
Community Vertical Composition of the Laguna Negra Hypersaline Microbial Mat, Puna Region (Argentinean Andes)


 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
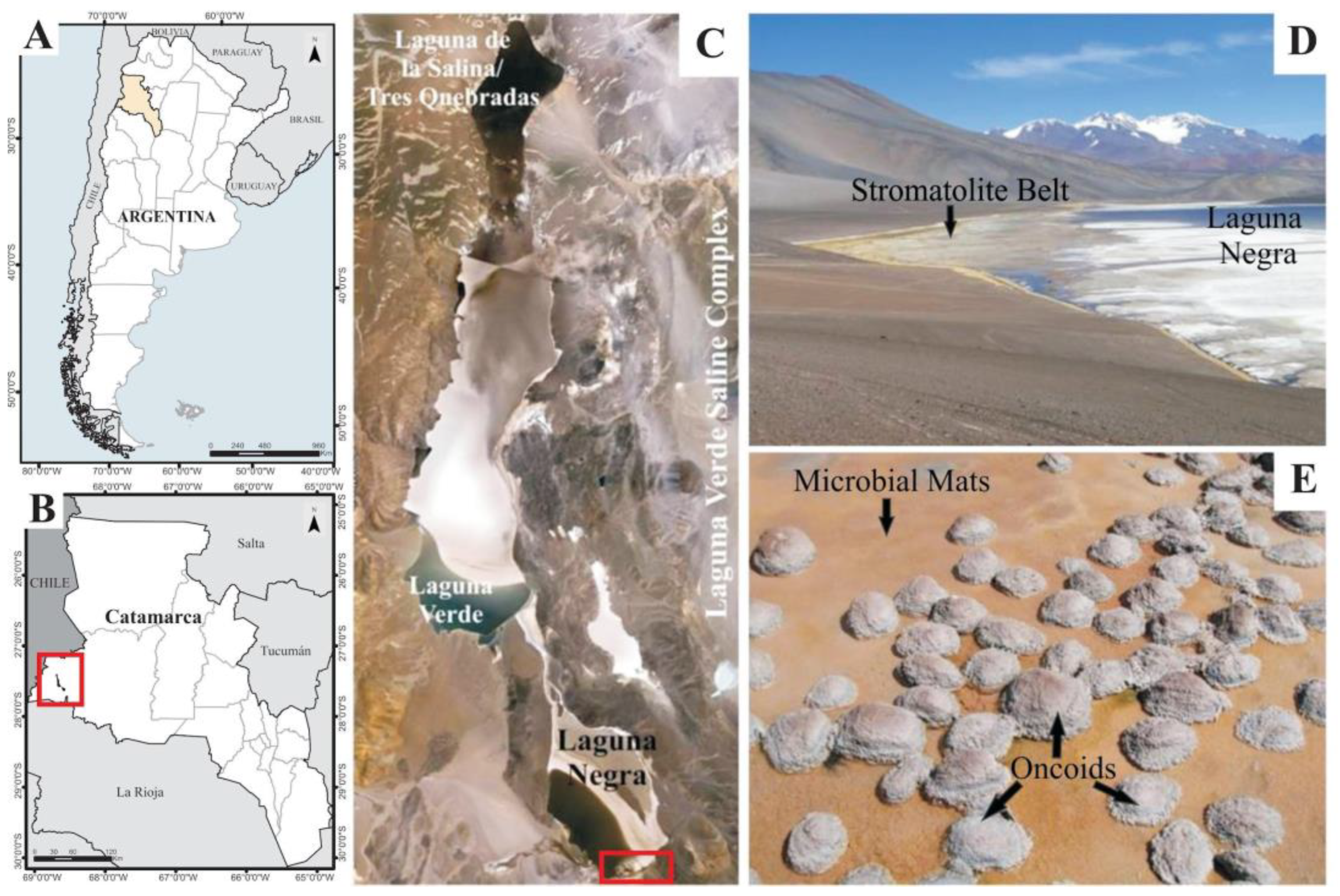
2.1. Study Site
2.2. Sample Collection
2.3. DNA Extraction and 454 Pyrosequencing
2.4. Data Analysis
2.5. Pigment Identification
2.6. Scanning Electron Microscopy and Mineralogy
2.7. Imaging by Confocal Laser Scanning Microscopy
3. Results
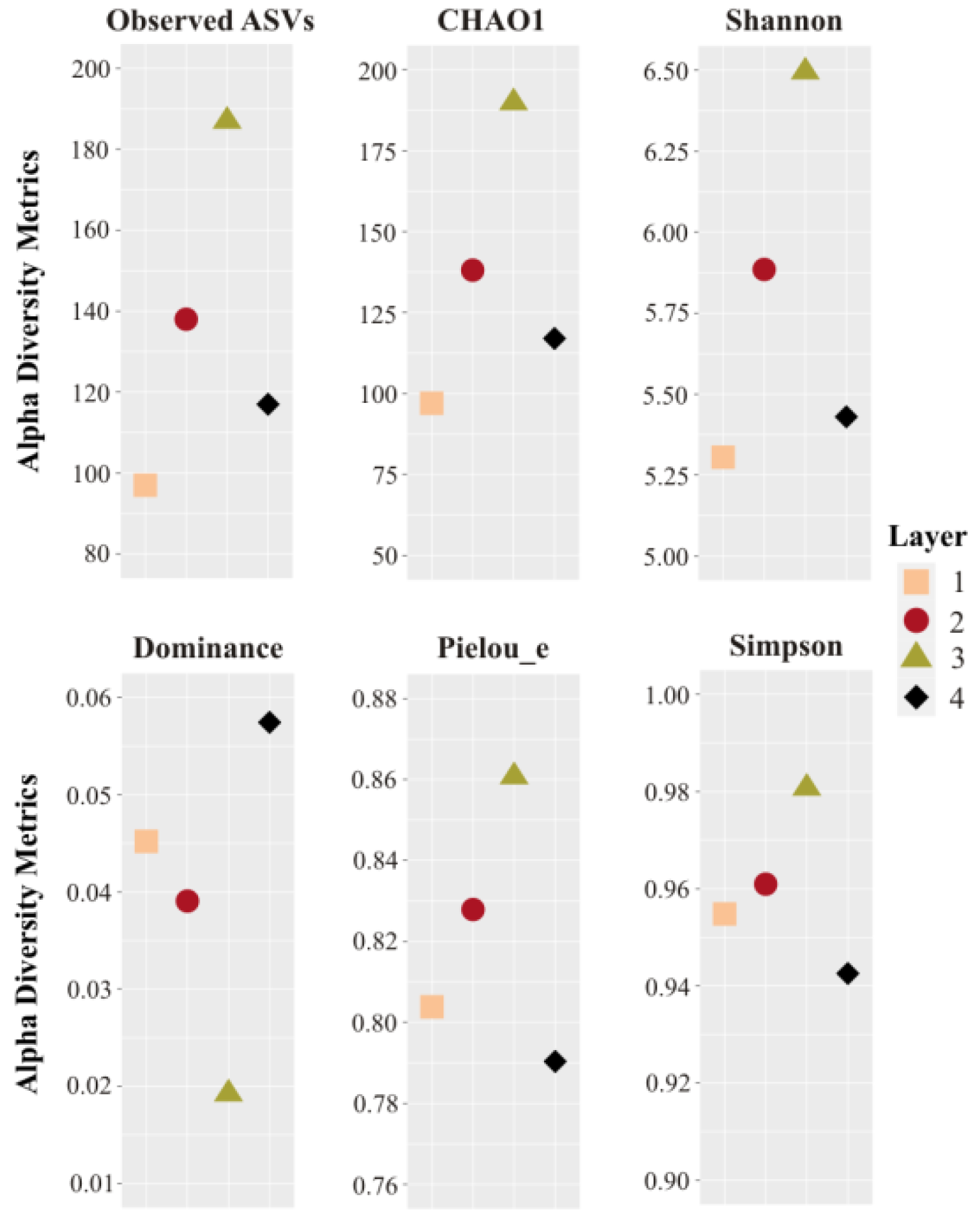
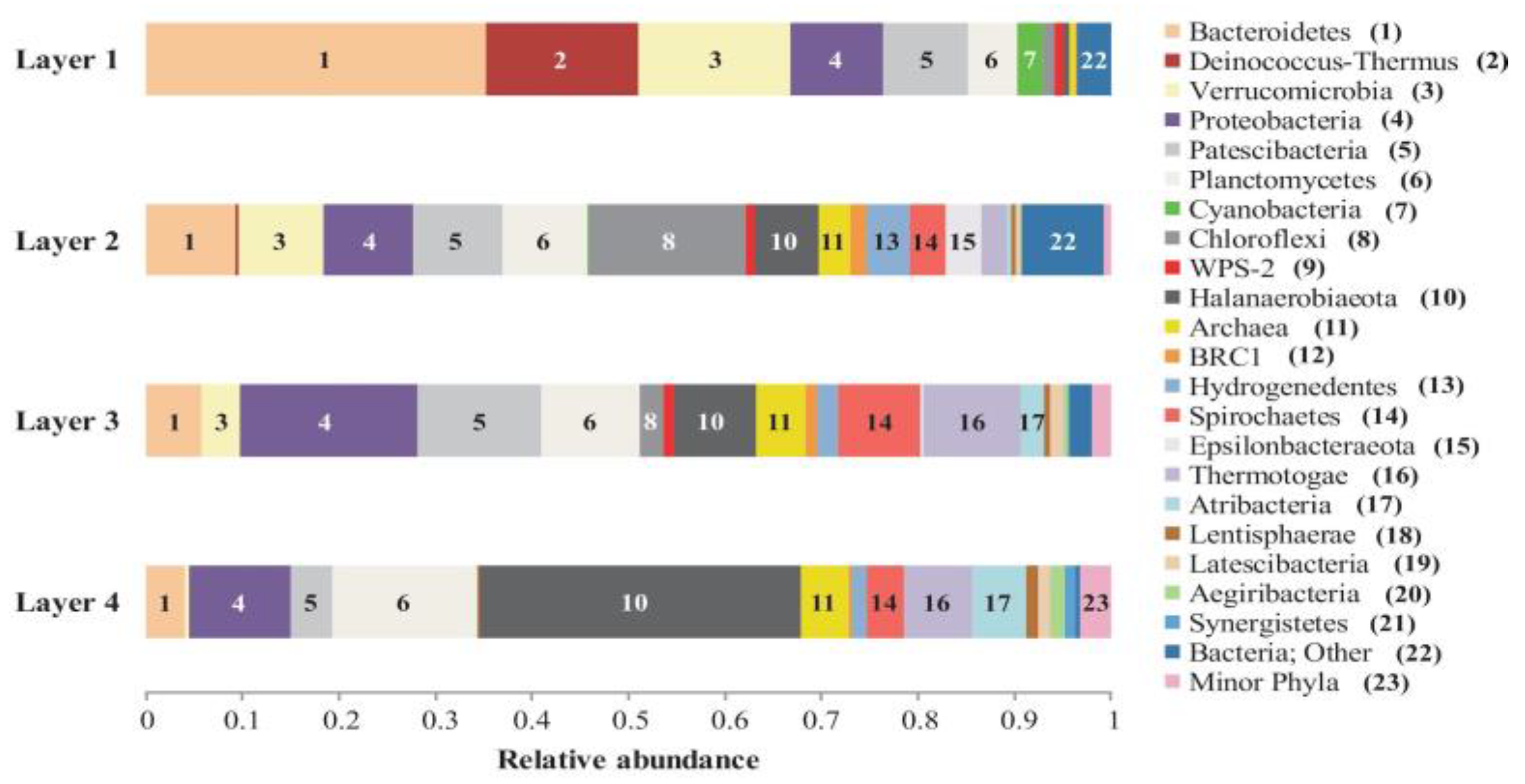
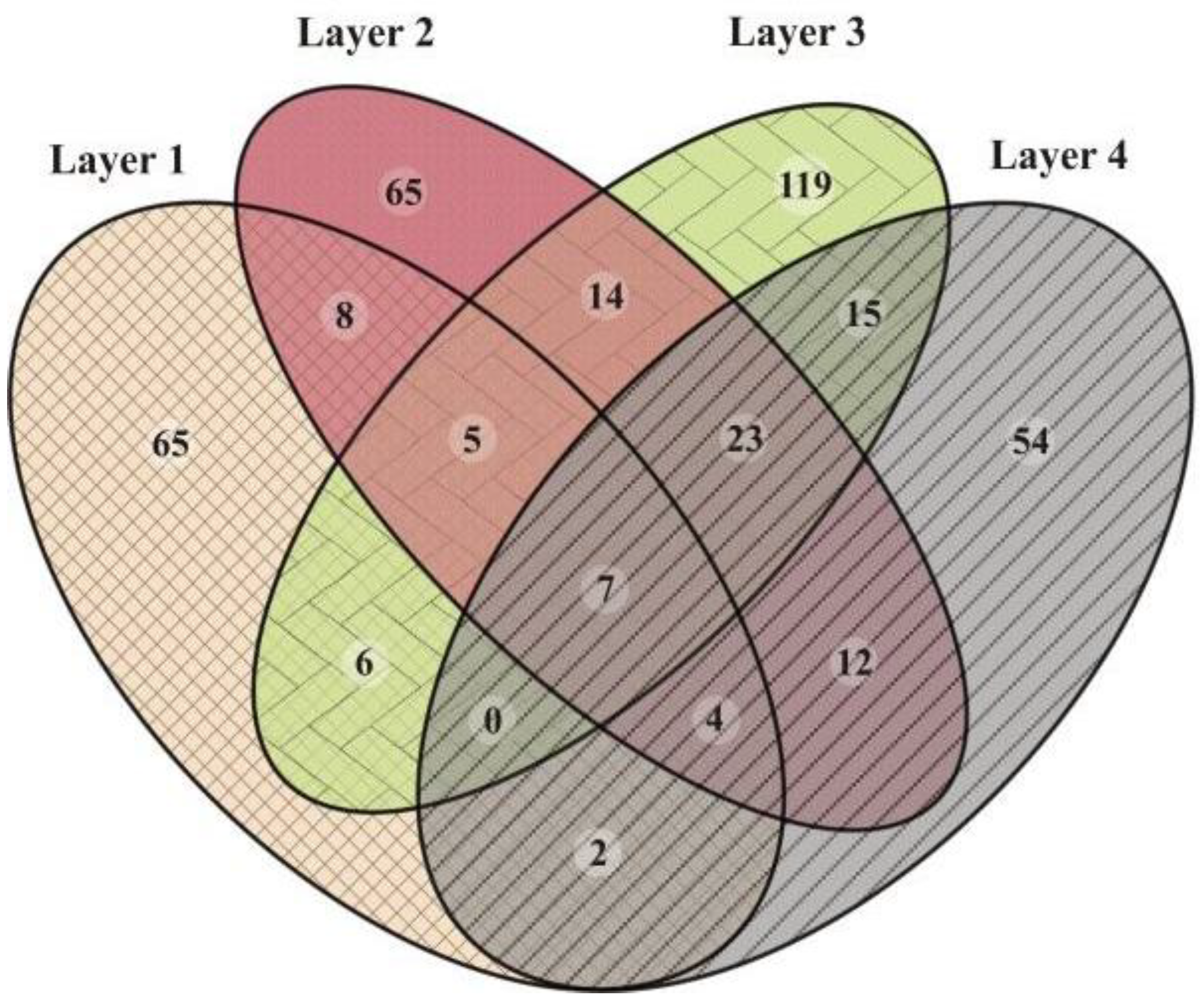
3.1. Prokaryotic Microbial Diversity
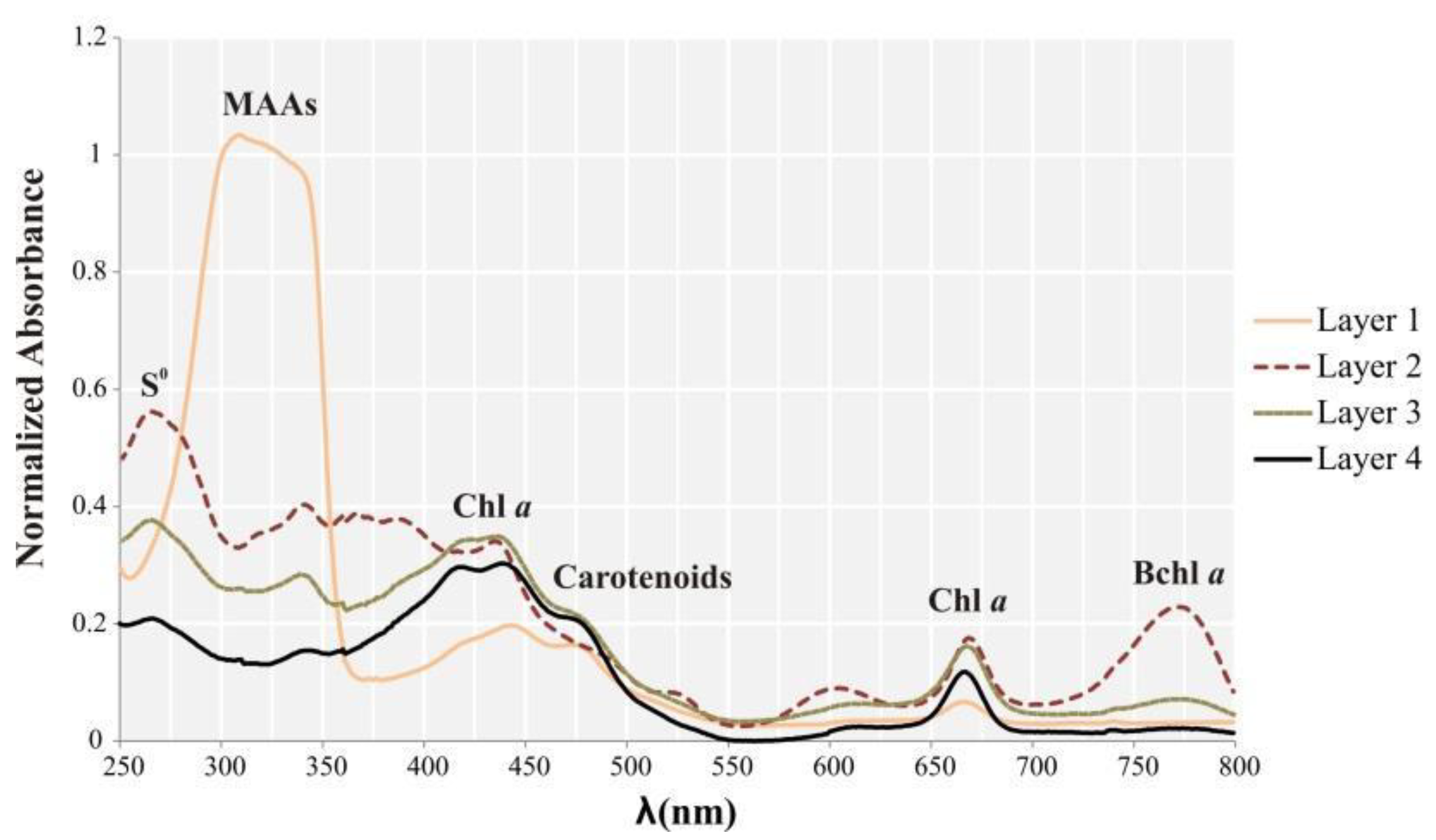
3.2. Pigments Analyses
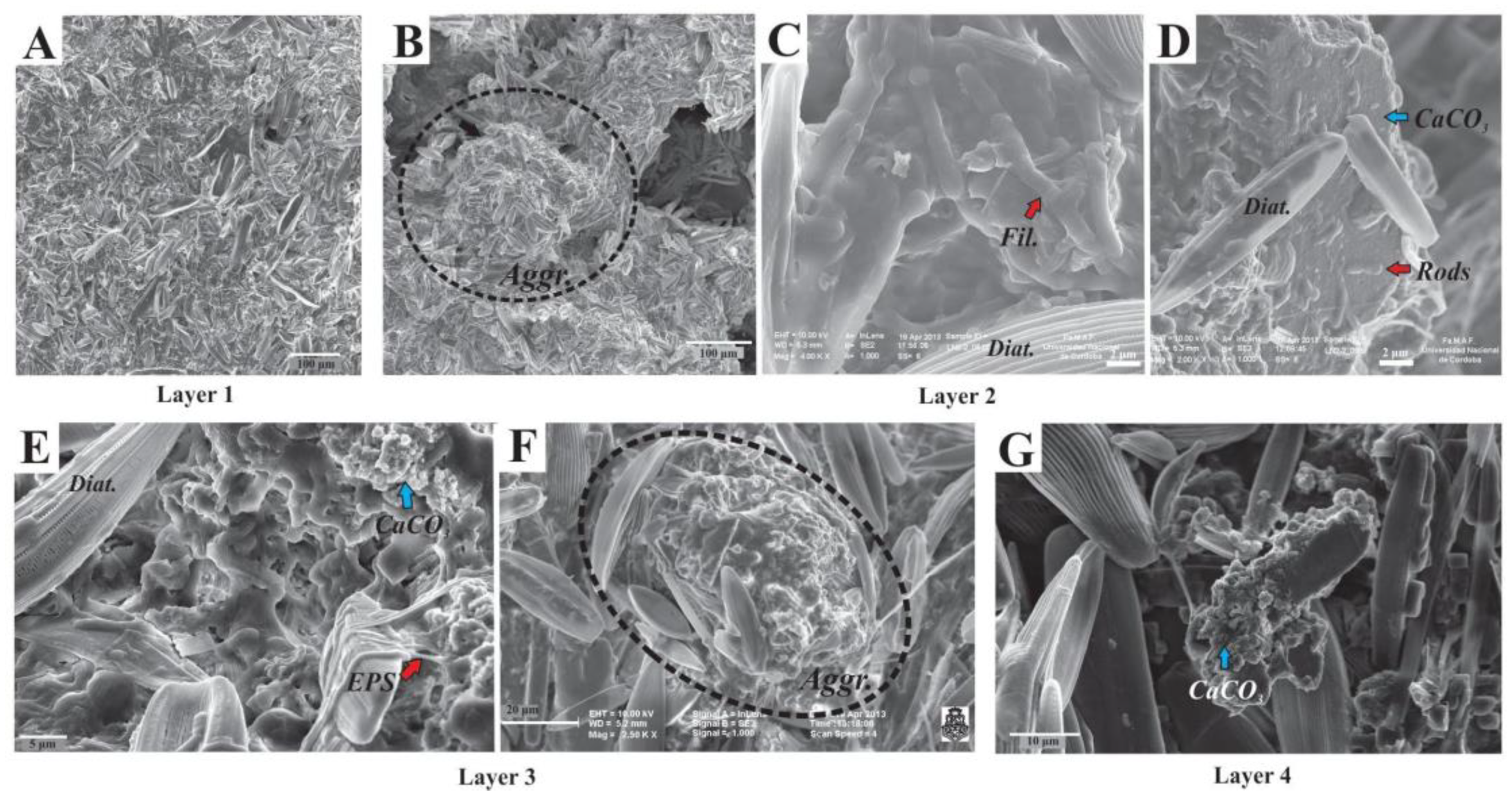
3.3. Scanning Electron Microscopy and XRD Mineralogy
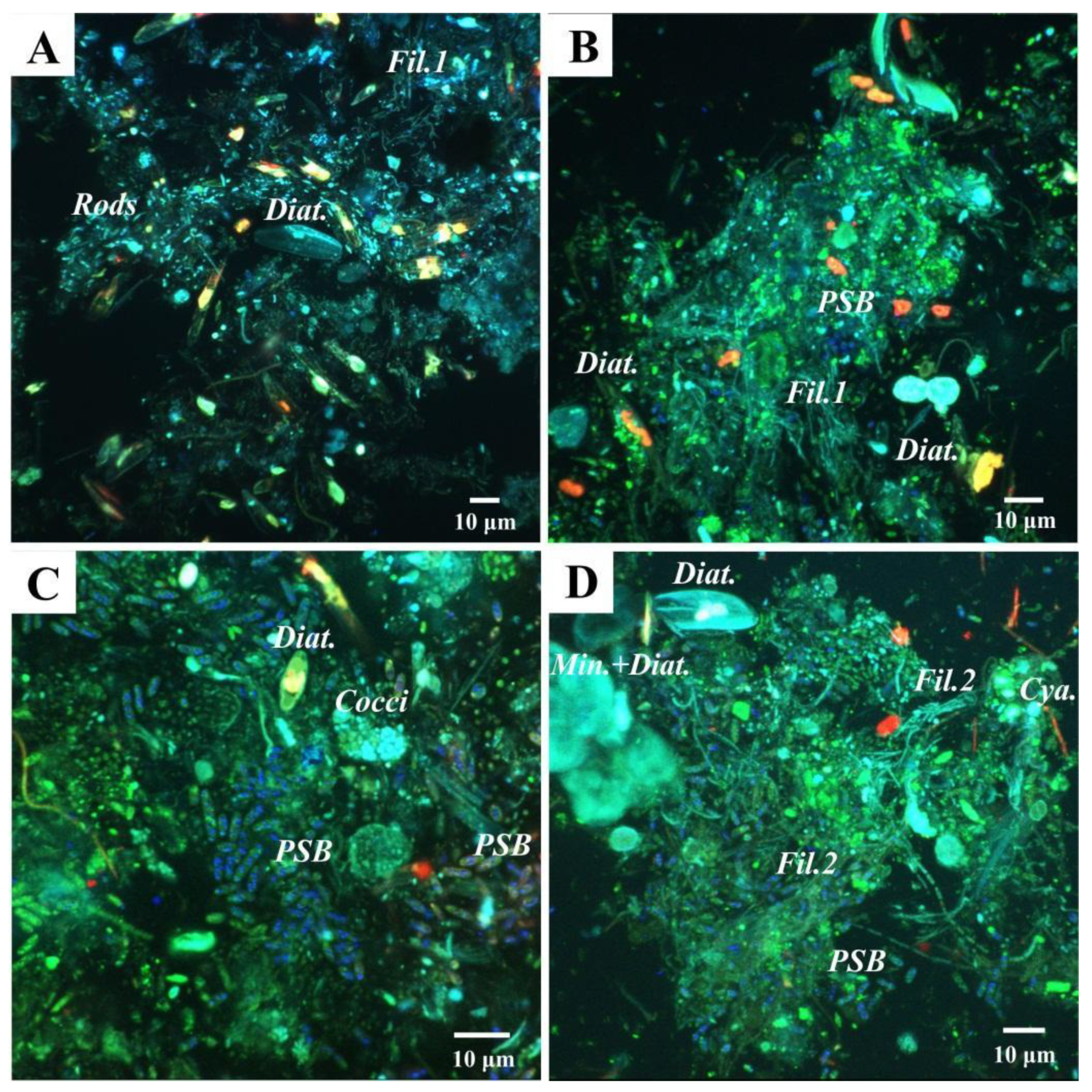
3.4. Confocal Laser Scanning Microscopy Analyses
4. Discussion
4.1. Microbial Diversity of the Laguna Negra Microbial Mat and Other Andean Wetlands
4.2. Microbial Taxa and Pigments Related with High UV Radiation in the Top Layer
4.3. The Laguna Negra Microbial Mat Community Harbors Halophilic Members of Chloroflexi and Halanaerobiaeota
4.4. Aggregates as Main Structural Component with Geomicrobiological Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franks, J.; Stolz, J.F. Flat laminated microbial mat communities. Earth-Sci. Rev. 2009, 96, 163–172. [Google Scholar] [CrossRef]
- Gerdes, G. What Are Microbial Mats? In Microbial Mats: Modern and Ancient Microorganisms in Stratified Systems, 1st ed.; Seckbach, J., Oren, A., Eds.; Springer Science & Business Media B.V.: Dordrecht, The Netherlands, 2010; pp. 5–25. [Google Scholar]
- Prieto-Barajas, C.M.; Valencia-Cantero, E.; Santoyo, G. Microbial mat ecosystems: Structure types, functional diversity, and biotechnological application. Electron. J. Biotechnol. 2018, 31, 48–56. [Google Scholar] [CrossRef]
- Marais, D.J.D. Microbial mats and the early evolution of life. Trends Ecol. Evol. 1990, 5, 140–144. [Google Scholar] [CrossRef]
- Seckbach, J.; Oren, A. Microbial Mats: Modern and Ancient Microorganisms in Stratified Systems, 1st ed.; Springer Science & Business Media B.V.: Dordrecht, The Netherlands, 2010; pp. 389–539. [Google Scholar]
- Ando, N.; Barquera, B.; Bartlett, D.H.; Boyd, E.; Burnim, A.A.; Byer, A.S.; Colman, D.; Gillilan, R.E.; Gruebele, M.; Makhatadze, G.; et al. The Molecular Basis for Life in Extreme Environments. Annu. Rev. Biophys. 2021, 50, 343–372. [Google Scholar] [CrossRef] [PubMed]
- Gregory-Wodzicki, K.M. Uplift history of the central and northern Andes: A review. GSA Bull. 2000, 112, 1091–1105. [Google Scholar] [CrossRef]
- Tapia, J.; Murray, J.; Ormachea-Muñoz, M.; Bhattacharya, P. The Unique Altiplano-Puna Plateau: Environmental Perspectives. J. South Am. Earth Sci. 2022, 115, 103725. [Google Scholar] [CrossRef]
- Piacentini, R.D.; Cede, A.; Bárcena, H. Extreme solar total and UV irradiances due to cloud effect measured near the summer solstice at the high-altitude desertic plateau Puna of Atacama (Argentina). J. Atmos. Sol. -Terr. Phys. 2003, 65, 727–731. [Google Scholar] [CrossRef]
- Strecker, M.R.; Alonso, R.N.; Bookhagen, B.; Carrapa, B.; Hilley, G.E.; Sobel, E.R.; Trauth, M.H. Tectonics and Climate of the Southern Central Andes. Annu. Rev. Earth Planet. Sci. 2007, 35, 747–787. [Google Scholar] [CrossRef] [Green Version]
- Farías, M.E.; Poiré, D.G.; Arrouy, M.J.; Albarracin, V.H. Modern Stromatolite Ecosystems at Alkaline and Hypersaline High-Altitude Lakes in the Argentinean Puna. In Stromatolites: Interaction of Microbes with Sediments; Tewari, V., Seckbach, J., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 431–441. [Google Scholar] [CrossRef]
- Albarracin, V.H.; Kurth, D.; Ordoñez, O.F.; Belfiore, C.; Luccini, E.; Salum, G.; Piacentini, R.D.; Farias, M.E. High-Up: A Remote Reservoir of Microbial Extremophiles in Central Andean Wetlands. Front. Microbiol. 2015, 6, 1404. [Google Scholar] [CrossRef] [Green Version]
- Blumthaler, M.; Ambach, W.; Ellinger, R. Increase in solar UV radiation with altitude. J. Photochem. Photobiol. B Biol. 1997, 39, 130–134. [Google Scholar] [CrossRef]
- Liley, J.B.; Mckenzie, R.L. Where on Earth has the highest UV? Natl. Inst. Water Atmos. Res. 2006, 1, 36–37. [Google Scholar]
- Cordero, R.R.; Seckmeyer, G.; Damiani, A.; Riechelmann, S.; Rayas, J.; Labbe, F.; Laroze, D. The world’s highest levels of surface UV. Photochem. Photobiol. Sci. 2014, 13, 70–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häder, D.P.; Cabrol, N.A. Monitoring of Solar Irradiance in the High Andes. Photochem. Photobiol. 2020, 96, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Cabrol, N.A.; Feister, U.; Häder, D.P.; Piazena, H.; Grin, E.A.; Klein, A. Record solar UV irradiance in the tropical Andes. Front. Environ. Sci. 2014, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Cabrol, N.A.; Grin, E.A.; Chong, G.; Minkley, E.; Hock, A.N.; Yu, Y.; Bebout, L.; Fleming, E.; Häder, D.P.; Demergasso, C.; et al. The High-Lakes Project. J. Geophys. Res. Earth Surf. 2009, 114, G00D06. [Google Scholar] [CrossRef] [Green Version]
- Gomez, F.J.; Kah, L.C.; Bartley, J.; Astini, R.A. Microbialites in a high-altitude andean lake: Multiple controls in carbonate precipitation and lamina accretion. Palaios 2014, 29, 233–249. [Google Scholar] [CrossRef]
- Farías, M.E.; Rascovan, N.; Toneatti, D.M.; Albarracin, V.; Flores, M.R.; Poiré, D.G.; Collavino, M.M.; Aguilar, O.M.; Vazquez, M.P.; Polerecky, L. The Discovery of Stromatolites Developing at 3570 m above Sea Level in a High-Altitude Volcanic Lake Socompa, Argentinean Andes. PLoS ONE 2013, 8, e53497. [Google Scholar] [CrossRef]
- Farías, M.E.; Contreras, M.; Rasuk, M.C.; Kurth, D.; Flores, M.R.; Poiré, D.G.; Novoa, F.; Visscher, P.T. Characterization of bacterial diversity associated with microbial mats, gypsum evaporites and carbonate microbialites in thalassic wetlands: Tebenquiche and La Brava, Salar de Atacama, Chile. Extremophiles 2014, 18, 311–329. [Google Scholar] [CrossRef]
- Farias, M.E.; Rasuk, M.C.; Gallagher, K.; Contreras, M.; Kurth, D.; Fernandez, A.B.; Poiré, D.G.; Novoa, F.; Visscher, P. Prokaryotic diversity and biogeochemical characteristics of benthic microbial ecosystems at La Brava, a hypersaline lake at Salar de Atacama, Chile. PLoS ONE 2017, 12, e0186867. [Google Scholar] [CrossRef] [Green Version]
- Rasuk, M.C.; Fernández, A.B.; Kurth, D.; Contreras, M.; Novoa, F.; Poiré, D.; Farías, M.E. Bacterial Diversity in Microbial Mats and Sediments from the Atacama Desert. Microb. Ecol. 2016, 71, 44–56. [Google Scholar] [CrossRef]
- Fernandez, A.B.; Rasuk, M.C.; Visscher, P.T.; Contreras, M.; Novoa, F.; Poire, D.G.; Patterson, M.M.; Ventosa, A.; Farias, M.E. Microbial Diversity in Sediment Ecosystems (Evaporites Domes, Microbial Mats, and Crusts) of Hypersaline Laguna Tebenquiche, Salar de Atacama, Chile. Front. Microbiol. 2016, 7, 1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toneatti, D.M.; Albarracín, V.H.; Flores, M.R.; Polerecky, L.; Farías, M.E. Stratified Bacterial Diversity along Physico-chemical Gradients in High-Altitude Modern Stromatolites. Front. Microbiol. 2017, 8, 646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villafañe, P.G.; Cónsole-Gonella, C.; Cury, L.F.; Farías, M.E. Short-term microbialite resurgence as indicator of ecological resilience against crises (Catamarca, Argentine Puna). Environ. Microbiol. Rep. 2021, 13, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Rascovan, N.; Maldonado, J.; Vazquez, M.P.; Farías, M.E. Metagenomic study of red biofilms from Diamante Lake reveals ancient arsenic bioenergetics in haloarchaea. ISME J. 2016, 10, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Zenoff, V.; Heredia, J.; Ferrero, M.; Siñeriz, F.; Farías, M.E. Diverse UV-B Resistance of Culturable Bacterial Community from High-Altitude Wetland Water. Curr. Microbiol. 2006, 52, 359–362. [Google Scholar] [CrossRef]
- Flores, M.R.; Ordoñez, O.F.; Maldonado, M.J.; Farías, M.E. Isolation of UV-B resistant bacteria from two high altitude Andean lakes (4,400 m) with saline and non-saline conditions. J. Gen. Appl. Microbiol. 2009, 55, 447–458. [Google Scholar] [CrossRef] [Green Version]
- Albarracín, V.H.; Gärtner, W.; Farías, M.E. Forged Under the Sun: Life and Art of Extremophiles from Andean Lakes. Photochem. Photobiol. 2016, 92, 14–28. [Google Scholar] [CrossRef]
- Albarracín, V.H.; Kraiselburd, I.; Bamann, C.; Wood, P.G.; Bamberg, E.; Farias, M.E.; Gärtner, W. Functional Green-Tuned Proteorhodopsin from Modern Stromatolites. PLoS ONE 2016, 11, e0154962. [Google Scholar] [CrossRef]
- Rasuk, M.C.; Ferrer, G.M.; Kurth, D.; Portero, L.R.; Farías, M.E.; Albarracín, V.H. UV-Resistant Actinobacteria from High-Altitude Andean Lakes: Isolation, Characterization and Antagonistic Activities. Photochem. Photobiol. 2017, 93, 865–880. [Google Scholar] [CrossRef]
- Portero, L.R.; Alonso-Reyes, D.G.; Zannier, F.; Vazquez, M.P.; Farías, M.E.; Gärtner, W.; Albarracín, V.H. Photolyases and Cryptochromes in UV-resistant Bacteria from High-altitude Andean Lakes. Photochem. Photobiol. 2019, 95, 315–330. [Google Scholar] [CrossRef]
- Gorriti, M.F.; Dias, G.M.; Chimetto, L.A.; Trindade-Silva, A.E.; Silva, B.S.; Mesquita, M.M.A.; Gregoracci, G.B.; Farias, M.E.; Thompson, C.C.; Thompson, F.L. Genomic and phenotypic attributes of novel salinivibrios from stromatolites, sediment and water from a high-altitude lake. BMC Genom. 2014, 15, 473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Reyes, D.; Farias, M.; Albarracín, V.H. Uncovering cryptochrome/photolyase gene diversity in aquatic microbiomes exposed to diverse UV B regimes. Aquat. Microbial. Ecol. 2020, 85, 141–154. [Google Scholar] [CrossRef]
- Alonso-Reyes, D.G.; Galván, F.S.; Portero, L.R.; Alvarado, N.N.; Farías, M.E.; Vazquez, M.P.; Albarracín, V.H. Genomic insights into an andean multiresistant soil actinobacterium of biotechnological interest. World J. Microbiol. Biotechnol. 2021, 37, 166. [Google Scholar] [CrossRef] [PubMed]
- Zannier, F.; Portero, L.R.; Douki, T.; Gärtner, W.; Farías, M.E.; Albarracín, V.H. Proteomic Signatures of Microbial Adaptation to the Highest Ultraviolet-Irradiation on Earth: Lessons From a Soil Actinobacterium. Front. Microbiol. 2022, 13, 791714. [Google Scholar] [CrossRef]
- Gutiérrez-Preciado, A.; Vargas-Chávez, C.; Reyes-Prieto, M.; Ordoñez, O.F.; Santos-García, D.; Rosas-Pérez, T.; Valdivia-Anistro, J.; Rebollar, E.A.; Saralegui, A.; Moya, A.; et al. The genomic sequence of Exiguobacterium chiriqhuchastr. N139 reveals a species that thrives in cold waters and extreme environmental conditions. PeerJ 2017, 5, e3162. [Google Scholar] [CrossRef] [Green Version]
- Gomez, F.J.; Mlewski, C.; Boidi, F.J.; Farías, M.E.; Gérard, E. Calcium Carbonate Precipitation in Diatom-rich Microbial Mats: The Laguna Negra Hypersaline Lake, Catamarca, Argentina. J. Sediment. Res. 2018, 88, 727–742. [Google Scholar] [CrossRef]
- Boidi, F.J.; Mlewski, E.C.; Gomez, F.J.; Gérard, E. Characterization of Microbialites and Microbial Mats of the Laguna Negra Hypersaline Lake (Puna of Catamarca, Argentina). In Microbial Ecosystems in Central Andes Extreme Environments; Farías, M.E., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 183–203. [Google Scholar]
- Harris, J.K.; Caporaso, J.G.; Walker, J.J.; Spear, J.R.; Gold, N.J.; Robertson, C.E.; Hugenholtz, P.; Goodrich, J.; McDonald, D.; Knights, D.; et al. Phylogenetic stratigraphy in the Guerrero Negro hypersaline microbial mat. ISME J. 2013, 7, 50–60. [Google Scholar] [CrossRef]
- Armitage, D.W.; Gallagher, K.L.; Youngblut, N.D.; Buckley, D.H.; Zinder, S.H. Millimeter-scale patterns of phylogenetic and trait diversity in a salt marsh microbial mat. Front. Microbiol. 2012, 3, 293. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. Accurate taxonomy assignments from 16S rRNA sequences produced by highly parallel pyrosequencers. Nucleic Acids Res. 2008, 36, e120. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.; McMurdie, P.; Rosen, M.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing 2020. Available online: https://www.R-project.org/ (accessed on 10 October 2021).
- Holm-Hansen, O.; Riemann, B. Chlorophyll a Determination: Improvements in Methodology. Oikos 1978, 30, 438–447. [Google Scholar] [CrossRef]
- Borrego, C.; Garcia-Gil, L. Separation of bacteriochlorophyll homologues from green photosynthetic sulfur bacteria by reversed-phase HPLC. Photosynth. Res. 1994, 41, 157–164. [Google Scholar] [CrossRef]
- LipidBank. Available online: http://www.lipidbank.jp (accessed on 20 November 2019).
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Stal, L.J.; van Gemerden, H.; Krumbein, W.E. The simultaneous assay of chlorophyll and bacteriochlorophyll in natural microbial communities. J. Microbiol. Methods 1984, 2, 295–306. [Google Scholar] [CrossRef]
- Pierson, B.K.; Sands, V.M.; Frederick, J.L. Spectral irradiance and distribution of pigments in a highly layered marine microbial mat. Appl. Environ. Microbiol. 1990, 56, 2327–2340. [Google Scholar] [CrossRef] [Green Version]
- Asker, D.; Beppu, T.; Ueda, K. Unique diversity of carotenoid-producing bacteria isolated from Misasa, a radioactive site in Japan. Appl. Microbiol. Biotechnol. 2007, 77, 383–392. [Google Scholar] [CrossRef]
- Lemee, L.; Peuchant, E.; Clerc, M.; Brunner, M.; Pfander, H. Deinoxanthin: A new carotenoid isolated from Deinococcus radiodurans. Tetrahedron 1997, 53, 919–926. [Google Scholar] [CrossRef]
- Mareš, J.; Hrouzek, P.; Kana, R.; Ventura, S.; Strunecky, O.; Komarek, J. The Primitive Thylakoid-Less Cyanobacterium Gloeobacter Is a Common Rock-Dwelling Organism. PLoS ONE 2013, 8, e66323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuczynska, P.; Jemiola-Rzeminska, M.; Strzalka, K. Photosynthetic Pigments in Diatoms. Mar. Drugs 2015, 13, 5847–5881. [Google Scholar] [CrossRef] [PubMed]
- George, D.M.; Vincent, A.S.; Mackey, H.R. An overview of anoxygenic phototrophic bacteria and their applications in environmental biotechnology for sustainable Resource recovery. Biotechnol. Rep. 2020, 28, e00563. [Google Scholar] [CrossRef]
- Jeong, S.-W.; Kim, J.-H.; Kim, J.-W.; Kim, C.Y.; Kim, S.Y.; Choi, Y.J. Metabolic Engineering of Extremophilic Bacterium Deinococcus radiodurans for the Production of the Novel Carotenoid Deinoxanthin. Microorganisms 2021, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids, 1st ed; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 2004. [Google Scholar]
- Lohr, M.; Wilhelm, C. Xanthophyll synthesis in diatoms: Quantification of putative intermediates and comparison of pigment conversion kinetics with rate constants derived from a model. Planta 2001, 212, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Esatbeyoglu, T.; Rimbach, G. Canthaxanthin: From molecule to function. Mol. Nutr. Food Res. 2016, 61, 1600469. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, R.E.; Olson, J.M.; Miller, M. Antenna Complexes from Green Photosynthetic Bacteria. In Anoxygenic Photosynthetic Bacteria. Advances in Photosynthesis and Respiration; Blankenship, R.E., Madigan, M.T., Bauer, C.E., Eds.; Springer: Dordrecht, The Netherlands, 1995; Volume 2, pp. 399–435. [Google Scholar]
- Parusel, A.B.J.; Grimme, S. A Theoretical Study of the Excited States of Chlorophyll a and Pheophytin a. J. Phys. Chem. B 2000, 104, 5395–5398. [Google Scholar] [CrossRef]
- Maturrano, L.; Santos, F.; Rosselló-Mora, R.; Antón, J. Microbial Diversity in Maras Salterns, a Hypersaline Environment in the Peruvian Andes. Appl. Environ. Microbiol. 2006, 72, 3887–3895. [Google Scholar] [CrossRef] [Green Version]
- Borsodi, A.K.; Aszalós, J.M.; Megyes, M.; Nagy, B. Benthic Bacterial Diversity of High-Altitude Athalassohaline Lakes of the Puna de Atacama (Central Andes). Geomicrobiol. J. 2022, 39, 28–38. [Google Scholar] [CrossRef]
- Aguilar, P.; Acosta, E.; Dorador, C.; Sommaruga, R. Large Differences in Bacterial Community Composition among Three Nearby Extreme Waterbodies of the High Andean Plateau. Front. Microbiol. 2016, 7, 976. [Google Scholar] [CrossRef]
- Rasuk, M.C.; Kurth, D.; Flores, M.R.; Contreras, M.; Novoa, F.; Poire, D.; Farías, M.E. Microbial Characterization of Microbial Ecosystems Associated to Evaporites Domes of Gypsum in Salar de Llamara in Atacama Desert. Microb. Ecol. 2014, 68, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Stivaletta, N.; Barbieri, R.; Cevenini, F.; López-García, P. Physicochemical Conditions and Microbial Diversity Associated with the Evaporite Deposits in the Laguna de la Piedra (Salar de Atacama, Chile). Geomicrobiol. J. 2011, 28, 83–95. [Google Scholar] [CrossRef]
- Mlewski, E.C.; Pisapia, C.; Gomez, F.; Lecourt, L.; Rueda, E.S.; Benzerara, K.; Ménez, B.; Borensztajn, S.; Jamme, F.; Réfrégiers, M.; et al. Characterization of Pustular Mats and Related Rivularia-Rich Laminations in Oncoids From the Laguna Negra Lake (Argentina). Front. Microbiol. 2018, 9, 996. [Google Scholar] [CrossRef] [PubMed]
- Shalygin, S.; Pietrasiak, N.; Gomez, F.; Mlewski, C.; Gerard, E.; Johansen, J.R. Rivularia halophila sp. nov. (Nostocales, Cyanobacteria): The first species of Rivularia described with the modern polyphasic approach. Eur. J. Phycol. 2018, 53, 537–548. [Google Scholar] [CrossRef]
- Sylvestre, F.; Servant-Vildary, S.; Roux, M. Diatom-based ionic concentration and salinity models from the south Bolivian Altiplano (15–23° S). J. Paleolimnol. 2001, 25, 279–295. [Google Scholar] [CrossRef]
- Tapia, P.M.; Fritz, S.C.; Seltzer, G.O.; Rodbell, D.T.; Metivier, S.P. Contemporary distribution and late-quaternary stratigraphy of diatoms in the Junin plain, central Andes, Peru. Bol. Soc. Geol. Perú 2006, 101, 19–42. [Google Scholar]
- Arrage, A.A.; Phelps, T.J.; Benoit, R.E.; White, D.C. Survival of subsurface microorganisms exposed to UV radiation and hydrogen peroxide. Appl. Environ. Microbiol. 1993, 59, 3545–3550. [Google Scholar] [CrossRef] [Green Version]
- Davis, S.J.; Vener, A.V.; Vierstra, R.D. Bacteriophytochromes: Phytochrome-like photoreceptors from nonphotosynthetic Eubacteria. Science 1999, 286, 2517–2520. [Google Scholar] [CrossRef]
- Makarova, K.S.; Aravind, L.; Wolf, Y.I.; Tatusov, R.L.; Minton, K.W.; Koonin, E.V.; Daly, M.J. Genome of the Extremely Radiation-Resistant Bacterium Deinococcus radiodurans Viewed from the Perspective of Comparative Genomics. Microbiol. Mol. Biol. Rev. 2001, 65, 44–79. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, L.; Simãµes, C.; Nobre, M.F.; Pino, N.M.; Battista, J.R.; Silva, M.T.; Rainey, F.A.; de Costa, M.S. Truepera radiovictrix gen. nov., sp. nov., a new radiation resistant species and the proposal of Trueperaceae fam. nov. FEMS Microbiol. Lett. 2005, 247, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, N.; Rohde, C.; Munk, C.; Nolan, M.; Lucas, S.; Del Rio, T.G.; Tice, H.; Deshpande, S.; Cheng, J.-F.; Tapia, R.; et al. Complete genome sequence of Truepera radiovictrix type strain (RQ-24T). Stand. Genom. Sci. 2011, 4, 91–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berelson, W.M.; Corsetti, F.A.; Pepe-Ranney, C.; Hammond, D.E.; Beaumont, W.; Spear, J.R. Hot spring siliceous stromatolites from Yellowstone National Park: Assessing growth rate and laminae formation. Geobiology 2011, 9, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Harris, J.K.; Wilcox, J.; Spear, J.R.; Miller, S.R.; Bebout, B.M.; Maresca, J.A.; Bryant, D.A.; Sogin, M.L.; Pace, N.R. Unexpected Diversity and Complexity of the Guerrero Negro Hypersaline Microbial Mat. Appl. Environ. Microbiol. 2006, 72, 3685–3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, H.L.; Smith, D.L.; Visscher, P.T.; Burns, B.P. Niche differentiation of bacterial communities at a millimetre scale in Shark Bay microbial mats. Sci. Rep. 2015, 5, 15607. [Google Scholar] [CrossRef]
- Mobberley, J.M.; Ortega, M.C.; Foster, J.S. Comparative microbial diversity analyses of modern marine thrombolitic mats by barcoded pyrosequencing. Environ. Microbiol. 2012, 14, 82–100. [Google Scholar] [CrossRef]
- Korbee, N.; Figueroa, F.; Aguilera, J. Accumulation of mycosporine-like amino acids (MAAs): Biosynthesis, photocontrol and ecophysiological functions. Rev. Chil. Hist. Nat. 2006, 79, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Pichel, F.; Castenholz, R.W. Occurrence of UV-absorbing, mycosporine-like compounds among cyanobacterial isolates and an estimate of their screening capacity. Appl. Environ. Microbiol. 1993, 59, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Shick, J.M.; Dunlap, W.C. Mycosporine-Like Amino Acids and Related Gadusols: Biosynthesis, Accumulation, and UV-Protective Functions in Aquatic Organisms. Annu. Rev. Physiol. 2002, 64, 223–262. [Google Scholar] [CrossRef] [Green Version]
- Oren, A.; Gunde-Cimerman, N. Mycosporines and mycosporine-like amino acids: UV protectants or multipurpose secondary metabolites? FEMS Microbiol. Lett. 2007, 269, 1–10. [Google Scholar] [CrossRef]
- Ingalls, A.E.; Whitehead, K.; Bridoux, M.C. Tinted windows: The presence of the UV absorbing compounds called mycosporine-like amino acids embedded in the frustules of marine diatoms. Geochim. Cosmochim. Acta 2010, 74, 104–115. [Google Scholar] [CrossRef]
- Klappenbach, J.A.; Pierson, B.K. Phylogenetic and physiological characterization of a filamentous anoxygenic photoautotrophic bacterium “Candidatus Chlorothrix halophila” gen. nov., sp. nov., recovered from hypersaline microbial mats. Arch. Microbiol. 2004, 181, 17–25. [Google Scholar] [CrossRef] [PubMed]
- van de Meene, A.M.L.; Le Olson, T.; Collins, A.M.; Blankenship, R.E. Initial Characterization of the Photosynthetic Apparatus of “Candidatus Chlorothrix halophila,” a Filamentous, Anoxygenic Photoautotroph. J. Bacteriol. 2007, 189, 4196–4203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anil Kumar, P.; Srinivas, T.N.R.; Sasikala, C.; Ramana, C.V. Halochromatium roseum sp. nov., a non-motile phototrophic gammaproteobacterium with gas vesicles, and emended description of the genus Halochromatium. Int. J. Syst. Evol. Microbiol. 2007, 57, 2110–2113. [Google Scholar] [CrossRef] [PubMed]
- Maki, J.S. Bacterial Intracellular Sulfur Globules: Structure and Function. J. Mol. Microbiol. Biotechnol. 2013, 23, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F. The Family Chromatiaceae. In The Prokaryotes, 4th ed; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 151–178. [Google Scholar]
- Mitchell, H.M.; Rocha, G.A.; Kaakoush, N.O.; O’Rourke, J.L.; Queiroz, D.M.M. The Family Helicobacteraceae. In The Prokaryotes, 4th ed; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 337–392. [Google Scholar]
- Nakai, R.; Nishijima, M.; Tazato, N.; Handa, Y.; Karray, F.; Sayadi, S.; Isoda, H.; Naganuma, T. Oligoflexus tunisiensis gen. nov., sp. nov., a Gram-negative, aerobic, filamentous bacterium of a novel proteobacterial lineage, and description of Oligoflexaceae fam. nov., Oligoflexales ord. nov. and Oligoflexia classis nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 3353–3359. [Google Scholar] [CrossRef]
- Waite, D.W.; Chuvochina, M.; Pelikan, C.; Parks, D.H.; Yilmaz, P.; Wagner, M.; Loy, A.; Naganuma, T.; Nakai, R.; Whitman, W.B.; et al. Proposal to reclassify the proteobacterial classes Deltaproteobacteria and Oligoflexia, and the phylum Thermodesulfobacteria into four phyla reflecting major functional capabilities. Int. J. Syst. Evol. Microbiol. 2020, 70, 5972–6016. [Google Scholar] [CrossRef]
- Ivanova, N.; Sikorski, J.; Chertkov, O.; Nolan, M.; Lucas, S.; Hammon, N.; Deshpande, S.; Cheng, J.-F.; Tapia, R.; Han, C.; et al. Complete genome sequence of the extremely halophilic Halanaerobium praevalens type strain (GSLT). Stand. Genom. Sci. 2011, 4, 312–321. [Google Scholar] [CrossRef] [Green Version]
- Kivistö, A.; Santala, V.; Karp, M. 1,3-Propanediol production and tolerance of a halophilic fermentative bacterium, Halanaerobium saccharolyticum subsp. saccharolyticum. J. Biotechnol. 2012, 158, 242–247. [Google Scholar] [CrossRef]
- Abdeljabbar, H.; Cayol, J.L.; Hania, W.B.; Boudabous, A.; Sadfi, N.; Fardeau, M.L. Halanaerobium sehlinense sp. nov., an extremely halophilic, fermentative, strictly anaerobic bacterium from sediments of the hypersaline lake Sehline Sebkha. Int. J. Syst. Evol. Microbiol. 2013, 63, 2069–2074. [Google Scholar] [CrossRef]
- Ravot, G.; Casalot, L.; Ollivier, B.; Loison, G.; Magot, M. rdlA, a new gene encoding a rhodanese-like protein in Halanaerobium congolense and other thiosulfate-reducing anaerobes. Res. Microbiol. 2005, 156, 1031–1038. [Google Scholar] [CrossRef]
- Liang, R.; Davidova, I.A.; Marks, C.R.; Stamps, B.W.; Harriman, B.H.; Stevenson, B.S.; Duncan, K.E.; Suflita, J.M. Metabolic Capability of a Predominant Halanaerobium sp. in Hydraulically Fractured Gas Wells and Its Implication in Pipeline Corrosion. Front. Microbiol. 2016, 7, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booker, A.E.; Borton, M.A.; Daly, R.A.; Welch, S.A.; Nicora, C.D.; Hoyt, D.W.; Wilson, T.; Purvine, S.O.; Wolfe, R.A.; Sharma, S.; et al. Sulfide generation by dominant Halanaerobium microorganisms in hydraulically fractured shales. mSphere 2017, 2, e00257-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupraz, C.; Reid, R.P.; Braissant, O.; Decho, A.W.; Norman, R.S.; Visscher, P.T. Processes of carbonate precipitation in modern microbial mats. Earth-Sci. Rev. 2009, 96, 141–162. [Google Scholar] [CrossRef]
- Bolhuis, H.; Cretoiu, M.S.; Stal, L.J. Molecular ecology of microbial mats. FEMS Microbiol. Ecol. 2014, 90, 335–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Wit, R.; Van Den Ende, F.P.; Van Gemerden, H. Mathematical simulation of the interactions among cyanobacteria, purple sulfur bacteria and chemotropic sulfur bacteria in microbial mat communities. FEMS Microbiol. Ecol. 1995, 17, 117–135. [Google Scholar] [CrossRef]
- Decker, K.L.M.; Potter, C.S.; Bebout, B.M.; Des Marais, D.J.; Carpenter, S.; Discipulo, M.; Hoehler, T.M.; Miller, S.R.; Thamdrup, B.; Turk, K.A.; et al. Mathematical simulation of the diel O, S, and C biogeochemistry of a hypersaline microbial mat. FEMS Microbiol. Ecol. 2005, 52, 377–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, E.K.; Kump, L.R. Biogeochemistry of microbial mats under Precambrian environmental conditions: A modelling study. Geobiology 2005, 3, 77–92. [Google Scholar] [CrossRef]
- Jørgensen, B.B. Bacterial sulfate reduction within reduced microniches of oxidized marine sediments. Mar. Biol. 1977, 41, 7. [Google Scholar] [CrossRef]
- Sigalevich, P.; Meshorer, E.; Helman, Y.; Cohen, Y. Transition for anaerobic to aerobic growth conditions for the sulfate-reducing bacterium Desulfovibrio oxyclinae results in flocculation. Appl. Environ. Microbiol. 2000, 66, 5005–5012. [Google Scholar] [CrossRef] [Green Version]
- Fourçans, A.; Ranchou-Peyruse, A.; Caumette, P.; Duran, R. Molecular Analysis of the Spatio-temporal Distribution of Sulfate-reducing Bacteria (SRB) in Camargue (France) Hypersaline Microbial Mat. Microb. Ecol. 2008, 56, 90–100. [Google Scholar] [CrossRef]
- Wilbanks, E.G.; Jaekel, U.; Salman, V.; Humphrey, P.T.; Eisen, J.A.; Facciotti, M.T.; Buckley, D.H.; Zinder, S.H.; Druschel, G.K.; Fike, D.; et al. Microscale sulfur cycling in the phototrophic pink berry consortia of the S ippewissett S alt M arsh. Environ. Microbiol. 2014, 16, 3398–3415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escudero, L.V.; Casamayor, E.O.; Chong, G.; Pedrós-Alió, C.; Demergasso, C. Distribution of Microbial Arsenic Reduction, Oxidation and Extrusion Genes along a Wide Range of Environmental Arsenic Concentrations. PLoS ONE 2013, 8, e78890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, K.L.; Mikulski, C.M.; Barnhorst, A.; Doucette, G.J. Comparative analysis of bacterioplankton assemblages from Karenia brevis bloom and nonbloom water on the west Florida shelf (Gulf of Mexico, USA) using 16S rRNA gene clone libraries. FEMS Microbiol. Ecol. 2010, 73, 468–485. [Google Scholar] [CrossRef] [PubMed]
- Fasca, H.; De Castilho, L.V.A.; De Castilho, J.F.M.; Pasqualino, I.; Alvarez, V.M.; Jurelevicius, D.D.A.; Seldin, L. Response of marine bacteria to oil contamination and to high pressure and low temperature deep sea conditions. MicrobiologyOpen 2018, 7, e00550. [Google Scholar] [CrossRef] [PubMed]
- Glavaš, N.; Défarge, C.; Gautret, P.; Joulian, C.; Penhoud, P.; Motelica, M.; Kovač, N. The structure and role of the “petola” microbial mat in sea salt production of the Sečovlje (Slovenia). Sci. Total Environ. 2018, 644, 1254–1267. [Google Scholar] [CrossRef]
- Ghotra, S.K. Novel Bacterial Lineages in the Uncultured Candidate Division SR1. Master’s Thesis, San José State University, San José, CA, USA, August 2014. [Google Scholar] [CrossRef]
- Dojka, M.A.; Hugenholtz, P.; Haack, S.K.; Pace, N.R. Microbial diversity in a hydrocarbon- and chlorinated-solvent-contaminated aquifer undergoing intrinsic bioremediation. Appl. Environ. Microbiol. 1998, 64, 3869–3877. [Google Scholar] [CrossRef] [Green Version]
- Emmerich, M.; Bhansali, A.; Lösekann-Behrens, T.; Schröder, C.; Kappler, A.; Behrens, S. Abundance, distribution, and activity of Fe(II)-oxidizing and Fe(III)-reducing microorganisms in hypersaline sediments of Lake Kasin, southern Russia. Appl. Environ. Microbiol. 2012, 78, 4386–4399. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Musarrat, J.; Alkhedhairy, A.A.; Kazuo, S. Diversity of bacteria and polyketide synthase associated with marine sponge Haliclona sp. Ann. Microbiol. 2014, 64, 199–207. [Google Scholar] [CrossRef]
- Vogt, J.C.; Abed, R.M.M.; Albach, D.C.; Palinska, K.A. Bacterial and Archaeal Diversity in Hypersaline Cyanobacterial Mats Along a Transect in the Intertidal Flats of the Sultanate of Oman. Microb. Ecol. 2018, 75, 331–347. [Google Scholar] [CrossRef]
- Dorador, C.; Vila, I.; Witzel, K.-P.; Imhoff, J.F. Bacterial and archaeal diversity in high altitude wetlands of the Chilean Altiplano. Fundam. Appl. Limnol. 2013, 182, 135–159. [Google Scholar] [CrossRef]
- Tomitani, A.; (Japan Agency for Marine-Earth Science and Technology, Institute of Biogeosciences, Yokosuka, Japan); Stal, L.J.; (Institute for Biodiversity and Ecosystem Dynamics, Faculty of Science, University of Amsterdam, Amsterdam, The Netherlands). Photosynthetic activity and community structure in coastal microbial mats. Unpublished work. 2013. [Google Scholar]
- Câmara dos Reis, M.; Lacativa Bagatini, I.; de Oliveira Vidal, L.; Bonnet, M.-P.; da Motta Marques, D.; Sarmento, H. Spatial heterogeneity and hydrological fluctuations drive bacterioplankton community composition in an Amazon floodplain system. PLoS ONE 2020, 15, e0228721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corsellis, Y.; Militon, C.; Sylvi, L.; Cuny, P.; (Mediterranean Institute of Oceanography, Marseille, France). High-throughput screening of the seasonal dynamics of prokaryotic communities of a Mediterranean thalassohaline lake. Unpublished work. 2017. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time (min) | Pigment | Layer 1 | Layer 2 | Layer 3 | Layer 4 | Related Organisms According to Bibliography |
|---|---|---|---|---|---|---|
| 3.08 | Unknown | - | - | - | X | |
| 5.067 | Fucoxanthin | XX | - | XXX | XXX | Diatoms, brown algae [58] |
| 6.883 | Diadinoxanthin | XX | X | - | - | Diatoms, phaeophytes, dinophytes, haptophytes [58] |
| 7.547 | Bacteriochlorophyll d | - | XXX | XX | - | Chloroflexi, Chlorobi [59] |
| 7.553 | Deinoxanthin | XX | - | - | - | Deinococcus spp. [60] |
| 7.85 | Carotenoid | X | - | - | - | Algae, diatoms, bacteria, plants, fungi [61] |
| 8.187 | Diatoxanthin | XXX | X | XXX | XXX | Diatoms, phaeophytes, dinophytes, haptophytes [58] |
| 8.417 | Zeaxanthin | X | - | XX | - | Algae, diatoms, bacteria [58], plants [62] |
| 9.151 | Bacteriochlorophyll c | - | XXX | XXX | X | Chloroflexi, Chlorobi, Acidobacteria [59] |
| 9.2 | Cantaxanthin | X | - | - | - | Bacteria (Acidobacteria) [59], algae, fungi, plants [63] |
| 9.504 | Bacteriochlorophyll d derivative | - | XX | X | - | |
| 9.784 | Bacteriochlorophyll a | - | XXXX | X | X | Alpha, Beta, and Gamma-proteobacteria, Chloroflexi, Gemmatimonadota [59], Chlorobi [64] |
| 10.283 | Bacteriochlorophyll a derivative | - | - | XX | XXX | |
| 10.787 | (2S,29S)-oscillol 2,29-difucoside (match 96%) | - | XX | - | - | Cyanobacteria [57] |
| 11.633 | Chlorophyll a | XXX | - | XX | XXX | Cyanobacteria, diatoms, plants [58] |
| 13.882 | Pheophytin | - | - | - | XXX | Bacteria, algae, plants [65] |
| 14.084 | Betacarotene | - | X | - | - | Bacteria (Chloroflexi, Acidobacteria) [59], algae, diatoms, plants [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boidi, F.J.; Mlewski, E.C.; Fernández, G.C.; Flores, M.R.; Gérard, E.; Farías, M.E.; Gomez, F.J. Community Vertical Composition of the Laguna Negra Hypersaline Microbial Mat, Puna Region (Argentinean Andes). Biology 2022, 11, 831. https://doi.org/10.3390/biology11060831
Boidi FJ, Mlewski EC, Fernández GC, Flores MR, Gérard E, Farías ME, Gomez FJ. Community Vertical Composition of the Laguna Negra Hypersaline Microbial Mat, Puna Region (Argentinean Andes). Biology. 2022; 11(6):831. https://doi.org/10.3390/biology11060831
Chicago/Turabian StyleBoidi, Flavia Jaquelina, Estela Cecilia Mlewski, Guillermo César Fernández, María Regina Flores, Emmanuelle Gérard, María Eugenia Farías, and Fernando Javier Gomez. 2022. "Community Vertical Composition of the Laguna Negra Hypersaline Microbial Mat, Puna Region (Argentinean Andes)" Biology 11, no. 6: 831. https://doi.org/10.3390/biology11060831
APA StyleBoidi, F. J., Mlewski, E. C., Fernández, G. C., Flores, M. R., Gérard, E., Farías, M. E., & Gomez, F. J. (2022). Community Vertical Composition of the Laguna Negra Hypersaline Microbial Mat, Puna Region (Argentinean Andes). Biology, 11(6), 831. https://doi.org/10.3390/biology11060831