Vitrification | 60–90 | [122] |
| Lilium spp. | Shoot tips | Droplet-vitrification | 42–87 | [123] |
| Lilium ledebourii | Seeds a,c,d,e Embryogenic axes a,b,c Lateral buds a,c Bulblet a,c Shoot tips f | Direct immersion in LN a/Vitrification b/Encapsulation-vitrificationc/Encapsulation-dehydration d/Preculture-desiccation e Droplet-vitrification f | 0 a/10 c/50 d/75 e 0 a/b/10 c 0 a/c 0 a/c 58–90 (53–88) f | [48,60,61] a–e, [124] f |
| Lilium× siberia | Apical meristems | Vitrification/Droplet-vitrification | 35–45/35–84 | [88] |
| Magnolia macrophylla | Shoot tips | Droplet-vitrification | 30 | [125] |
| Magnolia sinica | Seeds | Desiccation | 58 | [126] |
| Magnolia sirindhorniae | Shoot tips | Encapsulation-vitrification | 33 | [127] |
| Oncidium sp. | PLBs | Preculture-desiccation | 30 | [128] |
| Oncidium bifolium | Seeds Protocorms | Encapsulation-dehydration | 67 (5) 82 (11) | [129] |
| Oncidium flexuosum | Seeds | Vitrification | 78 | [130] |
| Phaius tankervillae | Seeds | Vitrification | 62 | [131] |
| Phalaenopsis bellina | PLBs | Encapsulation-dehydration | 47 | [64] |
| Preculture-desiccation | 30 | [64] | ||
| Pinus nigra | Proembryogenic masses | Slow cooling | 88 | [132] |
| Rhynchostylis gigantean | Protocorms | Vitrification | 19 | [115] |
| Rosa × hybrida | Shoot tips Axillary buds | Droplet-vitrification a Encapsulation-dehydration b | (58–64) a (12) a (0) b | [133] a, [134] b |
| Seidenfadenia mitrata | Protocorms | Vitrification | 67 | [115] |
| Vanda coerulea | PLBs | Droplet-vitrification | 5 | [135] |
| Vanda coerulea | Protocorms | Encapsulation-dehydration | 40 | [136] |
| Vanda coerulea | Seeds | Vitrification | 67 | [137] |
| Vanda tricolor | Mature seeds | Direct immersion in LN vitrification | 10 14 | [68] |
| Vanilla planifolia Andrews | Shoot apices | Droplet-vitrification | 30 (10) | [138] |
| Plant Species | Explant | Cryopreservation Technique | Survival (%) | Reference |
|---|---|---|---|---|
| Actinidia spp. | Shoot tips | Droplet-vitrification | 59–88 | [139] |
| Encapsulation-dehydration | 85–95 | [140] | ||
| Agave peacockii | Shoot tips | Droplet-vitrification | 96 | [141] |
| Agave tequilana | Somatic embryos | V cryo-plate technique | 83 | [142] |
| Ananas spp. | Shoot tips | Droplet-vitrification | 51 | [143] |
| Ananas (wild genus) | Pollen | Dehydration | 62 | [144] |
| Citrus sinensis | Callus | Modified aluminum cryo-plate | 88 | [145] |
| Citrus spp. | Shoot tips | Droplet-vitrification | 56 | [146] |
| Cocos nucifera | Shoot tips | Droplet-vitrification | 50 | [147] |
| Diospyros kaki | Shoot tips | D cryo-plate | 67–97 | [99] |
| Diospyros kaki | Shoot tips | Encapsulation-droplet-vitrification a Slow freezing b Vitrification c | 80 a 70–76 b 86 c | [148] a, [149] b, [150] b, [151] c |
| Diospyros spp. | Shoot tips | Vitrification a | 30 a 100 b | [152] a, [153] b |
| Musa spp. | Apical meristems | Droplet-vitrification | 39 | [36] |
| Musa spp. | Cell suspensions | Classical (slow) freezing | No data | [154] |
| Passiflora edulis | Zygotic embryos | Dehydration | 100 | [155] |
| Passiflora suberosa | Shoot tips | Encapsulation-vitrification | 28 | [156] |
| Passiflora pohlii | Nodal segments | Vitrification | 65 | [157] |
| Persea americana | Shoot tips | Vitrification | 73–80 | [158] |
| Persea americana | Somatic embryos | Cryovial-vitrification a Droplet-vitrification b | 73–91 a 85–100 b | [159] |
| Prunus cerasifera | Shoot tips | V and D cryo-plates | 56.1% (V cryo-plate) and 77.5% (D cryo-plate) | [97] |
| Prunus cerasus | Dormant bud | No data | No data | [160] |
| Prunus domestica | Shoot tips | V and D cryo-plates | 44.6% (V cryo-plate) and 47.5% (D cryo-plate) | [97] |
| Prunus spp. | Shoot tips | Encapsulation-dehydration a Vitrification b Droplet-vitrification c Two-step freezing d | 14–76 a 60–88 b 20–52 c 74 d | [161] a, [162] b, [97] c, [163] c, [163] d |
| Pyrus spp. | Shoot tips | Encapsulation-dehydration a Vitrification b Two step freezing c | 30–82 a 71 b 75–83 c | [164] a, [165] b, [166] c, [167] c |
5. Rewarming and Recovery
6. Cryopreservation and Omics Technologies
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roque-Borda, C.A.; Kulus, D.; de Souza, A.V.; Kaviani, B.; Vicente, E.F. Cryopreservation of agronomic plant germplasm using vitrification-based methods: An overview of selected case studies. Int. J. Mol. Sci. 2021, 22, 6157. [Google Scholar] [CrossRef]
- Krupnick, G.A. Conservation of tropical plant biodiversity: What have we done, where are we going? BioTropica 2013, 45, 693–708. [Google Scholar] [CrossRef]
- Corlett, R.T. Safeguarding our future by protecting biodiversity. Plant Diver. 2020, 42, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Reed, B.M. Implementing cryopreservation storage of clonally propagated plants. Cryo-Letters 2001, 22, 97–104. [Google Scholar]
- Panis, B. Sixty years of plant cryopreservation: From freezing hardy mulberry twigs to establishing reference crop collections for future generations. Acta Hortic. 2019, 1234, 1–7. [Google Scholar] [CrossRef]
- Wang, M.R.; Lambardi, M.; Engelmann, F.; Pathirana, R.; Panis, B.; Volk, G.M.; Wang, Q.C. Advances in cryopreservation of in vitro-derived propagules: Technologies and explant sources. Plant Cell Tissue Org. Cult. 2020, 144, 7–20. [Google Scholar] [CrossRef]
- Bernard, F.; Shaker-Bazarnov, H.; Kaviani, B. Effect of salicylic acid on cold preservation and cryopreservation of encapsulated embryonic axes of Persian lilac (Melia azedarach L.). Euphytica 2002, 123, 85–88. [Google Scholar] [CrossRef]
- Engelmann, F. Use of biotechnologies for the conservation of plant biodiversity. In Vitro Cell. Dev. Biol. Plant 2011, 47, 5–16. [Google Scholar] [CrossRef]
- Li, J.-W.; Zhang, X.-C.; Wang, M.-R.; Bi, W.-L.; Faisal, M.; da Teixeira Silva, J.A.; Volk, J.M.; Wang, Q.-C. Development, progress and future prospects in cryobiotechnology of Lilium spp. Plant Methods 2019, 15, 125. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, F. Plant cryopreservation: Progress and prospects. In Vitro Cell. Dev. Biol. Plant 2004, 40, 427–433. [Google Scholar] [CrossRef]
- Panis, B.; Piette, B.; Andreé, E.; Van den Houwe, I.; Swennen, R. Droplet vitrification: The first generic cryopreservation protocol for organized plant tissues? Acta Hortic. 2011, 908, 157–162. [Google Scholar] [CrossRef]
- Kaviani, B. Conservation of plant genetic resources by cryopreservation. Aust. J. Crop Sci. 2011, 5, 778–800. [Google Scholar]
- Cruz-Cruz, C.A.; González-Arnao, M.T.; Engelmann, F. Biotechnology and conservation of plant biodiversity. Resources 2013, 2, 73–95. [Google Scholar] [CrossRef]
- Acker, J.P.; Adkins, S.; Alves, A.; Horna, D.; Toll, J. Feasibility study for a safety back-up cryopreservation facility. In Independent Expert Report; Bioversity International: Rome, Italy, 2017; p. 100. [Google Scholar]
- Kulus, D.; Zalewska, M. Cryopreservation as a tool used in long-term storage of ornamental species—A review. Sci. Hortic. 2014, 168, 88–107. [Google Scholar] [CrossRef]
- Kaviani, B.; Negahdar, N. Propagation, micropropagation and cryopreservation of Buxus sempervirens Pojark., an endangered ornamental shrub. S. Afr. J. Bot. 2017, 111, 326–335. [Google Scholar] [CrossRef]
- Fukai, S. Plant regeneration from shoot tips of Dianthus hybrid cryopreserved in liquid nitrogen up to 2 years. Plant Tissue Cult. Lett. 1989, 6, 177–178. [Google Scholar] [CrossRef]
- Sakai, A. Survival of plant tissue of super-low temperature. Contrib. Inst. Temp. Sci. Haikkaido Univ. Ser. B 1956, 14, 17. [Google Scholar]
- González-Arnao, M.T.; Martinez-Montero, M.E.; Cruz-Cruz, C.A.; Engelmann, F. Advances in cryogenic techniques for the long-term preservation of plant biodiversity Maria Teresa. In Biotechnology and Biodiversity, Sustainable Development and Biodiversity; Ahuja, M.R., Ramawat, K.G., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 129–170. [Google Scholar]
- Wang, B.; Zhang, Z.; Yin, Z.; Feng, C.; Wang, Q. Novel and potential application of cryopreservation to plant genetic transformation. Biotechnol. Adv. 2012, 30, 604–612. [Google Scholar] [CrossRef]
- Harding, K. Genetic integrity of cryopreservaed plant cells: A review. CryoLetters 2004, 25, 3–22. [Google Scholar]
- Martinez-Montero, M.E.; Harding, K. Cryobionomics: Evaluating the concept in plant cryopreservation. In Plant Omics: The Omics of Plant Science; Barh, D., Khan, M., Davies, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 655–682. [Google Scholar]
- Gabellini, S.; Scaramuzzi, S. Evolving consumption trends, marketing strategies, and governance settings in ornamental horticulture: A grey literature review. Horticulturae 2022, 8, 234. [Google Scholar] [CrossRef]
- Salachna, P. Trends in ornamental plant production. Horticulturae 2022, 8, 413. [Google Scholar] [CrossRef]
- Normah, M.N.; Sulong, N.; Reed, B.M. Cryopreservation of shoot tips of recalcitrant and tropical species: Advances and strategies. Cryobiology 2019, 87, 1–14. [Google Scholar] [CrossRef]
- Malik, S.K.; Chaudhury, R. Cryopreservation techniques for conservation of tropical horticultural species using various explants. In Conservation and Utilization of Horticultural Genetic Resources; Rajasekharan, P., Rao, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 579–594. [Google Scholar]
- Viracheva, L.L.; Goncharova, O.A.; Kirillova, N.R.; Nosatenko, O.Y.; Trostenyuk, N.N. Rare and disappearing plants in the introductive collection of the Polar-Alpine Botanical Garden and Institute. Hort. Bot. 2019, 14, 387–402. [Google Scholar] [CrossRef]
- Global Fruit Production in 2020. Available online: www.statista.com (accessed on 2 May 2022).
- Zamecnik, J.; Faltus, M.; Bilavcik, A. Vitrification solutions for plant cryopreservation: Modification and properties. Plants 2021, 10, 2623. [Google Scholar] [CrossRef]
- Kulus, D.; Abratowska, A.; Mikuła, A. Morphogenetic response of shoot tips to cryopreservation by encapsulation-dehydration in a solid mutant and periclinal chimeras of Chrysanthemum × grandiflorum/Ramat./Kitam. Acta Physiol. Plant 2018, 40, 18. [Google Scholar] [CrossRef]
- Barh, D.; Sarwar Khan, M.; Davies, E. Plant Omics: The Omics of Plant Science; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Benson, E.E. Cryopreservation of phytodiversity: A critical appraisal of theory & practice. Crit. Rev. Plant Sci. 2008, 27, 141–219. [Google Scholar]
- Uemura, M.; Minami, A.; Kawamura, Y. Effect of low temperature and cryoprotectants on plant plasma membrane. In 1st International Symposium: Cryopreservation in Horticultural Species, Book of Abstracts; ISHS: Leuven, Belgium, 2009; p. 15. [Google Scholar]
- Ciani, F.; Cocchia, N.; Esposito, L.; Avallone, L. Fertility cryopreservation. In Advances in Embryo Transfer; Wu, B., Ed.; In Tech: Rijeka, Croatia, 2012. [Google Scholar]
- Benson, E.E.; Betsou, F.; Fuller, B.J.; Harding, K.; Kofanova, O. Translating cryobiology principles into transdisciplinary storage guidelines for biorepositories and biobanks: A concept paper. CryoLetters 2013, 34, 277–312. [Google Scholar]
- Panis, B.; Piette, B.; Swennen, R. Droplet vitrification of apical meristems: A cryopreservation protocol applicable to all Musaceae. Plant Sci. 2005, 168, 45–55. [Google Scholar] [CrossRef]
- Panis, B.; Lambardi, M. Status of cryopreservation technologies in plants (crops and forest trees). In The Role of Biotechnology; Villa Gualino: Turin, Italy, 2005; pp. 43–54. [Google Scholar]
- Jiroutová, P.; Sedlák, J. Cryobiotechnology of plants: A hot topic not only for gene banks. Appl. Sci. 2020, 10, 4677. [Google Scholar] [CrossRef]
- Panis, B.; Nagel, M.; Van den Houwe, I. Challenges and prospects for the conservation of crop genetic resources in field genebanks, in in vitro collections and/or in liquid nitrogen. Plants 2020, 9, 1634. [Google Scholar] [CrossRef]
- Benson, E.; Harding, K.; Debouck, D.; Dumet, D.; Escobar, R.; Mafla, G.; Panis, B.; Panta, A.; Tay, D.; Van den Houwe, I.; et al. Refinement and standardization of storage procedures for clonal Crops-Global Public Goods Phase 2: Part II. In Status of In Vitro Conservation Technologies for: Andean Root and Tuber Crops, Cassava, Musa, Potato, Sweet Potato and Yam; System-Wide Genetic Resources Programme: Rome, Italy, 2011; pp. 15–30. [Google Scholar]
- Engelmann, F. Germplasm collection, storage and preservation. In Plant Biotechnology and Agriculture—Prospects for the 21st Century; Altman, A., Hazegawa, P.M., Eds.; Academic Press: Oxford, UK, 2012; pp. 255–268. [Google Scholar]
- González-Arnao, M.T.; Panta, A.; Roca, W.M.; Escobar, R.H.; Engelmann, F. Development and large scale application of cryopreservation techniques for shoot and somatic embryo cultures of tropical crops. Plant Cell Tissue Org. Cult. 2008, 92, 1–13. [Google Scholar] [CrossRef]
- Benson, E.E.; Johnston, J.; Muthusamy, J.; Harding, K. Physical and engineering perspectives of in vitro plant cryopreservation. In Plant Tissue Culture Engineering; Gupta, S., Ibaraki, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 6, pp. 441–476. [Google Scholar]
- Heine-Dobbernack, E.; Seufert, S.; Schumacher, H.M. Controlled rate freezing of dedifferentiated plant cell lines—A mini-test system for quick evaluation of parameters. Cryobiology 2006, 53, 367–446. [Google Scholar] [CrossRef]
- Reed, B.M. Cryopreservation-practical considerations. In Plant Cryopreservation: A Practical Guide; Reed, B.M., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 3–13. [Google Scholar]
- Engelmann, F.; Takagi, H. Cryopreservation of Tropical Plant Germplasm: Current Research Progress and Application; IPGRI: Tsukuba, Japan, 2000; pp. 8–20. [Google Scholar]
- Hamilton, K.N.; Ashmore, S.E.; Pritchard, H.W. Thermal analysis and cryopreservation of seeds of Australian Wild Citrus species (Rutaceae): Citrus australasica, C. inodora and C. garrawayi. CryoLetters 2009, 30, 268–279. [Google Scholar]
- Kaviani, B.; Abadi, D.H.; Torkashvand, A.M.; Hoor, S.S. Cryopreservation of seeds of lily (Lilium ledebourii Baker Bioss.): Use of sucrose and dehydration. Afr. J. Biotechnol. 2009, 8, 3809–3810. [Google Scholar]
- Suzuki, M.; Ishikawa, M.; Okuda, H.; Noda, K.; Kishimoto, T.; Nakamura, T.; Ogiwara, I.; Shimura, I.; Akihama, T. Physiological changes in Gentian axillary buds during two-step preculturing with sucrose that conferred high levels of tolerance to desiccation and cryopreservation. Ann. Bot. 2006, 97, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, F. In vitro conservation methods. In Biotechnology and Plant Genetic Resources; Callow, J.A., Ford-Lloyd, B.V., Newbury, H.J., Eds.; CAB International: Oxford, UK, 1997; pp. 119–161. [Google Scholar]
- Wilkinson, T.; Wetten, A.; Prychid, C.; Fay, M.F. Suitability of cryopreservation for the long-term storage of rare and endangered plant species: A case history of Cosmos atrosanguineus. Ann. Bot. 2003, 91, 65–74. [Google Scholar] [CrossRef] [PubMed]
- González-Arnao, M.T.; Lazaro-Vallejo, C.E.; Valencia, M.G.; Ortiz Patraca, N.M.; Tex-cahua Martinez, H. Adjustment of cryoprotective conditions for vanilla (Vanilla planifolia) shoot-tips subjected to a droplet-vitrification protocol. Cryobiology 2010, 61, 362–408. [Google Scholar] [CrossRef]
- Engelmann, F. Encapsulation-dehydration: Past, present and future. Acta Hortic. 2009, 908, 165–171. [Google Scholar]
- da Teixeira Silva, J.A.; Zeng, S.; Galdiano, R.F.; Dobránszki, J.; Cardoso, J.C.; Vendrame, W.A. In vitro conservation of Dendrobium germplasm. Plant Cell Rep. 2014, 33, 1413–1423. [Google Scholar] [CrossRef]
- Ozden-Tokatli, Y.; De Carlo, A.; Gumusel1, F.; Pignattelli, S.; Lambardi, M. Development of encapsulation techniques for the production and conservation of synthetic seeds in ornamental species. Propag. Ornam. Plants 2008, 8, 17–22. [Google Scholar]
- Kaviani, B.; Bernard, F.; Shakeri, H.; Hadadchi, G.H.R. Effects of salic acid on enhancing the resistance of embryonic axes of Persian lilac (Melia azedarach L.) against cold and cryopreservation. Agron. Hortic. 2005, 67, 44–49. [Google Scholar]
- Kaviani, B. Effects of salicylic acid and encapsulation on enhancing the resistance of embryonic axes of Persian lilac (Melia azedarach L.) against cryopreservation. Int. J. Agri. Biol. 2007, 9, 625–627. [Google Scholar]
- Mohanty, P.; Das, M.C.; Kumaria, S.; Tandon, P. High-efficiency cryopreservation of the medicinal orchid Dendrobium nobile Lindl. Plant Cell Tissue Org. Cult. 2012, 109, 297–305. [Google Scholar] [CrossRef]
- Sopalun, K.; Kanchit, K.; Ishikawa, K. Vitrification-based cryopreservation of Grammatophyllum speciosum protocorm. CryoLetters 2010, 31, 347–357. [Google Scholar]
- Kaviani, B.; Safari-Motlagh, M.R.; Padasht-Dehkaei, M.N.; Darabi, A.H.; Rafizadeh, A. Cryopreservation of lily [Lilium ledebourii (Baker) Bioss.] germplasm by encapsulation-dehydration. Int. J. Bot. 2008, 4, 491–493. [Google Scholar] [CrossRef]
- Kaviani, B.; Dahkaei, M.; Hashemabadi, D.; Darabi, A. Cryopreservation of Lilium ledebourii (Baker) Bioss. by encapsulation-vitrification and in vivo media for planting of germplasm. Am.-Eur. J. Agric. Environ. Sci. 2010, 8, 556–560. [Google Scholar]
- Kaviani, B. Cryopreservation by encapsulation-dehydration for long-term storage of some important germplasm: Seed of lily [Lilium ledebourii (Baker) Bioss.], embryonic axe of Persian lilac (Melia azedarach L.), and tea (Camellia sinensis L.). Plant Omics J. 2010, 3, 177–182. [Google Scholar]
- Yin, L.L.; Poobathy, R.; James, J.; Julkifle, A.L.; Subramaniam, S. Preliminary investigation of cryopreservation by encapsulation-dehydration technique on Brassidium Shooting Star orchid hybrid. Afr. J. Biotechnol. 2011, 10, 4665–4672. [Google Scholar]
- Khoddamzadeh, A.A.; Sinniah, U.R.; Lynch, P.; Kadir, M.A.; Kadzimin, S.B.; Mahmood, M. Cryopreservation of protocorm-like bodies (PLBs) of Phalaenopsis bellina (Rchb. f.) Christenson by encapsulation-dehydration. Plant Cell Tissue Org. Cult. 2011, 107, 471–481. [Google Scholar] [CrossRef]
- Subramaniam, S.; Sinniah, U.R.; Khoddamzadeh, A.L.; Periasamy, S.; James, J.J. Fundamental concept of cryopreservation using Dendrobium Sonia-17 protocorm-like bodies by encapsulation-dehydration technique. Afr. J. Biotechnol. 2011, 10, 3902–3907. [Google Scholar]
- Bonnart, R.; Volk, G.M. Increased efficiency using the encapsulation-dehydration cryopreservation technique for Arabidopsis thaliana. CryoLetters 2010, 31, 95–100. [Google Scholar]
- Benelli, C.; De Carlo, A.; Engelmann, F. Recent advances in the cryopreservation of shoot-derived germplasm of economically important fruit trees of Actinidia, Diospyros, Malus, Olea, Prunus. Biotechnol. Adv. 2013, 31, 175–185. [Google Scholar] [CrossRef]
- Jitsopakul, N.; Thammasiri, K.; Yukawa, C.; Ishikawa, K. Effect of cryopreservation on seed germination and protocorm development of Vanda tricolor. Sci. Asia. 2012, 38, 244–249. [Google Scholar] [CrossRef]
- Sekizawa, K.; Yamamoto, S.; Rafique, T.; Fukui, K.; Niino, T. Cryopreservation of in vitro-grown shoot tips of carnation (Dianthus caryophyllus L.) by vitrification method using aluminum cryo-plates. Plant Biotechnol. 2011, 28, 401–405. [Google Scholar] [CrossRef]
- Kim, H.-H.; Popova, E.; No, N.-Y.; Back, H.-J.; Kim, C.-K.; Cho, E.-G.; Engelmann, F. Application of alternative loading solutions to garlic and chrysanthemum in droplet-vitrification procedures. Acta Hortic. 2011, 908, 173–180. [Google Scholar] [CrossRef]
- Matsumoto, T.; Sakai, A.; Yamada, K. Cryopreservation of in vitro-grown apical meristems of lily by vitrification. Plant Cell Tissue Org. Cult. 1995, 41, 237–241. [Google Scholar] [CrossRef]
- Moges, A.D.; Shibli, R.A.; Karam, N.S. Cryopreservation of African violet (Saintpaulia ionantha Wendl.) shoot tips. In Vitro Cell. Dev. Biol. Plant 2004, 40, 389–395. [Google Scholar] [CrossRef]
- Takagi, H.; Tien Thinh, N.T.; Islam, O.M.; Senboku, T.; Sakai, A. Cryopreservation of in vitro-grown shoot tips of taro (Colocasia esculenta (L.) Schott) by vitrification. 1. Investigation of basic conditions of the vitrification procedure. Plant Cell Rep. 1997, 16, 594–599. [Google Scholar] [CrossRef]
- Thinh, N.T.; Takagi, H. Cryopreservation of Colocasia esculenta L. Schott (Taro). In Biotechnology in Agriculture and Forestry; Towill, L.E., Bajaj, Y.P.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; Volume 50, pp. 96–119. [Google Scholar]
- Antony, J.J.J.; Keng, C.; Rathinam, X.; Marimuthu, S.; Subramaniam, S. Effect of preculture and PVS2 incubation conditions followed by histological analysis in the cryopreservaed PLBs of Dendrobium Bobby Messina orchid. Aust. J. Crop Sci. 2011, 5, 1557–1564. [Google Scholar]
- Thinh, N.T. Cryopreservation of Germplasm of Vegetatively Propagated Tropical Monocots by Vitrification. Ph.D. Thesis, Kobe University, Kobe, Japan, 1997. [Google Scholar]
- Cripps, R.F.; McGregor, K. Determination of the optimal dehydration period for the protocorms of Paralophia epipyhtica (Orchidaceae) using differential scanning calorimetry. Cryobiology 2009, 53, 367–446. [Google Scholar]
- Kim, H.H.; Lee, Y.G.; Shin, D.J.; Ko, H.C.; Gwag, J.G.; Cho, E.G.; Engelmann, F. Development of alternative plant vitrification solutions in droplet-vitrification procedures. Cryobiology 2009, 59, 370–418. [Google Scholar] [CrossRef]
- Sakai, A.; Engelmann, F. Vitrification, encapsulation-vitrification and droplet-vitrification: A review. CryoLetters 2007, 28, 151–172. [Google Scholar]
- Yin, M.; Hong, S. Cryopreservation of Dendrobium candidum Wall. ex Lindl. protocorm-like bodies by encapsulation-vitrification. Plant Cell Tissue Org. Cult. 2009, 98, 179–185. [Google Scholar] [CrossRef]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification. Plant Cell Rep. 1990, 9, 30–33. [Google Scholar] [CrossRef]
- Pennycooke, J.C.; Towill, L.E. Cryopreservation of shoot tips from in vitro plants of sweet potato [Ipomoea batatas (L.) Lam.] by vitrification. Plant Cell Rep. 2000, 19, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Niino, T.; Yamamoto, S.; Matsumoto, T.; Engelmann, F.; Arizaga, M.V.; Tanaka, D. Development of V and D cryo-plate methods as effective protocols for cryobanking. Acta Hortic. 2019, 1234, 249–262. [Google Scholar] [CrossRef]
- Towill, L.E.; Bonnart, R. Cracking in a vitrification solution during cooling or warming does not affect growth of cryopreserved mint shoot tips. CryoLetters 2003, 24, 341–346. [Google Scholar] [PubMed]
- Rahmah, R.; Mubbarakh, S.A.; Sinniah, U.R.; Subramaniam, S. Effects of droplet-vitrification on Brassidium Shooting Star’s orchid protocorm-like bodies (PLBs). Sci. Hortic. 2015, 197, 254–260. [Google Scholar] [CrossRef]
- Kim, H.M.; Shin, J.H.; Sohn, J.K. Cryopreservation of somatic embryos of the herbaceous peony (Paeonia lactiflora Pall.) by air drying. Cryobiology 2006, 1, 69–74. [Google Scholar] [CrossRef]
- Sant, R.; Panis, B.; Taylor, M.; Tyagi, A. Cryopreservation of shoot tips by droplet vitrification applicable to all taro (Colocasia esculenta var. esculenta) accessions. Plant Cell Tissue Org. Cult. 2008, 92, 107–111. [Google Scholar] [CrossRef]
- Chen, X.L.; Li, J.H.; Xin, X.; Zhang, Z.E.; Xin, P.P.; Lu, X.X. Cryopreservation of in vitro-grown apical meristems of Lilium by droplet-vitrification. S. Afr. J. Bot. 2011, 77, 397–403. [Google Scholar] [CrossRef]
- Wang, M.R.; Chen, L.; da Teixeira Silva, J.A.; Volk, G.M.; Wang, Q.-C. Cryobiotechnology of apple (Malus spp.): Development, progress and future prospects. Plant Cell Rep. 2018, 37, 689–709. [Google Scholar] [CrossRef]
- Yamamoto, S.; Fukui, K.; Niino, T. A new cryopreservation method for vegetatively propagated plant genetic resources using aluminum cryo-plates. Dev. Technol. 2011, 10, 10–11. [Google Scholar]
- Niino, T.; Yamamoto, S.; Fukui, K.; Martínez, C.R.C.; Arizaga, M.V.; Matsumoto, T.; Engelmann, F. Dehydration improves cryopreservation of mat rush (Juncus decipiens Nakai) basal stem buds on cryo-plates. CryoLetters 2013, 34, 549–560. [Google Scholar] [PubMed]
- Matsumoto, T. Cryopreservation of plant genetic resources: Conventional and new methods. Rev. Agric. Sci. 2017, 5, 13–20. [Google Scholar] [CrossRef]
- Salma, M.; Fki, L.; Engelmann-Sylvestre, I.; Niino, T.; Engelmann, F. Comparison of droplet-vitrification and D-cryoplate for cryopreservation of date palm (Phoenix dactylifera L.) polyembryonic masses. Sci. Hortic. 2014, 179, 91–97. [Google Scholar] [CrossRef]
- Matsumoto, T.; Yamamoto, S.; Fukui, K.; Niino, T. Cryopreservation of blueberry dormant shoot tips using V cryoplate method. Ann. Conf. Am. Soc. Hortic. Sci. 2014, 49, S337. [Google Scholar]
- Rafique, T.; Yamamoto, S.; Fukui, K.; Mahmood, Z.; Niino, T. Cryopreservation of sugarcane using the V cryo-plate technique. CryoLetters 2015, 36, 51–59. [Google Scholar]
- Engelmann-Sylvestre, I.; Engelmann, F. Cryopreservation of in vitro-grown shoot tips of Clinopodium odorum using aluminum cryo-plates. In Vitro Cell. Dev. Biol. Plant 2015, 51, 185–191. [Google Scholar] [CrossRef]
- Vujović, T.; Chatelet, P.; Ružića, D.; Engelmann, F. Cryopreservation of Prunus spp. using aluminum cryo-plates. Sci. Hortic. 2015, 195, 173–182. [Google Scholar] [CrossRef]
- Arizaga, M.V.; Yamamoto, S.I.; Tanaka, D.; Fukui, K.; Nohara, N.; Nishikawa, T.; Watanabe, K.N. Cryopreservation of in vitro shoot tips of ulluco (Ullucus tuberosus Cal.) using D cryo-plate method. CryoLetters 2017, 38, 419–427. [Google Scholar]
- Matsumoto, T.; Yamamoto, S.; Fukui, K.; Rafique, T.; Engelmann, F.; Niino, T. Cryopreservation of persimmon shoot tips from dormant buds using the D cryo-plate technique. Hortic. J. 2015, 84, 106–110. [Google Scholar] [CrossRef]
- Funnekotter, B.; Bunn, E.; Mancera, R.L. Cryo-mesh: A simple alternative cryopreservation protocol. CryoLetters 2017, 38, 155–159. [Google Scholar] [PubMed]
- Topal-Celikkan, F.; Ozkavukcu, S.; Balci, D.; Serin-Kilicoglu, S.; Atabenli-Erdemli, E. Mouse ovarian tissue vitrification on copper electron microscope grids versus slow freezing: A comparative ultrastructural study. Reprod. Fertil. Dev. 2015, 27, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- Reed, B.M.; Sarasan, V.; Kane, M.; Bunn, E.; Pence, V.C. Biodiversity conservation and biotechnology tools. In Vitro Cell. Dev. Biol. Plant 2011, 47, 1–4. [Google Scholar] [CrossRef]
- Lambardi, M.; Benelli, C.; De Carlo, A.; Ozudogru, E.A.; Previati, A.; Ellis, D. Cryopreservation of ancient apple cultivars of Veneto: A comparison between PVS2-vitrification and dormant-bud techniques. Acta Hortic. 2011, 908, 191–198. [Google Scholar] [CrossRef]
- Fukui, K.; Shirata, K.; Niino, T.; Kashif, I.M. Cryopreservation of mulberry winter buds in Japan. Int. Symp. Cryopreserv. Hortic. Species 2011, 908, 483–488. [Google Scholar] [CrossRef]
- Hirano, T.; Godo, T.; Mii, M.; Ishikawa, K. Cryopreservation of immature seeds of Bletilla striata by vitrification. Plant Cell Rep. 2005, 23, 534–539. [Google Scholar] [CrossRef]
- Jitsopakul, N.; Thammasiri, K.; Ishikawa, K. Cryopreservation of Bletilla striata mature seeds, 3-day germinating seeds and protocorms by droplet-vitrification. CryoLetters 2008, 29, 517–526. [Google Scholar]
- Antony, J.J.J.; Burkhan, H.; Sinniah, U.R.; Poobathy, R.; Subramaniam, S. Effect of PVS2 vitrification on Brassidium shooting star orchid using protocorm-like bodies (PLBs). Aust. J. Crop Sci. 2013, 7, 1078–1084. [Google Scholar]
- Mubbarakh, S.A.; Rahmah, S.; Rahman, Z.A.; Sah, N.N.M.; Subramaniam, S. Cryopreservation of Brassidium Shooting Star orchid using the PVS3 method supported with preliminary histological analysis. Appl. Biochem. Biotechnol. 2014, 172, 1131–1145. [Google Scholar] [CrossRef]
- Negahdar, N.; Hashemabadi, D.; Kaviani, B. In vitro conservation and cryopreservation of Buxus sempervirens L., a critically endangered ornamental shrub. Rus. J. Plant Physiol. 2021, 68, 661–668. [Google Scholar] [CrossRef]
- Vettorazzi, R.G.; Silva Carvalho, V.; Carvalho Teixeira, M.; Campostrini, E.; Da Cunha, M.; de Monteiro Matos, E.; Facio Viccini, L. Cryopreservation of immature and mature seeds of Brazilian orchids of the genus Cattleya. Sci. Hortic. 2019, 256, 108603. [Google Scholar] [CrossRef]
- Maneerattanarungroj, P.; Bunnag, S.; Monthatong, M. In vitro conservation of Cleisostoma areitinum (Rchb.f.) Garay, rare Thai orchid species by an encapsulation-dehydration method. Asian J. Plant Sci. 2007, 6, 1235–1240. [Google Scholar]
- González-Benito, M.E.; Pérez, C.; Viviani, A.B. Cryopreservation of nodal explants of an endangered plant species (Centaurium rigualii Esteve) using the encapsulation-dehydration method. Biodiver. Conserv. 1997, 6, 583–590. [Google Scholar] [CrossRef]
- Gogoi, K.; Kumaria, S.; Tandon, P. Cryopreservation of Cymbidium eburneum Lindl. and C. hookerianum Rchb.f., two threatened and vulnerable orchids via encapsulation–dehydration. In Vitro Cell. Dev. Biol. Plant 2013, 49, 248–254. [Google Scholar] [CrossRef]
- Surenciski, M.R.; Flachsland, E.D.; Terada, G.; Mroginski, L.A.; Rey, H.Y. Cryopreservation of Cyrtopodium hatschbachii Pabst (Orchidaceae) immature seeds by encapsulation-dehydration. Biocell 2012, 36, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Thammasiri, K. Cryopreservation of some Thai orchid species. Acta Hort. 2008, 788, 53–62. [Google Scholar] [CrossRef]
- Pimda, W.; Bunnag, S. Cryopreservation of Dendrobium heterocarpum Lindl. via encapsulation-dehydration method. Elba Bioflux. 2010, 2, 7–14. [Google Scholar]
- Lurswijidjarus, W.; Thammasiri, K. Cryopreservation of shoot tips of Dendrobium Walter Oumae by encapsulation/dehydration. Sci. Asia 2004, 30, 293–299. [Google Scholar] [CrossRef]
- Thammasiri, K. Cryopreservation of seeds of a Thai orchid (Doritis pulcherrima Lindl.) by vitrification. CryoLetters 2000, 21, 237–244. [Google Scholar]
- Seydi, S.; Sedaghathoor, S.; Kaviani, B. In vitro storage techniques of Fritillaria imperialis Lubra Maxima, a wild rare and critically endangered ornamental species. CMU J. Nat. Sci. 2020, 19, 612–628. [Google Scholar] [CrossRef]
- Mikuła, A.; Tomiczak, K.; Rybczýnski, J.J. Cryopreservation enhances embryo-genic capacity of Gentiana cruciata (L.) suspension culture and maintains (epi)genetic uniformity of regenerants. Plant Cell Rep. 2011, 30, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Mikuła, A.; Fiuk, A.; Rybczýnski, J.J. Induction, maintenance and preservation of embryogenic competence of Gentiana cruciata L. cultures. Acta Biol. Crac. Bot. 2005, 47, 227–236. [Google Scholar]
- Bouman, H.; Tiekstra, A.; Petutschnig, E.; Homan, M.; Schreurs, R. Cryopreservation of Lilium species and cultivars. Acta Hortic. 2003, 612, 147–154. [Google Scholar] [CrossRef]
- Yin, Z.-F.; Bi, W.-L.; Chen, L.; Zhao, B.; Wang, Q.-C. An efficient, widely applicable cryopreservation of Lilium shoot tips by droplet-vitrification. Acta Physiol. Plant. 2014, 36, 1683–1692. [Google Scholar] [CrossRef]
- Yi, J.Y.; Lee, G.A.; Chung, J.W.; Lee, S.Y.; Lim, K.B. Efficient cryopreservation of Lilium spp. shoot tips using droplet-vitrification. Plant Breed. Biotech. 2013, 1, 131–136. [Google Scholar] [CrossRef]
- Folgado, R.; Panis, B. Cryopreservation of Ashe magnolia shoot-tips by droplet-vitrification. Acta Hortic. 2019, 1234, 233–240. [Google Scholar] [CrossRef]
- Lin, L.; Cai, L.; Fan, L.; Ma, J.-C.; Yang, X.-Y.; Hu, X.-J. Seed dormancy, germination and storage behavior of Magnolia sinica, a plant species with extremely small populations of Magnoliaceae. Plant Divers. 2022, 44, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Kesa, S.; Limpanavech, P.; Kongchuensin, S.; Chareonsap, P.; Juthamas, P. In vitro germplasm conservation of Magnolia sirindhorniae Noot. & Chalermglin in minimal growth condition and by cryopreservation. In Proceedings of the 18th Annual Meeting of the Thai Society for Biotechnology. Biotechnology: Benefits & Bioethics, Bangkok, Thailand, 2–3 November 2006. [Google Scholar]
- Miao, N.H.; Kaneko, Y.; Sugawara, Y. Ultrastructural implications of pretreatment for successful cryopreservation of Oncidium protocorm-like body. CryoLetters 2005, 26, 333–340. [Google Scholar] [PubMed]
- Flachsland, E.; Terada, G.; Scocchi, A.; Rey, H.; Mroginski, L.; Engelmann, F. Cryopreservation of seeds and in vitro-cultured protocorms of Oncidium bifolium sims. (Orchidaceae) by encapsulation-dehydration. CryoLetters 2006, 27, 235–242. [Google Scholar] [PubMed]
- Galdiano, R.F.J.; Lemos, E.G.M.; Faria, R.T.; Vendrame, W.A. Cryopreservation of Dendrobium hybrid seeds and protocorms as affected by phloroglucinol and supercool X1000. Sci. Hortic. 2012, 148, 154–160. [Google Scholar] [CrossRef]
- Hirano, T.; Godo, T.; Miyoshi, K.; Ishikawa, K.; Ishikawa, M.; Mii, M. Cryopreservation and low-temperature storage of seeds of Phaius tankervilleae. Plant Biotechnol. Rep. 2009, 3, 103–109. [Google Scholar] [CrossRef]
- Salaj, T.; Matusikova, I.; Fraterova, L.; Pirselova, B.; Salaj, J. Regrowth of embryogenic tissues of Pinus nigra following cryopreservation. Plant Cell Tissue Org. Cult. 2011, 106, 55–61. [Google Scholar] [CrossRef]
- Halmagyi, A.; Pinker, I. Plant regeneration from Rosa shoot tips cryopreserved by a combined droplet vitrification method. Plant Cell Tissue Org. Cult. 2006, 84, 100129–100137. [Google Scholar] [CrossRef]
- Pawłowska, B.; Bach, A. Cryopreservation by encapsulation-dehydration of in vitro grown shoot buds of Rosa ‘New Dawn’. Acta Hortic. 2011, 908, 303–308. [Google Scholar] [CrossRef]
- Jitsopakul, N.; Thammasiri, K.; Ishikawa, K. Cryopreservation of Vanda coerulea protocorm-like bodies by droplet-vitrification. In 1st International Symposium: Cryopreservation in Horticultural Species, Book of Abstract; ISHS: Leuven, Belgium, 2009; p. 92. [Google Scholar]
- Jitsopakul, N.; Thammasiri, K.; Ishikawa, K. Cryopreservation of Vanda coerulea protocorms by encapsulation-dehydration. CryoLetters 2008, 29, 253–260. [Google Scholar] [PubMed]
- Thammasiri, K.; Soamkul, L. Cryopreservation of Vanda coerulea Griff. ex Lindl. seeds by vitrification. Sci. Asia 2007, 33, 223–227. [Google Scholar] [CrossRef]
- González-Arnao, M.T.; Lazaro-Vallejo, C.E.; Engelmann, F.; Gamez-Pastrana, R.; Martinez-Ocampo, Y.M.; Pastelin-Solano, M.C.; Diaz-Ramos, C. Multiplication and cryopreservation of vanilla (Vanilla planifolia ‘Andrews’). In Vitro Cell. Dev. Biol.-Plant 2009, 45, 574–582. [Google Scholar] [CrossRef]
- Pathirana, R.; Mathew, L.; McLachlan, A. A simplified method for high recovery of kiwifruit (Actinidia spp.) shoot tips after droplet vitrification cryopreservation suitable for long-term conservation. Plant Cell Tissue Organ Cult. 2021, 144, 97–102. [Google Scholar] [CrossRef]
- Bachiri, Y.; Song, G.Q.; Plessis, P.; Shoar-Ghaffari, A.; Rekab, T.; Morisset, C. Routine cryopreservation of kiwifruit (Actinidia spp) germplasm by encapsulation-dehydration: Importance of plant growth regulators. Cryoletters 2001, 22, 61–74. [Google Scholar] [PubMed]
- Delgado-Aceves, L.; Portillo, L.; Folgado, R.; Romo-Paz, F.J.; González-Arnao, M.T. New approaches for micropropagation and cryopreservation of Agave peacockii, an endangered species. Plant Cell Tissue Organ Cult. 2022. [Google Scholar] [CrossRef]
- Delgado-Aceves, L.; González-Arnao, M.T.; Santacruz-Ruvalcaba, F.; Folgado, R.; Portillo, L. Indirect somatic embryogenesis and cryopreservation of Agave tequilana Weber cultivar ‘Chato’. Plants 2021, 10, 249. [Google Scholar] [CrossRef]
- Souza, F.V.D.; Kaya, E.; Vieira, L.J.; Souza, E.H.; Amorim, V.B.O.; Skogerboe, D.; Matsumoto, T.; Alves, A.A.C.; da Silva Ledo, C.A.; Jenderek, M.M. Droplet-vitrification and morphohistological studies of cryopreserved shoot tips of cultivated and wild pineapple genotypes. Plant Cell Tissue Org. Cult. 2016, 126, 351–360. [Google Scholar] [CrossRef]
- da Silva, R.L.; de Souza, E.H.; Vieira, L.d.J.; Pelacani, C.R.; Souza, F.V.D. Cryopreservation of pollen of wild pineapple accessions. Sci. Hortic. 2017, 219, 326–334. [Google Scholar] [CrossRef]
- Souza, F.V.D.; Kaya, E.; de Jesus Vieira, L.; da Silva Souza, A.; de Jesus da Silva Carvalho, M.; Barbosa Santos, E.; Cunha Alves, A.A.; Ellis, D. Cryopreservation of Hamilin sweet potato [(Citrus sinensis (L.) Osbeck)] embryonic calli using a modified aluminum cryo-plate technique. Sci. Hortic. 2017, 224, 302–305. [Google Scholar] [CrossRef]
- Volk, G.M.; Bonnart, R.; Shepherd, A.; Yin, Z.F.; Lee, L.; Polek, M.; Krueger, R. Citrus cryopreservation: Viability of diverse taxa and histological observations. Plant Cell Tissue Org. Cult. 2017, 128, 327–334. [Google Scholar] [CrossRef]
- Wilms, H.; Rhee, J.H.; Rivera, R.L.; Longin, K.; Panis, B. Developing coconut cryopreservation protocols and establishing cryogenebank at RDA; a collaborative project between RDA and biodiversity international. Acta Hortic. 2019, 1234, 343–348. [Google Scholar] [CrossRef]
- Niu, Y.L.; Luo, Z.R.; Zhang, Y.F.; Zhang, Q.L. Cryopreservation of in vitro grown shoot tips of Diospyros kaki thunb. using different methods. CryoLetters 2012, 33, 69–74. [Google Scholar]
- Matsumoto, T.; Niino, T.; Shirata, K.; Kurahashi, T.; Matsumoto, S.; Maki, S. Long-term conservation of Diospyros germplasm using dormant buds by a prefreezing method. Plant Biotechnol. 2004, 21, 229–232. [Google Scholar] [CrossRef]
- Ai, P.; Luo, Z. Cryopreservation of dormant vegetative buds and genetic stability analysis of regenerated plantlets in persimmon. Acta Hortic. 2005, 685, 85–92. [Google Scholar] [CrossRef]
- Benelli, C.; De Carlo, A.; Giordani, E.; Pecchioli, S.; Bellini, E.; Kochanova, Z. Vitrification/one-step freezing procedure for cryopreservation of persimmon dormant buds. Acta Hortic. 2009, 833, 163–168. [Google Scholar] [CrossRef]
- Ai, P.; Luo, Z. Cryopreservation of dormant shoot-tips of persimmon by vitrification and plant regeneration. Sci. Agric. Sin. 2003, 36, 553–556, (In Chinese with English Summary). [Google Scholar]
- Matsumoto, T.; Mochida, K.; Itamura, H.; Sakai, A. Cryopreservation of persimmon (Diospyros kaki Thunb.) by vitrification of dormant shoot tips. Plant Cell Rep. 2001, 20, 398–402. [Google Scholar] [CrossRef]
- Panis, B.; Withers, L.A.; De Langhe, E. Cryopreservation of Musa suspension cultures and subsequent regeneration of plants. CryoLetters 1990, 11, 337–350. [Google Scholar]
- Generoso, A.L.; Carvalho, V.S.; Walter, R.; Campbell, G.; da Silva Araújo, L.; Silva Santana, G.; Cunha, M. Mature-embryo culture in the cryopreservation of passion fruit (Passiflora edulis Sims) seeds. Sci. Hort. 2019, 256, 108638. [Google Scholar] [CrossRef]
- Garcia, R.O.; Pacheco, G.; Vianna, M.G.; Mansur, E. In vitro conservation of Passiflora suberosa L., slow and cryopreservation. CryoLetters 2011, 32, 377–388. [Google Scholar] [PubMed]
- Merhy, T.S.M.; Vianna, M.G.; Garcia, R.O.; Pacheco, G.; Mansur, E. Cryopreservation of Passiflora pohblii nodal segments and assessment of genetic stability of regenerated plants. CryoLetters 2014, 35, 204–215. [Google Scholar]
- O’Brien, C.; Hiti-Bandaralage, J.; Folgado, R.; Lahmeyer, S.; Hayward, A.; Folsom, J.; Mitter, N. A method to increase regrowth of vitrified shoot tips of avocado (Persea americana Mill.): First critical step in developing a cryopreservation protocol. Sci. Hortic. 2020, 266, 109305. [Google Scholar] [CrossRef]
- O’Brien, C.; Parisi, A.; Lim Yiing Yuan, J.; Constantin, M.; Mitter, N. Cryopreservation of avocado (Persea americana Mill.) using somatic embryos. Acta Hortic. 2016, 1113, 265–270. [Google Scholar] [CrossRef]
- Towill, L.E.; Forsline, P.L. Cryopreservation of sour cherry (Prunus ceraceus L.) using a dormant vegetative bud method. CryoLetters 1999, 20, 215–222. [Google Scholar]
- Shatnawi, M.A.; Shibli, R.; Qrunfleh, I.; Bataeineh, K.; Obeidat, M. In vitro propagation and cryopreservation of Prunus avium using vitrification and encapsulation dehydration methods. J. Food Agric. Environ. 2007, 5, 204–208. [Google Scholar]
- Channuntapipat, C.; Collins, G.; Bertozzi, T.; Sedgley, M. Cryopreservation of in vitro almond shoot tips by vitrification. J. Hort. Sci. Biotech. 2000, 75, 228–232. [Google Scholar] [CrossRef]
- De Boucaud, M.T.; Brison, M.; Helliot, B.; Herve-Paulus, V. Cryopreservation in Prunus. In Cryopreservation of Plant Germplasm II—Biotechnology in Agriculture and Forestry; Towill, L.E., Bajaj, Y.P.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 287–311. [Google Scholar]
- Hao, Y.J.; Cheng, Y.J.; Deng, X. Stable maintenance and expression of a foreign gene in transgenic pear shoots retrieved from in vitro conservation. J. Plant Physiol. 2005, 162, 237–243. [Google Scholar] [CrossRef]
- Wang, B.; Yin, Z.-F.; Feng, C.-H.; Shi, X.; Li, Y.-P.; Wang, Q.-C. Cryopreservation of potato shoot tips. In Potato I Fruit, Vegetable and Cereal Science and Biotechnology, 2 (Special Issue 1); Benkeblia, N., Tennant, P., Eds.; Global Science Book: London, UK, 2008; pp. 46–53. [Google Scholar]
- Chang, Y.; Reed, B.M. Preculture conditions influence cold hardiness and regrowth of Pyrus cordata shoot tips after cryopreservation. HortScience 2001, 36, 1329–1333. [Google Scholar] [CrossRef]
- Chang, Y.; Reed, B.M. Extended alternating-temperature cold acclimation and culture duration improve pear shoot cryopreservation. Cryobiology 2000, 40, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Wang, M.R.; Wang, Q.C. ROS-induced oxidative stress in plant cryopreservation: Occurrence and alleviation. Planta 2021, 254, 124. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A. Gene ontology consortium. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Morrison, N.; Cochrane, G.; Faruque, N.; Tatusova, T.; Tateno, Y.; Hancock, D.; Field, D. Concept of sample in omics technology. OMICS J. Integr. Biol. 2006, 10, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Harding, K.; Benson, E.E. Biomarkers from molecules to ecosystems and biobanks to genebanks. In Conservation of Tropical Plant Species; Normah, N.M., Chin, H.F., Reed, B.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Volk, G.M. Application of functional genomics and proteomics to plant cryopreservation. Curr. Genom. 2010, 11, 24–29. [Google Scholar] [CrossRef]
- Walters, C.; Wheeler, L.; Stanwood, P.C. Longevity of cryogenically stored seeds. Cryobiology 2004, 48, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.R.; Bi, W.; Shukla, M.R.; Ren, L.; Hamborg, Z.; Blystad, D.R.; Saxena, P.K.; Wang, Q.C. Epigenetic and genetic integrity, metabolic stability, and field performance of cryopreserved plants. Plants 2021, 10, 1889. [Google Scholar] [CrossRef]
- Martinez-Montero, M.E.; González-Arnao, M.T.; Engelmann, F. Cryopreservation of tropical plant germplasm with vegetative propagation—Review of sugarcane (Saccharum spp.) and pineapple (Ananas comosus (L.) Merrill) cases. In Current Frontiers in Cryopreservation; Katkov, I.I., Ed.; In Tech: Rijeka, Croatia, 2012. [Google Scholar]
- Panis, B.; Van den Houwe, I.; Swennen, R.; Rhee, J.; Roux, N. Securing plant genetic resources for perpetuity through cryopreservation. Indian J. Plant Genet. Resour. 2016, 29, 300–302. [Google Scholar] [CrossRef]
- Seki, M.; Narusaka, M.; Ishida, J. Monitoring the expression profiles of 7000 Arabidopsis genes under drought, cold, and high-salinity stresses using a full length cDNA microarray. Plant J. 2002, 31, 279–292. [Google Scholar] [CrossRef]
- Takagi, H. Recent development in cryopreservation of shoot apices of tropical species. In Cryopreservation of Tropical Plant Germplasm Progress and Application; Engelmann, F., Takai, H., Eds.; JIRCAS/IPGRI Publication: Tsukuba, Japan, 2000; pp. 178–193. [Google Scholar]
- Fowler, S.; Thomashow, M.F. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 2002, 14, 1675–1690. [Google Scholar] [CrossRef]
- Kim, C.Y.; Liu, Y.; Thorne, E.T.; Yang, H.; Fukushige, H.; Gassmann, W.; Hildebrand, D.; Sharp, R.E.; Zhang, S. Activation of a stress-responsive mitogen activated protein kinase cascade induces the biosynthesis of ethylene in plants. Plant Cell 2003, 15, 2707–2718. [Google Scholar] [CrossRef]
- Lynch, P.T.; Siddika, A.; Johnston, J.W.; Mehra, A.; Benelli, C.; Lambardi, M.; Benson, E.E. Effects of osmotic pretreatments on oxidative stress and antioxidant profiles of cryopreserved olive somatic embryos. Plant Sci. 2011, 181, 47–56. [Google Scholar] [CrossRef]
- Grapin, A.; Dorion, N.; Verdeil, J.L.; Escoute, J. Histo-cytological changes in Pelargonium apices during the cryopreservation process: Effect of the osmotic agent chosen for the preculture step. Acta Hortic. 2007, 760, 195–201. [Google Scholar] [CrossRef]
- Gamez-Pastrana, R.; Gonzalez-Arnao, M.T.; Martinez-Ocampo, Y.; Engelmann, F. Thermal events in calcium alginate beads during encapsulation dehydration and encapsulation-vitrification protocols. Acta Hortic. 2011, 908, 47–54. [Google Scholar] [CrossRef]
- Yang, Z.; Sheng, J.; Lv, K.; Ren, L.; Zhang, D. Y2SK2 and SK3 type dehydrins from Agapanthus praecox can improve plant stress tolerance and act as multifunctional protectants. Plant Sci. 2019, 284, 143–160. [Google Scholar] [CrossRef]
- Chen, G.Q.; Ren, L.; Zhang, D.; Shen, X.H. Glutathione improves survival of cryopreserved embryogenic calli of Agapanthus praecox subsp. orientalis. Acta Physiol. Plant. 2016, 38, 250. [Google Scholar] [CrossRef]
- Di, W.; Jiang, X.; Xu, J.; Jia, M.; Li, B.; Liu, Y. Stress and damage mechanisms in Dendrobium nobile Lindl. protocorm-like bodies during pre- and post-liquid nitrogen exposure in cryopreservation revealed by iTRAQ proteomic analysis. In Vitro Cell. Dev. Biol. Plant 2018, 54, 253–272. [Google Scholar] [CrossRef]
- Jiang, X.; Ren, R.; Di, W.; Jia, M.; Li, Z.; Liu, Y.; Gao, R. Hydrogen peroxide and nitric oxide are involved in programmed cell death induced by cryopreservation in Dendrobium protocorm-like bodies. Plant Cell Tissue Organ Cult. 2019, 137, 553–563. [Google Scholar] [CrossRef]
- Poobathy, R.; Sinniah, U.R.; Xavier, R.; Subramaniam, S. Catalase and superoxide dismutase activities and the total protein content of protocorm-like bodies of Dendrobium Sonia-28 subjected to vitrification. Appl. Biochem. Biotechnol. 2013, 170, 1066–1079. [Google Scholar] [CrossRef]
- Rahmah, S.; Mubbarakh, A.; Ping, S.; Subramaniam, K.S. Effects of droplet-vitrification cryopreservation based on physiological and antioxidant enzyme activities of Brassidium shooting star orchid. Sci. World J. 2015, 961793, 1–10. [Google Scholar] [CrossRef]
- Antony, J.J.J.; Zakaria, S.; Zakaria, R.; Ujang, J.A.; Othman, N.; Subramaniam, S. Biochemical analyses of Dendrobium Sabin Blue PLBs during cryopreservation by vitrification. Physiol. Mol. Biol. Plant 2019, 25, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Vianna, M.G.; Garcia, R.O.; Mansur, E.; Engelmann, F.; Pacheco, G. Oxidative stress during the cryopreservation of Passiflora suberosa L. shoot tips using the V-Cryo-plate technique: Determination of the critical stages of the protocol. Plant Cell Tissue Organ Cult. 2019, 139, 369–379. [Google Scholar] [CrossRef]
- Johnston, J.W.; Benson, E.E.; Harding, K. Cryopreservation of in vitro Ribes shoots induces temporal changes in DNA methylation. Plant Physiol. Biochem. 2009, 47, 123–131. [Google Scholar] [CrossRef]
- Castillo, N.R.F.; Bassil, N.V.; Wada, S.; Reed, B.M. Genetic stability of cryopreserved shoot tips of Rubus germplasm. In Vitro Cell. Dev. Biol. Plant 2010, 46, 246–256. [Google Scholar] [CrossRef]
- Maki, S.; Hirai, Y.; Niino, T.; Matsumoto, T. Assessment of molecular genetic stability between long-term cryopreserved and tissue cultured wasabi (Wasabia japonica) plants. CryoLetters 2015, 36, 318–324. [Google Scholar]
- Li, J.-W.; Ozudogru, E.A.; Li, J.; Wang, M.-R.; Bi, W.-L.; Lambardi, M.; Wang, Q.-C. Cryobiotechnology of forest trees: Recent advances and future prospects. Biodivers. Conserv. 2017, 27, 795–814. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2016. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaviani, B.; Kulus, D. Cryopreservation of Endangered Ornamental Plants and Fruit Crops from Tropical and Subtropical Regions. Biology 2022, 11, 847. https://doi.org/10.3390/biology11060847
Kaviani B, Kulus D. Cryopreservation of Endangered Ornamental Plants and Fruit Crops from Tropical and Subtropical Regions. Biology. 2022; 11(6):847. https://doi.org/10.3390/biology11060847
Chicago/Turabian StyleKaviani, Behzad, and Dariusz Kulus. 2022. "Cryopreservation of Endangered Ornamental Plants and Fruit Crops from Tropical and Subtropical Regions" Biology 11, no. 6: 847. https://doi.org/10.3390/biology11060847
APA StyleKaviani, B., & Kulus, D. (2022). Cryopreservation of Endangered Ornamental Plants and Fruit Crops from Tropical and Subtropical Regions. Biology, 11(6), 847. https://doi.org/10.3390/biology11060847