
Model-Guided Metabolic Rewiring for Gamma-Aminobutyric Acid and Butyrolactam Biosynthesis in Corynebacterium glutamicum ATCC13032

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Media and Cultivations
2.3. Genetic Manipulation
2.4. Constraint-Based Metabolic Flux Analysis
2.5. Analytical Methods
2.6. SDS-PAGE Analysis
2.7. RNA Preparation and Quantitative Real-Time RT-PCR
3. Results
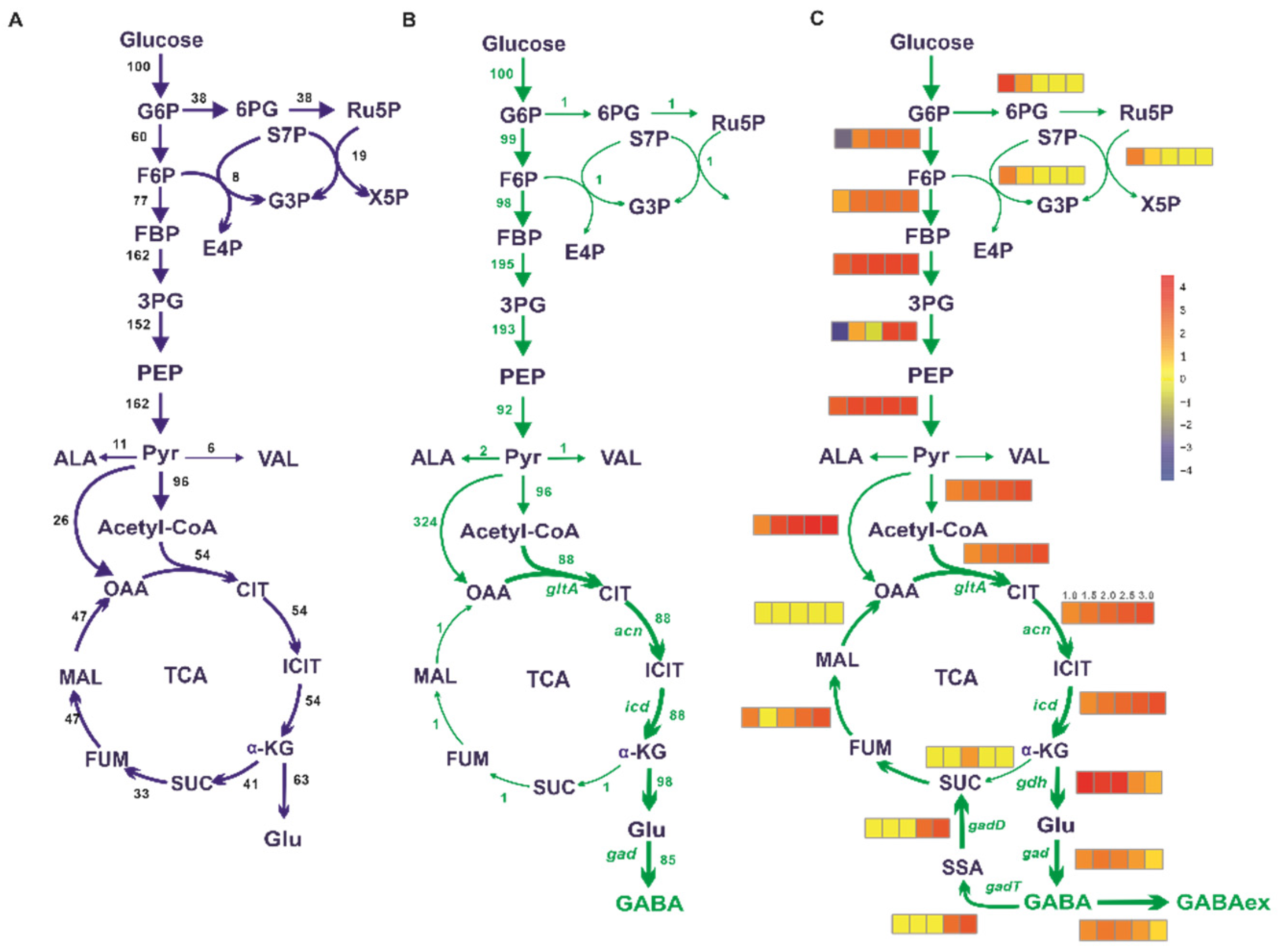
3.1. In Silico Simulation of Flux Distribution for GABA Overproduction
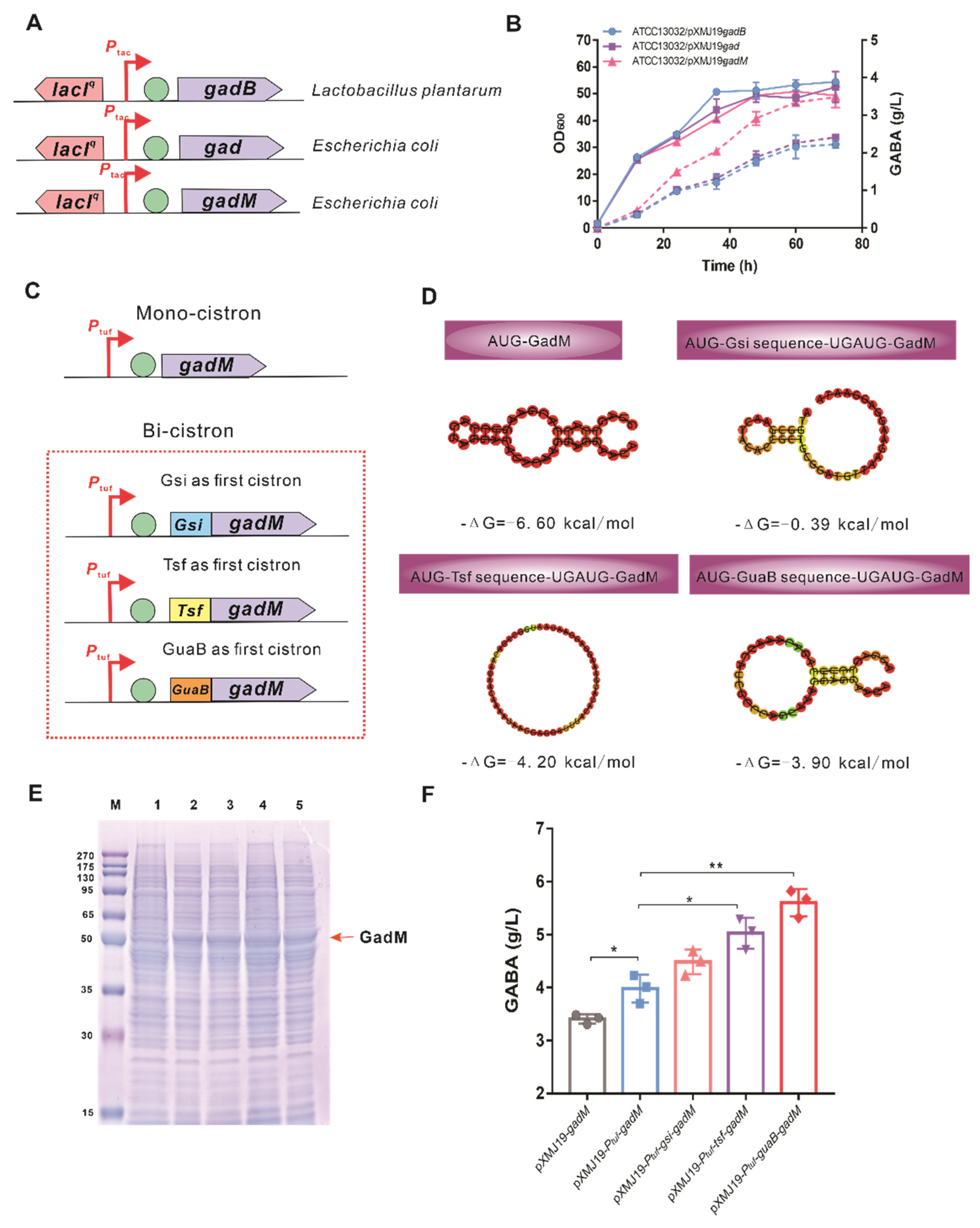
3.2. Constructing a GABA Biosynthetic Pathway by a Bi-Cistronic Expression Cassette
3.3. Reprogramming TCA Cycle to Improve GABA Production
3.4. Blocking Decomposition and Rewiring Transport of GABA
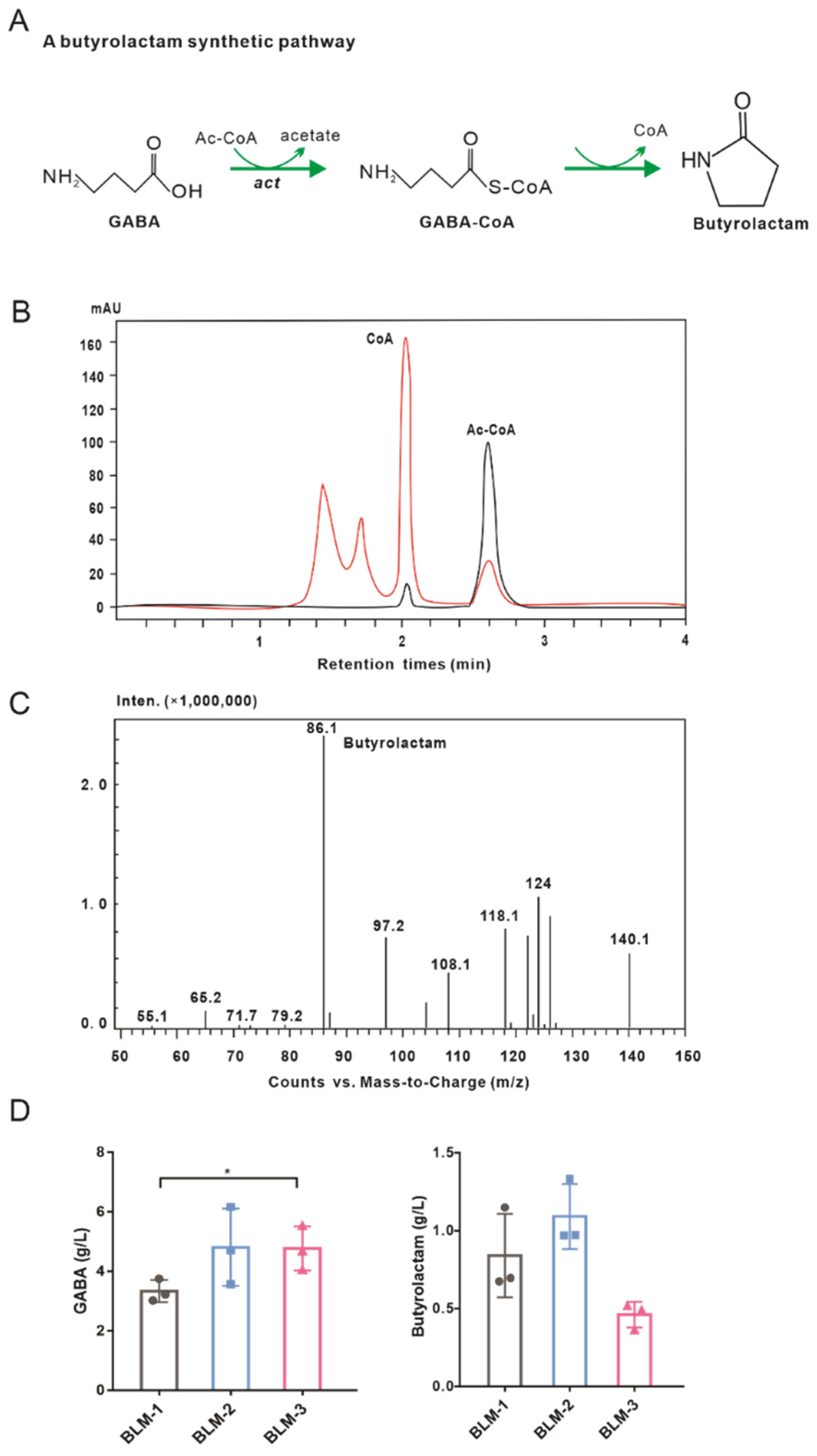
3.5. Construction of a Metabolic Pathway for Butyrolactam Biosynthesis
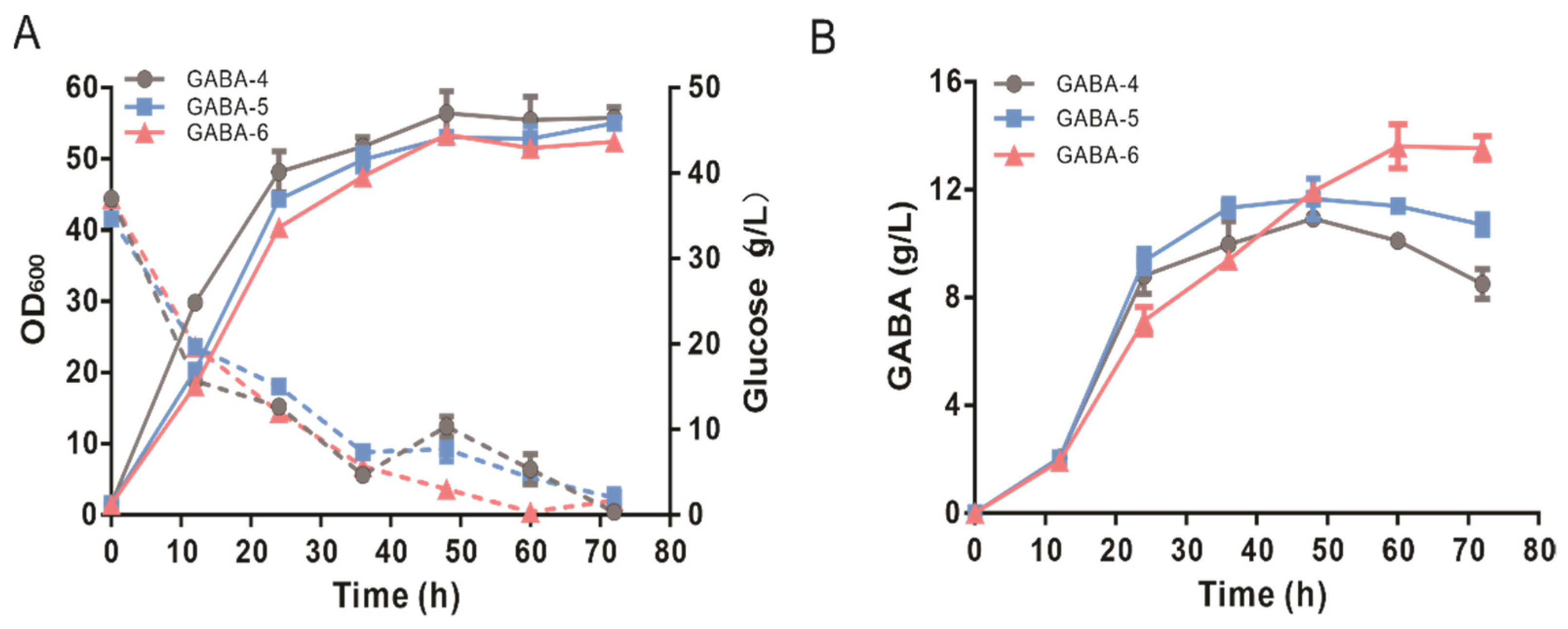
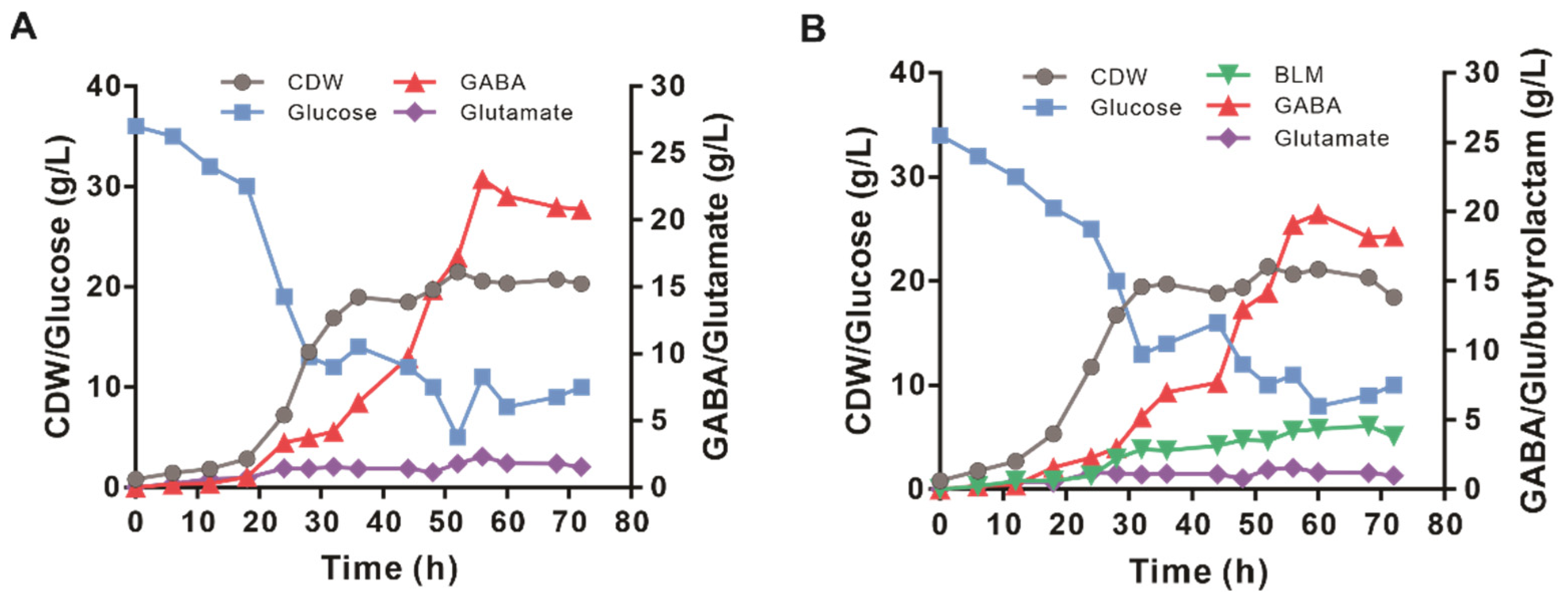
3.6. The Performance of Engineered GABA-6 and BLM-2 Strains in Fed-Batch Fermentation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pham, J.V.; Yilma, M.A.; Feliz, A.; Majid, M.T.; Maffetone, N.; Walker, J.R.; Kim, E.; Cho, H.J.; Reynolds, J.M.; Song, M.C.; et al. A review of the microbial production of bioactive natural products and biologics. Front. Microbiol. 2019, 10, 1404. [Google Scholar]
- Tsuge, Y.; Kawaguchi, H.; Sasaki, K.; Kondo, A. Engineering cell factories for producing building block chemicals for bio-polymer synthesis. Microb. Cell Fact. 2016, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Tsuge, Y.; Matsuzawa, H. Recent progress in production of amino acid-derived chemicals using Corynebacterium glutamicum. World J. Microbiol. Biotechnol. 2021, 37, 49. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhou, L.; Tian, K.; Kumar, A.; Singh, S.; Prior, B.A.; Wang, Z. Metabolic engineering of Escherichia coli: A sustainable industrial platform for bio-based chemical production. Biotechnol. Adv. 2013, 31, 1200–1223. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Yang, J.E.; Ha, J.Y.; Chae, T.U.; Shin, J.H.; Gustavsson, M.; Lee, S.Y. Bio-based production of monomers and polymers by metabolically engineered microorganisms. Curr. Opin. Biotechnol. 2015, 36, 73–84. [Google Scholar] [CrossRef]
- Lee, J.Y.; Na, Y.A.; Kim, E.; Lee, H.S.; Kim, P. The actinobacterium Corynebacterium glutamicum, an industrial workhorse. J. Microbiol. Biotechnol. 2016, 26, 807–822. [Google Scholar] [CrossRef]
- Becker, J.; Giesselmann, G.; Hoffmann, S.L.; Wittmann, C. Corynebacterium glutamicum for sustainable bioproduction: From metabolic physiology to systems metabolic engineering. Adv. Biochem. Eng. Biotechnol. 2018, 162, 217–263. [Google Scholar] [PubMed]
- Wang, B.; Hu, Q.; Zhang, Y.; Shi, R.; Chai, X.; Liu, Z.; Shang, X.; Zhang, Y.; Wen, T. A RecET-assisted CRISPR–Cas9 genome editing in Corynebacterium glutamicum. Microb. Cell Factories 2018, 17, 63. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cai, J.; Shang, X.; Wang, B.; Liu, S.; Chai, X.; Tan, T.; Zhang, Y.; Wen, T. A new genome-scale metabolic model of Corynebacterium glutamicum and its application. Biotechnol. Biofuels 2017, 10, 169. [Google Scholar] [CrossRef]
- Shang, X.; Chai, X.; Lu, X.; Li, Y.; Zhang, Y.; Wang, G.; Zhang, C.; Liu, S.; Zhang, Y.; Ma, J.; et al. Native promoters of Corynebacterium glutamicum and its application in l-lysine production. Biotechnol. Lett. 2018, 40, 383–391. [Google Scholar] [CrossRef]
- Zhang, Y.; Shang, X.; Lai, S.; Zhang, Y.; Hu, Q.; Chai, X.; Wang, B.; Liu, S.; Wen, T. Reprogramming one-carbon metabolic pathways to decouple l-serine catabolism from cell growth in Corynebacterium glutamicum. ACS Synth. Biol. 2018, 7, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhou, N.; Liu, Y.M.; Liu, C.; Lou, C.B.; Jiang, C.Y.; Liu, S.J. Ribosome binding site libraries and pathway modules for shikimic acid synthesis with Corynebacterium glutamicum. Microb. Cell Fact. 2015, 14, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Liu, D.; Mao, Z.; Mao, Y.; Ma, H.; Chen, T.; Zhao, X.; Wang, Z. Model-based reconstruction of synthetic promoter library in Corynebacterium glutamicum. Biotechnol. Lett. 2018, 40, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Rytter, J.V.; Helmark, S.; Chen, J.; Lezyk, M.J.; Solem, C.; Jensen, P.R. Synthetic promoter libraries for Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2014, 98, 2617–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Gao, X.; Zhao, Z.; Li, A.; Wang, Y.; Yang, Y.; Liu, X.; Bai, Z. Enhanced production of recombinant proteins in Corynebacterium glutamicum by constructing a bicistronic gene expression system. Microb. Cell Fact. 2020, 19, 113. [Google Scholar] [CrossRef]
- Jiang, Y.; Qian, F.; Yang, J.; Liu, Y.; Dong, F.; Xu, C.; Sun, B.; Chen, B.; Xu, X.; Li, Y.; et al. CRISPR-Cpf1 assisted genome editing of Corynebacterium glutamicum. Nat. Commun. 2017, 8, 15179. [Google Scholar] [CrossRef] [Green Version]
- Abdou, A.M.; Higashiguchi, S.; Horie, K.; Kim, M.; Hatta, H.; Yokogoshi, H. Relaxation and immunity enhancement effects of gamma-aminobutyric acid (GABA) administration in humans. Biofactors 2006, 26, 201–208. [Google Scholar] [CrossRef]
- Ge, S.; Goh, E.L.; Sailor, K.A.; Kitabatake, Y.; Ming, G.L.; Song, H. GABA regulates synaptic integration of newly generated neurons in the adult brain. Nature 2006, 439, 589–593. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, K.; Kimura, M.; Kasaha, K.; Matsumoto, K.; Sansawa, H.; Yamori, Y. Effect of a gamma-aminobutyric acid-enriched dairy product on the blood pressure of spontaneously hypertensive and normotensive Wistar-Kyoto rats. Br. J. Nutr. 2004, 92, 411–417. [Google Scholar] [CrossRef]
- Park, S.J.; Kim, E.Y.; Noh, W.; Oh, Y.H.; Kim, H.Y.; Song, B.K.; Cho, K.M.; Hong, S.H.; Lee, S.H.; Jegal, J. Synthesis of nylon 4 from gamma-aminobutyrate (GABA) produced by recombinant Escherichia coli. Bioprocess. Biosyst. Eng. 2013, 36, 885–892. [Google Scholar] [CrossRef]
- Takahashi, C.; Shirakawa, J.; Tsuchidate, T.; Okai, N.; Hatada, K.; Nakayama, H.; Tateno, T.; Ogino, C.; Kondo, A. Robust production of gamma-amino butyric acid using recombinant Corynebacterium glutamicum expressing glutamate decarboxylase from Escherichia coli. Enzyme Microb. Technol. 2012, 51, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Rao, Z.; Li, X.; Xu, M.; Zhang, X.; Xu, Z. One-step fermentation converting glucose in gamma-aminobutyric acid by a recombinant Corynebacterium crenatum. Acta Microbiol. Sin. 2013, 53, 817–824. [Google Scholar]
- Yang, T.; Rao, Z.; Kimani, B.G.; Xu, M.; Zhang, X.; Yang, S.T. Two-step production of gamma-aminobutyric acid from cassava powder using Corynebacterium glutamicum and Lactobacillus plantarum. J. Ind. Microbiol. Biotechnol. 2015, 42, 1157–1165. [Google Scholar] [CrossRef]
- Zhao, A.; Hu, X.; Wang, X. Metabolic engineering of Escherichia coli to produce gamma-aminobutyric acid using xylose. Appl. Microbiol. Biotechnol. 2017, 101, 3587–3603. [Google Scholar] [CrossRef]
- Ke, C.; Yang, X.; Rao, H.; Zeng, W.; Hu, M.; Tao, Y.; Huang, J. Whole-cell conversion of l-glutamic acid into gamma-aminobutyric acid by metabolically engineered Escherichia coli. Springerplus 2016, 5, 591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.Y.; Kim, S.K.; Ra, C.H. Evaluation of gamma-aminobutyric acid (GABA) production by Lactobacillus plantarum using two-step fermentation. Bioprocess. Biosyst. Eng. 2021, 44, 2099–2108. [Google Scholar] [CrossRef]
- Choi, J.W.; Yim, S.S.; Lee, S.H.; Kang, T.J.; Park, S.J.; Jeong, K.J. Enhanced production of gamma-aminobutyrate (GABA) in recombinant Corynebacterium glutamicum by expressing glutamate decarboxylase active in expanded pH range. Microb. Cell Fact. 2015, 14, 21. [Google Scholar] [CrossRef] [Green Version]
- Shi, F.; Xie, Y.; Jiang, J.; Wang, N.; Li, Y.; Wang, X. Directed evolution and mutagenesis of glutamate decarboxylase from Lactobacillus brevis Lb85 to broaden the range of its activity toward a near-neutral pH. Enzyme Microb. Technol. 2014, 61–62, 35–43. [Google Scholar] [CrossRef]
- Thu Ho, N.A.; Hou, C.Y.; Kim, W.H.; Kang, T.J. Expanding the active pH range of Escherichia coli glutamate decarboxylase by breaking the cooperativeness. J. Biosci. Bioeng. 2013, 115, 154–158. [Google Scholar] [CrossRef]
- Shi, F.; Jiang, J.; Li, Y.; Li, Y.; Xie, Y. Enhancement of gamma-aminobutyric acid production in recombinant Corynebacterium glutamicum by co-expressing two glutamate decarboxylase genes from Lactobacillus brevis. J. Ind. Microbiol. Biotechnol. 2013, 40, 1285–1296. [Google Scholar] [CrossRef]
- Wang, N.; Ni, Y.; Shi, F. Deletion of odhA or pyc improves production of gamma-aminobutyric acid and its precursor l-glutamate in recombinant Corynebacterium glutamicum. Biotechnol. Lett. 2015, 37, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Luan, M.; Li, Y. Ribosomal binding site sequences and promoters for expressing glutamate decarboxylase and producing gamma-aminobutyrate in Corynebacterium glutamicum. AMB Express 2018, 8, 61. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Shi, F.; Wang, X. Chromosomal editing of Corynebacterium glutamicum ATCC 13032 to produce gamma-aminobutyric acid. Biotechnol. Appl. Biochem. 2022. [Google Scholar] [CrossRef] [PubMed]
- Jorge, J.M.; Leggewie, C.; Wendisch, V.F. A new metabolic route for the production of gamma-aminobutyric acid by Corynebacterium glutamicum from glucose. Amino Acids 2016, 48, 2519–2531. [Google Scholar] [CrossRef] [PubMed]
- Okai, N.; Takahashi, C.; Hatada, K.; Ogino, C.; Kondo, A. Disruption of pknG enhances production of gamma-aminobutyric acid by Corynebacterium glutamicum expressing glutamate decarboxylase. AMB Express 2014, 4, 20. [Google Scholar] [CrossRef] [Green Version]
- Jorge, J.M.; Nguyen, A.Q.; Perez-Garcia, F.; Kind, S.; Wendisch, V.F. Improved fermentative production of gamma-aminobutyric acid via the putrescine route: Systems metabolic engineering for production from glucose, amino sugars, and xylose. Biotechnol. Bioeng. 2017, 114, 862–873. [Google Scholar] [CrossRef]
- Shi, F.; Zhang, M.; Li, Y. Overexpression of ppc or deletion of mdh for improving production of gamma-aminobutyric acid in recombinant Corynebacterium glutamicum. World J. Microbiol. Biotechnol. 2017, 33, 122. [Google Scholar] [CrossRef]
- Ni, Y.; Shi, F.; Wang, N. Specific gamma-aminobutyric acid decomposition by gabP and gabT under neutral pH in recombinant Corynebacterium glutamicum. Biotechnol. Lett. 2015, 37, 2219–2227. [Google Scholar] [CrossRef]
- Wen, J.; Bao, J. Improved fermentative gamma-aminobutyric acid production by secretory expression of glutamate decarboxylase by Corynebacterium glutamicum. J. Biotechnol. 2021, 331, 19–25. [Google Scholar] [CrossRef]
- Baritugo, K.A.; Kim, H.T.; David, Y.; Khang, T.U.; Hyun, S.M.; Kang, K.H.; Yu, J.H.; Choi, J.H.; Song, J.J.; Joo, J.C.; et al. Enhanced production of gamma-aminobutyrate (GABA) in recombinant Corynebacterium glutamicum strains from empty fruit bunch biosugar solution. Microb. Cell Fact. 2018, 17, 129. [Google Scholar] [CrossRef]
- Wei, L.; Zhao, J.; Wang, Y.; Gao, J.; Du, M.; Zhang, Y.; Xu, N.; Du, H.; Ju, J.; Liu, Q.; et al. Engineering of Corynebacterium glutamicum for high-level gamma-aminobutyric acid production from glycerol by dynamic metabolic control. Metab. Eng. 2022, 69, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Tokiwa, Y.; Calabia, B.P.; Ugwu, C.U.; Aiba, S. Biodegradability of plastics. Int. J. Mol. Sci. 2009, 10, 3722–3742. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kao, E.; Wang, G.; Baidoo, E.E.K.; Chen, M.; Keasling, J.D. Metabolic engineering of Escherichia coli for the biosynthesis of 2-pyrrolidone. Metab. Eng. Commun. 2016, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chae, T.U.; Ko, Y.S.; Hwang, K.S.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of four-, five- and six-carbon lactams. Metab. Eng. 2017, 41, 82–91. [Google Scholar] [CrossRef]
- Zhang, J.; Barajas, J.F.; Burdu, M.; Wang, G.; Baidoo, E.E.; Keasling, J.D. Application of an acyl-coa ligase from Streptomyces aizunensis for lactam biosynthesis. ACS Synth. Biol. 2017, 6, 884–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeom, S.J.; Kim, M.; Kwon, K.K.; Fu, Y.; Rha, E.; Park, S.H.; Lee, H.; Kim, H.; Lee, D.H.; Kim, D.M.; et al. A synthetic microbial biosensor for high-throughput screening of lactam biocatalysts. Nat. Commun. 2018, 9, 5053. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Wang, W.; Deng, A.; Sun, Z.; Zhang, Y.; Liang, Y.; Che, Y.; Wen, T. A mimicking-of-DNA-methylation-patterns pipeline for overcoming the restriction barrier of bacteria. PLoS Genet. 2012, 8, e1002987. [Google Scholar] [CrossRef] [Green Version]
- Jakoby, M.; Ngouoto-Nkili, C.; Burkovski, A. Construction and application of new Corynebacterium glutamicum vectors. Biotechnol. Tech. 1999, 13, 437–441. [Google Scholar] [CrossRef]
- Schafer, A.; Tauch, A.; Jager, W.; Kalinowski, J.; Thierbach, G.; Puhler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Cremer, J.; Eggeling, L.; Sahm, H. Control of the Lysine Biosynthesis Sequence in Corynebacterium glutamicum as analyzed by overexpression of the individual corresponding genes. Appl. Environ. Microbiol. 1991, 57, 1746–1752. [Google Scholar] [CrossRef] [Green Version]
- Koffas, M.A.G.; Jung, G.Y.; Aon, J.C.; Stephanopoulos, G. Effect of pyruvate carboxylase overexpression on the physiology of Corynebacterium glutamicum. Appl. Environ. Microbiol. 2002, 68, 5422–5428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugan, R.E.; Schmidt, M.J.; Hoganson, G.E.; Steele, J.; Gilles, B.A.; Shug, A.L. High-performance liquid chromatography of coenzyme A esters formed by transesterification of short-chain acylcarnitines: Diagnosis of acidemias by urinary analysis. Anal. Biochem. 1987, 160, 275–280. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Bott, M. Offering surprises: TCA cycle regulation in Corynebacterium glutamicum. Trends Microbiol. 2007, 15, 417–425. [Google Scholar] [CrossRef]
- Han, S.O.; Inui, M.; Yukawa, H. Effect of carbon source availability and growth phase on expression of Corynebacterium glutamicum genes involved in the tricarboxylic acid cycle and glyoxylate bypass. Microbiology 2008, 154, 3073–3083. [Google Scholar] [CrossRef] [Green Version]
- Roy, V.; Roth, R.; Berge, M.; Chitta, R.; Vajrala, S.; Kuntumalla, S.; Schmelzer, A.E.; Schoner, R. A bicistronic vector with destabilized mRNA secondary structure yields scalable higher titer expression of human neurturin in E. coli. Biotechnol. Bioeng. 2017, 114, 1753–1761. [Google Scholar] [CrossRef]
- Bentele, K.; Saffert, P.; Rauscher, R.; Ignatova, Z.; Bluthgen, N. Efficient translation initiation dictates codon usage at gene start. Mol. Syst Biol. 2013, 9, 675. [Google Scholar] [CrossRef]
- Goodman, D.B.; Church, G.M.; Kosuri, S. Causes and effects of N-terminal codon bias in bacterial genes. Science 2013, 342, 475–479. [Google Scholar] [CrossRef] [Green Version]
- Osterman, I.A.; Evfratov, S.A.; Sergiev, P.V.; Dontsova, O.A. Comparison of mRNA features affecting translation initiation and reinitiation. Nucleic Acids Res. 2013, 41, 474–486. [Google Scholar] [CrossRef] [Green Version]
- Kind, S.; Becker, J.; Wittmann, C. Increased lysine production by flux coupling of the tricarboxylic acid cycle and the lysine biosynthetic pathway--metabolic engineering of the availability of succinyl-CoA in Corynebacterium glutamicum. Metab. Eng. 2013, 15, 184–195. [Google Scholar] [CrossRef]
- Theodosiou, E.; Breisch, M.; Julsing, M.K.; Falcioni, F.; Buhler, B.; Schmid, A. An artificial TCA cycle selects for efficient alpha-ketoglutarate dependent hydroxylase catalysis in engineered Escherichia coli. Biotechnol. Bioeng. 2017, 114, 1511–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Ding, J.Y.; Ma, W.H.; Zhou, N.Y.; Liu, S.J. Identification and characterization of gamma-aminobutyric acid uptake system GabPCg (NCgl0464) in Corynebacterium glutamicum. Appl. Environ. Microbiol. 2012, 78, 2596–2601. [Google Scholar] [CrossRef] [Green Version]
- Kashiwagi, K.; Shibuya, S.; Tomitori, H.; Kuraishi, A.; Igarashi, K. Excretion and uptake of putrescine by the PotE protein in Escherichia coli. J. Biol. Chem. 1997, 272, 6318–6323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Ma, L.; Shen, X.; Wang, J.; Feng, Q.; Liu, L.; Zheng, G.; Yan, Y.; Sun, X.; Yuan, Q. Targeting metabolic driving and intermediate influx in lysine catabolism for high-level glutarate production. Nat. Commun. 2019, 10, 3337. [Google Scholar] [CrossRef] [PubMed]
- Salis, H.M.; Mirsky, E.A.; Voigt, C.A. Automated design of synthetic ribosome binding sites to control protein expression. Nat. Biotechnol. 2009, 27, 946–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, D.; Fey, J.P.; Eberhard, S.; Hicks, A.J.; Stern, D.B. In vivo evidence for the prokaryotic model of extended codon-anticodon interaction in translation initiation. EMBO J. 2003, 22, 651–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, C.; Niebisch, A.; Gebel, L.; Bott, M. Glutamate production by Corynebacterium glutamicum: Dependence on the oxoglutarate dehydrogenase inhibitor protein OdhI and protein kinase PknG. Appl. Microbiol. Biotechnol. 2007, 76, 691–700. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Shang, X.; Wang, B.; Hu, Q.; Liu, S.; Wen, T. Reconstruction of tricarboxylic acid cycle in Corynebacterium glutamicum with a genome-scale metabolic network model for trans-4-hydroxyproline production. Biotechnol. Bioeng. 2019, 116, 99–109. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhao, J.; Wang, X.; Tang, Y.; Liu, S.; Wen, T. Model-Guided Metabolic Rewiring for Gamma-Aminobutyric Acid and Butyrolactam Biosynthesis in Corynebacterium glutamicum ATCC13032. Biology 2022, 11, 846. https://doi.org/10.3390/biology11060846
Zhang Y, Zhao J, Wang X, Tang Y, Liu S, Wen T. Model-Guided Metabolic Rewiring for Gamma-Aminobutyric Acid and Butyrolactam Biosynthesis in Corynebacterium glutamicum ATCC13032. Biology. 2022; 11(6):846. https://doi.org/10.3390/biology11060846
Chicago/Turabian StyleZhang, Yun, Jing Zhao, Xueliang Wang, Yuan Tang, Shuwen Liu, and Tingyi Wen. 2022. "Model-Guided Metabolic Rewiring for Gamma-Aminobutyric Acid and Butyrolactam Biosynthesis in Corynebacterium glutamicum ATCC13032" Biology 11, no. 6: 846. https://doi.org/10.3390/biology11060846
APA StyleZhang, Y., Zhao, J., Wang, X., Tang, Y., Liu, S., & Wen, T. (2022). Model-Guided Metabolic Rewiring for Gamma-Aminobutyric Acid and Butyrolactam Biosynthesis in Corynebacterium glutamicum ATCC13032. Biology, 11(6), 846. https://doi.org/10.3390/biology11060846