
Effects of Field Simulated Marine Heatwaves on Sedimentary Organic Matter Quantity, Biochemical Composition, and Degradation Rates

 ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling Strategy
2.2. Biochemical Composition of Sediment Organic Matter
2.3. Extracellular Enzymatic Activities, C Degradation Rates and Turnover
2.4. Effect Size
2.5. Statistical Analyses
3. Results
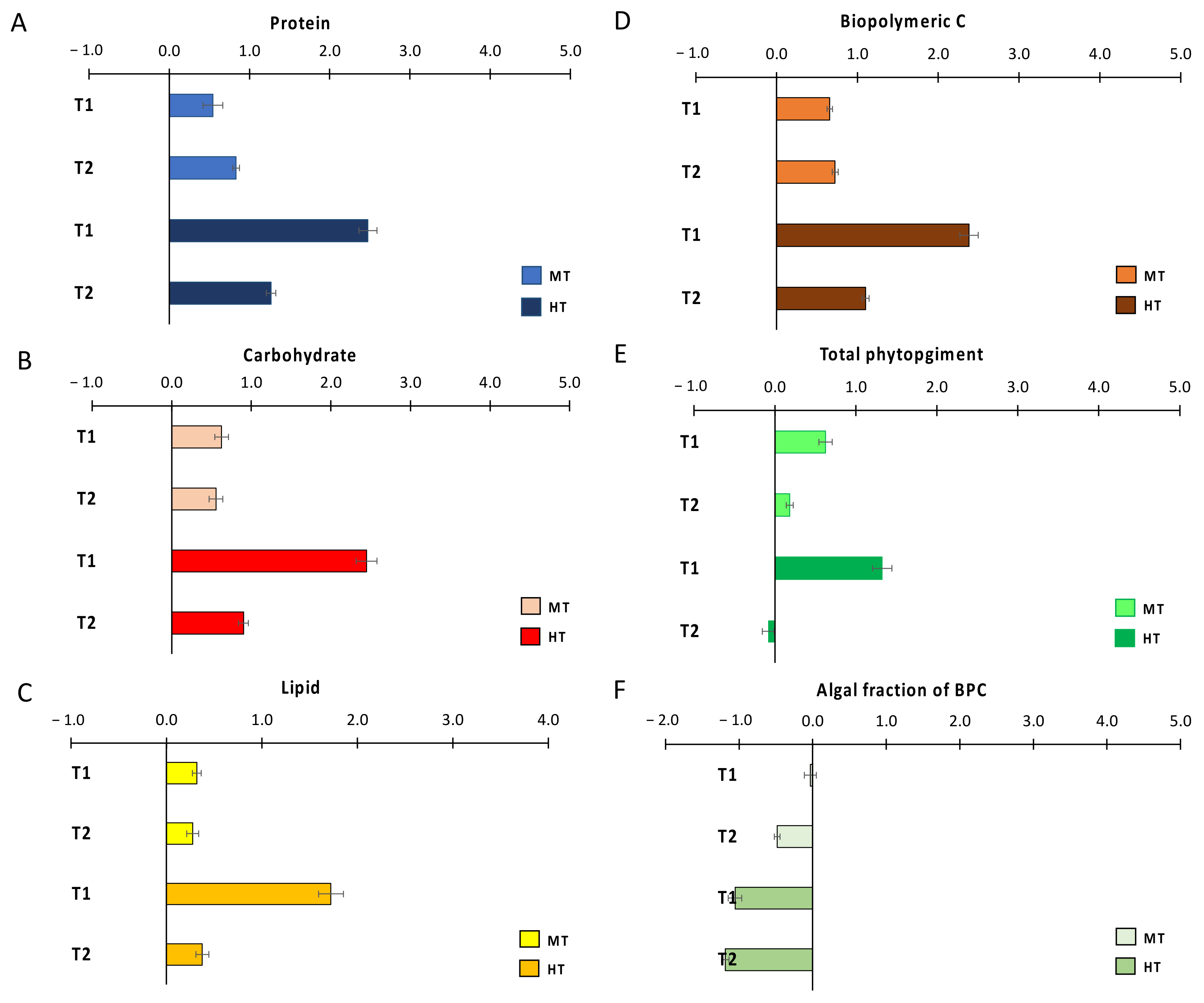
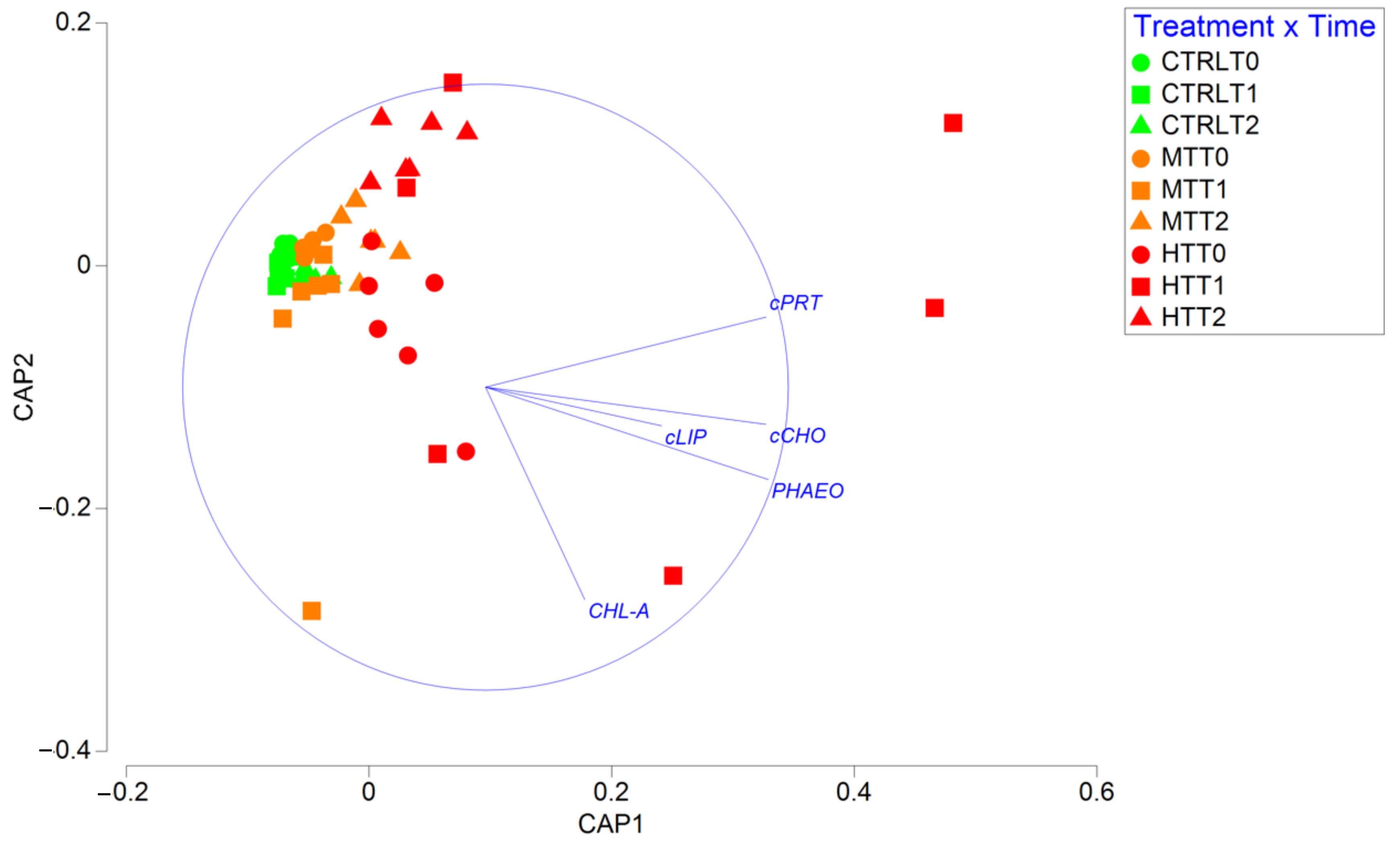
3.1. Effects of the Thermal Anomalies on Organic Matter Quantity and Biochemical Composition
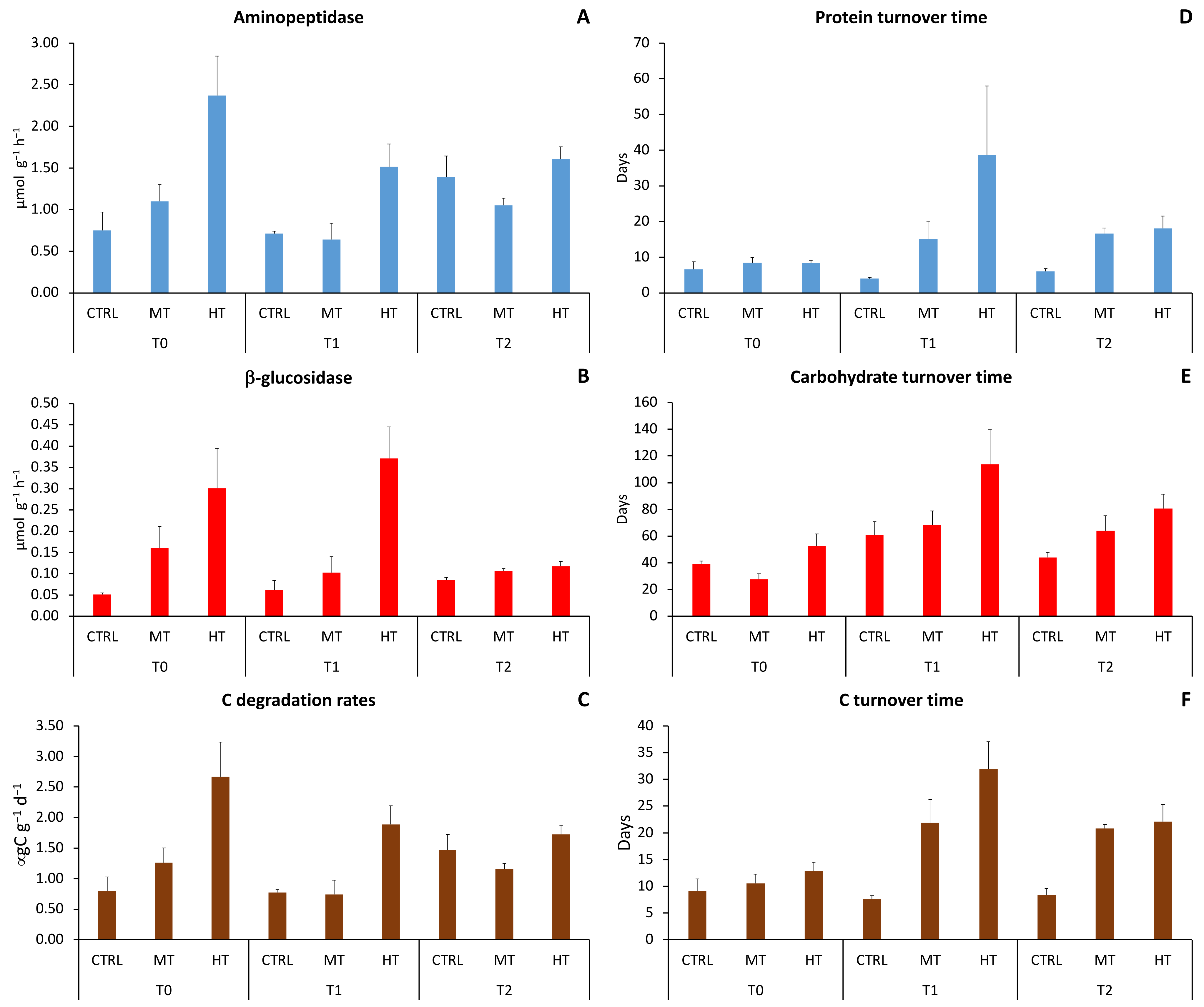
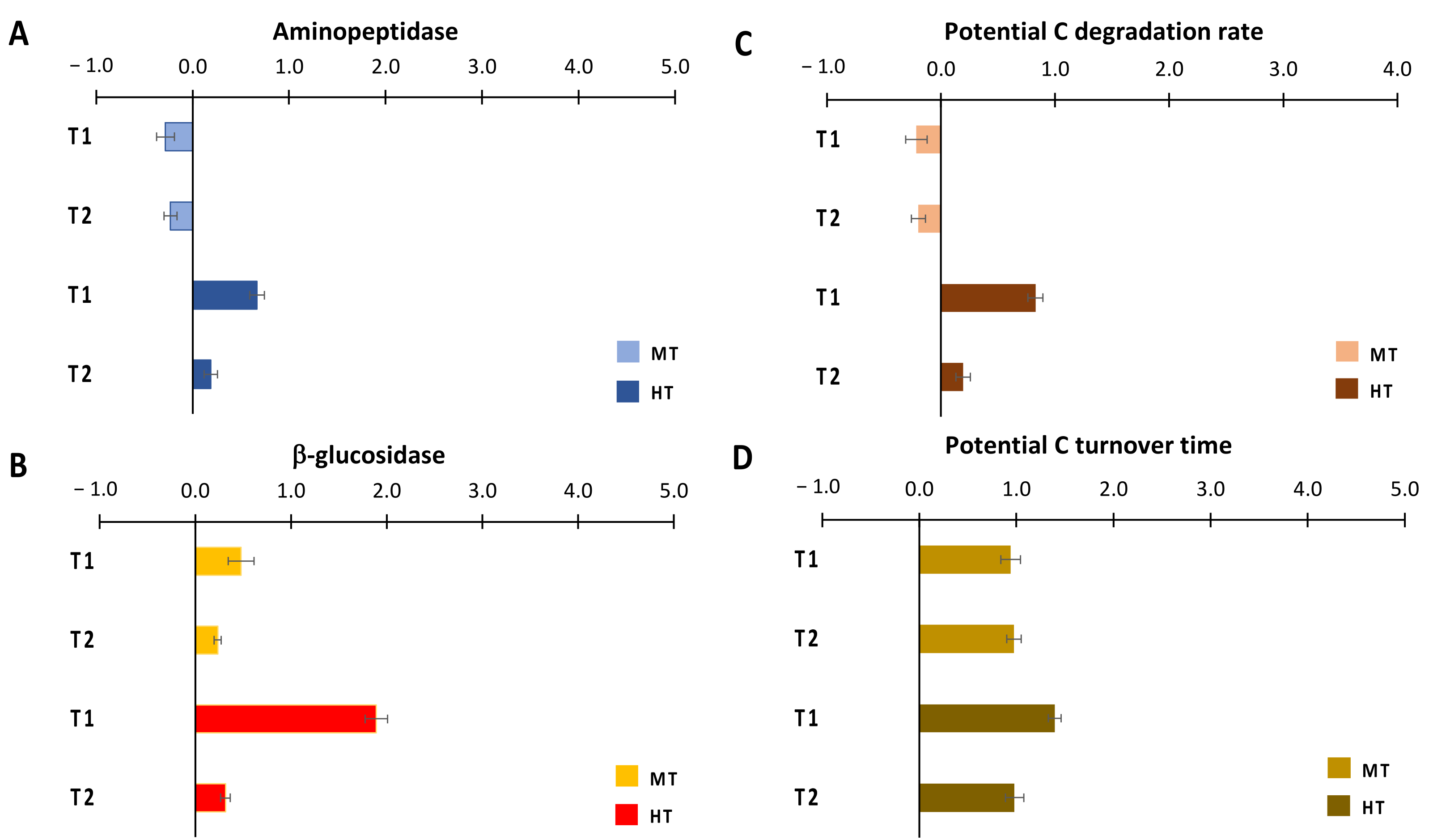
3.2. Sedimentary Organic Matter Degradation Rates
4. Discussion
4.1. MHWs Effects on Sedimentary Organic Matter Quantity, Biochemical Composition, and Nutritional Quality
4.2. MHWs’ Effects on Organic C Degradation Rates
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global Warming Transforms Coral Reef Assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M.; Bindi, M.; Brown, S.; Camilloni, I.; Diedhiou, A.; Djalante, R.; Ebi, K.L.; Engelbrecht, F.; et al. Impacts of 1.5 °C Global Warming on Natural and Human Systems. In Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; IPCC: Geneva, Switzerland, 2018; pp. 175–311. [Google Scholar]
- Hobday, A.J.; Alexander, L.V.; Perkins, S.E.; Smale, D.A.; Straub, S.C.; Oliver, E.C.J.; Benthuysen, J.A.; Burrows, M.T.; Donat, M.G.; Feng, M.; et al. A Hierarchical Approach to Defining Marine Heatwaves. Prog. Oceanogr. 2016, 141, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Frölicher, T.L.; Fischer, E.M.; Gruber, N. Marine Heatwaves under Global Warming. Nature 2018, 560, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Oliver, E.C.J.; Burrows, M.T.; Donat, M.G.; Sen Gupta, A.; Alexander, L.V.; Perkins-Kirkpatrick, S.E.; Benthuysen, J.A.; Hobday, A.J.; Holbrook, N.J.; Moore, P.J.; et al. Projected Marine Heatwaves in the 21st Century and the Potential for Ecological Impact. Front. Mar. Sci. 2019, 6, 734. [Google Scholar] [CrossRef] [Green Version]
- Benedetti-Cecchi, L. Complex Networks of Marine Heatwaves Reveal Abrupt Transitions in the Global Ocean. Sci. Rep. 2021, 11, 1739. [Google Scholar] [CrossRef]
- Sen Gupta, A.; Thomsen, M.; Benthuysen, J.A.; Hobday, A.J.; Oliver, E.; Alexander, L.V.; Burrows, M.T.; Donat, M.G.; Feng, M.; Holbrook, N.J.; et al. Drivers and Impacts of the Most Extreme Marine Heatwave Events. Sci. Rep. 2020, 10, 19359. [Google Scholar] [CrossRef]
- Kuglitsch, F.G.; Toreti, A.; Xoplaki, E.; Della-Marta, P.M.; Zerefos, C.S.; Türkeş, M.; Luterbacher, J. Heat Wave Changes in the Eastern Mediterranean since 1960. Geophys. Res. Lett. 2010, 37, L04802. [Google Scholar] [CrossRef] [Green Version]
- Darmaraki, S.; Somot, S.; Sevault, F.; Nabat, P.; Cabos Narvaez, W.D.; Cavicchia, L.; Djurdjevic, V.; Li, L.; Sannino, G.; Sein, D.V. Future Evolution of Marine Heatwaves in the Mediterranean Sea. Clim. Dyn. 2019, 53, 1371–1392. [Google Scholar] [CrossRef] [Green Version]
- Darmaraki, S.; Somot, S.; Sevault, F.; Nabat, P. Past Variability of Mediterranean Sea Marine Heatwaves. Geophys. Res. Lett. 2019, 46, 9813–9823. [Google Scholar] [CrossRef] [Green Version]
- Lejeusne, C.; Chevaldonné, P.; Pergent-Martini, C.; Boudouresque, C.F.; Pérez, T. Climate Change Effects on a Miniature Ocean: The Highly Diverse, Highly Impacted Mediterranean Sea. Trends Ecol. Evol. 2010, 25, 250–260. [Google Scholar] [CrossRef]
- Jacox, M.G.; Alexander, M.A.; Amaya, D.; Becker, E.; Bograd, S.J.; Brodie, S.; Hazen, E.L.; Pozo Buil, M.; Tommasi, D. Global Seasonal Forecasts of Marine Heatwaves. Nature 2022, 604, 486–490. [Google Scholar] [CrossRef]
- Wernberg, T.; Smale, D.A.; Tuya, F.; Thomsen, M.S.; Langlois, T.J.; de Bettignies, T.; Bennett, S.; Rousseaux, C.S. An Extreme Climatic Event Alters Marine Ecosystem Structure in a Global Biodiversity Hotspot. Nat. Clim. Chang. 2013, 3, 78–82. [Google Scholar] [CrossRef]
- Wernberg, T.; Bennett, S.; Babcock, R.C.; de Bettignies, T.; Cure, K.; Depczynski, M.; Dufois, F.; Fromont, J.; Fulton, C.J.; Hovey, R.K.; et al. Climate-Driven Regime Shift of a Temperate Marine Ecosystem. Science 2016, 353, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Cavole, L.; Demko, A.; Diner, R.; Giddings, A.; Koester, I.; Pagniello, C.; Paulsen, M.-L.; Ramirez-Valdez, A.; Schwenck, S.; Yen, N.; et al. Biological Impacts of the 2013–2015 Warm-Water Anomaly in the Northeast Pacific: Winners, Losers, and the Future. Oceanography 2016, 29, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Oliver, E.C.J.; Benthuysen, J.A.; Bindoff, N.L.; Hobday, A.J.; Holbrook, N.J.; Mundy, C.N.; Perkins-Kirkpatrick, S.E. The Unprecedented 2015/16 Tasman Sea Marine Heatwave. Nat. Commun. 2017, 8, 16101. [Google Scholar] [CrossRef] [Green Version]
- Smale, D.; Wernberg, T.; Vanderklift, M. Regional-Scale Variability in the Response of Benthic Macroinvertebrate Assemblages to a Marine Heatwave. Mar. Ecol. Prog. Ser. 2017, 568, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Cerrano, C.; Bavestrello, G.; Bianchi, C.N.; Cattaneo-vietti, R.; Bava, S.; Morganti, C.; Morri, C.; Picco, P.; Sara, G.; Schiaparelli, S.; et al. A Catastrophic Mass-Mortality Episode of Gorgonians and Other Organisms in the Ligurian Sea (North-Western Mediterranean), Summer 1999. Ecol. Lett. 2000, 3, 284–293. [Google Scholar] [CrossRef]
- Garrabou, J.; Perez, T.; Sartoretto, S.; Harmelin, J. Mass Mortality Event in Red Coral Corallium rubrum Populations in the Provence Region (France, NW Mediterranean). Mar. Ecol. Prog. Ser. 2001, 217, 263–272. [Google Scholar] [CrossRef]
- Coma, R.; Ribes, M.; Serrano, E.; Jiménez, E.; Salat, J.; Pascual, J. Global Warming-Enhanced Stratification and Mass Mortality Events in the Mediterranean. Proc. Natl. Acad. Sci. USA 2009, 106, 6176–6181. [Google Scholar] [CrossRef] [Green Version]
- Garrabou, J.; Coma, R.; Bensoussan, N.; Bally, M.; Chevaldonné, P.; Cigliano, M.; Diaz, D.; Harmelin, J.G.; Gambi, M.C.; Kersting, D.K.; et al. Mass Mortality in Northwestern Mediterranean Rocky Benthic Communities: Effects of the 2003 Heat Wave. Glob. Chang. Biol. 2009, 15, 1090–1103. [Google Scholar] [CrossRef]
- Marbà, N.; Duarte, C.M. Mediterranean Warming Triggers Seagrass (Posidonia oceanica) Shoot Mortality. Glob. Chang. Biol. 2010, 16, 2366–2375. [Google Scholar] [CrossRef]
- Rivetti, I.; Fraschetti, S.; Lionello, P.; Zambianchi, E.; Boero, F. Global Warming and Mass Mortalities of Benthic Invertebrates in the Mediterranean Sea. PLoS ONE 2014, 9, e115655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodeur, R.D.; Auth, T.D.; Phillips, A.J. Major Shifts in Pelagic Micronekton and Macrozooplankton Community Structure in an Upwelling Ecosystem Related to an Unprecedented Marine Heatwave. Front. Mar. Sci. 2019, 6, 212. [Google Scholar] [CrossRef] [Green Version]
- Sorte, C.J.B.; Fuller, A.; Bracken, M.E.S. Impacts of a Simulated Heat Wave on Composition of a Marine Community. Oikos 2010, 119, 1909–1918. [Google Scholar] [CrossRef]
- Smale, D.A.; Wernberg, T.; Peck, L.S.; Barnes, D.K.A. Turning on the Heat: Ecological Response to Simulated Warming in the Sea. PLoS ONE 2011, 6, e16050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smale, D.A.; Yunnie, A.L.E.; Vance, T.; Widdicombe, S. Disentangling the Impacts of Heat Wave Magnitude, Duration and Timing on the Structure and Diversity of Sessile Marine Assemblages. PeerJ 2015, 3, e863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuels, T.; Rynearson, T.A.; Collins, S. Surviving Heatwaves: Thermal Experience Predicts Life and Death in a Southern Ocean Diatom. Front. Mar. Sci. 2021, 8, 600343. [Google Scholar] [CrossRef]
- Stipcich, P.; Marín-Guirao, L.; Pansini, A.; Pinna, F.; Procaccini, G.; Pusceddu, A.; Soru, S.; Ceccherelli, G. Effects of Current and Future Summer Marine Heat Waves on Posidonia oceanica: Plant Origin Matters? Front. Clim. 2022, 4, 844831. [Google Scholar] [CrossRef]
- Sanz-Lázaro, C.; Valdemarsen, T.; Holmer, M. Effects of Temperature and Organic Pollution on Nutrient Cycling in Marine Sediments. Biogeosciences 2015, 12, 4565–4575. [Google Scholar] [CrossRef] [Green Version]
- Alsterberg, C.; Sundbäck, K.; Hulth, S. Functioning of a Shallow-Water Sediment System during Experimental Warming and Nutrient Enrichment. PLoS ONE 2012, 7, e51503. [Google Scholar] [CrossRef]
- Duarte, C.M.; Middelburg, J.J.; Caraco, N. Major Role of Marine Vegetation on the Oceanic Carbon Cycle. Biogeosciences 2005, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Abd-Elgawad, A.; Abram, N.; Adler, C.; Alegría, A.; Arístegui, J.; Bindoff, N.L.; Bouwer, L.; Cáceres, B.; Cai, R.; Cassotta, S.; et al. IPCC Technical Summary. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Pörtner, H.-O., Roberts, D.C., Masson-Delmotte, V., Zhai, P., Tignor, M., Poloczanska, E., Mintenbeck, K., Alegría, A., Nicolai, M., Okem, A., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2019. [Google Scholar]
- Danovaro, R. Methods for the Study of Deep-Sea Sediments, Their Functioning and Biodiversity, 1st ed.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2010; ISBN 9781439811375. [Google Scholar]
- Pusceddu, A.; Bianchelli, S.; Canals, M.; Sanchez-Vidal, A.; Durrieu De Madron, X.; Heussner, S.; Lykousis, V.; de Stigter, H.; Trincardi, F.; Danovaro, R. Organic Matter in Sediments of Canyons and Open Slopes of the Portuguese, Catalan, Southern Adriatic and Cretan Sea Margins. Deep. Sea Res. Part I Oceanogr. Res. 2010, 57, 441–457. [Google Scholar] [CrossRef]
- Pusceddu, A.; Dell’Anno, A.; Fabiano, M.; Danovaro, R. Quantity and Bioavailability of Sediment Organic Matter as Signatures of Benthic Trophic Status. Mar. Ecol. Prog. Ser. 2009, 375, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Hartree, E.F. Determination of Proteins: A Modification of the Lowry Method That Gives a Linear Photometric Response. Anal. Biochem. 1972, 48, 422–427. [Google Scholar] [CrossRef]
- Rice, D.L. The Detritus Nitrogen Problem: New Observations and Perspectives from Organic Geochemistry. Mar. Ecol. Prog. Ser. 1982, 9, 153–162. [Google Scholar] [CrossRef]
- Gerchakov, S.M.; Hatcher, P.G. Improved Technique for Analysis of Carbohydrates in Sediments. Limnol. Oceanogr. 1972, 17, 938–943. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Marsh, J.B.; Weinstein, W.J. A Simple Charring Method for Determination of Lipids. J. Lipid Res. 1966, 7, 574–576. [Google Scholar] [CrossRef]
- Fabiano, M.; Danovaro, R.; Fraschetti, S. A Three-Year Time Series of Elemental and Biochemical Composition of Organic Matter in Subtidal Sandy Sediments of the Ligurian Sea (Northwestern Mediterranean). Cont. Shelf Res. 1995, 15, 1453–1469. [Google Scholar] [CrossRef]
- Bianchelli, S.; Nizzoli, D.; Bartoli, M.; Viaroli, P.; Rastelli, E.; Pusceddu, A. Sedimentary Organic Matter, Prokaryotes, and Meiofauna across a River-Lagoon-Sea Gradient. Diversity 2020, 12, 189. [Google Scholar] [CrossRef]
- Pusceddu, A.; Bianchelli, S.; Martín, J.; Puig, P.; Palanques, A.; Masqué, P.; Danovaro, R. Chronic and Intensive Bottom Trawling Impairs Deep-Sea Biodiversity and Ecosystem Functioning. Proc. Natl. Acad. Sci. USA 2014, 111, 8861–8866. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.J. A New Method for Non-Parametric Multivariate Analysis of Variance: Non-Parametric Manova for Ecology. Austr. Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J.; Millar, R.B. Spatial Variation and Effects of Habitat on Temperate Reef Fish Assemblages in Northeastern New Zealand. J. Exp. Mar. Biol. Ecol. 2004, 305, 191–221. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; Balakrishnan, N., Colton, T., Everitt, B., Piegorsch, W., Ruggeri, F., Teugels, J.L., Eds.; Wiley: Hoboken, NJ, USA; pp. 1–15. ISBN 978-1-118-44511-2.
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods, 1st ed.; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Levitus, S.; Antonov, J.; Boyer, T. Warming of the World Ocean. Science 2005, 287, 2225–2229. [Google Scholar] [CrossRef] [Green Version]
- Lyman, J.M.; Good, S.A.; Gouretski, V.V.; Ishii, M.; Johnson, G.C.; Palmer, M.D.; Smith, D.M.; Willis, J.K. Robust Warming of the Global Upper Ocean. Nature 2010, 465, 334–337. [Google Scholar] [CrossRef]
- Harley, C.D.G.; Randall Hughes, A.; Hultgren, K.M.; Miner, B.G.; Sorte, C.J.B.; Thornber, C.S.; Rodriguez, L.F.; Tomanek, L.; Williams, S.L. The Impacts of Climate Change in Coastal Marine Systems: Climate Change in Coastal Marine Systems. Ecol. Lett. 2006, 9, 228–241. [Google Scholar] [CrossRef] [Green Version]
- Oliver, E.C.J.; Donat, M.G.; Burrows, M.T.; Moore, P.J.; Smale, D.A.; Alexander, L.V.; Benthuysen, J.A.; Feng, M.; Sen Gupta, A.; Hobday, A.J.; et al. Longer and More Frequent Marine Heatwaves over the Past Century. Nat. Commun. 2018, 9, 1324. [Google Scholar] [CrossRef]
- Traving, S.J.; Kellogg, C.T.E.; Ross, T.; McLaughlin, R.; Kieft, B.; Ho, G.Y.; Peña, A.; Krzywinski, M.; Robert, M.; Hallam, S.J. Prokaryotic Responses to a Warm Temperature Anomaly in Northeast Subarctic Pacific Waters. Commun. Biol. 2021, 4, 1217. [Google Scholar] [CrossRef]
- Dell’Anno, A.; Mei, M.L.; Pusceddu, A.; Danovaro, R. Assessing the Trophic State and Eutrophication of Coastal Marine Systems: A New Approach Based on the Biochemical Composition of Sediment Organic Matter. Mar. Pollut. Bull. 2002, 44, 611–622. [Google Scholar] [CrossRef]
- Bianchelli, S.; Pusceddu, A.; Buschi, E.; Danovaro, R. Trophic Status and Meiofauna Biodiversity in the Northern Adriatic Sea: Insights for the Assessment of Good Environmental Status. Mar. Environ. Res. 2016, 113, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Finkel, Z.V.; Beardall, J.; Flynn, K.J.; Quigg, A.; Rees, T.A.V.; Raven, J.A. Phytoplankton in a Changing World: Cell Size and Elemental Stoichiometry. J. Plankton Res. 2010, 32, 119–137. [Google Scholar] [CrossRef] [Green Version]
- Lewandowska, A.M.; Breithaupt, P.; Hillebrand, H.; Hoppe, H.-G.; Jürgens, K.; Sommer, U. Responses of Primary Productivity to Increased Temperature and Phytoplankton Diversity. J. Sea Res. 2012, 72, 87–93. [Google Scholar] [CrossRef]
- Novak, T.; Godrijan, J.; Pfannkuchen, D.M.; Djakovac, T.; Medić, N.; Ivančić, I.; Mlakar, M.; Gašparović, B. Global Warming and Oligotrophication Lead to Increased Lipid Production in Marine Phytoplankton. Sci. Total Environ. 2019, 668, 171–183. [Google Scholar] [CrossRef]
- Remy, M.; Hillebrand, H.; Flöder, S. Stability of Marine Phytoplankton Communities Facing Stress Related to Global Change: Interactive Effects of Heat Waves and Turbidity. J. Exp. Mar. Biol. Ecol. 2017, 497, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Pusceddu, A.; Dell’Anno, A.; Danovaro, R.; Manini, E.; Sara, G.; Fabiano, M. Enzymatically Hydrolyzable Protein and Carbohydrate Sedimentary Pools as Indicators of the Trophic State of Detritus Sink Systems: A Case Study in a Mediterranean Coastal Lagoon. Estuaries 2003, 26, 641–650. [Google Scholar] [CrossRef]
- Pansch, C.; Scotti, M.; Barboza, F.R.; Al-Janabi, B.; Brakel, J.; Briski, E.; Bucholz, B.; Franz, M.; Ito, M.; Paiva, F.; et al. Heat Waves and Their Significance for a Temperate Benthic Community: A near-Natural Experimental Approach. Glob. Chang. Biol. 2018, 24, 4357–4367. [Google Scholar] [CrossRef]
- Richon, C.; Dutay, J.-C.; Bopp, L.; Le Vu, B.; Orr, J.C.; Somot, S.; Dulac, F. Biogeochemical Response of the Mediterranean Sea to the Transient SRES-A2 Climate Change Scenario. Biogeosciences 2019, 16, 135–165. [Google Scholar] [CrossRef] [Green Version]
- Chust, G.; Allen, J.I.; Bopp, L.; Schrum, C.; Holt, J.; Tsiaras, K.; Zavatarelli, M.; Chifflet, M.; Cannaby, H.; Dadou, I.; et al. Biomass Changes and Trophic Amplification of Plankton in a Warmer Ocean. Glob. Chang. Biol. 2014, 20, 2124–2139. [Google Scholar] [CrossRef]
- Solidoro, C.; Bastianini, M.; Bandelj, V.; Codermatz, R.; Cossarini, G.; Melaku Canu, D.; Ravagnan, E.; Salon, S.; Trevisani, S. Current State, Scales of Variability, and Trends of Biogeochemical Properties in the Northern Adriatic Sea. J. Geophys. Res. 2009, 114, C07S91. [Google Scholar] [CrossRef]
- Stephens, D.W.; Krebs, J.R. Foraging Theory; Princeton University Press: Princeton, NJ, USA, 1986; ISBN 9780691084428. [Google Scholar]
- Li, Y.; Sun, L.-L.; Sun, Y.-Y.; Cha, Q.-Q.; Li, C.-Y.; Zhao, D.-L.; Song, X.-Y.; Wang, M.; McMinn, A.; Chen, X.-L.; et al. Extracellular Enzyme Activity and Its Implications for Organic Matter Cycling in Northern Chinese Marginal Seas. Front. Microbiol. 2019, 10, 2137. [Google Scholar] [CrossRef] [Green Version]
- Arndt, S.; Jørgensen, B.B.; LaRowe, D.E.; Middelburg, J.J.; Pancost, R.D.; Regnier, P. Quantifying the Degradation of Organic Matter in Marine Sediments: A Review and Synthesis. Earth-Sci. Rev. 2013, 123, 53–86. [Google Scholar] [CrossRef]
- Christian, J.R.; Karl, D.M. Bacterial Ectoenzymes in Marine Waters: Activity Ratios and Temperature Responses in Three Oceanographic Provinces. Limnol. Oceanogr. 1995, 40, 1042–1049. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.B.; Fukami, K.; Nishijima, T. Regulation of Seasonal Variability of Aminopeptidase Activities in Surface and Bottom Waters of Uranouchi Inlet, Japan. Aquat. Microb. Ecol. 2000, 21, 139–149. [Google Scholar] [CrossRef]
- Arnosti, C.; Jørgensen, B. High Activity and Low Temperature Optima of Extracellular Enzymes in Arctic Sediments: Implications for Carbon Cycling by Heterotrophic Microbial Communities. Mar. Ecol. Prog. Ser. 2003, 249, 15–24. [Google Scholar] [CrossRef]
- Franzo, A.; Celussi, M.; Bazzaro, M.; Relitti, F.; Del Negro, P. Microbial Processing of Sedimentary Organic Matter at a Shallow LTER Site in the Northern Adriatic Sea: An 8-Year Case Study. Nat. Conserv. 2019, 34, 397–415. [Google Scholar] [CrossRef]
- Azam, F.; Fenchel, T.; Field, J.; Gray, J.; Meyer-Reil, L.; Thingstad, F. The Ecological Role of Water-Column Microbes in the Sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Amon, R.M.W.; Benner, R. Bacterial Utilization of Different Size Classes of Dissolved Organic Matter. Limnol. Oceanogr. 1996, 41, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Benner, R.; Amon, R.M.W. The Size-Reactivity Continuum of Major Bioelements in the Ocean. Annu. Rev. Mar. Sci. 2015, 7, 185–205. [Google Scholar] [CrossRef]
- Weiss, M.S.; Abele, U.; Weckesser, J.; Welte, W.; Schiltz, E.; Schulz, G.E. Molecular Architecture and Electrostatic Properties of a Bacterial Porin. Science 1991, 254, 1627–1630. [Google Scholar] [CrossRef]
- Thomson, B.; Hepburn, C.D.; Lamare, M.; Baltar, F. Temperature and UV Light Affect the Activity of Marine Cell-Free Enzymes. Biogeosciences 2017, 14, 3971–3977. [Google Scholar] [CrossRef] [Green Version]
- van Nugteren, P.; Moodley, L.; Brummer, G.-J.; Heip, C.H.R.; Herman, P.M.J.; Middelburg, J.J. Seafloor Ecosystem Functioning: The Importance of Organic Matter Priming. Mar. Biol. 2009, 156, 2277–2287. [Google Scholar] [CrossRef] [Green Version]
- Lionello, P.; Scarascia, L. The Relation between Climate Change in the Mediterranean Region and Global Warming. Reg. Environ. Chang. 2018, 18, 1481–1493. [Google Scholar] [CrossRef]
- Juza, M.; Fernández-Mora, À.; Tintoré, J. Sub-Regional Marine Heat Waves in the Mediterranean Sea from Observations: Long-Term Surface Changes, Sub-Surface and Coastal Responses. Front. Mar. Sci. 2022, 9, 785771. [Google Scholar] [CrossRef]
- Atwood, T.B.; Witt, A.; Mayorga, J.; Hammill, E.; Sala, E. Global Patterns in Marine Sediment Carbon Stocks. Front. Mar. Sci. 2020, 7, 165. [Google Scholar] [CrossRef]
- Verdura, J.; Linares, C.; Ballesteros, E.; Coma, R.; Uriz, M.J.; Bensoussan, N.; Cebrian, E. Biodiversity Loss in a Mediterranean Ecosystem Due to an Extreme Warming Event Unveils the Role of an Engineering Gorgonian Species. Sci. Rep. 2019, 9, 5911. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Source | Df | MS | F | P (MC) | Explained Variance (%) |
|---|---|---|---|---|---|---|
| Protein | Tr | 2 | 1.705 | 29.0 | ** | 50.9 |
| Ti | 2 | 0.149 | 2.5 | ns | 2.8 | |
| Tr × Ti | 4 | 0.205 | 3.5 | * | 13.6 | |
| Res | 45 | 0.058 | 32.6 | |||
| Carbohydrate | Tr | 2 | 1.149 | 11.8 | ** | 26.1 |
| Ti | 2 | 0.414 | 4.3 | * | 7.9 | |
| Tr × Ti | 4 | 0.403 | 4.2 | ** | 22.7 | |
| Res | 45 | 0.097 | 43.3 | |||
| Lipid | Tr | 2 | 0.186 | 9.8 | ** | 28.0 |
| Ti | 2 | 0.040 | 2.1 | ns | 3.6 | |
| Tr × Ti | 4 | 0.042 | 2.2 | ns | 11.5 | |
| Res | 45 | 0.019 | 56.9 | |||
| Chlorophyll-a | Tr | 2 | 1.479 | 2.9 | ns | 7.6 |
| Ti | 2 | 0.740 | 1.4 | ns | 1.8 | |
| Tr × Ti | 4 | 1.237 | 2.4 | ns | 17.1 | |
| Res | 45 | 0.516 | 73.5 | |||
| Phaeopigment | Tr | 2 | 8.211 | 16.231 | ** | 28.4 |
| Ti | 2 | 2.724 | 5.385 | ** | 8.2 | |
| Tr × Ti | 4 | 3.216 | 6.358 | ** | 29.9 | |
| Res | 45 | 0.506 | 33.5 | |||
| Total phytopigment | Tr | 2 | 16.554 | 12.5 | ** | 27.3 |
| Ti | 2 | 5.575 | 4.2 | * | 7.6 | |
| Tr × Ti | 4 | 5.499 | 4.2 | ** | 22.5 | |
| Res | 45 | 1.322 | 42.6 | |||
| Biopolymeric C | Tr | 2 | 7.861 | 23.8 | ** | 44.1 |
| Ti | 2 | 0.794 | 2.4 | ns | 2.7 | |
| Tr × Ti | 4 | 1.372 | 4.2 | ** | 18.3 | |
| Res | 45 | 0.330 | 34.8 | |||
| Autotrophic fraction of biopolymeric C | Tr | 2 | 0.064 | 16.9 | ** | 37.1 |
| Ti | 2 | 0.021 | 5.5 | ** | 10.6 | |
| Tr × Ti | 4 | 0.009 | 2.5 | ns | 10.4 | |
| Res | 45 | 0.004 | 41.9 | |||
| OM biochemical composition | Tr | 2 | 12.731 | 10.6 | ** | 24.2 |
| Ti | 2 | 4.068 | 3.4 | * | 6.0 | |
| Tr × Ti | 4 | 5.103 | 4.3 | ** | 24.6 | |
| Res | 45 | 1.196 | 45.2 |
| Anomaly | Contrast | Protein | Carbohydrate | Lipid | Phytopigment | Biopolymeric C |
|---|---|---|---|---|---|---|
| MT | T0 vs. T1 | ** | ns | ** | ** | ** |
| T0 vs. T2 | * | ns | ** | * | * | |
| T1 vs. T2 | * | ns | ns | ** | ns | |
| HT | T0 vs. T1 | ** | ** | * | ns | ** |
| T0 vs. T2 | ** | *** | *** | *** | *** | |
| T1 vs. T2 | *** | *** | *** | *** | *** |
| Variable | Source | df | MS | F | P (MC) | % of Explained Variance |
|---|---|---|---|---|---|---|
| Aminopeptidase | Tr | 2 | 4.760 | 13.7 | ** | 35.1 |
| Ti | 2 | 1.071 | 3.1 | ns | 5.7 | |
| Tr × Ti | 4 | 0.744 | 2.1 | ns | 9.5 | |
| Res | 45 | 0.347 | 49.7 | |||
| β-glucosidase | Tr | 2 | 0.184 | 14.7 | ** | 34.6 |
| Ti | 2 | 0.030 | 2.4 | ns | 3.6 | |
| Tr × Ti | 4 | 0.039 | 3.1 | * | 16.3 | |
| Res | 45 | 0.013 | 45.6 | |||
| C degradation rate | Tr | 2 | 6.744 | 15.0 | ** | 38.1 |
| Ti | 2 | 0.927 | 2.1 | ns | 2.9 | |
| Tr × Ti | 4 | 0.998 | 2.2 | ns | 9.9 | |
| Res | 45 | 0.451 | 49.1 | |||
| Protein turnover time (d) | Tr | 2 | 490.2 | 11.6 | ** | 30.4 |
| Ti | 2 | 178.7 | 4.2 | * | 9.2 | |
| Tr × Ti | 4 | 85.5 | 2.0 | ns | 8.8 | |
| Res | 45 | 42.3 | 51.6 | |||
| Carbohydrate turnover time (d) | Tr | 2 | 3842.8 | 6.9 | ** | 18.2 |
| Ti | 2 | 5574.0 | 10.0 | ** | 27.8 | |
| Tr × Ti | 4 | 438.2 | 0.8 | ns | 0.0 | |
| Res | 45 | 559.7 | 54.0 | |||
| C turnover time (d) | Tr | 2 | 905.4 | 19.6 | ** | 34.9 |
| Ti | 2 | 424.1 | 9.2 | ** | 15.3 | |
| Tr × Ti | 4 | 178.1 | 3.9 | * | 16.1 | |
| Res | 45 | 46.1 | 33.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soru, S.; Stipcich, P.; Ceccherelli, G.; Ennas, C.; Moccia, D.; Pusceddu, A. Effects of Field Simulated Marine Heatwaves on Sedimentary Organic Matter Quantity, Biochemical Composition, and Degradation Rates. Biology 2022, 11, 841. https://doi.org/10.3390/biology11060841
Soru S, Stipcich P, Ceccherelli G, Ennas C, Moccia D, Pusceddu A. Effects of Field Simulated Marine Heatwaves on Sedimentary Organic Matter Quantity, Biochemical Composition, and Degradation Rates. Biology. 2022; 11(6):841. https://doi.org/10.3390/biology11060841
Chicago/Turabian StyleSoru, Santina, Patrizia Stipcich, Giulia Ceccherelli, Claudia Ennas, Davide Moccia, and Antonio Pusceddu. 2022. "Effects of Field Simulated Marine Heatwaves on Sedimentary Organic Matter Quantity, Biochemical Composition, and Degradation Rates" Biology 11, no. 6: 841. https://doi.org/10.3390/biology11060841
APA StyleSoru, S., Stipcich, P., Ceccherelli, G., Ennas, C., Moccia, D., & Pusceddu, A. (2022). Effects of Field Simulated Marine Heatwaves on Sedimentary Organic Matter Quantity, Biochemical Composition, and Degradation Rates. Biology, 11(6), 841. https://doi.org/10.3390/biology11060841