
Preconditioning Methods to Improve Mesenchymal Stromal Cell-Derived Extracellular Vesicles in Bone Regeneration—A Systematic Review

 , and
, and
Abstract
:Simple Summary
Abstract
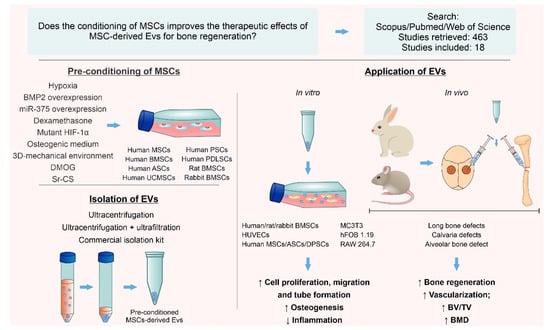
1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Eligibility Criteria
2.3. Study Selection
2.4. Data Extraction
2.5. Risk of Bias Assessment
3. Results
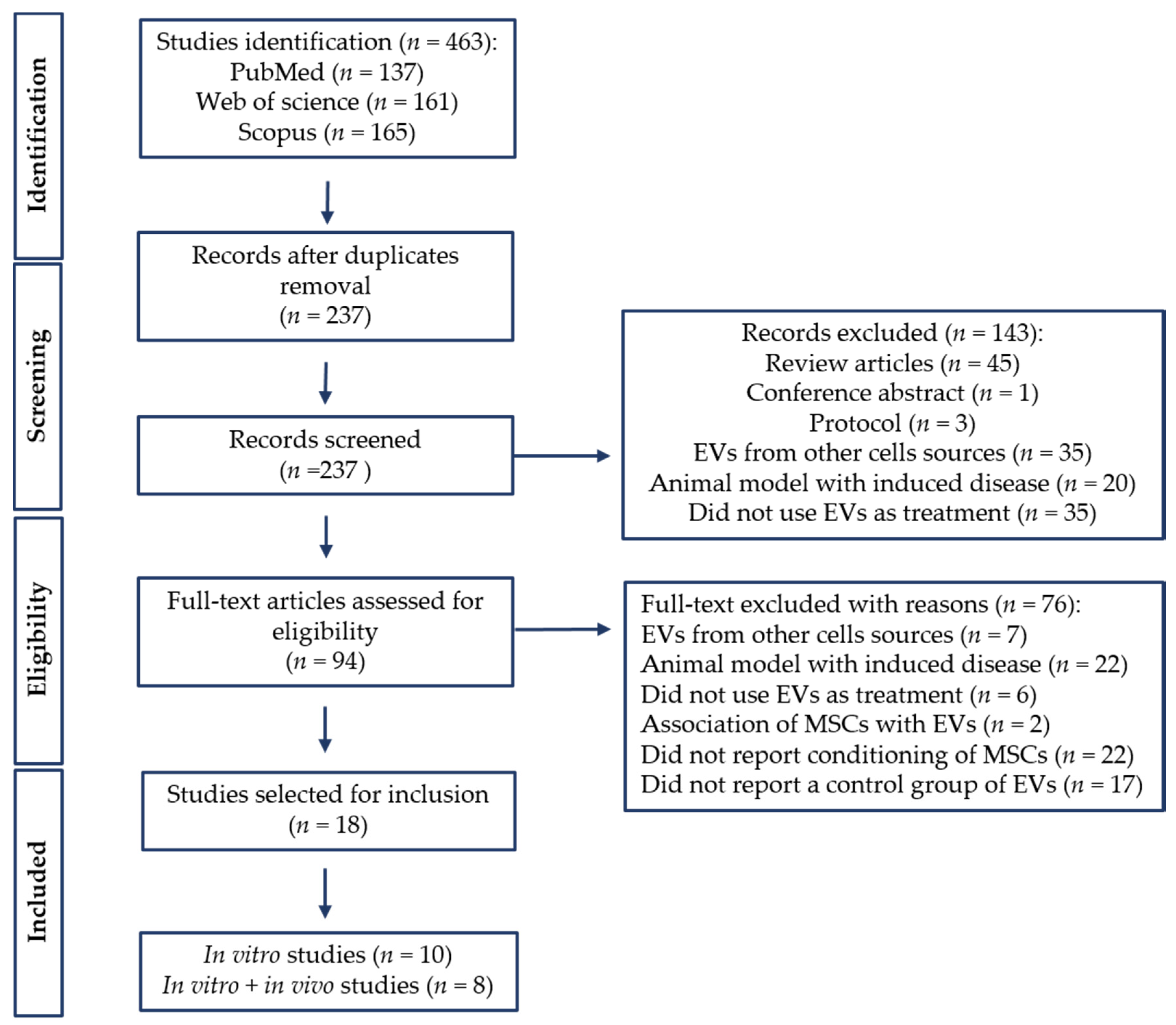
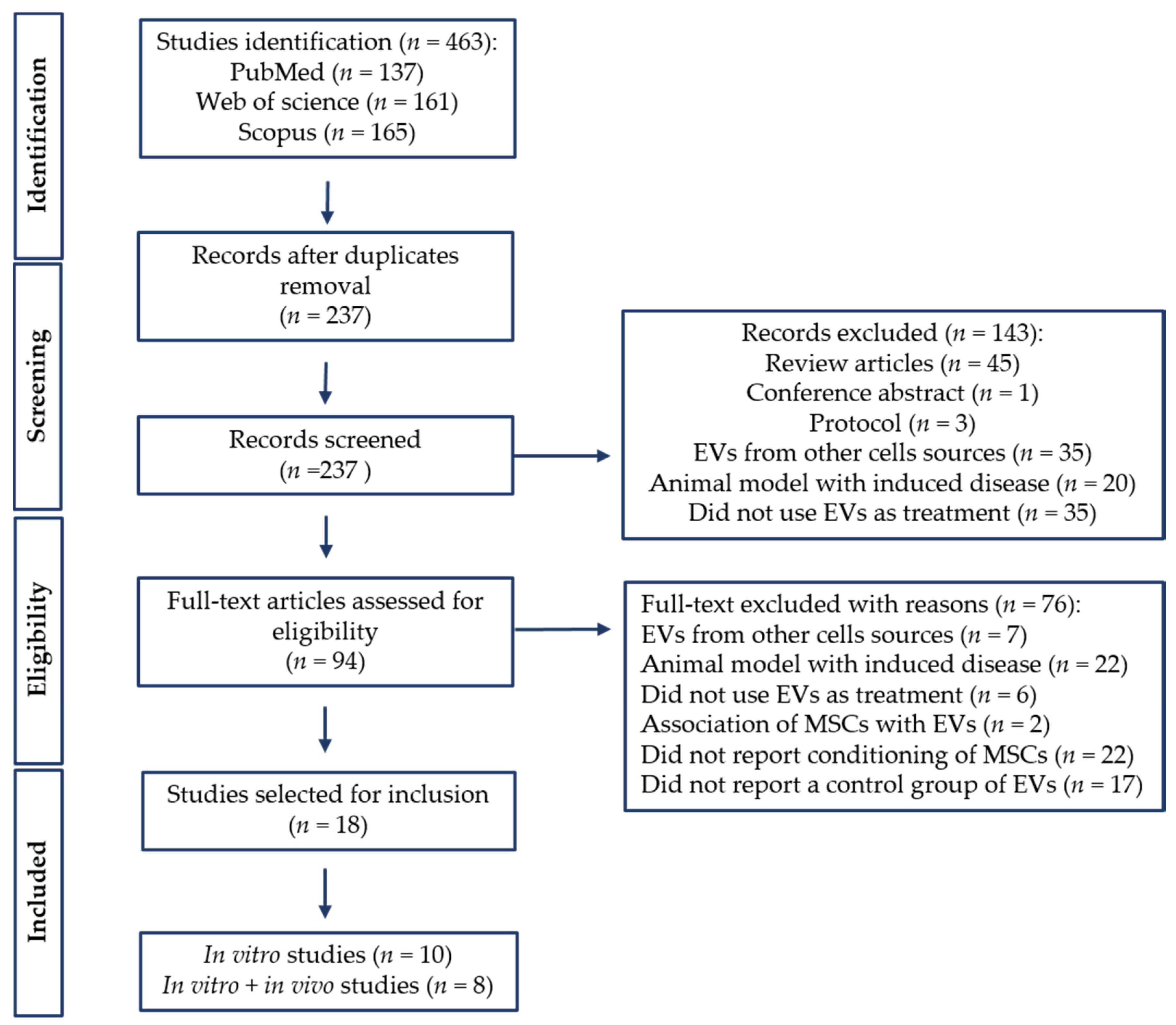
3.1. Search Results
3.2. Study Characteristics
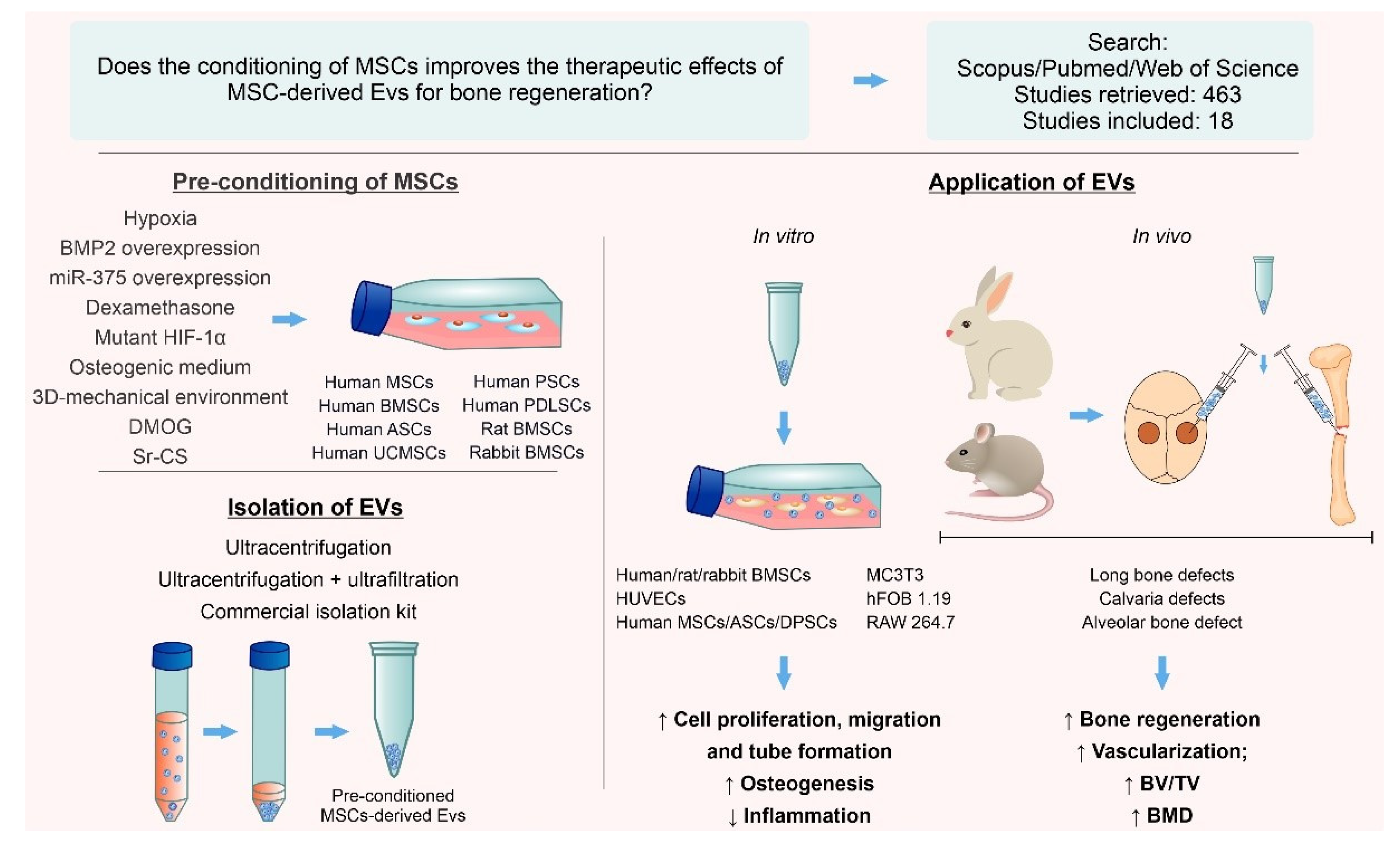
3.3. Conditioning Methods
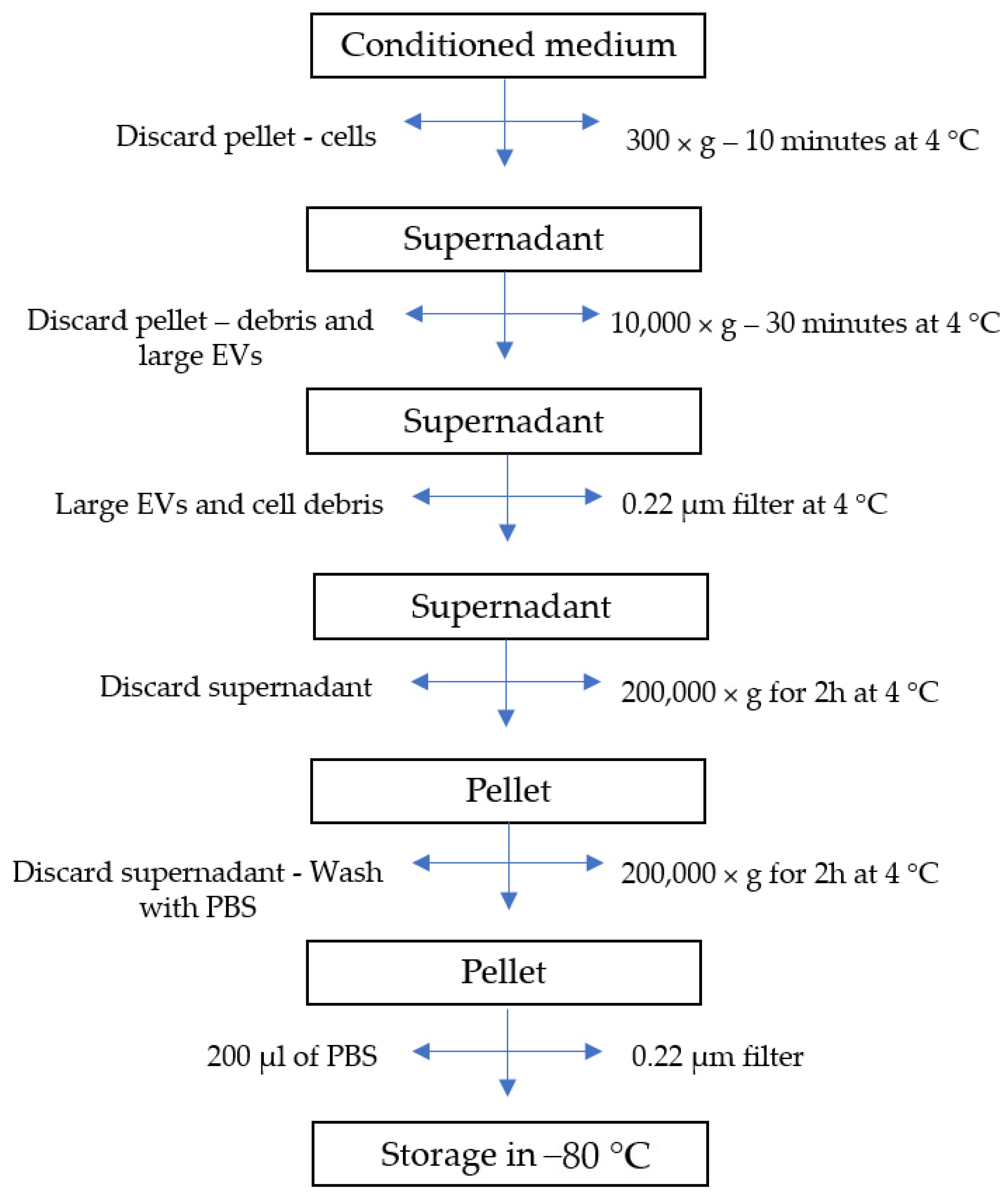
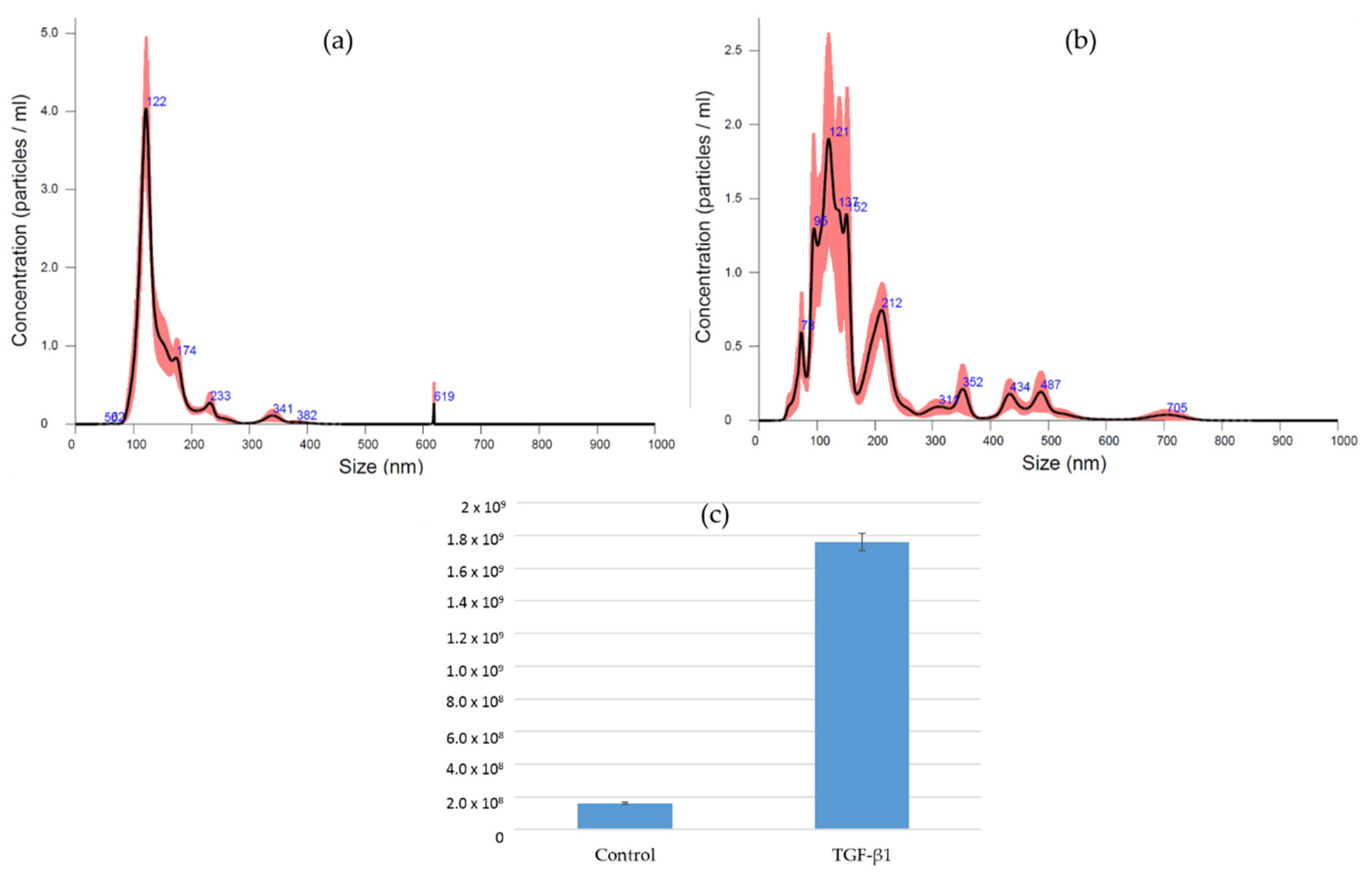
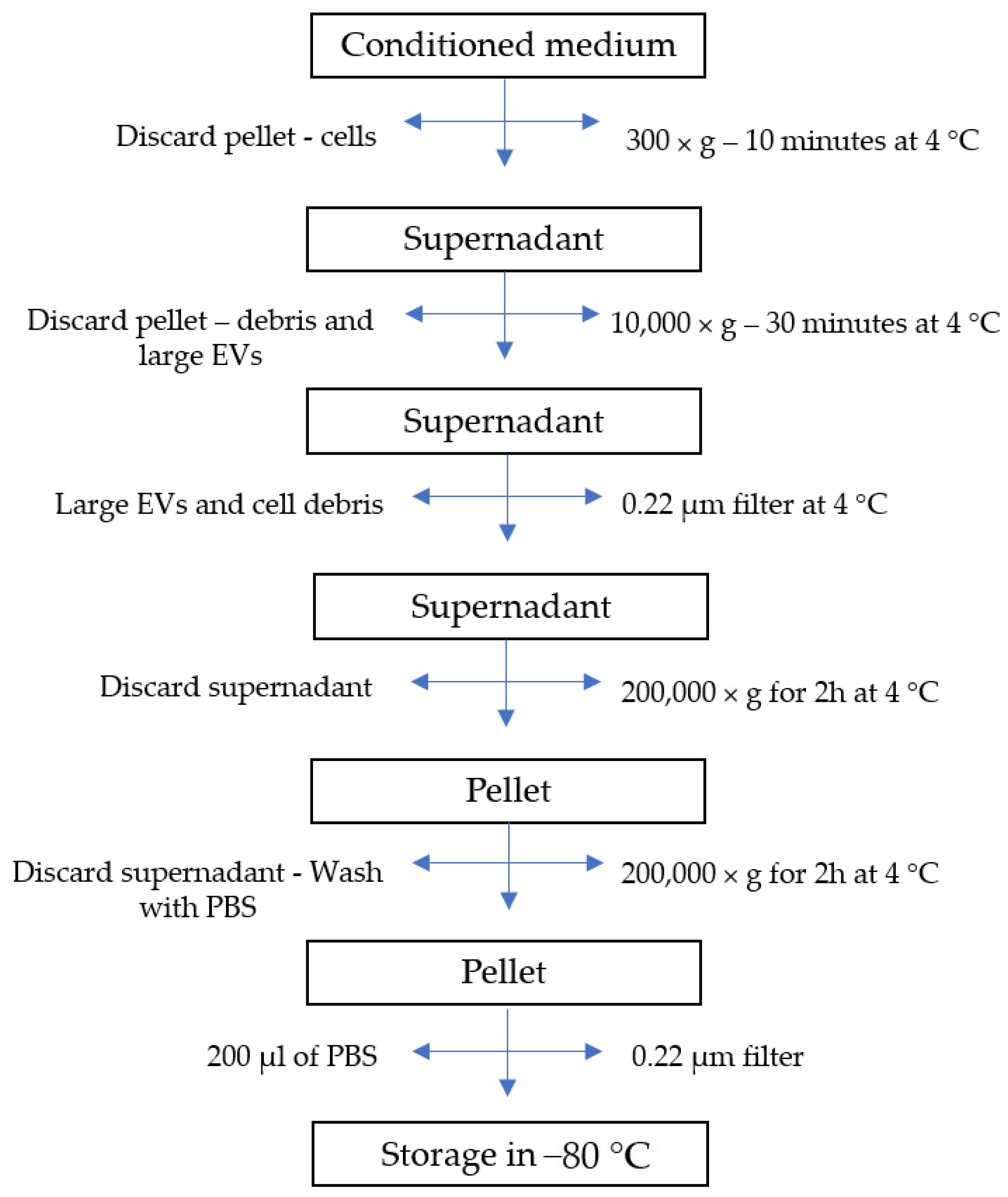
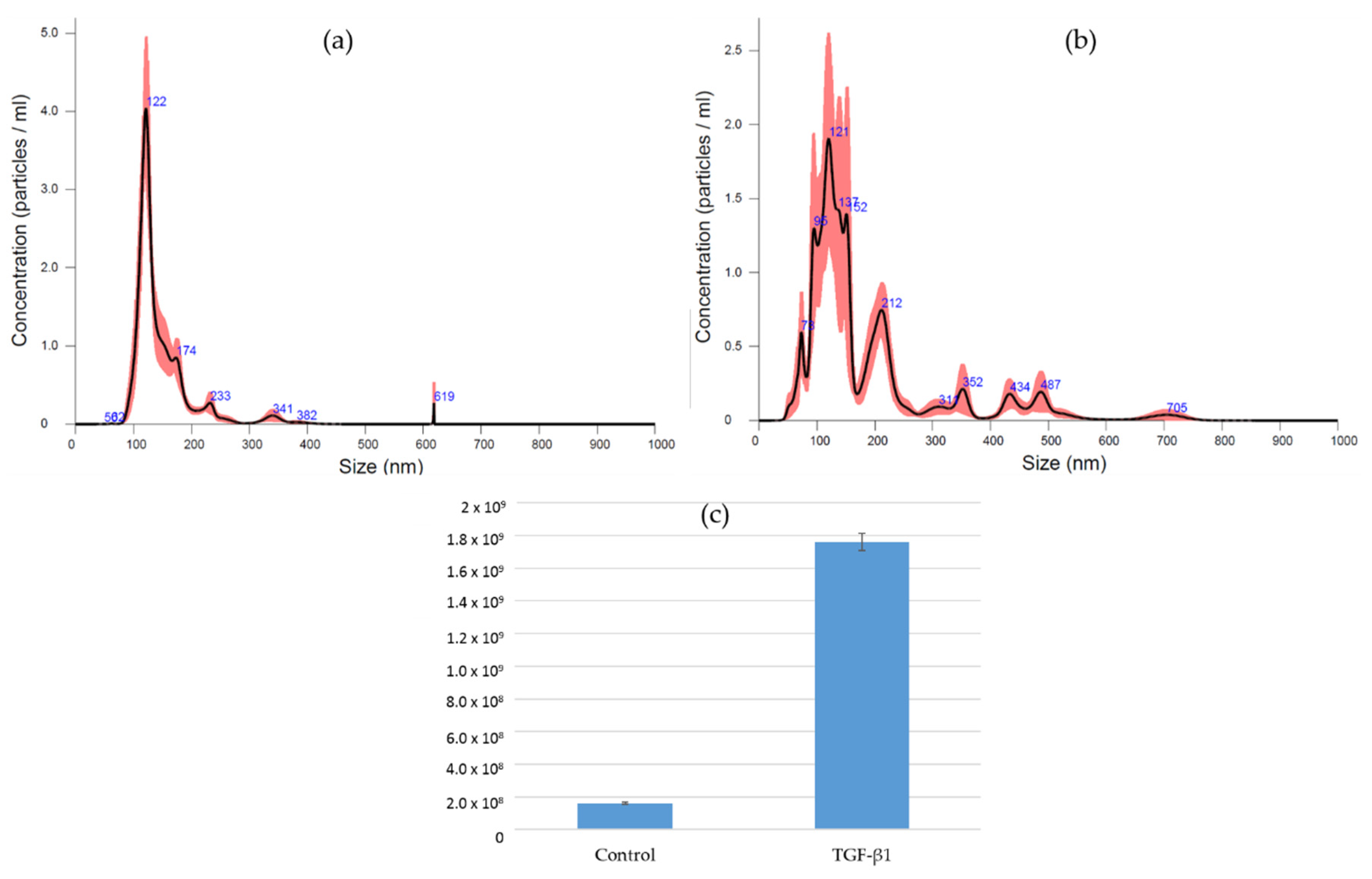
3.4. EV Isolation and Characterization
3.5. In Vitro Studies
3.6. In Vivo Studies
3.7. In Vitro and In Vivo Outcomes
3.7.1. Osteogenic Differentiation
3.7.2. Angiogenesis
3.7.3. Bone Regeneration
3.7.4. Vascularization
3.8. Secondary Outcomes
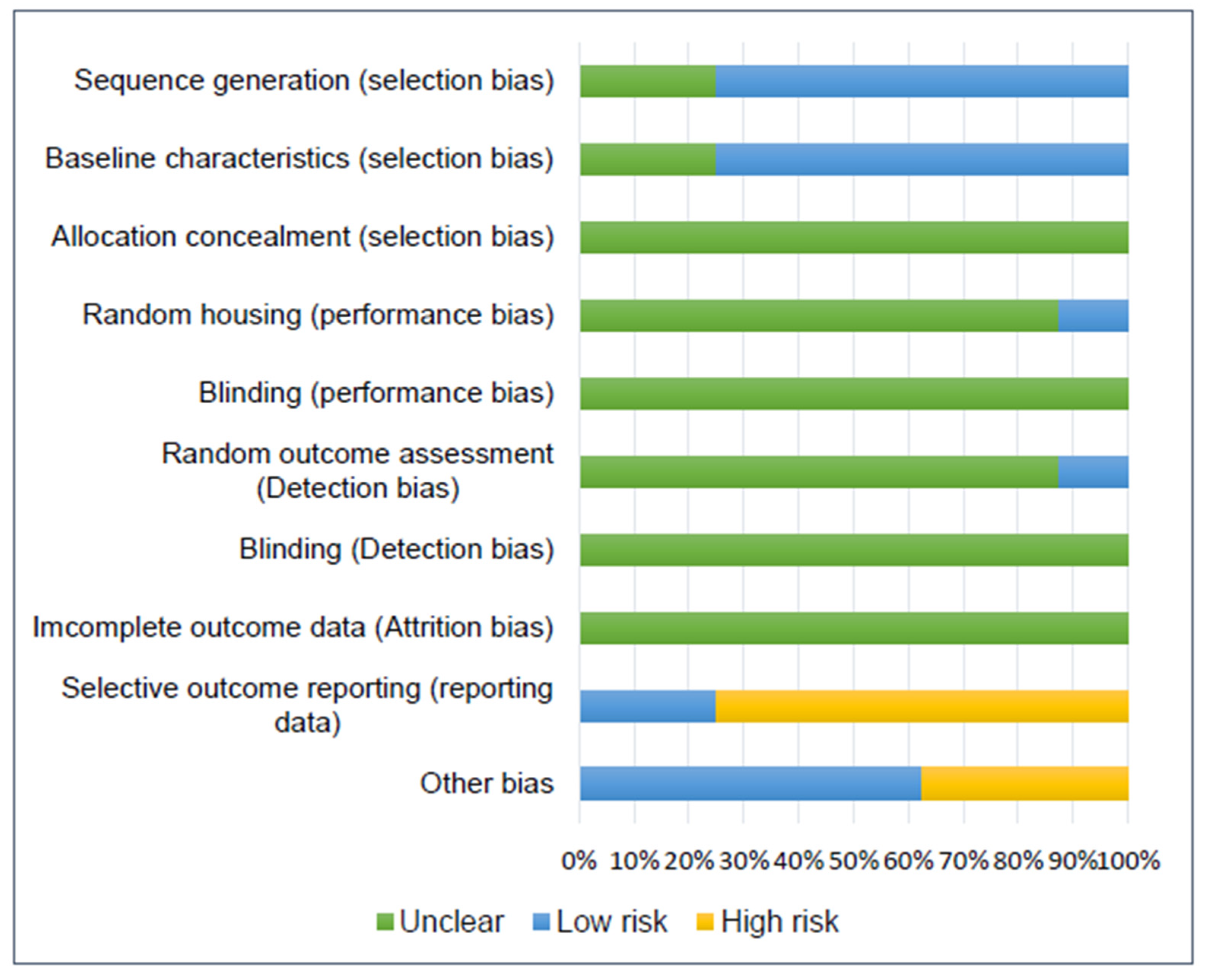
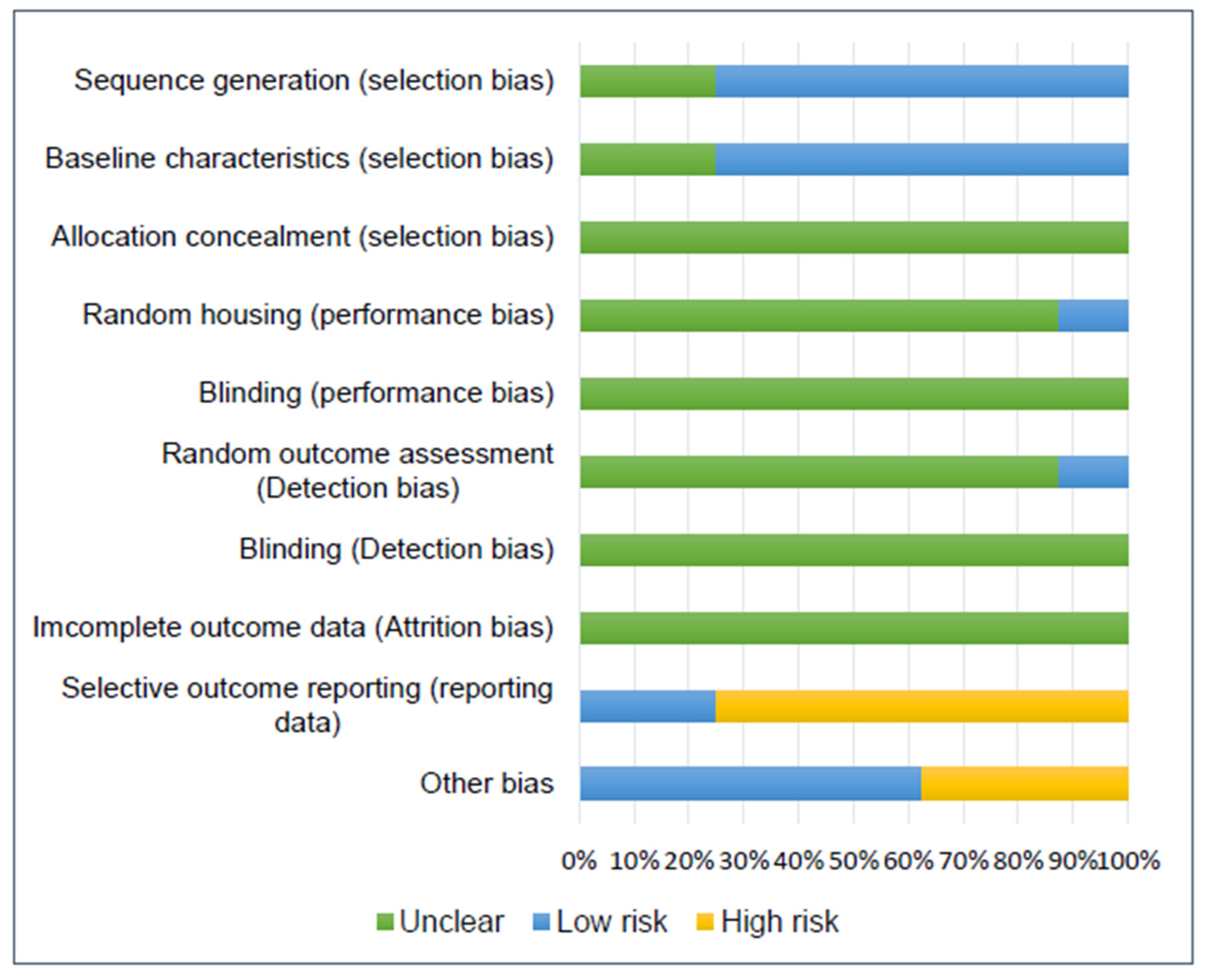
3.9. Risk of Bias Assessment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
- Electronic search strategy
- Pubmed:
- #1: (exosomes[Title/Abstract] OR “extracellular vesicles”[Title/Abstract]) OR (EVs[Title/Abstract])) OR (microvesicles[Title/Abstract])
- #2: (“mesenchymal stem cells”[Title/Abstract]) OR (“mesenchymal stromal cells”[Title/Abstract])) OR (“stem cells”[Title/Abstract])) OR (MSCs[Title/Abstract])))
- #3: (“Bone regeneration”[Title/Abstract]) OR (“Bone healing”[Title/Abstract])) OR (“Bone repair”[Title/Abstract])) OR (osteogenesis[Title/Abstract])) OR (fracture[Title/Abstract])
- #4: #1 AND #2 AND #3
- Scopus:
- #1: TITLE-ABS-KEY (exosomes OR “extracellular vesicles” OR EVs OR microvesicles)
- #2: TITLE-ABS-KEY (osteogenesis OR “bone regeneration” OR fracture OR “bone healing” OR “bone repair”)
- #3: TITLE-ABS-KEY (“mesenchymal stem cells” OR “mesenchymal stromal cells” OR “stem cells” OR mscs))
- #4: #1 AND #2 AND #3 (LIMIT-TO (DOCTYPE, “ar”)) AND (LIMIT-TO (LANGUAGE, “English”))
- Web of Science:
- #1: TS = (exosomes OR “extracellular vesicles” OR EV OR microvesicles)
- #2: TS = (osteogenesis OR “bone regeneration” OR fracture OR “bone healing” OR “bone repair”)
- #3: TS = (“mesenchymal stem cells” OR “mesenchymal stromal cells” OR “stem cells” OR MSCs)
- #4: #1 AND #2 AND #3 (Refined by: DOCUMENT TYPE: (ARTICLE OR EARLY ACCESS).
References
- Zhu, G.; Zhang, T.; Chen, M.; Yao, K.; Huang, X.; Zhang, B.; Li, Y.; Liu, J.; Wang, Y.; Zhao, Z. Bone physiological microenvironment and healing mechanism: Basis for future bone-tissue engineering scaffolds. Bioact. Mater. 2021, 6, 4110–4140. [Google Scholar] [CrossRef] [PubMed]
- Marsell, R.; Einhorn, T.A. The biology of fracture healing. Injury 2011, 42, 551–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roddy, E.; DeBaun, M.R.; Daoud-Gray, A.; Yang, Y.P.; Gardner, M.J. Treatment of critical-sized bone defects: Clinical and tissue engineering perspectives. Eur. J. Orthop. Surg. Traumatol. 2018, 28, 351–362. [Google Scholar] [CrossRef]
- Meirelles, L.D.S.; Fontes, A.M.; Covas, D.T.; Caplan, A.I. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009, 20, 419–427. [Google Scholar] [CrossRef]
- Caplan, A.I. Why are MSCs therapeutic? New data: New insight. J. Pathol. 2009, 217, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I.; Dennis, J.E. Mesenchymal stem cells as trophic mediators. J. Cell. Biochem. 2006, 98, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, Y.; Jiang, Y.; Du, J.; Xu, J.; Liu, Y. Systemic therapy of MSCs in bone regeneration: A systematic review and meta-analysis. Stem Cell Res. Ther. 2021, 12, 377. [Google Scholar] [CrossRef]
- Liao, Y.; Zhang, X.-L.; Li, L.; Shen, F.-M.; Zhong, M.-K. Stem cell therapy for bone repair: A systematic review and meta-analysis of preclinical studies with large animal models. Br. J. Clin. Pharmacol. 2014, 78, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Egido-Moreno, S.; Valls-Roca-Umbert, J.; Céspedes-Sánchez, J.; López-López, J.; Velasco-Ortega, E. Clinical Efficacy of Mesenchymal Stem Cells in Bone Regeneration in Oral Implantology. Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public Health 2021, 18, 894. [Google Scholar] [CrossRef]
- García-Bernal, D.; García-Arranz, M.; Yáñez, R.M.; Hervás-Salcedo, R.; Cortés, A.; Fernández-García, M.; Hernando-Rodríguez, M.; Quintana-Bustamante, Ó.; Bueren, J.A.; García-Olmo, D.; et al. The Current Status of Mesenchymal Stromal Cells: Controversies, Unresolved Issues and Some Promising Solutions to Improve Their Therapeutic Efficacy. Front. Cell Dev. Biol. 2021, 9, 650664. [Google Scholar] [CrossRef]
- Cocucci, E.; Meldolesi, J. Ectosomes and exosomes: Shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015, 25, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stahl, P.D. Extracellular vesicles: A new communication paradigm? Nat. Rev. Mol. Cell Biol. 2019, 20, 509–510. [Google Scholar] [CrossRef]
- Presen, D.M.; Traweger, A.; Gimona, M.; Redl, H. Mesenchymal Stromal Cell-Based Bone Regeneration Therapies: From Cell Transplantation and Tissue Engineering to Therapeutic Secretomes and Extracellular Vesicles. Front. Bioeng. Biotechnol. 2019, 7, 352. [Google Scholar] [CrossRef]
- Zhang, L.; Jiao, G.; Ren, S.; Zhang, X.; Li, C.; Wu, W.; Wang, H.; Liu, H.; Zhou, H.; Chen, Y. Exosomes from bone marrow mesenchymal stem cells enhance fracture healing through the promotion of osteogenesis and angiogenesis in a rat model of nonunion. Stem Cell Res. Ther. 2020, 11, 38. [Google Scholar] [CrossRef]
- Liu, W.; Li, L.; Rong, Y.; Qian, D.; Chen, J.; Zhou, Z.; Luo, Y.; Jiang, D.; Cheng, L.; Zhao, S.; et al. Hypoxic mesenchymal stem cell-derived exosomes promote bone fracture healing by the transfer of miR-126. Acta Biomater. 2020, 103, 196–212. [Google Scholar] [CrossRef]
- Ying, C.; Wang, R.; Wang, Z.; Tao, J.; Yin, W.; Zhang, J.; Yi, C.; Qi, X.; Han, D. BMSC-Exosomes Carry Mutant HIF-1α for Improving Angiogenesis and Osteogenesis in Critical-Sized Calvarial Defects. Front. Bioeng. Biotechnol. 2020, 8, 565561. [Google Scholar] [CrossRef]
- Chen, S.; Zheng, Y.; Zhang, S.; Jia, L.; Zhou, Y. Promotion Effects of miR-375 on the Osteogenic Differentiation of Human Adipose-Derived Mesenchymal Stem Cells. Stem Cell Rep. 2017, 8, 773–786. [Google Scholar] [CrossRef] [Green Version]
- Liang, B.; Liang, J.-M.; Ding, J.-N.; Xu, J.; Xu, J.-G.; Chai, Y.-M. Dimethyloxaloylglycine-stimulated human bone marrow mesenchymal stem cell-derived exosomes enhance bone regeneration through angiogenesis by targeting the AKT/mTOR pathway. Stem Cell Res. Ther. 2019, 10, 335. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-C.; Kang, M.; Lu, Y.; Shirazi, S.; Diaz, J.I.; Cooper, L.F.; Gajendrareddy, P.; Ravindran, S. Functionally engineered extracellular vesicles improve bone regeneration. Acta Biomater. 2020, 109, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Zhang, J.; Yuan, H.; Xu, Z.; Li, Q.; Niu, X.; Hu, B.; Wang, Y.; Li, X. Exosomes Secreted by Human-Induced Pluripotent Stem Cell-Derived Mesenchymal Stem Cells Repair Critical-Sized Bone Defects through Enhanced Angiogenesis and Osteogenesis in Osteoporotic Rats. Int. J. Biol. Sci. 2016, 12, 836–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, S.; Li, Y.; Chen, P. Osteogenic effect of bone marrow mesenchymal stem cell-derived exosomes on steroid-induced osteonecrosis of the femoral head. Drug Des. Dev. Ther. 2019, 13, 45–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.-C.; Kang, K.-S. Functional enhancement strategies for immunomodulation of mesenchymal stem cells and their therapeutic application. Stem Cell Res. Ther. 2020, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-S.; Bandeira, E.; Shelke, G.V.; Lässer, C.; Lötvall, J. Enhancement of therapeutic potential of mesenchymal stem cell-derived extracellular vesicles. Stem Cell Res. Ther. 2019, 10, 288. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; The PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.M.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, D.; Li, C.; Zhou, S.; Tian, D.; Xiao, D.; Zhang, H.; Gao, F.; Huang, J. Exosomes secreted from mutant-HIF-1α-modified bone-marrow-derived mesenchymal stem cells attenuate early steroid-induced avascular necrosis of femoral head in rabbit. Cell Biol. Int. 2017, 41, 1379–1390. [Google Scholar] [CrossRef]
- Li, W.; Liu, Y.; Zhang, P.; Tang, Y.; Zhou, M.; Jiang, W.; Zhang, X.; Wu, G.; Zhou, Y. Tissue-Engineered Bone Immobilized with Human Adipose Stem Cells-Derived Exosomes Promotes Bone Regeneration. ACS Appl. Mater. Interfaces 2018, 10, 5240–5254. [Google Scholar] [CrossRef]
- Zhao, M.; Li, P.; Xu, H.; Pan, Q.; Zeng, R.; Ma, X.; Li, Z.; Lin, H. Dexamethasone-Activated MSCs Release MVs for Stimulating Osteogenic Response. Stem Cells Int. 2018, 2018, 7231739. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Guan, Z.; Zhang, M.; Lyu, S.; Thuaksuban, N.; Kamolmattayakul, S.; Nuntanaranont, T.; Yazawa, T. Exosomal circLPAR1 Promoted Osteogenic Differentiation of Homotypic Dental Pulp Stem Cells by Competitively Binding to hsa-miR-31. BioMed Res. Int. 2020, 2020, 6319395. [Google Scholar] [CrossRef] [PubMed]
- Pishavar, E.; Copus, J.S.; Atala, A.; Lee, S.J. Comparison Study of Stem Cell-Derived Extracellular Vesicles for Enhanced Osteogenic Differentiation. Tissue Eng. Part A 2021, 27, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Hu, W.; Zou, X.; Xu, J.; He, S.; Chang, L.; Li, X.; Yin, Y.; Tian, M.; Li, Z.; et al. Human Periodontal Ligament Stem Cell-Derived Exosomes Promote Bone Regeneration by Altering MicroRNA Profiles. Stem Cells Int. 2020, 2020, 8852307. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Guo, S.; Tong, S.; Sun, X. Promoting Osteogenic Differentiation of Human Adipose-Derived Stem Cells by Altering the Expression of Exosomal miRNA. Stem Cells Int. 2019, 2019, 1351860. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Li, Z.; Crawford, R.; Xiao, Y.; Zhou, Y. Immunoregulatory role of exosomes derived from differentiating mesenchymal stromal cells on inflammation and osteogenesis. J. Tissue Eng. Regen. Med. 2019, 13, 1978–1991. [Google Scholar] [CrossRef]
- Wang, X.; Omar, O.; Vazirisani, F.; Thomsen, P.; Ekström, K. Mesenchymal stem cell-derived exosomes have altered microRNA profiles and induce osteogenic differentiation depending on the stage of differentiation. PLoS ONE 2018, 13, e0193059. [Google Scholar] [CrossRef]
- Narayanan, R.; Huang, C.-C.; Ravindran, S. Hijacking the Cellular Mail: Exosome Mediated Differentiation of Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 3808674. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Tang, Y.; Liu, Y.; Zhang, P.; Lv, L.; Zhang, X.; Jia, L.; Zhou, Y. Exosomes derived from miR-375-overexpressing human adipose mesenchymal stem cells promote bone regeneration. Cell Prolif. 2019, 52, e12669. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Su, X.; Li, M.; Wan, W.; Li, A.; Zhou, H.; Xu, F. Three-dimensional mechanical microenvironment enhanced osteogenic activity of mesenchymal stem cells-derived exosomes. Chem. Eng. J. 2020, 417, 128040. [Google Scholar] [CrossRef]
- Zhai, M.; Zhu, Y.; Yang, M.; Mao, C. Human Mesenchymal Stem Cell Derived Exosomes Enhance Cell-Free Bone Regeneration by Altering Their miRNAs Profiles. Adv. Sci. 2020, 7, 2001334. [Google Scholar] [CrossRef]
- Liu, L.; Yu, F.; Li, L.; Zhou, L.; Zhou, T.; Xu, Y.; Lin, K.; Fang, B.; Xia, L. Bone marrow stromal cells stimulated by strontium-substituted calcium silicate ceramics: Release of exosomal miR-146a regulates osteogenesis and angiogenesis. Acta Biomater. 2021, 119, 444–457. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.A.; Lemischka, I.R. Stem Cells and Their Niches. Science 2006, 311, 1880–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, P.N. Glucocorticoid-induced osteoporosis. Int. J. Rheum. Dis. 2008, 11, 381–385. [Google Scholar] [CrossRef]
- Dimitriou, R.; Tsiridis, E.; Giannoudis, P.V. Current concepts of molecular aspects of bone healing. Injury 2005, 36, 1392–1404. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Jiang, W.; Phillips, F.M.; Haydon, R.C.; Peng, Y.; Zhou, L.; Luu, H.H.; An, N.; Breyer, B.; Vanichakarn, P.; et al. Osteogenic activity of the fourteen types of human bone morphogenetic proteins (BMPs). J. Bone Jt. Surg. 2003, 85, 1544–1552. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Hammond, S.M. An overview of microRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Shen, X.J.; Zou, Q.; Wang, S.P.; Tang, S.M.; Zhang, G.Z. Biological functions of microRNAs: A review. J. Physiol. Biochem. 2011, 67, 129–139. [Google Scholar] [CrossRef]
- Su, P.; Tian, Y.; Yang, C.; Ma, X.; Wang, X.; Pei, J.; Qian, A. Mesenchymal Stem Cell Migration during Bone Formation and Bone Diseases Therapy. Int. J. Mol. Sci. 2018, 19, 2343. [Google Scholar] [CrossRef] [Green Version]
- Sorrell, J.M.; Baber, M.A.; Caplan, A.I. Influence of Adult Mesenchymal Stem Cells on In Vitro Vascular Formation. Tissue Eng. Part A 2009, 15, 1751–1761. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Meirelles, L.; Caplan, A.I.; Nardi, N.B. In Search of the In Vivo Identity of Mesenchymal Stem Cells. Stem Cells 2008, 26, 2287–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosso, A.; Burger, M.G.; Lunger, A.; Schaefer, D.J.; Banfi, A.; Di Maggio, N. It Takes Two to Tango: Coupling of Angiogenesis and Osteogenesis for Bone Regeneration. Front. Bioeng. Biotechnol. 2017, 5, 68. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.C.; Keith, B. The role of oxygen availability in embryonic development and stem cell function. Nat. Rev. Mol. Cell Biol. 2008, 9, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Lu, K.; Zhang, N.; Zhao, Y.; Ma, Q.; Shen, J.; Lin, Y.; Xiang, P.; Tang, Y.; Hu, X.; et al. Myocardial reparative functions of exosomes from mesenchymal stem cells are enhanced by hypoxia treatment of the cells via transferring microRNA-210 in an nSMase2-dependent way. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1659–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Cai, Y.; Luo, L.M.; Liu, H.B. Hypoxic adipose mesenchymal stem cells derived conditioned medium protects myocardial infarct in rat. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4397–4406. [Google Scholar] [PubMed]
- Zhang, Y.; Ma, L.; Su, Y.; Su, L.; Lan, X.; Wu, D.; Han, S.; Li, J.; Kvederis, L.; Corey, S.; et al. Hypoxia conditioning enhances neuroprotective effects of aged human bone marrow mesenchymal stem cell-derived conditioned medium against cerebral ischemia in vitro. Brain Res. 2019, 1725, 146432. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, Y.; Shen, L.; Ding, W.; Chen, X.; Wu, E.; Cai, K.; Wang, G. Hypoxic Preconditioning Augments the Therapeutic Efficacy of Bone Marrow Stromal Cells in a Rat Ischemic Stroke Model. Cell. Mol. Neurobiol. 2017, 37, 1115–1129. [Google Scholar] [CrossRef]
- Takahashi, H.; Ohnishi, S.; Yamamoto, Y.; Hayashi, T.; Murao, N.; Osawa, M.; Maeda, T.; Ishikawa, K.; Sakamoto, N.; Funayama, E. Topical Application of Conditioned Medium from Hypoxically Cultured Amnion-Derived Mesenchymal Stem Cells Promotes Wound Healing in Diabetic Mice. Plast. Reconstr. Surg. 2021, 147, 1342–1352. [Google Scholar] [CrossRef]
- Jun, E.K.; Zhang, Q.; Yoon, B.S.; Moon, J.-H.; Lee, G.; Park, G.; Kang, P.J.; Lee, J.H.; Kim, A.; You, S. Hypoxic Conditioned Medium from Human Amniotic Fluid-Derived Mesenchymal Stem Cells Accelerates Skin Wound Healing through TGF-β/SMAD2 and PI3K/Akt Pathways. Int. J. Mol. Sci. 2014, 15, 605–628. [Google Scholar] [CrossRef] [Green Version]
- Ho, S.S.; Hung, B.P.; Heyrani, N.; Lee, M.A.; Leach, J.K. Hypoxic Preconditioning of Mesenchymal Stem Cells with Subsequent Spheroid Formation Accelerates Repair of Segmental Bone Defects. Stem Cells 2018, 36, 1393–1403. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.; Kim, R.; Park, S.I.; Jung, Y.J.; Ham, O.; Lee, J.; Kim, J.H.; Oh, S.; Lee, M.Y.; Kim, J.; et al. Enhanced Healing of Rat Calvarial Bone Defects with Hypoxic Conditioned Medium from Mesenchymal Stem Cells through Increased Endogenous Stem Cell Migration via Regulation of ICAM-1 Targeted-microRNA-221. Mol. Cells 2015, 38, 643–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, T.-G.; Zhao, X.; Yang, Q.; Li, Y.; Ge, C.; Zhao, G.; Franceschi, R.T. Physical and functional interactions between Runx2 and HIF-1α induce vascular endothelial growth factor gene expression. J. Cell. Biochem. 2011, 112, 3582–3593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamiya, H.; Ikeda, T.; Jeong, J.-H.; Saito, T.; Yano, F.; Jung, Y.-K.; Ohba, S.; Kawaguchi, H.; Chung, U.-I.; Choi, J.-Y. Analysis of the Runx2 promoter in osseous and non-osseous cells and identification of HIF2A as a potent transcription activator. Gene 2008, 416, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Ahluwalia, A.; Tarnawski, A.S. Critical Role of Hypoxia Sensor—HIF-1α in VEGF Gene Activation. Implications for Angiogenesis and Tissue Injury Healing. Curr. Med. Chem. 2012, 19, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Jaakkola, P.; Mole, D.R.; Tian, Y.-M.; Wilson, M.I.; Gielbert, J.; Gaskell, S.J.; von Kriegsheim, A.; Hebestreit, H.F.; Mukherji, M.; Schofield, C.J.; et al. Targeting of HIF-alpha to the von Hippel-Lindau Ubiquitylation Complex by O2-Regulated Prolyl Hydroxylation. Science 2001, 292, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Ogle, M.E.; Gu, X.; Espinera, A.R.; Wei, L. Inhibition of prolyl hydroxylases by dimethyloxaloylglycine after stroke reduces ischemic brain injury and requires hypoxia inducible factor-1α. Neurobiol. Dis. 2012, 45, 733–742. [Google Scholar] [CrossRef] [Green Version]
- Esmaeilzadeh, B.; Artimani, T.; Amiri, I.; Najafi, R.; Shahidi, S.; Sabec, M.; Farzadinia, P.; Zare, M.; Zahiri, M.; Asl, S.S. Dimethyloxalylglycine preconditioning enhances protective effects of bone marrow-derived mesenchymal stem cells in Aβ- induced Alzheimer disease. Physiol. Behav. 2019, 199, 265–272. [Google Scholar] [CrossRef]
- Liu, X.-B.; Wang, J.-A.; Ji, X.-Y.; Yu, S.P.; Wei, L. Preconditioning of bone marrow mesenchymal stem cells by prolyl hydroxylase inhibition enhances cell survival and angiogenesis in vitro and after transplantation into the ischemic heart of rats. Stem Cell Res. Ther. 2014, 5, 111. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Chen, S.; Song, W.-Q.; Gao, Y.-S.; Guan, J.-J.; Wang, Y.; Sun, Y.; Zhang, C.-Q. Dimethyloxaloylglycine Improves Angiogenic Activity of Bone Marrow Stromal Cells in the Tissue-Engineered Bone. Int. J. Biol. Sci. 2014, 10, 746–756. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Guan, J.; Changqing, Z.; Ding, H.; Jieyuan, Z.; Xie, Z.; Chen, C.; Li, X.; Zhang, C.; Huang, Y. Dimethyloxaloylglycine Promotes the Angiogenic Activity of Mesenchymal Stem Cells Derived from iPSCs via Activation of the PI3K/Akt Pathway for Bone Regeneration. Int. J. Biol. Sci. 2016, 12, 639–652. [Google Scholar] [CrossRef] [Green Version]
- Marx, D.; Yazdi, A.R.; Papini, M.; Towler, M. A review of the latest insights into the mechanism of action of strontium in bone. Bone Rep. 2020, 12, 100273. [Google Scholar] [CrossRef] [PubMed]
- Cianferotti, L.; D’Asta, F.; Brandi, M.L. A review on strontium ranelate long-term antifracture efficacy in the treatment of postmenopausal osteoporosis. Ther. Adv. Musculoskelet. Dis. 2013, 5, 127–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saidak, Z.; Marie, P.J. Strontium signaling: Molecular mechanisms and therapeutic implications in osteoporosis. Pharmacol. Ther. 2012, 136, 216–226. [Google Scholar] [CrossRef]
- Lin, K.; Xia, L.; Li, H.; Jiang, X.; Pan, H.; Xu, Y.; Lu, W.W.; Zhang, Z.; Chang, J. Enhanced osteoporotic bone regeneration by strontium-substituted calcium silicate bioactive ceramics. Biomaterials 2013, 34, 10028–10042. [Google Scholar] [CrossRef] [PubMed]
- Thormann, U.; Ray, S.; Sommer, U.; El Khassawna, T.; Rehling, T.; Hundgeburth, M.; Henß, A.; Rohnke, M.; Janek, J.; Lips, K.S.; et al. Bone formation induced by strontium modified calcium phosphate cement in critical-size metaphyseal fracture defects in ovariectomized rats. Biomaterials 2013, 34, 8589–8598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glatt, V.; Evans, C.H.; Tetsworth, K. A Concert between Biology and Biomechanics: The Influence of the Mechanical Environment on Bone Healing. Front. Physiol. 2017, 7, 678. [Google Scholar] [CrossRef] [Green Version]
- Vogel, V.; Sheetz, M. Local force and geometry sensing regulate cell functions. Nat. Rev. Mol. Cell Biol. 2006, 7, 265–275. [Google Scholar] [CrossRef]
- Poniatowski, Ł.; Wojdasiewicz, P.; Gasik, R.; Szukiewicz, D. Transforming Growth Factor Beta Family: Insight into the Role of Growth Factors in Regulation of Fracture Healing Biology and Potential Clinical Applications. Mediat. Inflamm. 2015, 2015, 137823. [Google Scholar] [CrossRef] [Green Version]
- Dubon, M.J.; Yu, J.; Choi, S.; Park, K.S. Transforming growth factor β induces bone marrow mesenchymal stem cell migration via noncanonical signals and N-cadherin. J. Cell. Physiol. 2018, 233, 201–213. [Google Scholar] [CrossRef]
- Elsafadi, M.; Manikandan, M.; Almalki, S.; Mobarak, M.; Atteya, M.; Iqbal, Z.; Hashmi, J.A.; Shaheen, S.; Alajez, N.; Alfayez, M.; et al. TGFβ1-Induced Differentiation of Human Bone Marrow-Derived MSCs Is Mediated by Changes to the Actin Cytoskeleton. Stem Cells Int. 2018, 2018, 6913594. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal research: Reporting in vivo experiments: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Conditioning | Conditioning Method | Reference |
|---|---|---|
| Chemical induction | Standard osteogenic medium | [29,31,33,36,40] |
| Dimethyloxalylglycine | [20] | |
| Dexamethasone | [30] | |
| Strontium-substituted calcium silicate ceramics | [41] | |
| Chemical induction + culture conditions | Standart osteogenic medium + serum deprivation | [32,34,35,37] |
| Genetic modification | miR-375-overexpressing ASCs | [38] |
| Genetic modification + culture conditions | BMP2-overexpressing BMSCs + serum deprivation | [21] |
| Mutant HIF-1α-modified BMSCs + serum deprivation | [18,28] | |
| Culture conditions | Hypoxia | [17] |
| Three-dimensional mechanical microenvironment | [39] |
| EV Source/Origin | Type of EV | Isolation Method | Characterization Method | EV Markers | Size Distribution | Reference |
|---|---|---|---|---|---|---|
| Human ASCs | Exosomes | Ultracentrifugation | TEM, NTA, WB | CD63, CD9 | 33–177 nm | [29] |
| Human ASCs | Exosomes | Ultracentrifugation | TEM, NTA, WB | CD9, CD63 | ~75 nm | [38] |
| Human ASCs | Exosomes | Ultracentrifugation | TEM, WB | TSG101, CD9 | 30–150 nm | [34] |
| Human BMSCs | Exosomes | ExoQuick TC isolation kit, System Biosciences | TEM | NR | NR | [37] |
| Human BMSCs | Exosomes | Ultracentrifugation | TEM, WB, qNano platform | CD9, CD63, GM130, TSG101 | 30–100 nm | [20] |
| Human BMSCs | Exosomes | Ultracentrifugation | TEM, NTA | NR | ~100 nm | [35] |
| Human BMSCs | Extracellular vesicles | ExoQuick TC isolation kit, System Biosciences | TEM, NTA, immunoblotting | CD63, CD9 | 100–150 nm | [21] |
| Human BMSCs and Human PSCs | Extracellular vesicles | Ultrafiltration + ultracentrifugation | TEM, DLS | NR | BMSC: ~137–262 nm PSC: ~130–202 nm | [32] |
| Human DPSCs | Exosomes | Ultracentrifugation | TEM, NTA, flow cytometry | CD63, CD81 | 20–120 nm | [31] |
| Human MSCs * | Exosomes | Ultracentrifugation | TEM, NTA | NR | 30–150 nm | [36] |
| Human MSCs * | Exosomes | ExoQuick TC isolation kit, System Biosciences | AFM, TEM, NTA, WB | CD63 | ~143 nm | [40] |
| Human PDLSCs | Exosomes | ExoQuick TC isolation kit, System Biosciences | TEM, NTA, WB | CD63, CD81 | 30–200 nm | [39] |
| Human PDLSCs | Exosomes | Ultracentrifugation | TEM, NTA, WB | CD63, TSG101 | ~120 nm | [33] |
| Human UCMSCs | Exosomes | Ultrafiltration + sucrose + ultracentrifugation | TEM, NTA, WB | TSG101, CD9, CD63, CD81 | 50–150 nm | [17] |
| Rabbit BMSCs | Exosomes | Exosome isolation kit, Invitrogen | TEM, WB, qNano Platform | CD9, CD63, CD81 | 75–150 nm | [28] |
| Rat BMSCs | Microvesicles | Differential centrifugation ** | TEM, NTA, flow cytometry | CD90 | 100–400 nm | [30] |
| Rat BMSCs | Exosomes | Ultrafiltration + sucrose + ultracentrifugation | TRPS, WB | CD9, CD63, CD81 | 50–150 nm | [18] |
| Rat BMSCs | Exosomes | Ultracentrifugation and ultrafiltration | TEM, NTA, WB | CD9, CD63, CD81 | ~130 nm | [41] |
| MSC Conditioning Method | Culture Method for Control EVs | Exposure to EVs | Treatment/Groups | Analysis | EV Concentration | Outcome | Ref. |
|---|---|---|---|---|---|---|---|
| Human ACSs conditioned with osteogenic medium for 2, 4, 7, and 14 days (xd-Exo) | Human ASCs cultured with normal medium without stimuli (0d-Exo) | Human BMSCs | Cells cultured in OM or PM with 0d-Exo, 2d-Exo, 4d-Exo, 7d-Exo, and 14d-Exo | ALP activity | 10, 25, or 50 μg/mL | ↑ in cell differentiation in OM by osteogenically induced exosomes at 25 μg/mL exosomes. | [29] |
| Human ASCs conditioned with osteogenic medium for 14 days + serum-free medium for 24 h (Exo-D14) | Human ASCs cultured with serum-free medium for 24 h (Exo-D0) | Human ASCs | Exo-D0 and Exo-D14 (both cultured in OM); negative control (PM); positive control (OM) | ALP activity, ARS, osteogenic markers | 20 µg/mL | ↑ in cell differentiation in Exo-D14-treated ASCs compared to Exo-D0. | [34] |
| Human BMSCs conditioned with osteogenic medium for 2 and 4 weeks + serum-free medium for 24 h (two- or four-week exosomes) | Human BMSCs cultured in serum-free medium for 24 h (regular exosomes) | Human BMSCs | 2D culture: two-week exosomes; four-week exosomes; regular exosomes; control. 3D type I collagen: four-week exosomes; regular exosomes; control | Pro-osteogenic gene analysis | NR (exosomes isolated from 5 × 105 cells) | 2D culture: upregulation of pro-osteogenic genes by two- and four-week exosomes. 3D culture: both four-week exosomes and regular exosomes upregulate pro-osteogenic genes. This article did not compare the results between the three types of exosomes. | [37] |
| Human BMSCs conditioned with osteogenic medium for 3, 7, or 14 days + serum-free medium for 12 h (xd-Exo) | Human BMSCs cultured with serum-free medium for 12 h (0d-Exo) | hBMSCs RAW 264.7 | Human BMSCs cultured in OM treated with 0d-Exo, 3d-Exo, 7d-Exo, and 14d-Exo | ALP activity, osteogenic markers | 10 µg exosome protein | ↑ in cell migration by 3d-Exo, 7d-Exo, and 14d-Exo. ↑ in ALP activity, OPN, ALP, RUNX2, BMP2, and BMP7 when treated with 0d-Exo. ↓ in inflammation by 0d-Exo, 3d-Exo, and 7d-Exo. | [35] |
| Human BMSCs treated with 0d-Exo, 3d-Exo, 7d-Exo, and 14d-Exo | Cell viability, cell migration | Cell viability: NR cell migration:10 µg exosome protein | |||||
| RAW 264.7 treated with 0d-Exo, 3d-Exo, 7d-Exo, and 14d-Exo | Cytokine expression | 10 µg exosome protein | |||||
| Human BMSCs and PSCs conditioned with osteogenic medium for 7 and 21 days + serum-free medium for 72 h (Dx) | Human PSCs and BMSCs cultured in serum-free medium for 72 h (D0) | Human BMSCs | Cells cultured in OM treated with D0, D7, and D21 derived from PSCs and BMSCs | ALP activity, ARS | 10 μg/mL | ↑ in cell differentiation by both BMSC- and PSC-derived exosomes, mainly by 21-day osteogenically induced exosomes. | [32] |
| Human DPSCs conditioned with osteogenic medium for 7 days (Exo7) | Human DPSCs cultured in serum-free medium for 48 h (Exo0) | Human DPSCs | Cells cultured in PM treated with Exo7 and Exo0 positive control (PC; OM), negative control (NC; PM) | ALP activity, ARS, osteogenic markers | NR | ↑ in cell differentiation and expression of RUNX2, COL-1, and OCN by Exo7. | [31] |
| Human MSCs conditioned with osteogenic medium for 3, 6, 9, 12, 15, 18, and 21 days (Exo-Dx) | Human MSCs in passage 6, cultured in normal medium without stimuli (Exo-P6) | HumanMSCs | Cells culture in PM with human fibronectin-coated plates treated with exo-P6, D3, D6, D9, D12, D15, D18, and D21; negative control at day 0 (NCtrl-D0), NCtrl-D14, and NCtrl-D21 | ALP activity, ARS | NR | ↑ in cell differentiation by mid-to-late osteogenically induced exosomes (Exo-D15, Exo-D18, and Exo-21). | [36] |
| Human MSCs conditioned with osteogenic medium for 4, 10, 15, and 20 days (Exo-Dx) | Human MSCs cultured in normal medium without stimuli (Exo-D0) | Human MSCs | Cells cultured in PM treated with Exo-D0, Exo-D4, Exo-D10, Exo-D15, and Exo-D20 | ALP activity, ARS, osteogenic markers (immunofluorescence staining) | 100 μL of 1.0 × 1013 particles per mL | ↑ in cell differentiation by Exo-D10 and Exo-D15 compared to Exo-D0. | [40] |
| Human PDLSCs conditioned with osteogenic medium for 3, 7, and 14 days (Exo-Dx) | Undifferentiated human PDLSCs (Exo-NC) | Rat BMSCs | Cells cultured in OM or PM treated with Exo-D3, Exos_D7, Exos-D14, Exos-NC, and PBS | ALP activity, ARS, osteogenic markers | 50 μg/mL | ↑ in cell differentiation by Exo-D3 and Exo-D14 in PM. ↑ in cell differentiation by Exo-D3, Exo-D7, and Exo-D14 in OM. | [33] |
| Human BMSCs conditioned with dimethyloxaloylglycin (DMOG, 1000 µM) for 48 h (DMOG-MSC-Exos) | Human BMSCs cultured in normal medium without stimuli (MSC-Exos) | HUVECs | MSC-Exos, DMOG-MSC-Exos, or an equivalent volume of PBS | Cell proliferation, migration, and tube formation | 50 µg/mL | ↑ in cell migration and tube formation by DMOG-MSC-Exos with no difference in cell proliferation. | [20] |
| Rat BMSCs conditioned with Dexamethasone (10−8, 10−7, 10−6 M) for 48 h (DXM-MV) | Rat BMSCs cultured in normal medium without stimuli (n-MV) | MC3T3 | Cells cultured in PM with n-MV or DXM-MV (10−8, 10−7, 10−6 M) | ARS, osteogenic markers | NR | ↑ in cell differentiation, migration, and proliferation by DXM-MVs. | [30] |
| Rat BMSCs stimulated by strontium-substituted calcium silicate ceramics for 48 h (Sr-CS-Exo) | Rat BMSCs cultured in normal medium without stimuli (Exo) | HUVECs | Cells treated with PBS, Exo (without stimuli), β-TCP-Exo, CS-Exo, and Sr-CS-Exo | Cell proliferation, cell migration, tube formation, and angiogenesis-related gene expression | Cell proliferation: 50 or 100 μg/mL; cell migration, tube formation, and angiogenesis-related gene expression: NR | ↑ in cell proliferation, migration, tube formation, and gene and protein expression for VEGF and ANG1 by the Sr-CS-Exo. | [41] |
| miR-375-overexpressing human ASCs (Exo (miR-375)) | Human ASC-overexpressing control vector (Exo (NC)) | Human BMSCs | Cells culture in OM or PM treated with Exo (miR-375) and Exo (NC) | ALP activity, ARS, osteogenic markers, cell proliferation | Exo (miR-375): 50 μg/mL; Exo (NC): NR | ↑ in cell differentiation and RUNX2 and OCN expression by Exo(miR-375) in PM and OM. ALP and COL1A1 were upregulated by Exo(miR-375) only in OM. | [38] |
| BMP2-overexpressing human BMSCs + serum-free medium for 24 h (BMP2 FEEs) | Human BMSCs cultured with serum-free medium for 24 h (control EV) | Human BMSCs | BMSCs cultured in collagen sponge with PM treated with BMP2 FEEs or untreated group (PBS) | Osteogenic markers * | 1 × 108 EV particles | BMP2 FEEs increased the expression of BMP2, RUNX2, osterix, and BMP9 in 3D BMSC culture compared to untreated cells. There was no control EV treatment in this analysis. | [21] |
| Cells treated with control EVs and BMP2 FEEs; rhBMP2 (positive control) | SMAD1/5/8 phosphorylation | 6 × 106 EVs for every 30,000 HMSCs. | |||||
| Mutant HIF-1α-modified rabbit BMSCs (BMSC-ExosMU) + serum-free medium for 24 h | Rabbit BMSCs modified by wild-type HIF α (BMSC-ExosWT) + serum-free medium for 24 h | Rabbit BMSCs | BMSC-ExosMU BMSC-ExosWT | ARS, ALP activity, osteogenic markers | 80 μg/mL | ↑ in cell differentiation and expression of OCN and ALP by BMSC-ExosMU. ↑ in HUVEC proliferation, migration, and tube formation by BMSC-ExosMU. | [28] |
| HUVECs | BMSC-ExosMU BMSC-ExosWT | Cell proliferation, cell migration, and tube formation | 80 μg/mL, 40 μg/mL, and 20 μg/mL per group | ||||
| Mutant HIF-1α-modified rat BMSCs + serum-free medium for 48 h (BMSC-Exos-HIF1 α) | Culture method not reported (BMSC-Exos) | Rat BMSCs | Cells cultured in PM treated with: BMSC-Exos-HIF1α, BMSC-Exos, and control | Cell proliferation | 200 μg/mL | ↑ in cell differentiation; proliferation; and expression of ALP, RUNX2, and COL1-a1 by BMSC-Exos-HIF1α. | [18] |
| Cells cultured in OM and treated with BMSC-Exos-HIF1α, BMSC-Exos, and control | ALP activity, ARS, osteogenic markers | 200 μg/mL | |||||
| Human PDLSCs cultured in three-dimensional microscale magnetically stretched collagen hydrogels (SM-Exo) | Human PDLSCs cultured in 3D culture (Exo) | Human BMSCs | Cells cultured with PBS, SM-Exo, and Exo | Cell proliferation, cell migration | 100 μg/mL | ↑ in cell differentiation; proliferation; migration; and ALP, RUNX-2, OCN, and COL-1 expression by SM-Exo. | [39] |
| Cells cultured in OM treated with PBS, SM-Exo, and Exo | Osteogenic markers, ARS | 100 μg/mL | |||||
| Human UCMSCs conditioned with hypoxia (Hypo-Exos) | Human UCMSCs cultured in normoxia (Exos) | HUVECs hFOB 1.19 | Cells treated with PBS, Exos, or Hypo-Exos | Cell proliferation, migration, tube formation, and angiogenesis-related genes | Cell proliferation, migration, and tube formation: 100 μg/mL; angiogenesis-related genes: NR | ↑ in HUVEC proliferation, migration, tube formation, and VEGF expression by Hypo-Exos. ALP, COLA1, and OCN expression in hFOB 1.19 did not differ among groups. | [17] |
| Osteogenic markers | NR |
| Bone Defect Model | Animal Model | Treatment Groups | Concentration of EVs | Scaffold/Vehicle | Time Point Analysis | Analysis | Outcome | Ref. |
|---|---|---|---|---|---|---|---|---|
| Two 5 mm diameter calvarial defects | Sprague–Dawley rats (male) | Blank group: left side defect Hydrogel group (n = 12) Hydrogel + Exo (NC) (n = 12) Hydrogel + Exo (miR-375) (n = 12) | 20 μL Exo (miR-375)or Exo (NC) at 50 μg/mL | Hydrogel (250 μL) | 8 weeks | μCT, histology, and IHC | ↑ BV/TV ratio and BMD; new bone formation; mature osteoid, OCN, and BMP2 by Exo (miR-375). | [38] |
| Two 5 mm diameter calvarial defects | Sprague–Dawley rats (male) | HA group (n = 10) HA + MSC-Exos (n = 10) HA + DMOG-MSC-Exos (n = 10) | 100 μg of exosomes in 200 μL PBS or 200 μL PBS alone | Classical porous hydroxyapatite scaffolds | 8 weeks | μCT, histology, sequential fluorescent labeling, and immunofluorescence staining | ↑ BV/TV ratio and BMD, new bone area by DMOG-MSC-Exos. ↑ new vessel area and IHC for CD31 by DMOG-MSC-Exos. | [20] |
| Two 5 mm diameter calvarial defects | Rats (strain/sex NR) | Control group (collagen alone) Positive control group (rhBMP2 + collagen) Control EV group BMP2 EV group (n = 6 defects per group and time point) | 5 × 108 EVs/50 μL per defect 50 μg/50 μL rhBMP2 per defect | Collagen tape | 4, 8, and 12 weeks | μCT, histology, and IHC | ↑ BV/TV ratio; ongoing woven bone formation; and early expression of BMP2, BSP, DMP1, and OCN by BMP2 EV. Fatty marrow was not present in BMP2 EV group as in rhBMP2 group. | [21] |
| Two 5 mm diameter calvarial defects | Sprague–Dawley rats (sex NR) | β-TCP (n = 13) BMSC-Exos + β-TCP (n = 13) BMSC-Exos-HIF1a + β-TCP (n = 13) | 200 μg of exosomes | β-TCP | 12 weeks | μCT, sequential fluorescent labeling, histomorphology, and IHC | ↑ BMD and BV/TV analysis, new bone area, vessel number and volume, and OCN and CD31 expression by BMSC-Exos-HIF1a + β-TCP group. | [18] |
| Femoral fracture model with Kirschner’s wire | Mice (strain/sex NR) | PBS group (n = 8) Exos group (n = 8) Hypo-Exos group (n = 8) | 200 μg of exosomes in 200 μL of PBS or 200 μL PBS alone | PBS | 7 days | X-ray, μCT, histology, and immunofluorescence staining | ↑ in callus volume/tissue volume, vessel number, vessel volume, and Ki67/CD31-positive cells by Hypo-Exos. | [17] |
| Segmental radius defect (8 mm) | Sprague–Dawley rats (male) | Healthy group Negative group Exo-D0 group Exo-D4 group Exo-D10 group Exo-D15 group hMSC cell-seeded group (n = 5/group) | NR | Poly-L-lysine-coated 3D titanium scaffolds | 4 and 12 weeks | Histology | ↑ new bone formation, osteoblasts, and Haversian canal-like structures by Exo-10 and Exo-15. ↑ collagen by Exo-15. | [40] |
| Alveolar bone defects (4 mm length × 3 mm width × 2 mm height) | Sprague–Dawley rats (male) | Matrigel™ group Exo+ Matrigel™ group SM-Exo + Matrigel™ group Control (PBS) group (n = 8) | 100 μg/100 μL of Matrigel™ or 100 μL of Matrigel™ alone | Matrigel™ | 3 and 6 weeks | μCT, histology, and IHC | ↑ BV/TV ratio, new bone area, and RUNX-2 and OCN expression (IHC) in the SM-Exo + Matrigel group. | [39] |
| Distal femur defects (3.5 mm diameter × 4 mm depth) | Sprague–Dawley rats (male) | SF-PBS group SF-Exo group SF-β-TCP-Exo group SF-CS-Exo group SF-Sr-CS-Exo group (n = 30 femur defects in 15 rats) | 100 μL (1000 μg/mL) | Silk fibroin | 8 weeks | μCT, histology, and IHC | ↑ BV/TV and BMD values; new bone area; and expression levels of CD31, VEGF, and VE-ca in the SF-Sr-CS-Exo group. | [41] |
| microRNA/circRNA Profile | Functions/Signaling Pathways | Reference |
|---|---|---|
| Exo-D3 versus Exo-P6: one miRNA differentially expressed, which enriches three pathways related to osteogenic differentiation. Exo-D21 versus Exo-P6: nine differentially expressed miRNAs that enrich twenty pathways related to osteogenic differentiation. Exo-D21 versus Exo-D3: sixteen differentially expressed miRNAs that enrich twenty pathways related to osteogenic differentiation. | Exo-D3 versus Exo-P6: Hippo signaling pathway, adherens junction, ECM–receptor interaction. Exo-D21 versus Exo-P6: Wnt, Hippo, MAPK, cAMP, PI3K-Akt, TGF-beta, TNF, VEGF, insulin, and AMPK signaling pathways, among others. Exo-D21 versus Exo-D3: Wnt, Hippo, MAPK, cAMP, PI3K-Akt, TGF-β, TNF, HIF-1, insulin, and AMPK signaling pathways, among others. | [36] |
| 11 circRNAs were upregulated in Exo7. ↑ circLPAR1 and ↓ hsa-miR-31 in Exo7-treated DPSCs | circLPAR1 was predicted to bind to hsa-miR-31, a miRNA that showed an inhibitory effect against osteogenic differentiation. circLPAR1 would be the target of hsa-miR-31. Both downregulation of hsa-miR-31 and upregulation of circLPAR1 promoted osteogenic differentiation of DPSCs. | [31] |
| ↑ of 72 miRNAs and ↓ of 35 miRNAs in exosomes derived from osteogenically differentiated PDLSCs. ↑of miR-122-5p, miR-142-5p, miR-25-3p, miR-192-5p. ↓ of miR-125b-5p, let-7b-5p, and miR-100-5p. | Predicted functions: catalytic activity, protein binding, metabolic process, transport, and phosphate-containing compound metabolic process. Processes related to target genes: 2-oxocarboxylic acid metabolism, adipocytokine signaling pathway, AMPK signaling pathway, insulin signaling pathway, and MAPK signaling pathway. | [33] |
| Upregulation of 160, 166, 193, and 136 miRNAs and downregulation of 130, 139, 150, and 191 miRNAs were were observed in the Exo-D4, Exo-D10, Exo-D15, and Exo-D20 exosomes, respectively. ↑ expression of osteogenic miRNAs (Hsa-miR-146a-5p, Hsa-miR-503-5p, Hsa-miR-483-3p, and Hsa-miR-129-5p) and ↓ expression of anti-osteogenic miRNAs (Hsa-miR-32-5p, Hsa-miR-133a-3p, and Hsa-miR-204-5p) in 10- and 15-day osteogenically induced exosomes. | Predicted signaling pathways: PI3K/Akt and MAPK. | [40] |
| ↑ of 201 miRNAs and ↓ of 33 miRNAs in exosomes derived from osteogenically differentiated ASCs. ↑ of five miRNAs (miR-130a-3p, miR-30b-5p, miR-34a-5p, miR-324-5p, and miR-378f) and ↓ of miR-513b-5p. | Predicted processes affected: axon guidance, MAPK signaling, Wnt signaling, endocytosis, regulation of actin cytoskeleton, and TGF-β signaling pathway. Functions affected: enzyme binding, cell projection, transcription factor activity, regulation of gene expression, and cell metabolism. Mir-130a03p had the highest differential expression and was predicted to bind to SIRT7. The downregulation of SIRT7 may enhance the osteogenic differentiation of BMSCs. | [34] |
| NA | Phosphorylation of STAT6, GSK-3α/β, STAT5b, and STAT5a/b increased following stimulation with 0d-Exo, whereas phosphorylation levels of FAK, PRAS40, and WNK1 were downregulated. SMAD 4 and BMPR2 were upregulated in 0d-Exo treatment. SMAD1/8, BMPR1A, and BMPRIB showed no difference between treatments. | [35] |
| D21 versus D0 (do not specify the type of cell): ↑ of miR-186, miR-210, miR-181c-5p, and miR-146a-5p and ↓ of miR-133 and miR-485. | Potential signaling pathways: TGF-beta signaling pathway, Hippo signaling pathway, Map kinase, and Wnt signaling pathway, among others. | [32] |
| NA | ↓ PTEN in DMOG-MSC-Exos. The deficiency of PTEN is related to increased migration and invasion of HUVECs. The downstream target of PTEN, AKT/mTOR, was blocked, and DMOG-MSC-Exos lost their superior pro-angiogenic abilities. | [20] |
| ↑ of miR-146a in BMSCs by Sr-CS extracts | The miR-146a inhibition led to the downregulation of miR-146a in both BMSCs and BMSC-Exos. Treatment with Sr-CS + 146I-Exo (derived from miR-146a inhibition) diminished its angiogenic ability in HUVECs. Prediction targets showed that miR-146a directly targets Smad4 and NF2 (reported to inhibit angiogenesis). | [41] |
| ↓ of IGFBP3 associated with miR-375 overexpression. ↑ cell differentiation in IGFBP3-deficient cells cultured in OM. Furthermore, the increase in osteogenic differentiation induced by Exo (miR-375) treatment was reversed by IGFBP3 recombinant treatment. | [38] | |
| NA | BMP2 FEEs were able to trigger SMAD 1/5/8 phosphorylation, and control EVs had no effect beyond the resting cell state. | [21] |
| ↑ of 94 miRNAs and ↓ of 39 miRNAs in the Hypo-Exos group when compared to the Exos group. ↑ of miR-126, miR-855–5p, miR-146b, miR-223, and miR-451. | Knockdown of miR-126 inhibited Hypo-Exos-mediated proliferation, migration, and angiogenesis in vitro and in vivo. SPRED1 is a target of miR-126 and was increased in miR-126 knockdown Hypo-Exos-treated cells. Silencing of SPRED1 promoted cell proliferation, migration, and tube formation during treatments with miR-126 knockdown Hypo-Exos. Hypo-Exos suppressed SPRED1 by activating the Ras/Erk pathway. | [17] |
| 565 miRNAs were differentially expressed in SM-Exo. ↑ of 16 miRNAs and ↓ of 9 miRNAs in SM-Exo compared to Exos. ↑ of mir-10a-5p and mir-10b-5p and ↓ of mir-212-3p in SM-Exo. | NA | [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hertel, F.C.; Silva, A.S.d.; Sabino, A.d.P.; Valente, F.L.; Reis, E.C.C. Preconditioning Methods to Improve Mesenchymal Stromal Cell-Derived Extracellular Vesicles in Bone Regeneration—A Systematic Review. Biology 2022, 11, 733. https://doi.org/10.3390/biology11050733
Hertel FC, Silva ASd, Sabino AdP, Valente FL, Reis ECC. Preconditioning Methods to Improve Mesenchymal Stromal Cell-Derived Extracellular Vesicles in Bone Regeneration—A Systematic Review. Biology. 2022; 11(5):733. https://doi.org/10.3390/biology11050733
Chicago/Turabian StyleHertel, Fernanda Campos, Aline Silvestrini da Silva, Adriano de Paula Sabino, Fabrício Luciani Valente, and Emily Correna Carlo Reis. 2022. "Preconditioning Methods to Improve Mesenchymal Stromal Cell-Derived Extracellular Vesicles in Bone Regeneration—A Systematic Review" Biology 11, no. 5: 733. https://doi.org/10.3390/biology11050733
APA StyleHertel, F. C., Silva, A. S. d., Sabino, A. d. P., Valente, F. L., & Reis, E. C. C. (2022). Preconditioning Methods to Improve Mesenchymal Stromal Cell-Derived Extracellular Vesicles in Bone Regeneration—A Systematic Review. Biology, 11(5), 733. https://doi.org/10.3390/biology11050733