
Joint Transcriptome and Metabolome Analysis Prevails the Biological Mechanisms Underlying the Pro-Survival Fight in In Vitro Heat-Stressed Granulosa Cells

 ,
,  ,
,  ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Granulosa Cell Culture, Heat Treatment, and Cell Experiments
2.2. Transcriptomics Data and Differentially Expressed Genes
2.3. Metabolomics Data and Differentially Expressed Metabolites
2.4. Integrated Pathway Analysis of Genes and Metabolites
2.5. Interaction Network Analysis among Genes and Metabolites
3. Results
3.1. Effect of Heat Stress on Granulosa Cell Parameters
3.2. Transcriptome and Metabolome from Heat-Stressed Granulosa Cells
3.3. Integrated Pathways Analysis of Genes and Metabolites
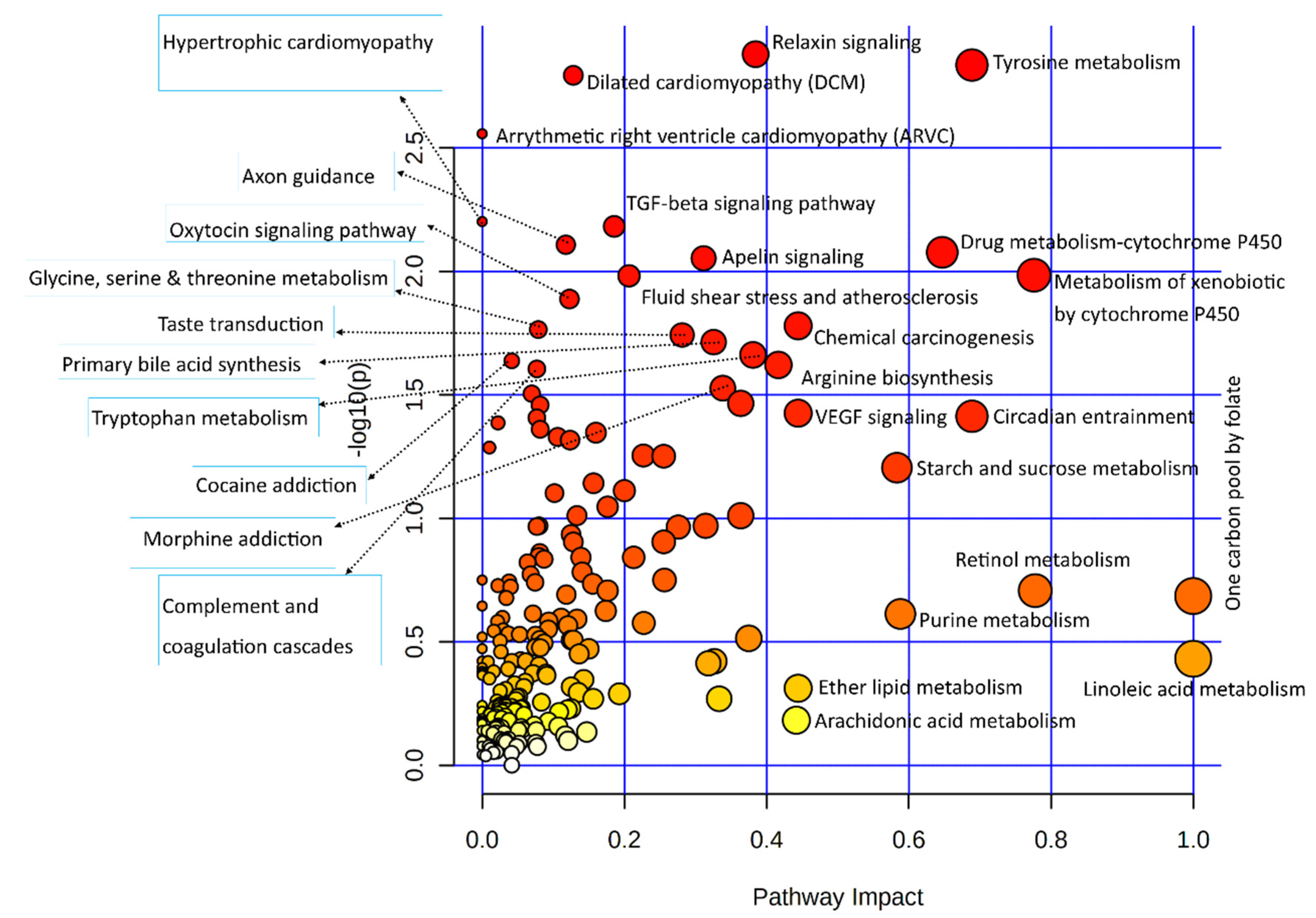
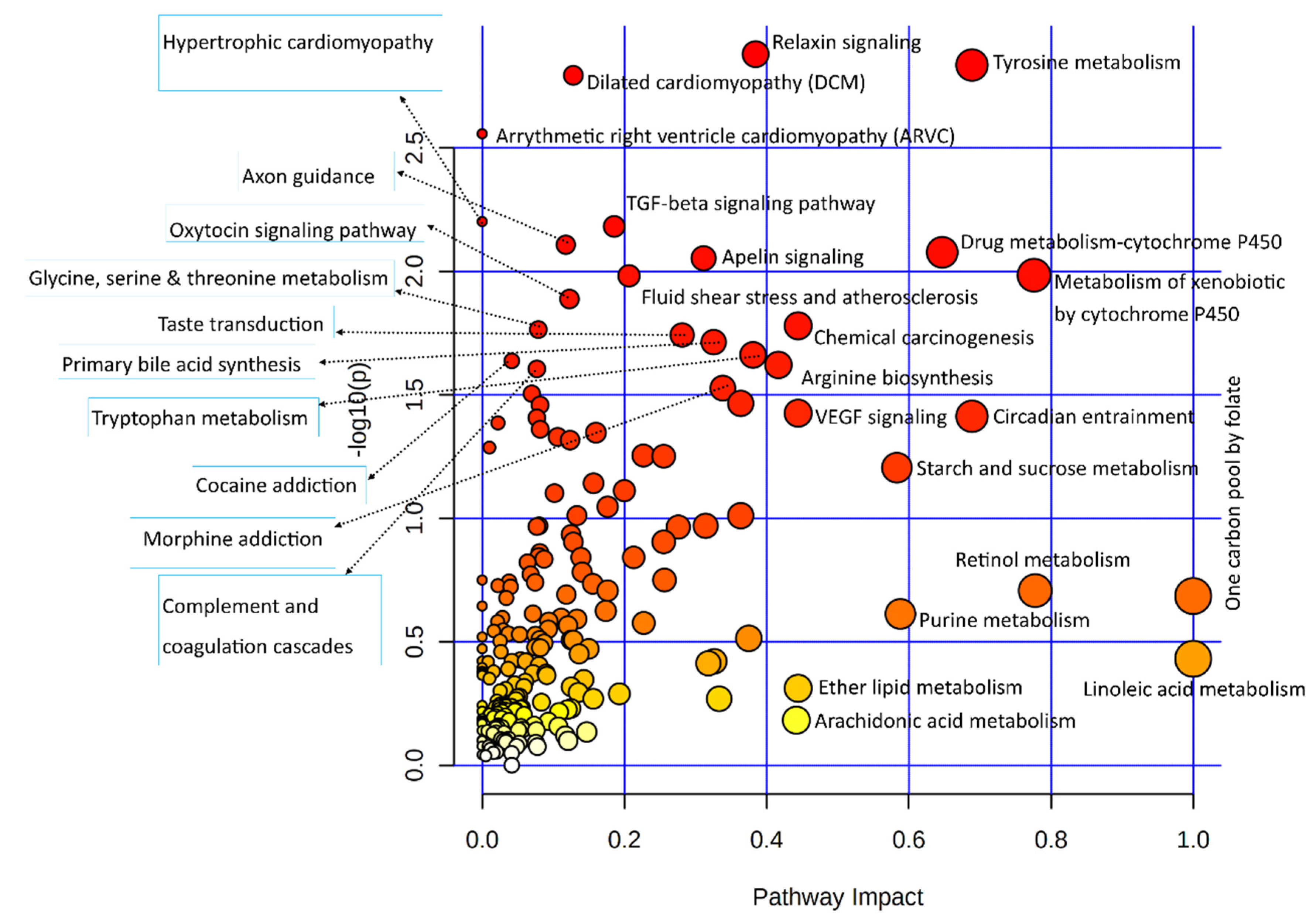
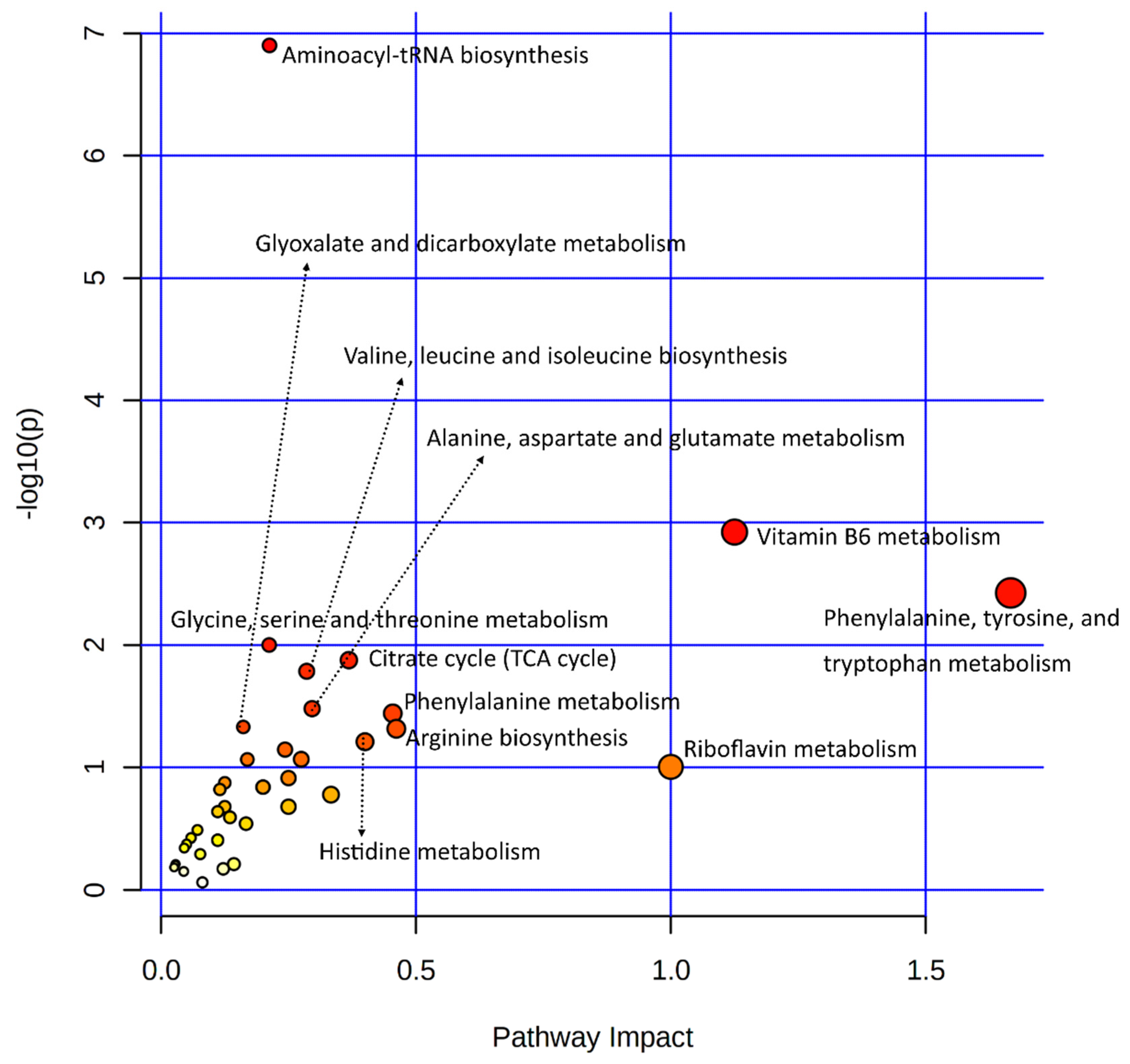
3.3.1. Joint Pathway Enrichment of Genes Used in This Study
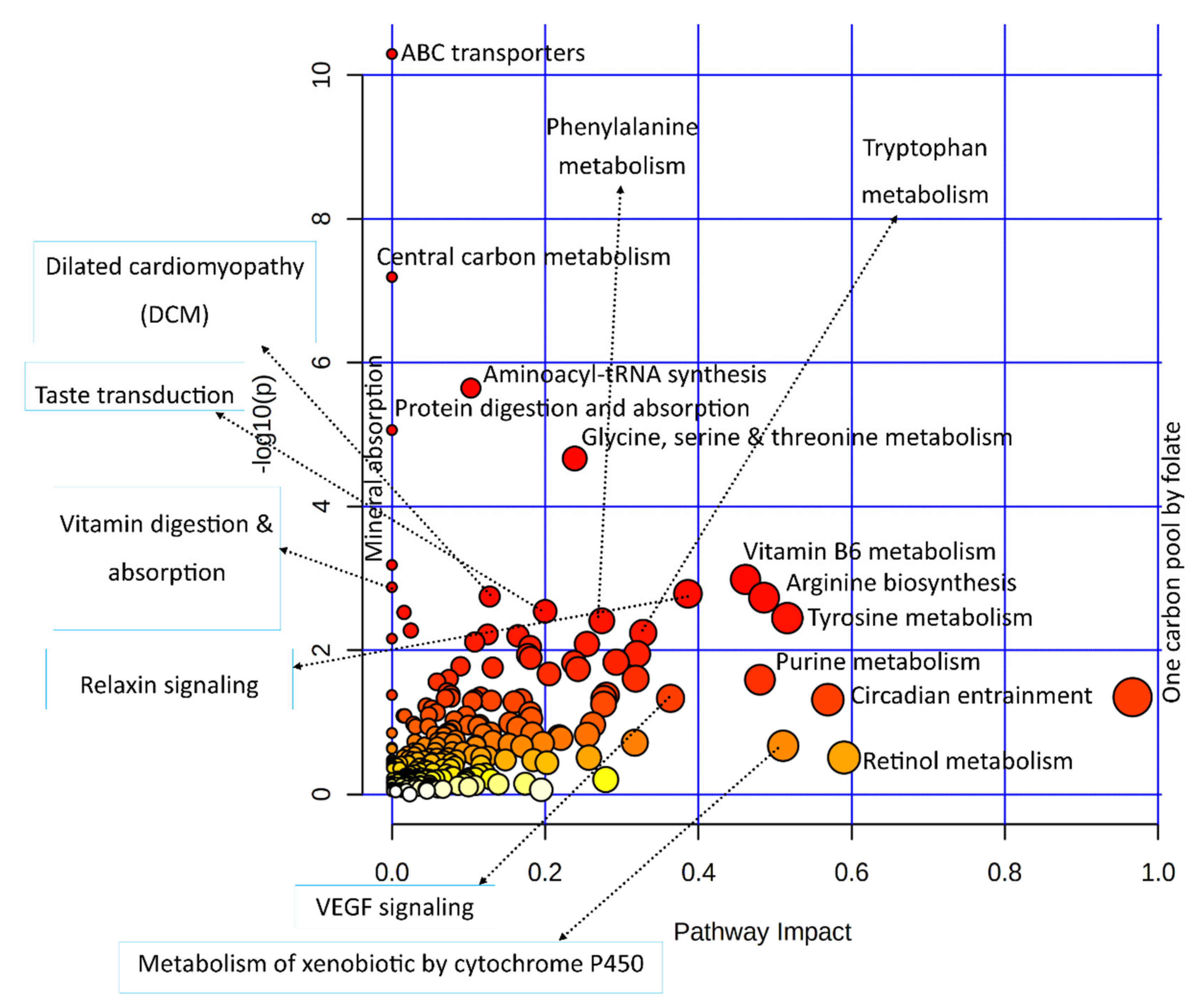
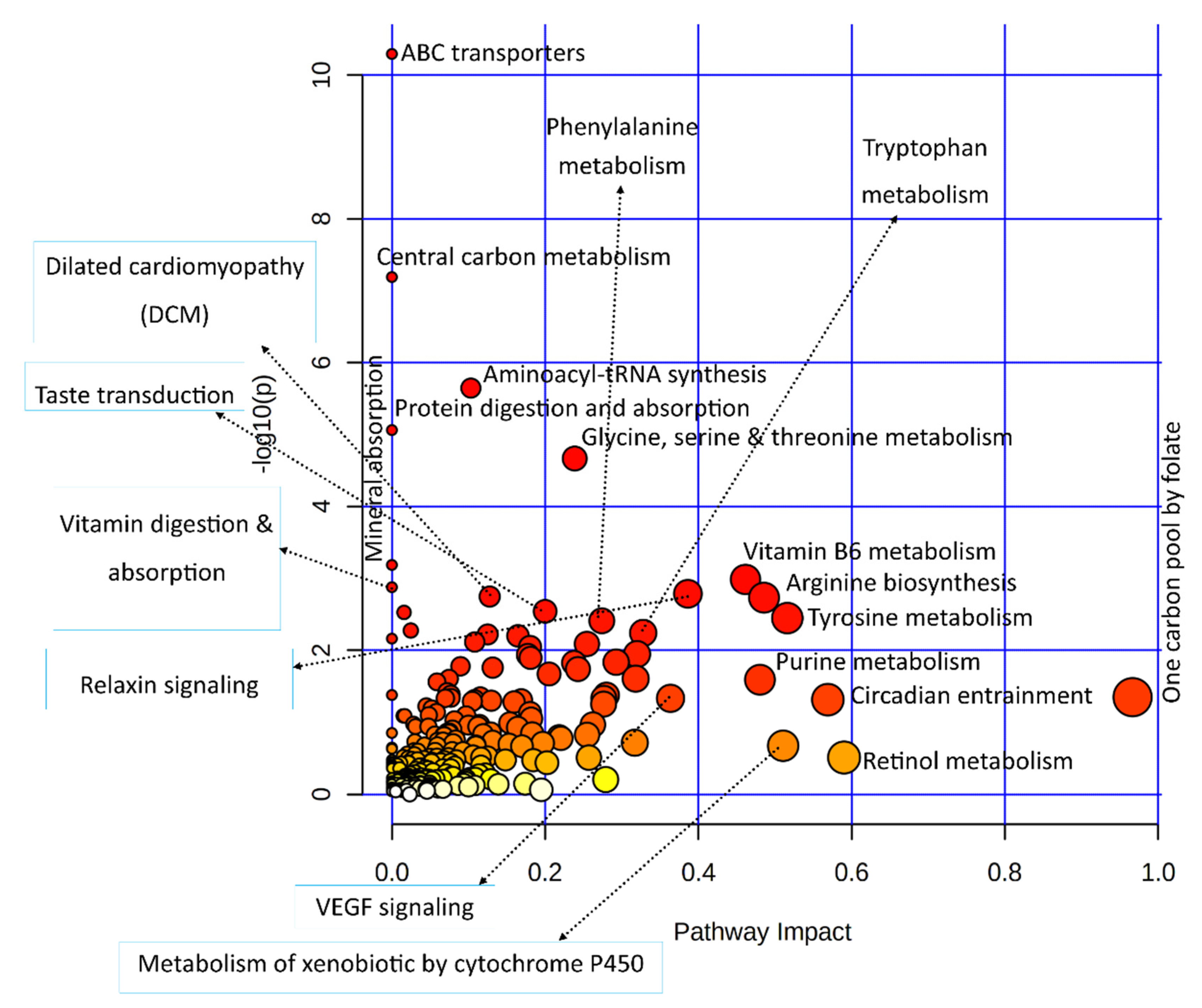
3.3.2. Combined Pathway Enrichment of Genes and Metabolites
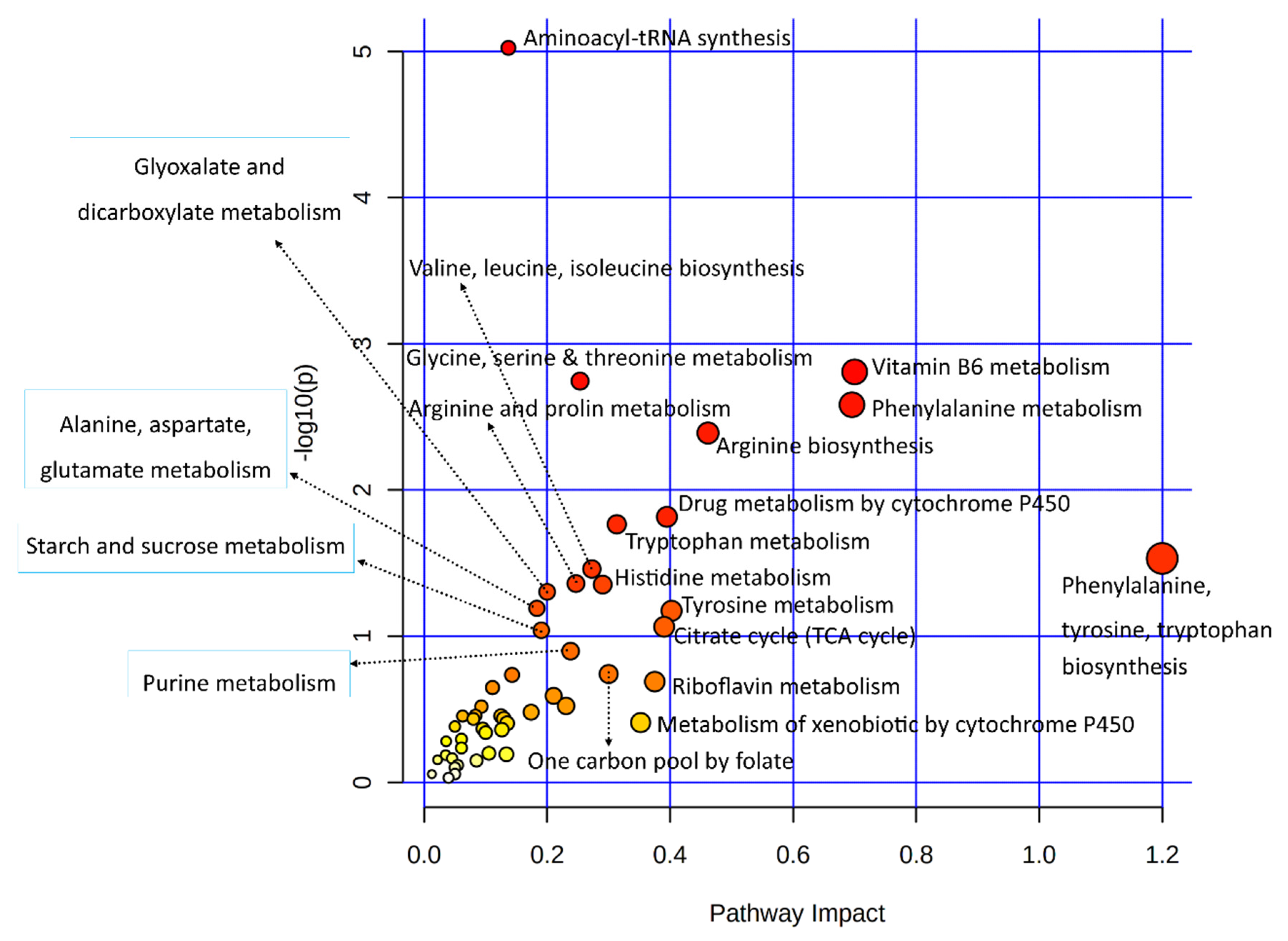
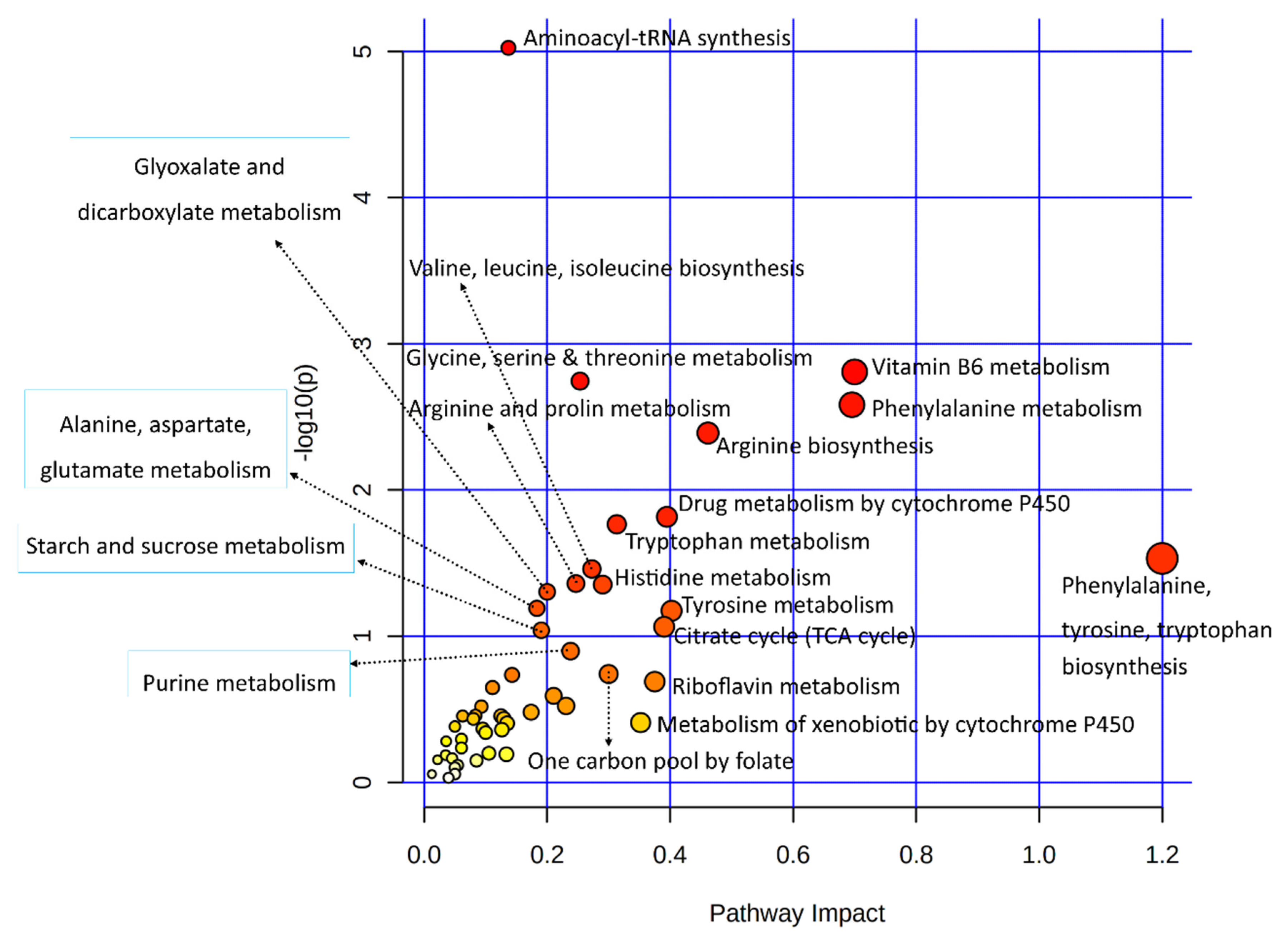
3.3.3. Joint Metabolic Pathway Analysis
3.4. Important Pathways, Metabolites, and Genes in the Joint Pathway Analysis
3.5. Interaction Network Analysis among Transcriptome and Metabolome Data
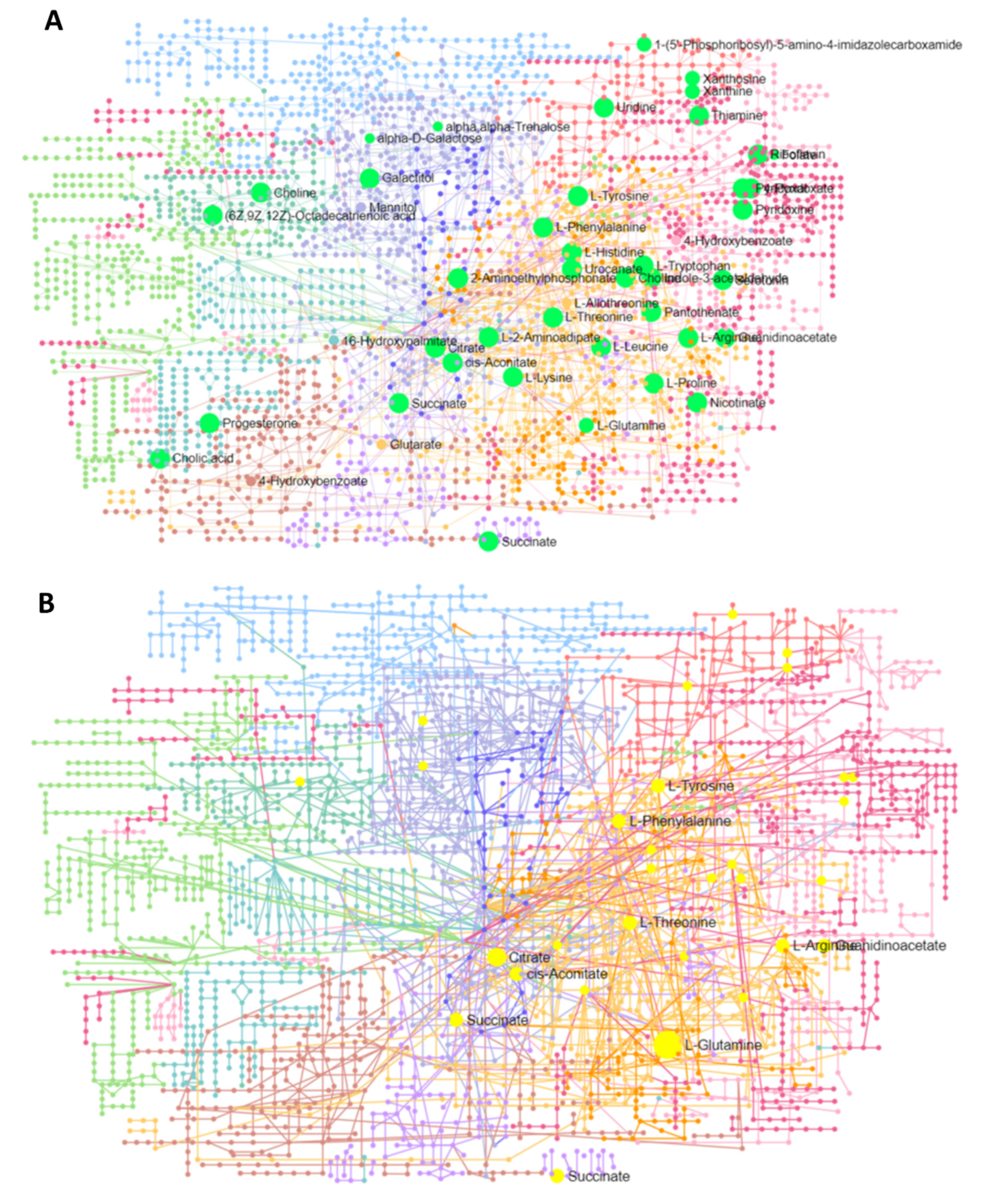
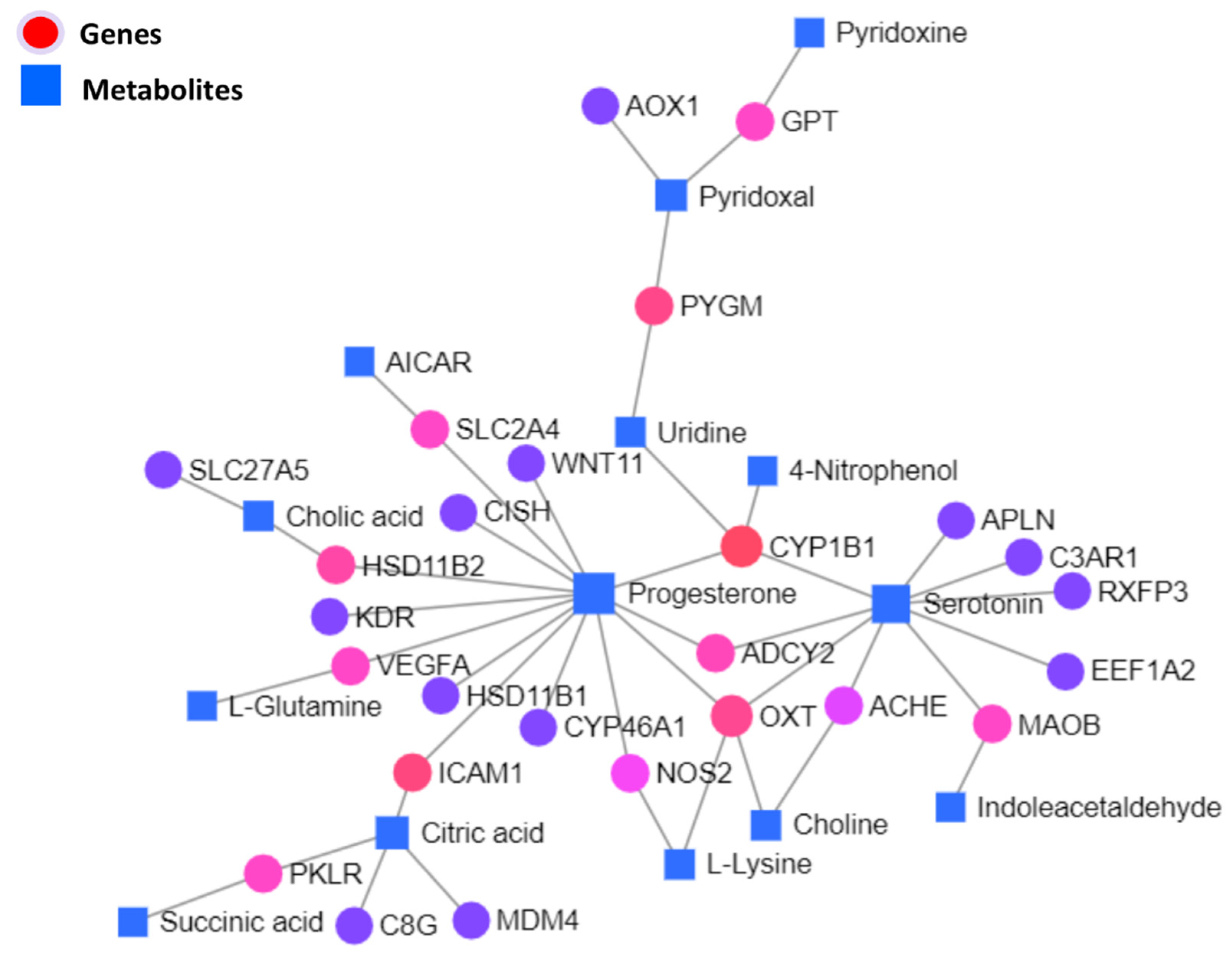
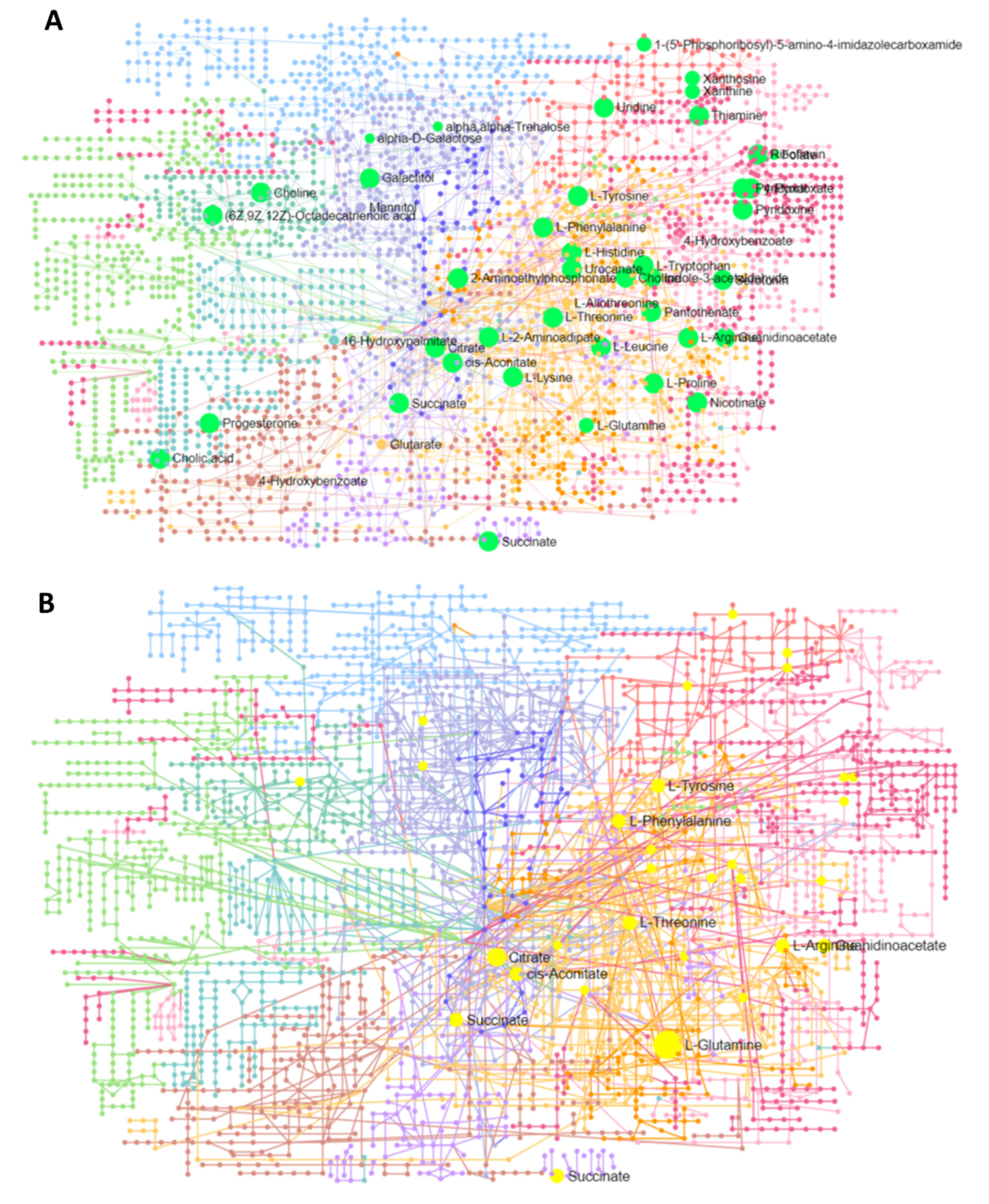
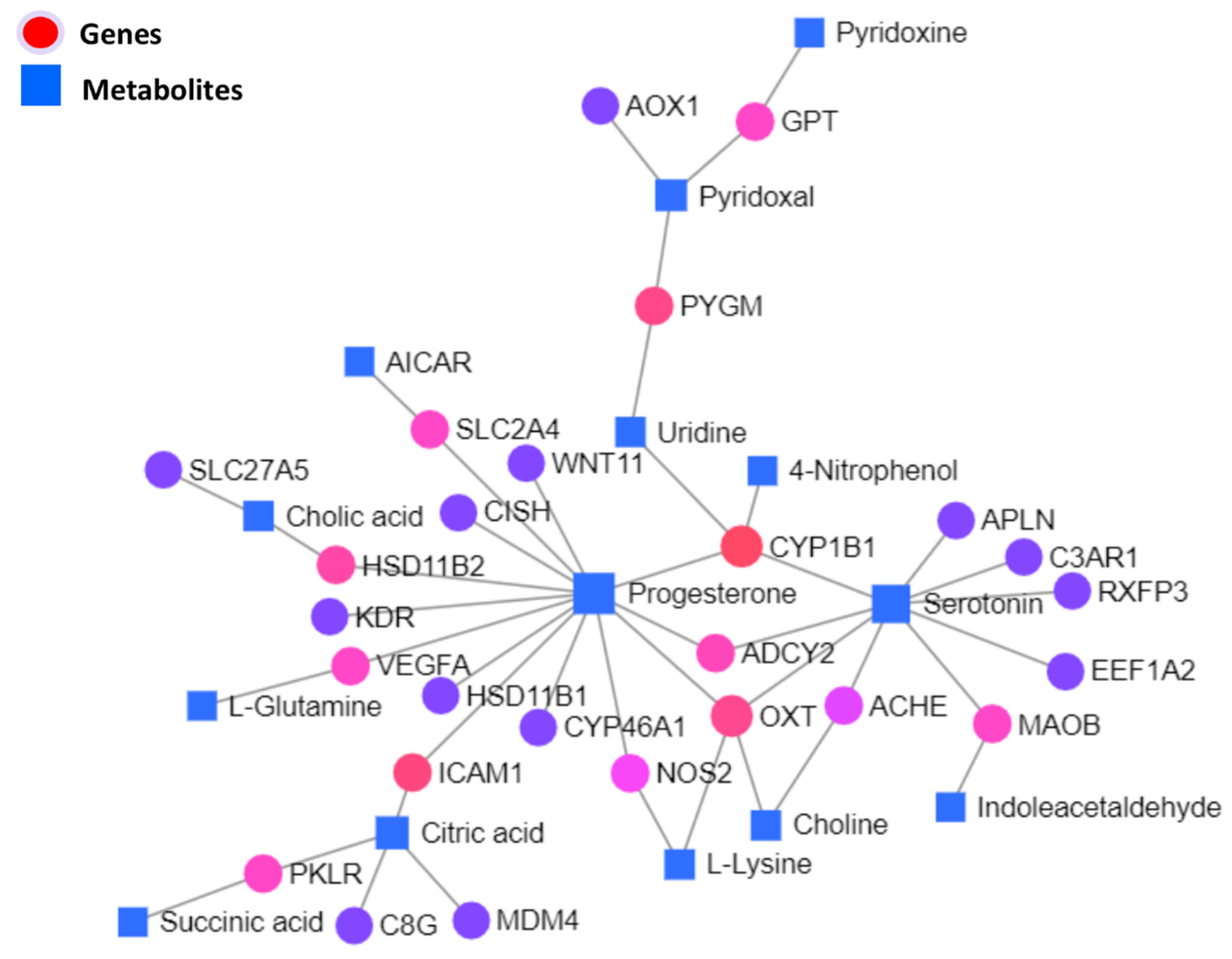
3.5.1. Interaction Network Analysis among Genes and Metabolites
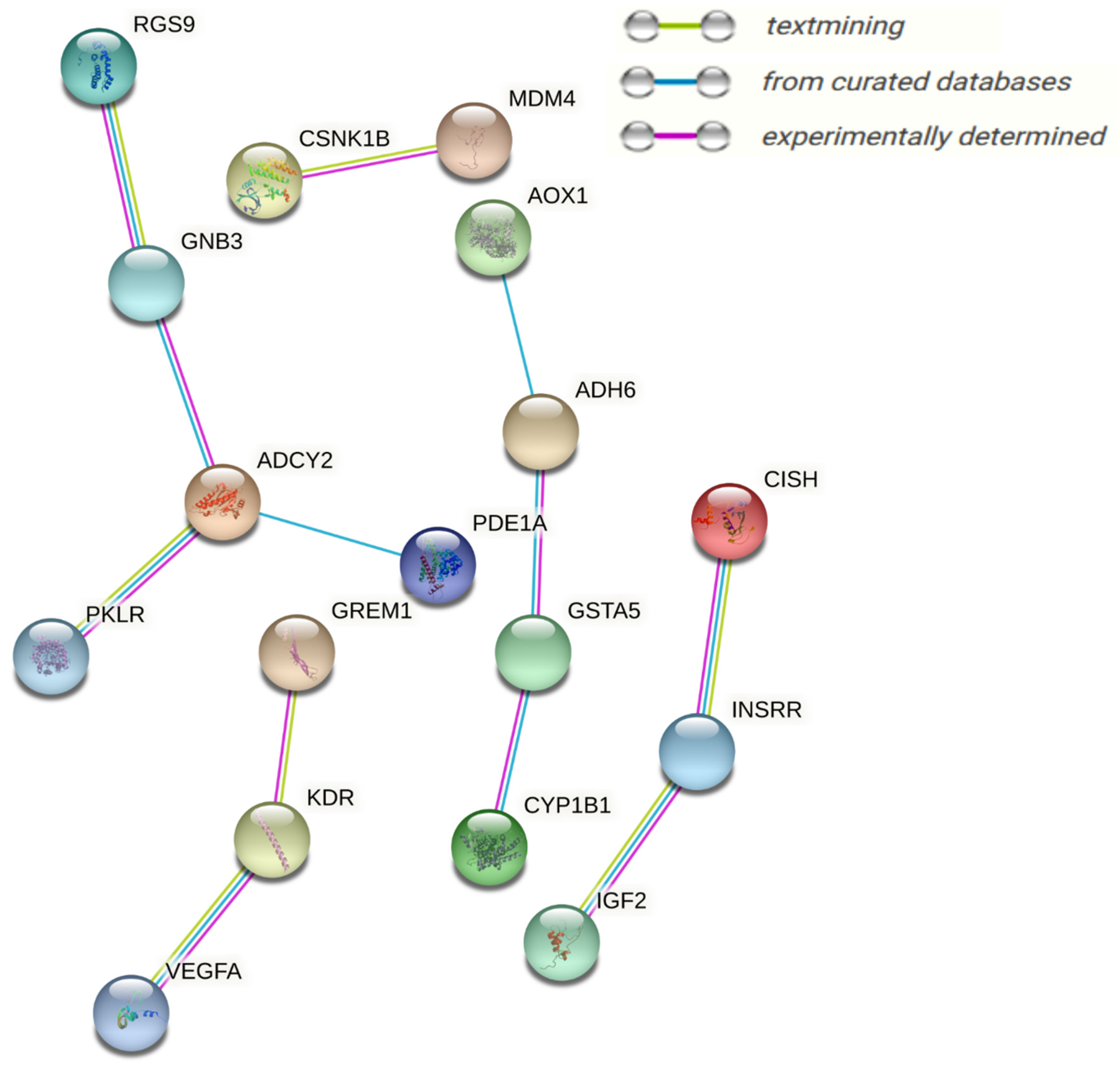
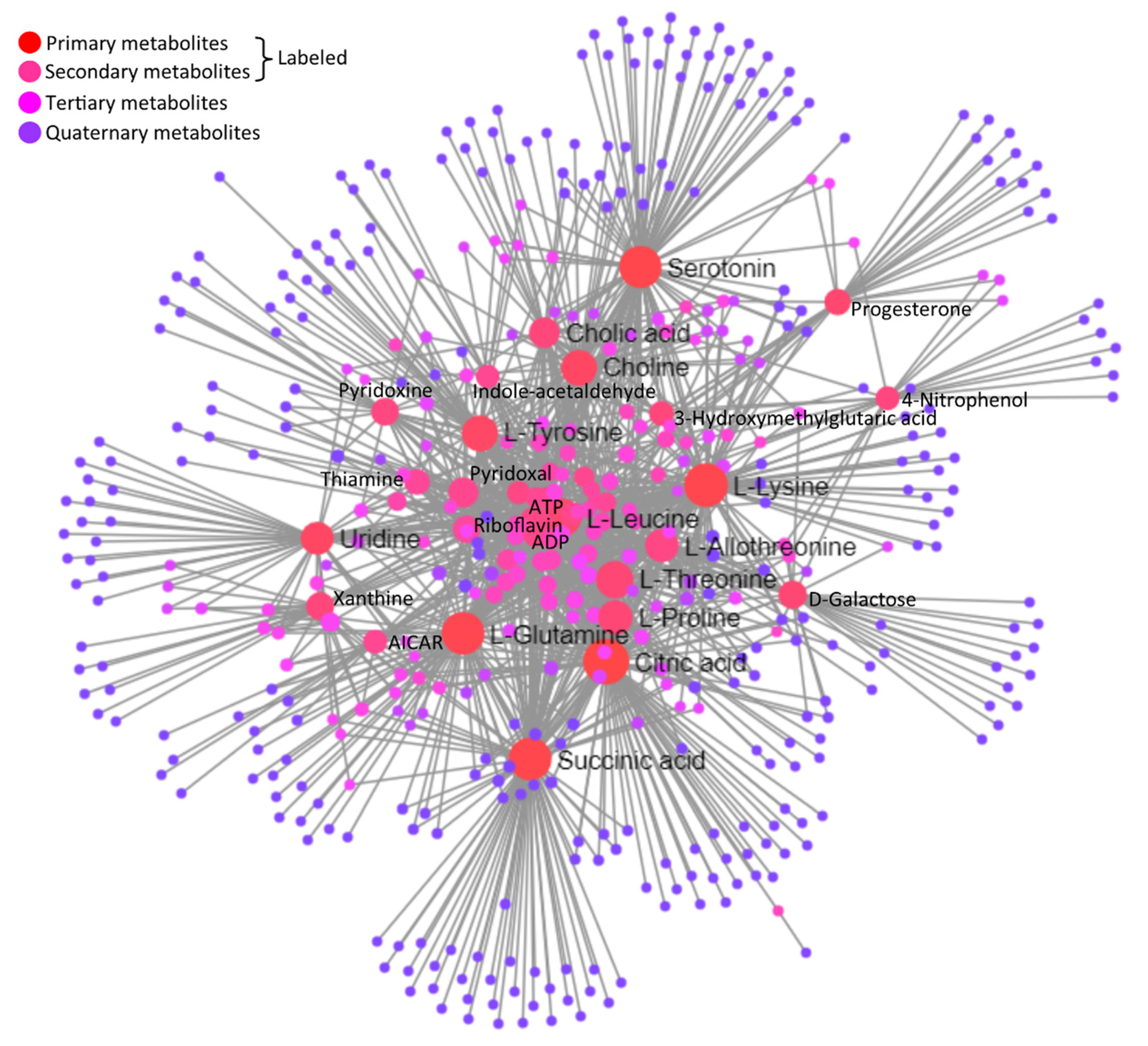
3.5.2. Interaction Network among Metabolites
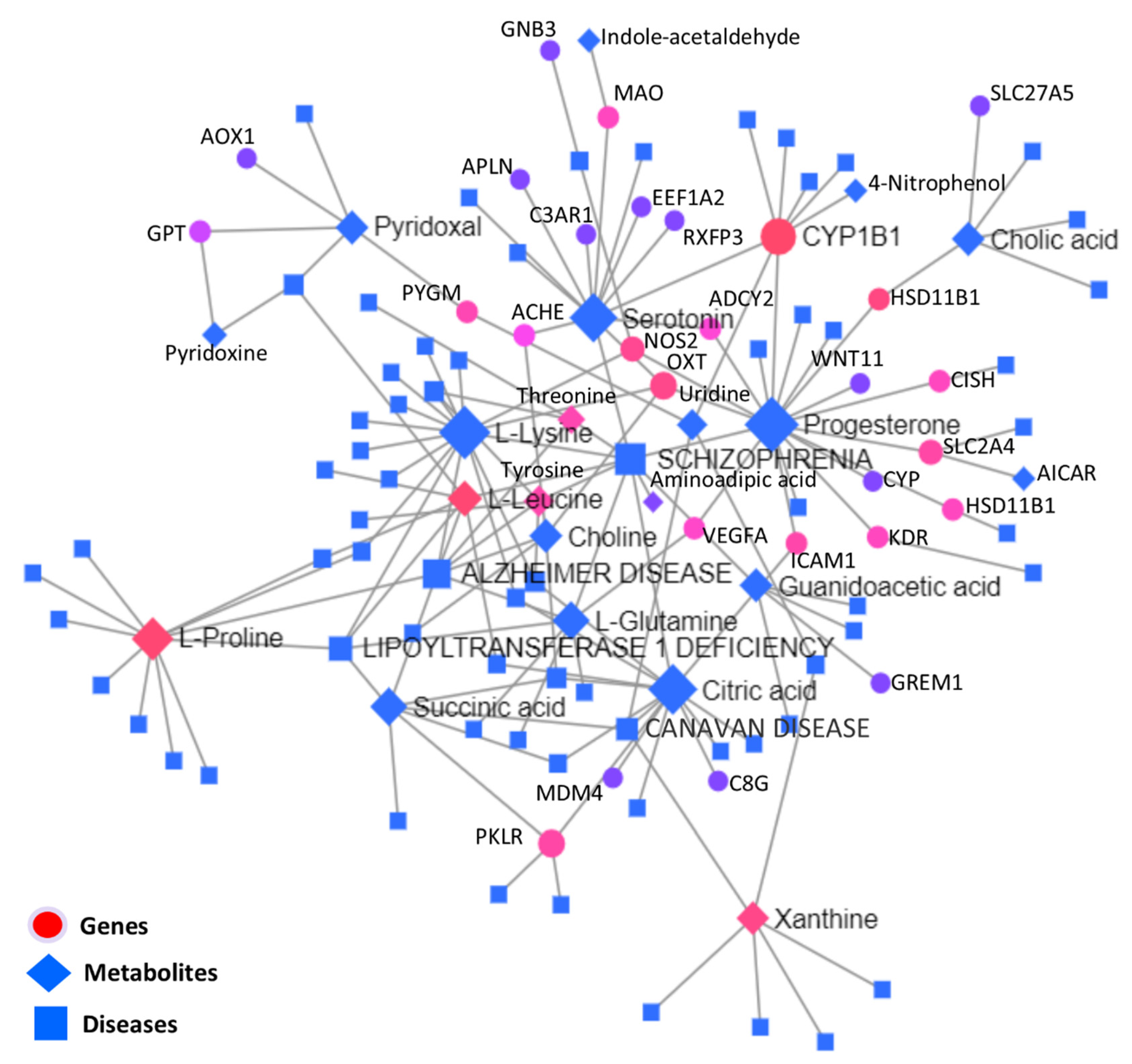
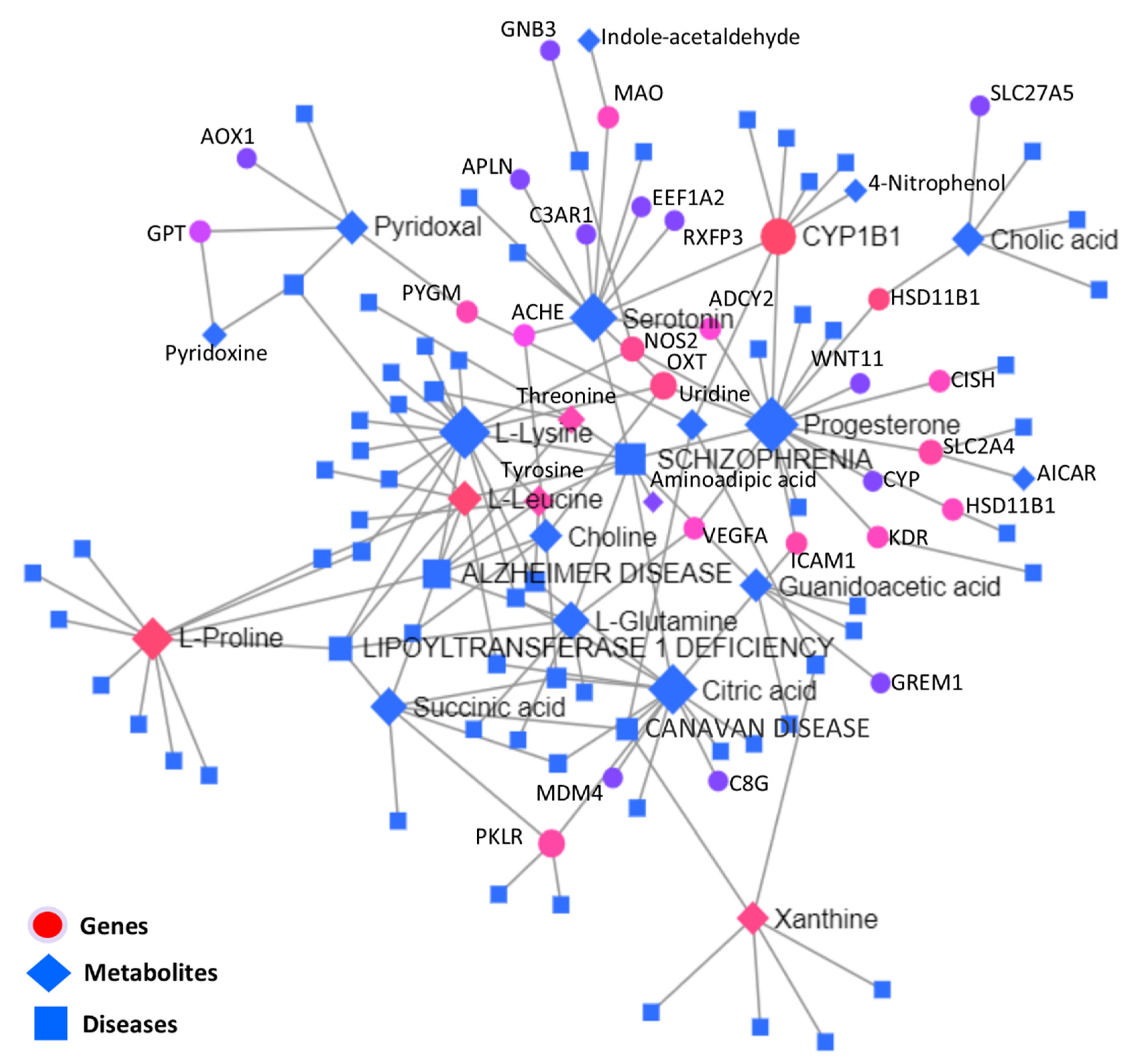
3.5.3. Interactive Network Analysis among Genes, Metabolites, and Diseases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jahromi, B.N.; Mosallanezhad, Z.; Matloob, N.; Davari, M.; Ghobadifar, M.A. The potential role of granulosa cells in the maturation rate of immature human oocytes and embryo development: A co-culture study. Clin. Exp. Reprod. Med. 2015, 42, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Diaz, F.J.; Wigglesworth, K.; Eppig, J.J. Oocytes determine cumulus cell lineage in mouse ovarian follicles. J. Cell Sci. 2007, 120, 1330–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alemu, T.W.; Pandey, H.O.; Salilew Wondim, D.; Gebremedhn, S.; Neuhof, C.; Tholen, E.; Holker, M.; Schellander, K.; Tesfaye, D. Oxidative and endoplasmic reticulum stress defense mechanisms of bovine granulosa cells exposed to heat stress. Theriogenology 2018, 110, 130–141. [Google Scholar] [CrossRef]
- Khan, A.; Dou, J.; Wang, Y.; Jiang, X.; Khan, M.Z.; Luo, H.; Usman, T.; Zhu, H. Evaluation of heat stress effects on cellular and transcriptional adaptation of bovine granulosa cells. J. Anim. Sci. Biotechnol. 2020, 11, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.L.; Liu, D.; Ding, G.R.; Liao, P.F.; Zhang, J.W. Hypoxia-inducible factor-1α and Wnt/β-catenin signaling pathways promote the invasion of hypoxic gastric cancer cells. Mol. Med. Rep. 2015, 12, 3365–3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, Z. Heat stress reduces maturation and developmental capacity in bovine oocytes. Reprod. Fertil. Dev. 2021, 33, 66–75. [Google Scholar] [CrossRef]
- Roth, Z.; Meiden, R.; Braw-Tal, R.; Wolfenson, D. Immediate and delayed effects of heat stress on follicular development and its association with plasma FSH and inhibin concentration in cows. J. Reprod. Fertil. 2000, 120, 83–90. [Google Scholar] [CrossRef]
- Sammad, A.; Khan, M.Z.; Abbas, Z.; Hu, L.; Ullah, Q.; Wang, Y.; Zhu, H.; Wang, Y. Major Nutritional Metabolic Alterations Influencing the Reproductive System of Postpartum Dairy Cows. Metabolites 2022, 12, 60. [Google Scholar] [CrossRef]
- Gaskins, A.J.; Mínguez-Alarcón, L.; VoPham, T.; Hart, J.E.; Chavarro, J.E.; Schwartz, J.; Souter, I.; Laden, F. Impact of ambient temperature on ovarian reserve. Fertil. Steril. 2021, 116, 1052–1060. [Google Scholar] [CrossRef]
- Bridges, P.J.; Brusie, M.A.; Fortune, J.E. Elevated temperature (heat stress) in vitro reduces androstenedione and estradiol and increases progesterone secretion by follicular cells from bovine dominant follicles. Domest. Anim. Endocrinol. 2005, 29, 508–522. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Wheelock, J.B.; Sanders, S.R.; Moore, C.E.; Green, H.B.; Waldron, M.R.; Rhoads, R.P. Postabsorptive carbohydrate adaptations to heat stress and monensin supplementation in lactating Holstein cows1. J. Dairy Sci. 2011, 94, 5620–5633. [Google Scholar] [CrossRef] [PubMed]
- Sammad, A.; Umer, S.; Shi, R.; Zhu, H.; Zhao, X.; Wang, Y. Dairy cow reproduction under the influence of heat stress. J. Anim. Physiol. Anim. Nutr. 2020, 104, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.K.; Jeong, J.K.; Choi, I.S.; Kang, H.G.; Hur, T.Y.; Jung, Y.H.; Kim, I.H. Relationships among ketosis, serum metabolites, body condition, and reproductive outcomes in dairy cows. Theriogenology 2015, 84, 252–260. [Google Scholar] [CrossRef]
- Leroy, J.L.M.R.; Vanholder, T.; Mateusen, B.; Christophe, A.; Opsomer, G.; de Kruif, A.; Genicot, G.; Van Soom, A. Non-esterified fatty acids in follicular fluid of dairy cows and their effect on developmental capacity of bovine oocytes in vitro. Reproduction 2005, 130, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Bromfield, J.J.; Sheldon, I.M. Lipopolysaccharide initiates inflammation in bovine granulosa cells via the TLR4 pathway and perturbs oocyte meiotic progression in vitro. Endocrinology 2011, 152, 5029–5040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shehab-El-Deen, M.A.M.M.; Leroy, J.L.M.R.; Fadel, M.S.; Saleh, S.Y.A.; Maes, D.; Van Soom, A. Biochemical changes in the follicular fluid of the dominant follicle of high producing dairy cows exposed to heat stress early post-partum. Anim. Reprod. Sci. 2010, 117, 189–200. [Google Scholar] [CrossRef]
- Paes, V.M.; Vieira, L.A.; Correia, H.H.V.; Sa, N.A.R.; Moura, A.A.A.; Sales, A.D.; Rodrigues, A.P.R.; Magalhães-Padilha, D.M.; Santos, F.W.; Apgar, G.A.; et al. Effect of heat stress on the survival and development of in vitro cultured bovine preantral follicles and on in vitro maturation of cumulus–oocyte complex. Theriogenology 2016, 86, 994–1003. [Google Scholar] [CrossRef]
- Vanholder, T.; Leroy, J.L.M.R.; Van Soom, A.; Opsomer, G.; Maes, D.; Coryn, M.; De Kruif, A. Effect of non-esterified fatty acids on bovine granulosa cell steroidogenesis and proliferation in vitro. Anim. Reprod. Sci. 2005, 87, 33–44. [Google Scholar] [CrossRef]
- Saadeldin, I.M.; Swelum, A.A.A.; Elsafadi, M.; Mahmood, A.; Osama, A.; Shikshaky, H.; Alfayez, M.; Alowaimer, A.N.; Magdeldin, S. Thermotolerance and plasticity of camel somatic cells exposed to acute and chronic heat stress. J. Adv. Res. 2020, 22, 105–118. [Google Scholar] [CrossRef]
- Saadeldin, I.M.; Swelum, A.A.A.; Elsafadi, M.; Mahmood, A.; Alfayez, M.; Alowaimer, A.N. Differences between the tolerance of camel oocytes and cumulus cells to acute and chronic hyperthermia. J. Therm. Biol. 2018, 74, 47–54. [Google Scholar] [CrossRef]
- Sammad, A.; Luo, H.; Hu, L.; Zhu, H.; Wang, Y. Transcriptomics Reveals Granulosa Cell’s Coping through Redox, Inflammatory, Metabolic and Cytoskeleton Mechanisms under Acute Heat Stress. Cells 2022, 11, 1443. [Google Scholar] [CrossRef] [PubMed]
- Sammad, A.; Hu, L.; Luo, H.; Abbas, Z.; Umer, S.; Zhao, S.; Xu, Q.; Khan, A.; Wang, Y.; Zhu, H.; et al. Investigation of Metabolome Underlying the Biological Mechanisms of Acute Heat Stressed Granulosa Cells. Int. J. Mol. Sci. 2022, 23, 2146. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khan, M.Z.; Dou, J.; Umer, S.; Xu, H.; Sammad, A.; Zhu, H.B.; Wang, Y. RNAi-Mediated Silencing of Catalase Gene Promotes Apoptosis and Impairs Proliferation of Bovine Granulosa Cells under Heat Stress. Animals 2020, 10, 1060. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Hansen, P.J. Embryonic mortality in cattle from the embryo’s perspective. J. Anim. Sci. 2002, 80, E33–E44. [Google Scholar] [CrossRef]
- Destaw, W.M.; Kefyalew, A.W. Evaluation of the reproductive performance of Holstein Friesian dairy cows in Alage ATVET college, Ethiopia. Int. J. Livest. Prod. 2018, 9, 131–139. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, C.; Elsheikh, N.A.H.; Li, C.; Yang, F.; Wang, G.; Li, L. HO-1 reduces heat stress-induced apoptosis in bovine granulosa cells by suppressing oxidative stress. Aging 2019, 11, 5535–5547. [Google Scholar] [CrossRef]
- Fu, Y.; He, C.-J.; Ji, P.-Y.; Zhuo, Z.-Y.; Tian, X.-Z.; Wang, F.; Tan, D.-X.; Liu, G.-S. Effects of Melatonin on the Proliferation and Apoptosis of Sheep Granulosa Cells under Thermal Stress. Int. J. Mol. Sci. 2014, 15, 21090–21104. [Google Scholar] [CrossRef] [Green Version]
- Abdelnour, S.A.; Swelum, A.A.; Abd El-Hack, M.E.; Khafaga, A.F.; Taha, A.E.; Abdo, M. Cellular and functional adaptation to thermal stress in ovarian granulosa cells in mammals. J. Therm. Biol. 2020, 92. [Google Scholar] [CrossRef] [PubMed]
- Abbas, Z.; Sammad, A.; Hu, L.; Fang, H.; Xu, Q.; Wang, Y. Glucose metabolism and dynamics of facilitative glucose transporters (Gluts) under the influence of heat stress in dairy cattle. Metabolites 2020, 10, 312. [Google Scholar] [CrossRef] [PubMed]
- Sammad, A.; Wang, Y.J.; Umer, S.; Lirong, H.; Khan, I.; Khan, A.; Ahmad, B.; Wang, Y. Nutritional Physiology and Biochemistry of Dairy Cattle under the Influence of Heat Stress: Consequences and Opportunities. Animals 2020, 10, 793. [Google Scholar] [CrossRef] [PubMed]
- Soto, P.; Smith, L.C. BH4 peptide derived from Bcl-xL and Bax-inhibitor peptide suppresses apoptotic mitochondrial changes in heat stressed bovine oocytes. Mol. Reprod. Dev. 2009, 76, 637–646. [Google Scholar] [CrossRef]
- Abdelnour, S.A.; Abd El-Hack, M.E.; Khafaga, A.F.; Arif, M.; Taha, A.E.; Noreldin, A.E. Stress biomarkers and proteomics alteration to thermal stress in ruminants: A review. J. Therm. Biol. 2019, 79, 120–134. [Google Scholar] [CrossRef]
- Guérin, P.; El Mouatassim, S.; Ménézo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef]
- Zhang, X.-Y.; Xiong, Y.-M.; Tan, Y.-J.; Wang, L.; Li, R.; Zhang, Y.; Liu, X.-M.; Lin, X.-H.; Jin, L.; Hu, Y.-T.; et al. Melatonin rescues impaired penetration ability of human spermatozoa induced by mitochondrial dysfunction. Reproduction 2019, 158, 465–475. [Google Scholar] [CrossRef]
- Pedruzzi, L.M.; Stockler-Pinto, M.B.; Leite, M.; Mafra, D. Nrf2–keap1 system versus NF-κB: The good and the evil in chronic kidney disease? Biochimie 2012, 94, 2461–2466. [Google Scholar] [CrossRef]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, S.; Summers, C.M.; Pearce, S.C.; Gabler, N.K.; Valentine, R.J.; Baumgard, L.H.; Rhoads, R.P.; Selsby, J.T. Short-term heat stress altered metabolism and insulin signaling in skeletal muscle. J. Anim. Sci. 2018, 96, 154–167. [Google Scholar] [CrossRef] [Green Version]
- Hambright, H.G.; Meng, P.; Kumar, A.P.; Ghosh, R. Inhibition of PI3K/AKT/mTOR axis disrupts oxidative stress-mediated survival of melanoma cells. Oncotarget 2015, 6, 7195–7208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, N.M.; Potteti, H.R.; Vegiraju, S.; Chen, H.J.; Tamatam, C.M.; Reddy, S.P. PI3K-AKT Signaling via Nrf2 Protects against Hyperoxia-Induced Acute Lung Injury, but Promotes Inflammation Post-Injury Independent of Nrf2 in Mice. PLoS ONE 2015, 10, e0129676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantó, C.; Auwerx, J. PGC-1α, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipidol. 2009, 20, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef]
- Li, Z.Y.; Chen, Z.L.; Zhang, T.; Wei, C.; Shi, W.Y. TGF-β and NF-κB signaling pathway crosstalk potentiates corneal epithelial senescence through an RNA stress response. Aging 2016, 8, 2337–2354. [Google Scholar] [CrossRef] [Green Version]
- Tominaga, K.; Suzuki, H.I. TGF-β signaling in cellular senescence and aging-related pathology. Int. J. Mol. Sci. 2019, 20, 5002. [Google Scholar] [CrossRef] [Green Version]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef] [Green Version]
- Dean, M.; Annilo, T. Evolution of the ATP-binding cassette (ABC) transporter superfamily in vertebrates. Annu. Rev. Genomics Hum. Genet. 2005, 6, 123–142. [Google Scholar] [CrossRef]
- He, F.; Antonucci, L.; Karin, M. NRF2 as a regulator of cell metabolism and inflammation in cancer. Carcinogenesis 2020, 41, 405–416. [Google Scholar] [CrossRef]
- Signorile, A.; Ferretta, A.; Ruggieri, M.; Paolicelli, D.; Lattanzio, P.; Trojano, M.; De Rasmo, D. Mitochondria, oxidative stress, camp signalling and apoptosis: A crossroads in lymphocytes of multiple sclerosis, a possible role of nutraceutics. Antioxidants 2021, 10, 21. [Google Scholar] [CrossRef]
- Lian, J.; Yue, Y.; Yu, W.; Zhang, Y. Immunosenescence: A key player in cancer development. J. Hematol. Oncol. 2020, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Blagosklonny, M.V. Cell cycle arrest is not yet senescence, which is not just cell cycle arrest: Terminology for TOR-driven aging. Aging 2012, 4, 159–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.; Levine, A.J. The regulation of energy metabolism and the IGF-1/mTOR pathways by the p53 protein. Trends Cell Biol. 2010, 20, 427–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihaylova, M.M.; Shaw, R.J. The AMP-activated protein kinase (AMPK) signaling pathway coordinates cell growth, autophagy, & metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [PubMed]
- Rhoads, M.L.; Rhoads, R.P.; VanBaale, M.J.; Collier, R.J.; Sanders, S.R.; Weber, W.J.; Crooker, B.A.; Baumgard, L.H. Effects of heat stress and plane of nutrition on lactating Holstein cows: I. Production, metabolism, and aspects of circulating somatotropin. J. Dairy Sci. 2009, 92, 1986–1997. [Google Scholar] [CrossRef] [Green Version]
- Rhoads, R.P.; La Noce, A.J.; Wheelock, J.B.; Baumgard, L.H. Short communication: Alterations in expression of gluconeogenic genes during heat stress and exogenous bovine somatotropin administration. J. Dairy Sci. 2011, 94, 1917–1921. [Google Scholar] [CrossRef]
- Wang, J.P.; Bu, D.P.; Wang, J.Q.; Huo, X.K.; Guo, T.J.; Wei, H.Y.; Zhou, L.Y.; Rastani, R.R.; Baumgard, L.H.; Li, F.D. Effect of saturated fatty acid supplementation on production and metabolism indices in heat-stressed mid-lactation dairy cows. J. Dairy Sci. 2010, 93, 4121–4127. [Google Scholar] [CrossRef] [Green Version]
- Migocka-Patrzałek, M.; Elias, M. Muscle glycogen phosphorylase and its functional partners in health and disease. Cells 2021, 10, 883. [Google Scholar] [CrossRef]
- De Groote, M.L.; Kazemier, H.G.; Huisman, C.; Van Der Gun, B.T.F.; Faas, M.M.; Rots, M.G. Upregulation of endogenous ICAM-1 reduces ovarian cancer cell growth in the absence of immune cells. Int. J. Cancer 2014, 134, 280–290. [Google Scholar] [CrossRef]
- DeNardo, G.L.; DeNardo, S.J. Update: Turning the Heat on Cancer. Cancer Biotherapy Radiopharm. 2009, 23, 671–679. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Liu, M.; Huang, H.; Li, B.; Zhao, H.; Feng, X.Q.; Zhao, H.P. Heat Stress-Induced Multiple Multipolar Divisions of Human Cancer Cells. Cells 2019, 8, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanamori, T.; Miyazaki, N.; Aoki, S.; Ito, K.; Hisaka, A.; Hatakeyama, H. Investigation of energy metabolic dynamism in hyperthermia-resistant ovarian and uterine cancer cells under heat stress. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Sun, W.; Zhang, C.; Dong, B.; Yang, J.; Hou, M.; Xiong, L.; Cai, B.; Liu, X.; Xue, W. Metabolic Control by Heat Stress Determining Cell Fate to Ferroptosis for Effective Cancer Therapy. ACS Nano 2021, 15, 7179–7194. [Google Scholar] [CrossRef] [PubMed]
- Payen, V.L.; Mina, E.; Van Hée, V.F.; Porporato, P.E.; Sonveaux, P. Monocarboxylate transporters in cancer. Mol. Metab. 2020, 33. [Google Scholar] [CrossRef]
- Yu, S.; Wu, Y.; Li, C.; Qu, Z.; Lou, G.; Guo, X.; Ji, J.; Li, N.; Guo, M.; Zhang, M.; et al. Comprehensive analysis of the SLC16A gene family in pancreatic cancer via integrated bioinformatics. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Wong, R.S.Y. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Kumari, R.; Jat, P. Mechanisms of Cellular Senescence: Cell Cycle Arrest and Senescence Associated Secretory Phenotype. Front. Cell Dev. Biol. 2021, 9, 485. [Google Scholar] [CrossRef]
- Meulmeester, E.; Jochemsen, A. p53: A Guide to Apoptosis. Curr. Cancer Drug Targets 2008, 8, 87–97. [Google Scholar] [CrossRef]
- Rayess, H.; Wang, M.B.; Srivatsan, E.S. Cellular senescence and tumor suppressor gene p16. Int. J. Cancer 2012, 130, 1715–1725. [Google Scholar] [CrossRef] [Green Version]
- Hashim, N.A.A.; Ab-Rahim, S.; Suddin, L.S.; Saman, M.S.A.; Mazlan, M. Global serum metabolomics profiling of colorectal cancer. Mol. Clin. Oncol. 2019, 11, 3–14. [Google Scholar]
- Vermeulen, K.; Berneman, Z.N.; Van Bockstaele, D.R. Cell cycle and apoptosis. Cell Prolif. 2003, 36, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Menendez, D.; Inga, A.; Resnick, M.A. The expanding universe of p53 targets. Nat. Rev. Cancer 2009, 9, 724–737. [Google Scholar] [CrossRef] [PubMed]
- Vousden, K.H.; Prives, C. Blinded by the Light: The Growing Complexity of p53. Cell 2009, 137, 413–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haley, V.L.; Barnes, D.J.; Sandovici, I.; Constancia, M.; Graham, C.F.; Pezzella, F.; Bühnemann, C.; Carter, E.J.; Hassan, A.B. Igf2 pathway dependency of the Trp53 developmental and tumour phenotypes. EMBO Mol. Med. 2012, 4, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.L.; Wang, H.L.; Jiang, L.Z.; Qian, Y.; Yang, C.X.; Chang, W.W.; Zhong, J.F.; Xing, G.D. Heat stress induces apoptosis through disruption of dynamic mitochondrial networks in dairy cow mammary epithelial cells. Vitr. Cell. Dev. Biol.-Anim. 2020, 56, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Sammad, A.; Luo, H.; Hu, L.; Zhu, H.; Wang, Y. Transcriptome Reveals Granulosa Cells Coping through Redox, Inflammatory and Metabolic Mechanisms under Acute Heat Stress. Cells 2022, 11, 1443. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathways | Total /Hits | Raw p-Value | Holm Adjust p-Value |
|---|---|---|---|
| ABC transporters | 198/18 | 5.1 × 10−11 | 1.7 × 10−8 |
| Central carbon metabolism in cancer | 103/11 | 6.5 × 10−8 | 2.1 × 10−5 |
| Aminoacyl-tRNA biosynthesis | 118/10 | 2.3 × 10−6 | 7.4 × 10−4 |
| Protein digestion and absorption | 168/11 | 8.7 × 10−6 | 2.8 × 10−3 |
| Glycine, serine, and threonine metabolism | 93/8 | 2.2 × 10−5 | 7.0 × 10−3 |
| Mineral absorption | 84/6 | 6.5 × 10−4 | 2.1 × 10−1 |
| Vitamin B6 metabolism | 36/4 | 1.0 × 10−3 | 3.3 × 10−1 |
| Vitamin digestion and absorption | 65/5 | 1.3 × 10−3 | 4.3 × 10−1 |
| Relaxin signaling pathway | 136/7 | 1.6 × 10−3 | 5.3 × 10−1 |
| Dilated cardiomyopathy (DCM) | 102/6 | 1.8 × 10−3 | 5.7 × 10−1 |
| Arginine biosynthesis | 42/4 | 1.9 × 10−3 | 5.9 × 10−1 |
| Taste transduction | 112/6 | 2.9 × 10−3 | 9.1 × 10−1 |
| Arrhythmogenic right ventricular cardiomyopathy (ARVC) | 78/5 | 3.0 × 10−3 | 9.4 × 10−1 |
| Tyrosine metabolism | 117/6 | 3.6 × 10−3 | 1.00000 |
| Phenylalanine metabolism | 83/5 | 3.9 × 10−3 | 1.00000 |
| Cocaine addiction | 56/4 | 5.3 × 10−3 | 1.00000 |
| Tryptophan metabolism | 129/6 | 5.8 × 10−3 | 1.00000 |
| Glyoxylate and dicarboxylate metabolism | 92/5 | 6.1 × 10−3 | 1.00000 |
| TGF-beta signaling pathway | 93/5 | 6.3 × 10−3 | 1.00000 |
| Hypertrophic cardiomyopathy (HCM) | 95/5 | 6.9 × 10−3 | 1.00000 |
| Axon guidance | 180/7 | 7.7 × 10−3 | 1.00000 |
| GABAergic synapse | 99/5 | 8.2 × 10−3 | 1.00000 |
| Alanine, aspartate and glutamate metabolism | 65/4 | 9.0 × 10−3 | 1.00000 |
| Apelin signaling pathway | 149/6 | 1.1 × 10−2 | 1.00000 |
| Histidine metabolism | 70/4 | 1.2 × 10−2 | 1.00000 |
| Joint Pathways | Total /Hits | Raw p-Value | Holm Adjust p-Value | Metabolites | Genes |
|---|---|---|---|---|---|
| Top functional pathways from the integrated analysis | |||||
| ABC transporters | 198/18 | 5.11 × 1011 | 1.67 × 10−8 | ↓Alpha-Trehalose; ↓D-Mannitol; ↑L-Lysine; ↑L-Arginine; ↑L-Glutamine; ↓L-Histidine; ↑L-Leucine; ↓L-Threonine; ↓Proline; ↑Choline; ↓Thiamine; ↓Ciliatine; ↓L-Phenylalanine; ↓Riboflavin; ↓Uridine; ↓Xanthosine | ↓ABCC5; ↓ABCA2; |
| Protein digestion and absorption | 168/11 | 8.70 × 10−6 | 0.002819 | ↑L-Leucine; ↓L-Phenylalanine; ↓L-Tryptophan; ↓L-Threonine; ↑L-Glutamine; ↑L-Arginine; ↑L-Lysine; ↓L-Histidine; ↓Proline; ↑L-Tyrosine | ↓COL11A2 |
| Taste transduction | 112/6 | 0.0028879 | 0.91259 | ↓Serotonin; ↑Citric Acid | ↓SCN9A; ↓PDE1A; ↓GABBR1; ↓GNB3 |
| Cocaine addiction | 56/4 | 0.0053408 | 1.00000 | ↑L-Tyrosine; | ↓MAOB; ↓RGS9; ↓GRIN2D |
| GABAergic synapse | 99/5 | 0.0082127 | 1.00000 | ↑L-Glutamine; ↓Succinic Acid | ↓GABBR1; ↓ADCY2; ↓GNB3 |
| Functional signaling pathways (manual query of integrated analysis) | |||||
| cAMP signaling pathway | 254/8 | 0.015042 | 1.00000 | ↓Serotonin; ↓Succinic Acid | ↓RYR2; ↓ATP2A1; ↓GRIN2D; ↓GABBR1; ↓ADCY2; ↓OXT |
| Ovarian steroidogenesis | 78/4 | 0.016783 | 1.00000 | ↓Progesterone | ↓ADCY2; ↓CYP1B1 ↓PLA2G4B |
| mTOR signaling pathway | 158/4 | 0.1386 | 1.00000 | ↑L-Leucine; ↑L-Arginine | ↓DEPTOR; ↓WNT11 |
| AMPK signaling pathway | 146/2 | 0.5515 | 1.00000 | ↓AICAR | ↑SLC2A4 |
| Cancer pathways (manual query of integrated analysis) | |||||
| Central carbon metabolism in cancer | 103/11 | 6.5 × 10−8 | 2.1 × 10−5 | ↑L-Glutamine; ↑Citric Acid; ↓Succinic Acid; ↑L-Leucine; ↓L-Phenylalanine; ↓L-Histidine; ↓L-Tryptophan; ↑L-Tyrosine; ↓Proline; ↑L-Arginine | ↑SLC16A3 |
| Breast cancer | 153/3 | 0.30314 | 1.00000 | ↓Progesterone | ↓CSNK1B; ↓WNT11 |
| Prostate cancer | 109/2 | 0.40000 | 1.00000 | ↓Progesterone | ↓INSRR |
| Pathways in cancer | 573/8 | 0.43410 | 1.00000 | ↓Progesterone | ↓WNT11; ↓IGF2; ↑NOS2; ↓VEGFA; ↓ADCY2; ↓GNB3; ↑GSTA5 |
| Metabolic Pathways | Total /Hits | Raw p-Value | Holm Adjust p-Value | Metabolites | Genes |
|---|---|---|---|---|---|
| Vitamin B6 metabolism | 21/4 | 0.00156 | 0.12918 | ↑Pyridoxine; ↓Pyridoxal; ↑4-Pyridoxic Acid | ↑AOX1 |
| Glycine, serine, and threonine metabolism | 72/7 | 0.00180 | 0.14759 | ↑Choline; ↓Glycocyamine; ↓L-Threonine; ↑L-Allo-Threonine; | ↓MAOB; ↓AOC2; ↓AMT |
| Phenylalanine metabolism | 24/4 | 0.00261 | 0.21157 | ↓L-Phenylalanine; ↑L-Tyrosine; | ↓AOC2; ↓MAOB |
| Arginine biosynthesis | 27/4 | 0.00408 | 0.32625 | ↑L-Arginine; ↑L-Glutamine; | ↑NOS2; ↓GPT |
| Tryptophan metabolism | 84/6 | 0.017185 | 1.00000 | ↓L-Tryptophan; ↓Serotonin; ↑Indole-3-Acetaldehyde; | ↓CYP1B1 ↓MAOB; ↑AOX1 |
| Arginine and proline metabolism | 78/5 | 0.04371 | 1.00000 | ↑L-Arginine; ↓Glycocyamine; ↓Proline | ↑NOS2; ↓MAOB |
| Histidine metabolism | 32/3 | 0.04444 | 1.00000 | ↓Urocanic Acid; ↓L-Histidine | ↓MAOB |
| Glyoxylate and dicarboxylate metabolism | 56/4 | 0.04982 | 1.00000 | ↓Cis-Aconitic Acid; ↑Citric Acid; ↑L-Glutamine | ↓AMT |
| Alanine, aspartate and glutamate metabolism | 61/4 | 0.06459 | 1.00000 | ↑L-Glutamine; ↑Citric Acid; ↓Succinic Acid | ↓GPT |
| Tyrosine metabolism | 88/5 | 0.06715 | 1.00000 | ↑L-Tyrosine | ↓MAOB; ↑ADH6; ↓AOC2; ↑AOX1 |
| Starch and sucrose metabolism | 43/3 | 0.09127 | 1.00000 | ↓Alpha-Trehalose | ↓AMY2B; ↑PYGM |
| Purine metabolism | 169/7 | 0.12673 | 1.00000 | ↓Xanthine; ↑L-Glutamine; ↓AICAR; ↓Xanthosine | ↓PDE1A; ↓ADCY2; ↓PKLR |
| One carbon pool by folate | 31/2 | 0.18155 | 1.00000 | ↑Folic Acid | ↓AMT |
| Id | Label | Regulation Status | Degree | Betweenness |
|---|---|---|---|---|
| C00410 | Progesterone | Down | 13 | 544.42 |
| C00780 | Serotonin | Down | 9 | 254.42 |
| 1545 | CYP1B1 | Down | 4 | 281 |
| C00158 | Citric acid | Up | 4 | 145 |
| 5020 | OXT | Down | 4 | 112 |
| C00250 | Pyridoxal | Down | 3 | 110 |
| 3383 | ICAM1 | Up | 2 | 170 |
| C00299 | Uridine | Down | 2 | 170 |
| 5837 | PYGM | Up | 2 | 140 |
| 3291 | HSD11B2 | Down | 2 | 74 |
| 108 | ADCY2 | Down | 2 | 51 |
| 4129 | MAOB | Down | 2 | 38 |
| 7422 | VEGFA | Down | 2 | 38 |
| 5313 | PKLR | Down | 2 | 38 |
| 6517 | SLC2A4 | Up | 2 | 38 |
| 2875 | GPT | Down | 2 | 38 |
| C00695 | Cholic acid | Down | 2 | 38 |
| 4843 | NOS2 | Up | 2 | 12.5 |
| 43 | ACHE | Down | 2 | 6.5 |
| C00114 | Choline | Up | 2 | 6.08 |
| C00047 | L-Lysine | Up | 2 | 3.08 |
| Id | Label | Regulation Status | Degree | Betweenness |
|---|---|---|---|---|
| C00158 | Citric acid | Up | 131 | 26,439.49 |
| C00047 | L-Lysine | Up | 112 | 17,872.77 |
| C00042 | Succinic acid | Down | 105 | 20,430.26 |
| C00064 | L-Glutamine | Up | 105 | 17,053.91 |
| C00780 | Serotonin | Down | 101 | 20,433.61 |
| C00123 | L-Leucine | Up | 76 | 7023.93 |
| C00188 | L-Threonine | Down | 72 | 5723.68 |
| C00082 | L-Tyrosine | Up | 65 | 9222.24 |
| C00114 | Choline | Up | 64 | 9738.79 |
| C00148 | L-Proline | Down | 56 | 5378.26 |
| C05519 | L-Allo-threonine | Up | 52 | 3135.83 |
| C00299 | Uridine | Down | 50 | 9847.77 |
| C00695 | Cholic acid | Down | 41 | 4475.47 |
| C00250 | Pyridoxal | Down | 39 | 2385.19 |
| C00984 | D-Galactose | Down | 33 | 6225.15 |
| Id | Disease | Degree | b/w | Metabolites | Genes |
|---|---|---|---|---|---|
| 181500 | SCHIZOPHRENIA | 10 | 1086 | ↓Progesterone, ↑L-Lysine, ↓Serotonin, ↑Citric acid, ↑L-Glutamine, ↓Threonine, ↓Aminoadipic acid | ↓VEGFA |
| 104300 | ALZHEIMER DISEASE | 8 | 542 | ↑L-Lysine, ↑L-Leucine, ↑L-Glutamine, ↑Choline, ↓L-Proline, ↓Threonine | - |
| 616299 | LIPOYLTRANSFERASE 1 DEFICIENCY | 5 | 286 | ↑L-Lysine, ↑L-Leucine, ↑L-Glutamine, ↓Succinic acid, ↓L-Proline, ↓Threonine | - |
| 271900 | CANAVAN DISEASE | 4 | 453 | ↓Uridine, ↓Xanthine, ↑Citric acid, ↓Succinic acid | - |
| 617290 | EPILEPSY, VITAMIN B6-DEPENDENT | 3 | 203 | ↓Pyridoxal, ↑Pyridoxine, ↑L-Leucine | - |
| 276700 | TYROSINEMIA, TYPE I | 3 | 56 | ↓Threonine, ↑L-Tyrosine, ↑L-Lysine | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sammad, A.; Luo, H.; Hu, L.; Zhao, S.; Gong, J.; Umer, S.; Khan, A.; Zhu, H.; Wang, Y. Joint Transcriptome and Metabolome Analysis Prevails the Biological Mechanisms Underlying the Pro-Survival Fight in In Vitro Heat-Stressed Granulosa Cells. Biology 2022, 11, 839. https://doi.org/10.3390/biology11060839
Sammad A, Luo H, Hu L, Zhao S, Gong J, Umer S, Khan A, Zhu H, Wang Y. Joint Transcriptome and Metabolome Analysis Prevails the Biological Mechanisms Underlying the Pro-Survival Fight in In Vitro Heat-Stressed Granulosa Cells. Biology. 2022; 11(6):839. https://doi.org/10.3390/biology11060839
Chicago/Turabian StyleSammad, Abdul, Hanpeng Luo, Lirong Hu, Shanjiang Zhao, Jianfei Gong, Saqib Umer, Adnan Khan, Huabin Zhu, and Yachun Wang. 2022. "Joint Transcriptome and Metabolome Analysis Prevails the Biological Mechanisms Underlying the Pro-Survival Fight in In Vitro Heat-Stressed Granulosa Cells" Biology 11, no. 6: 839. https://doi.org/10.3390/biology11060839
APA StyleSammad, A., Luo, H., Hu, L., Zhao, S., Gong, J., Umer, S., Khan, A., Zhu, H., & Wang, Y. (2022). Joint Transcriptome and Metabolome Analysis Prevails the Biological Mechanisms Underlying the Pro-Survival Fight in In Vitro Heat-Stressed Granulosa Cells. Biology, 11(6), 839. https://doi.org/10.3390/biology11060839