
Apothecial Ancestry, Evolution, and Re-Evolution in Thelebolales (Leotiomycetes, Fungi)

 ,
, 
 ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimens Studied and Bibliographic Review
2.2. Morphological Studies
2.3. Phylogenetic Studies
2.4. Morphological Character Evolution within Thelebolales
3. Results
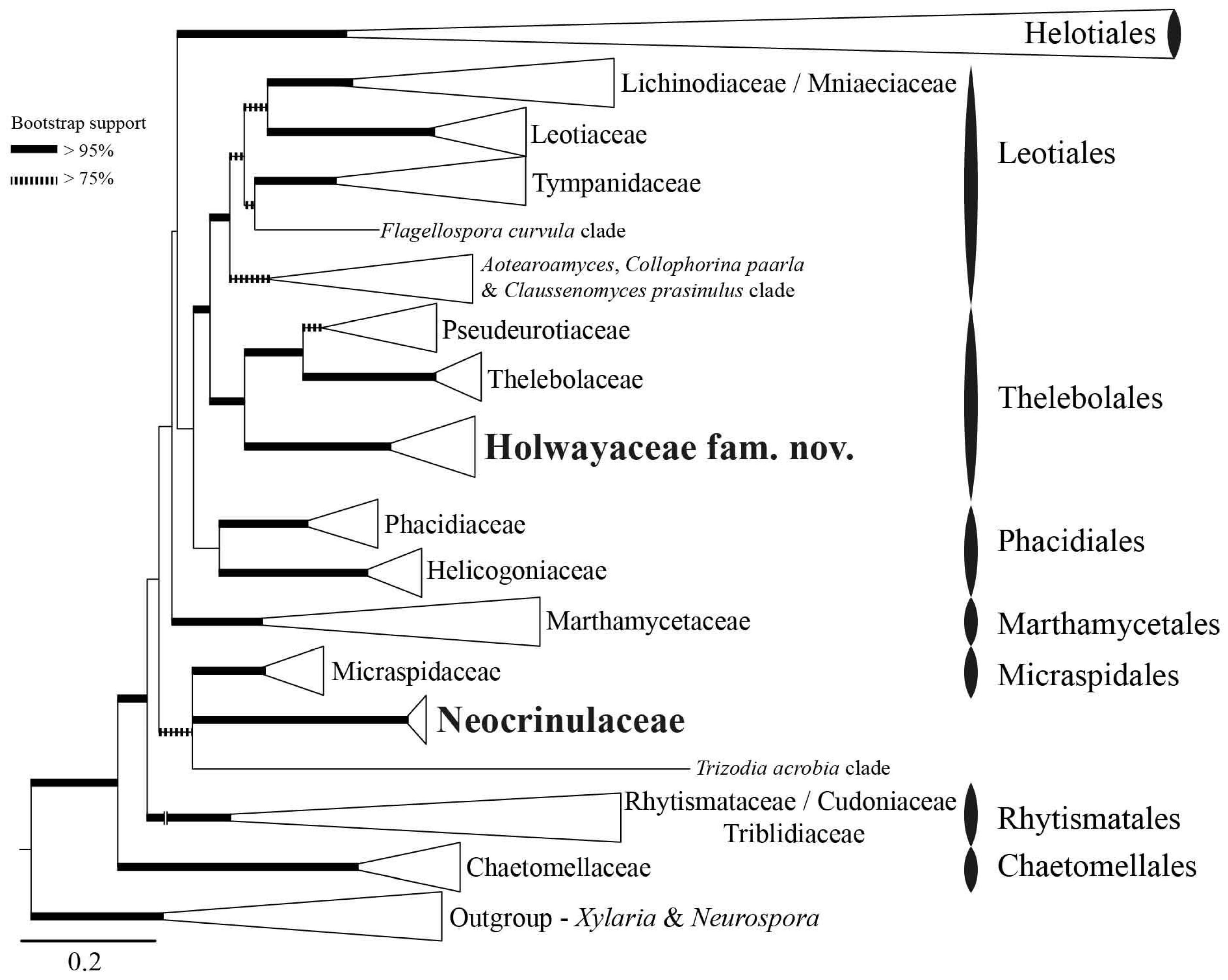
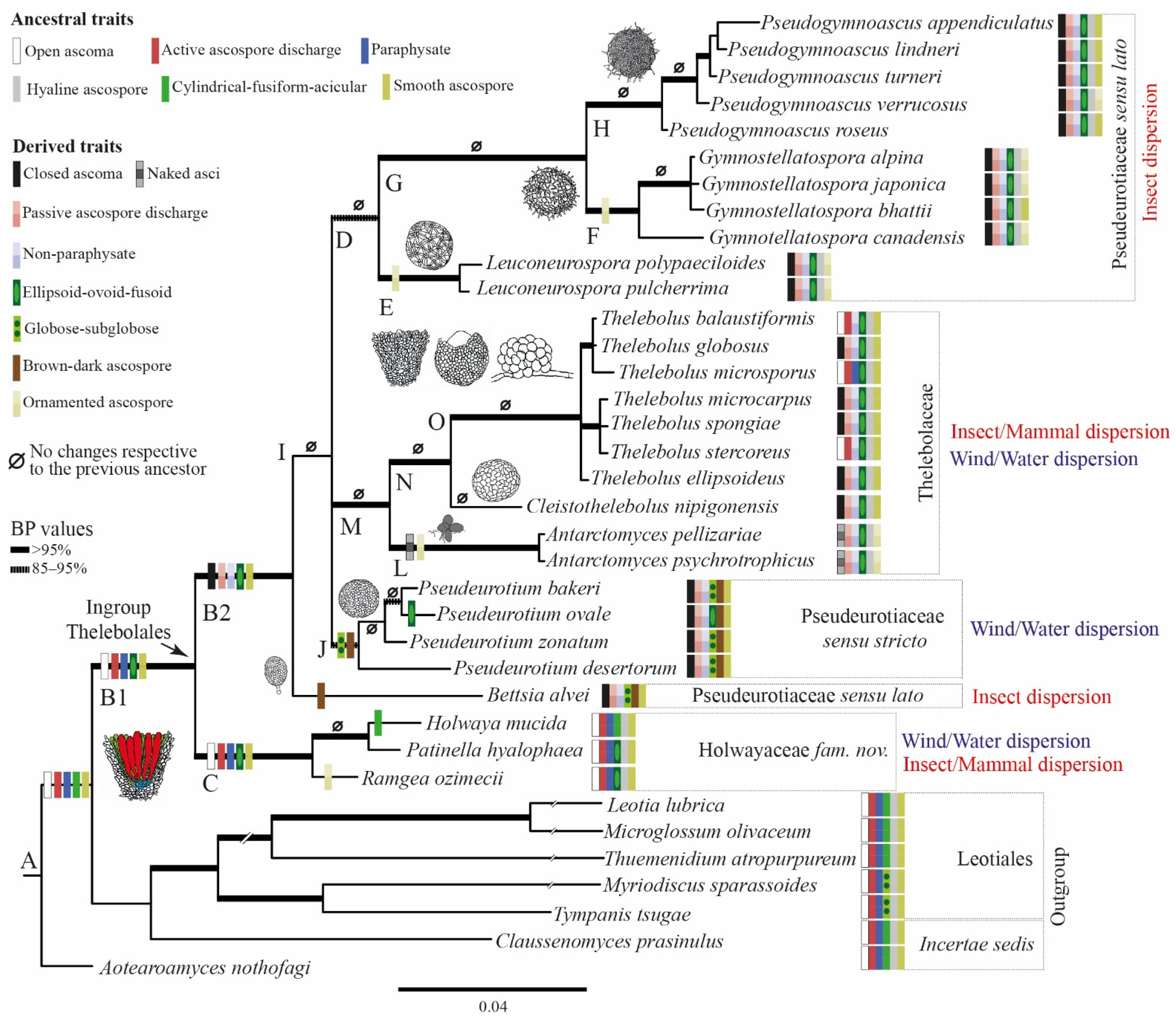
3.1. Phylogenetic Results
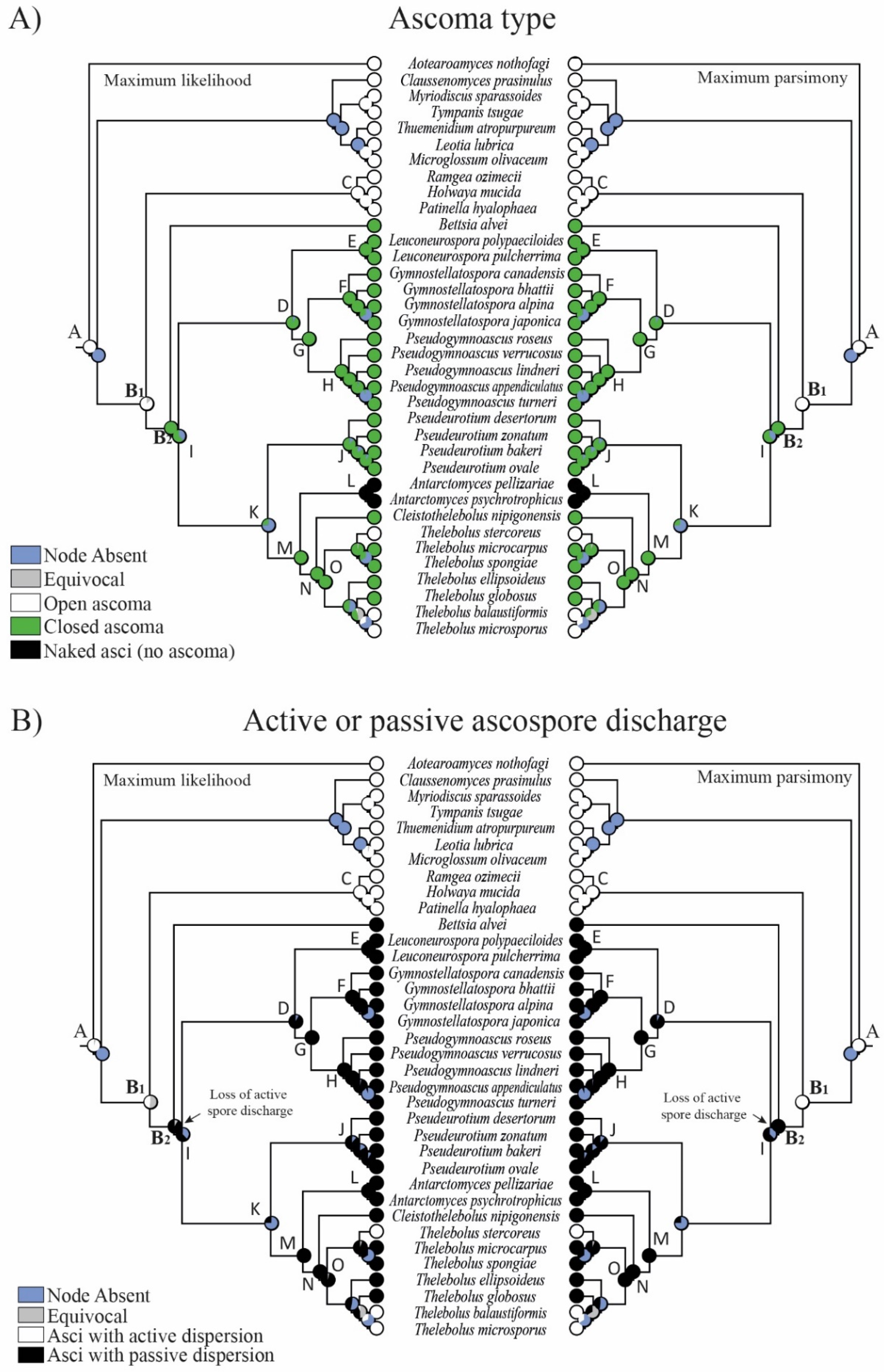
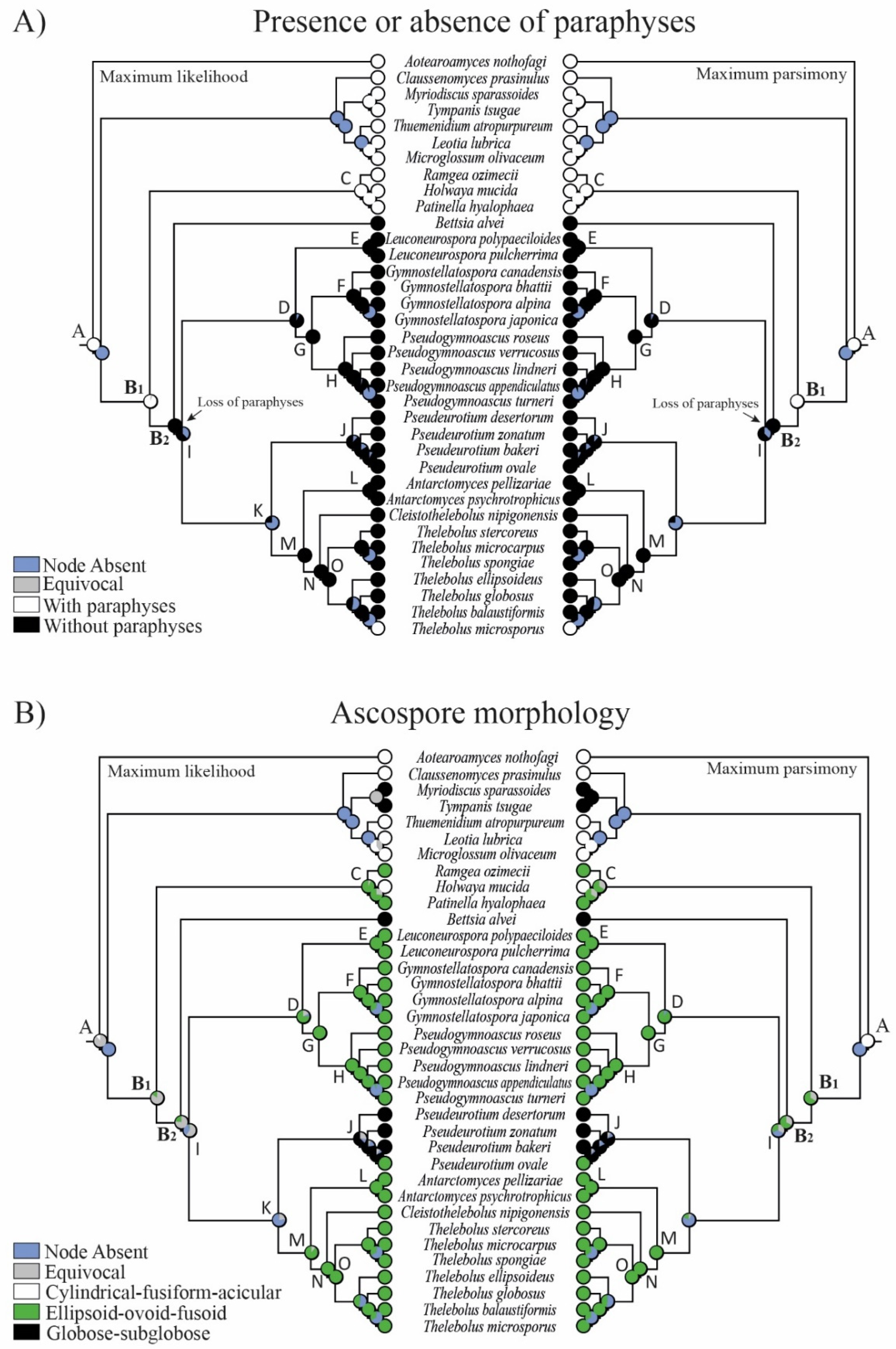
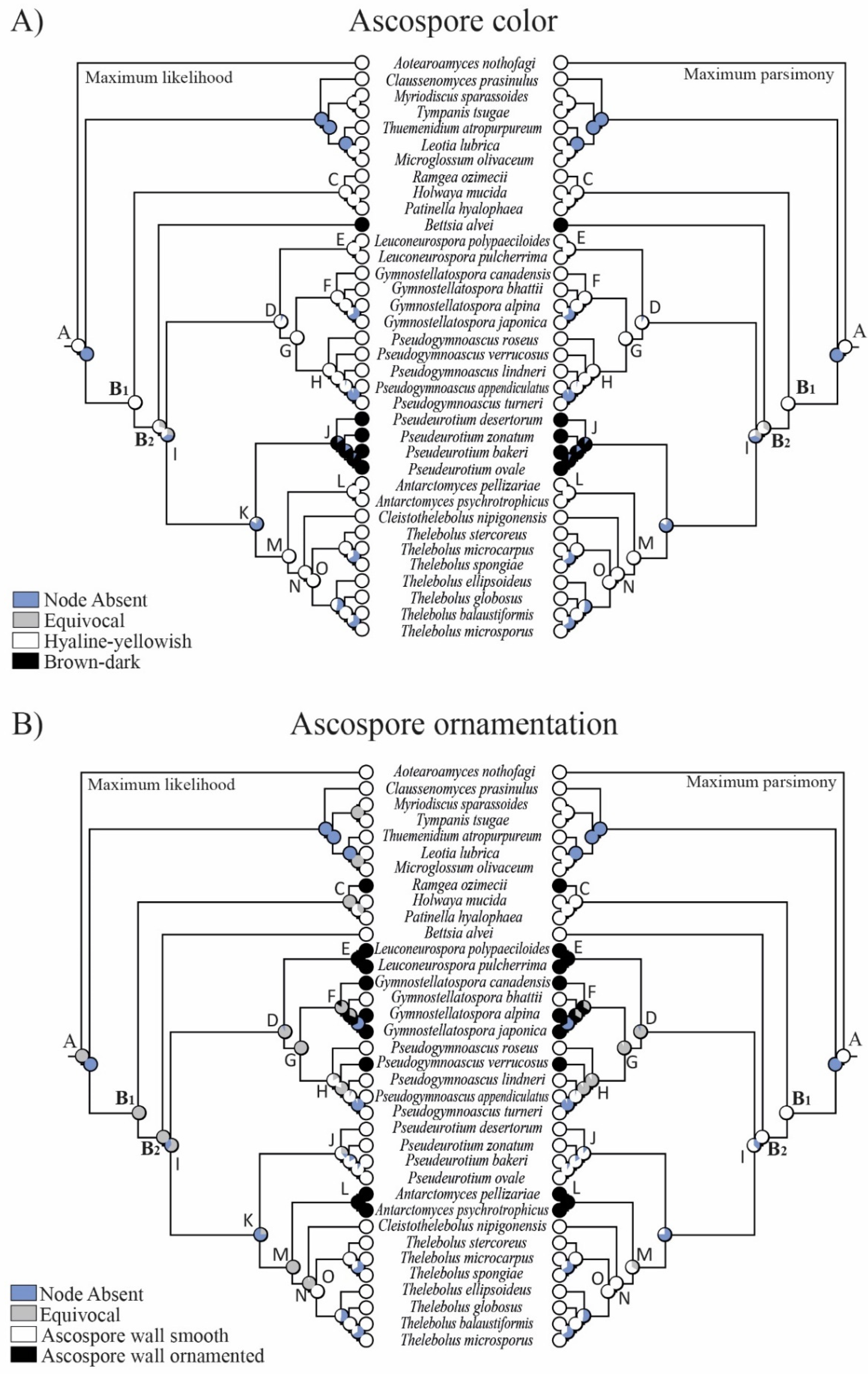
3.2. Reconstruction of Ancestral States
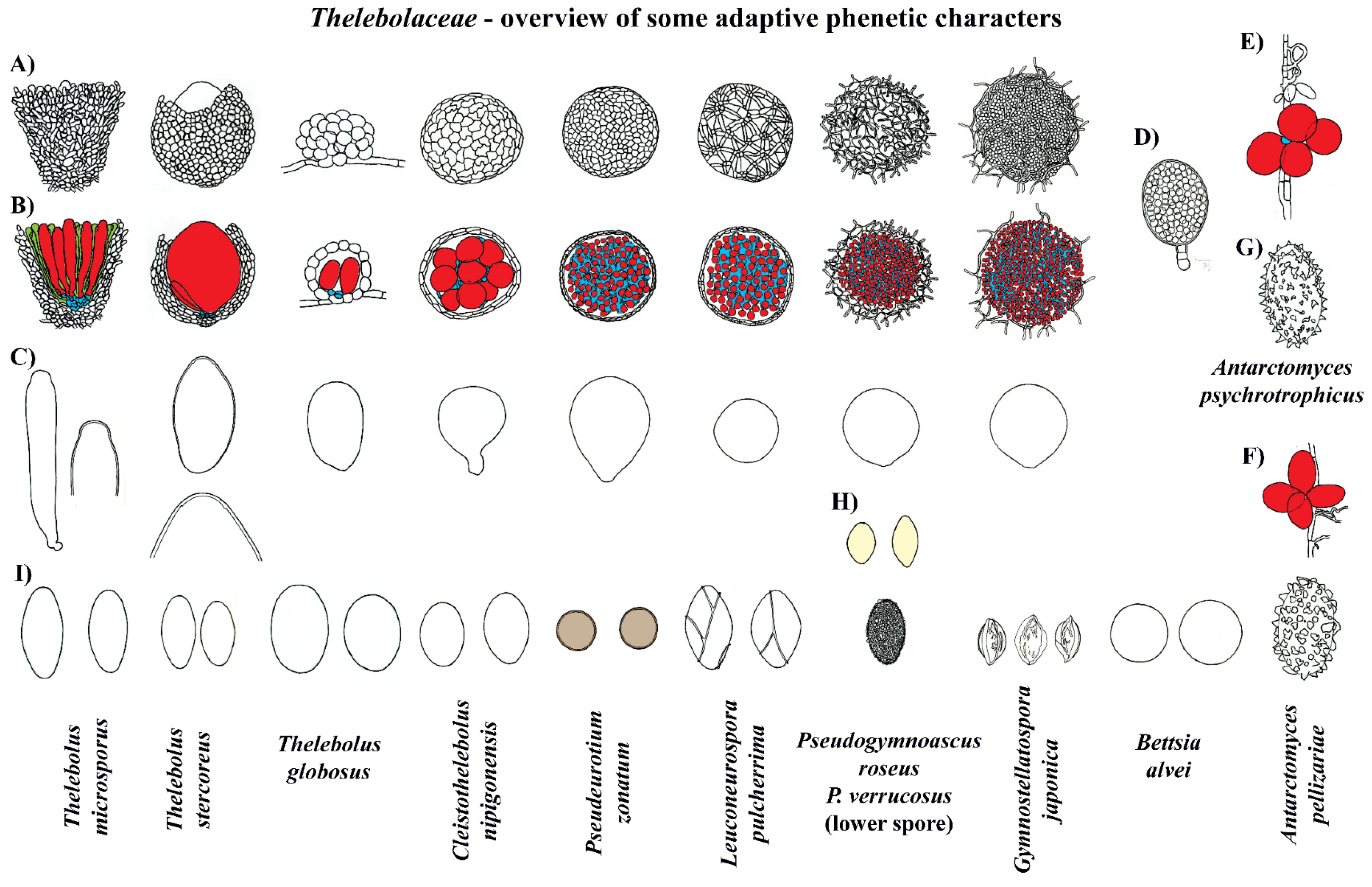
3.3. Taxonomy

4. Discussion
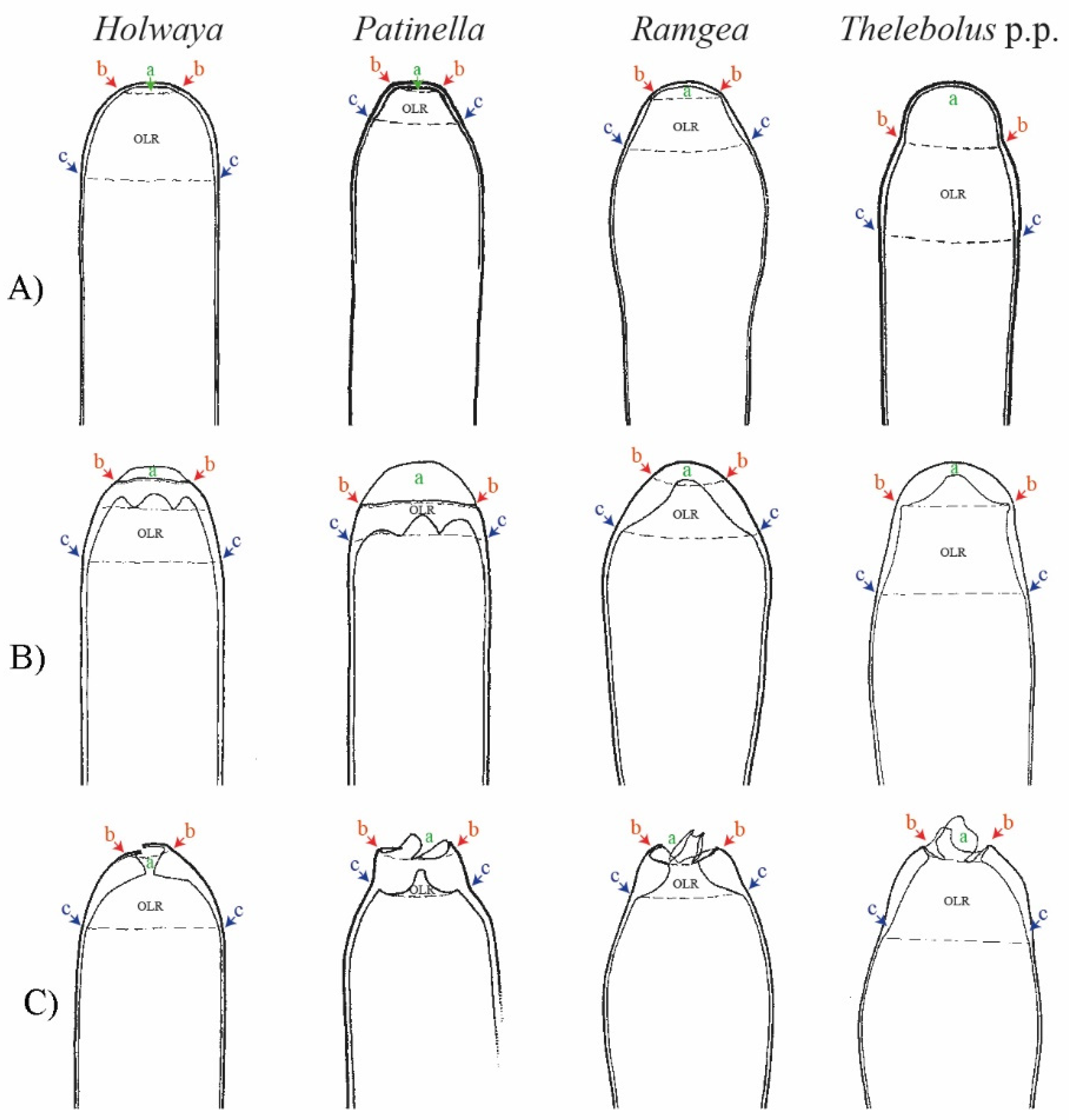
4.1. Diversity and Systematics of Holwayaceae
4.2. Ecology and Distribution in Holwayaceae
4.3. Evolution in Holwayaceae
4.4. Evolution in Thelebolales Lineages
4.5. Known Anamorphs and Conidia Dispersal
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, B.; Hussain, M.; Zhang, W.; Stadler, M.; Liu, X.; Xiang, M. Current insights into fungal species diversity and perspective on naming the environmental DNA sequences of fungi. Mycology 2019, 10, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Fry, C.; Smith, R.J.; Simmonds, M.S.J.; Kersey, P.J.; Pritchard, H.W.; Abbo, M.S.; Acedo, C.; Adams, J.; Ainsworth, A.M.; et al. State of the World’s Plants and Fungi 2020; Royal Botanic Gardens, Kew: London, UK, 2020; pp. 1–96. [Google Scholar] [CrossRef]
- Hawksworth, D.L. The magnitude of fungal diversity: The 1.5 million species estimate revisited. Mycol. Res. 2001, 105, 1422–1432. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Ohman, A.; Glotzer, D.; Nuhn, M.; Kirk, P.; Nilsson, R.H. Progress in molecular and morphological taxon discovery in Fungi and options for formal classification of environmental sequences. Fungal Biol. Rev. 2011, 25, 38–47. [Google Scholar] [CrossRef]
- Monastersky, R. Biodiversity: Life—A status report. Nature 2014, 516, 158–161. [Google Scholar] [CrossRef]
- Cannon, P.; Aguirre-Hudson, B.; Aime, M.C.; Ainsworth, A.M.; Bidartondo, M.I.; Gaya, E.; Hawksworth, D.; Kirk, P.; Leitch, I.J.; Lücking, R. Definition and diversity. In State of the World’s Fungi. Report; Willis, K.J., Ed.; Royal Botanic Gardens, Kew: London, UK, 2018; pp. 4–11. [Google Scholar]
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited: 2.2 to 3.8 million species. In The Fungal Kingdom; Heitman, J., Howlett, B.J., Crous, P.W., Stukenbrock, E.H., James, T.Y., Eds.; American Society for Microbiology: Washington, DC, USA, 2018; pp. 1–1136. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CABI: Wallingford, UK, 2008. [Google Scholar]
- Liu, Y.J.; Hall, B.D. Body plan evolution of ascomycetes, as inferred from an RNA polymerase II phylogeny. Proc. Natl. Acad. Sci. USA 2004, 101, 4507–4512. [Google Scholar] [CrossRef]
- Taylor, T.N.; Krings, M.; Taylor, E.L. Fossil Fungi; Academic Press: London, UK, 2015; pp. 1–384. [Google Scholar]
- Schmitt, I.; Prado, R.; Grube, M.; Lumbsch, H.T. Repeated evolution of closed fruiting bodies is linked to ascoma development in the largest group of lichenized fungi (Lecanoromycetes, Ascomycota). Mol. Phylogenet. Evol. 2009, 52, 34–44. [Google Scholar] [CrossRef]
- Berbee, M.L.; Taylor, J.W. Two ascomycete classes based on fruiting-body characters and ribosomal DNA sequence. Mol. Biol. Evol. 1992, 9, 278. [Google Scholar]
- Schoch, C.L.; Sung, G.H.; López-Giráldez, F.; Townsend, J.P.; Miadlikowska, J.; Hofstetter, V.; Robbertse, B.; Matheny, P.B.; Kauff, F.; Wang, Z.; et al. The Ascomycota tree of life: A phylum-wide phylogeny clarifies the origin and evolution of fundamental reproductive and ecological traits. Syst. Biol. 2009, 58, 224–239. [Google Scholar] [CrossRef] [PubMed]
- Johnston, P.R.; Quijada, L.; Smith, C.A.; Baral, H.O.; Hosoya, T.; Baschien, C.; Pärtel, K.; Zhuang, W.Y.; Haelewaters, D.; Park, D.; et al. A multigene phylogeny toward a new phylogenetic classification of Leotiomycetes. IMA Fungus 2019, 10, 1–22. [Google Scholar] [CrossRef]
- Greif, M.D.; Currah, R.S. A functional interpretation of the role of the reticuloperidium in whole-ascoma dispersal by arthropods. Mycol. Res. 2003, 107, 77–81. [Google Scholar] [CrossRef]
- Trail, F. Fungal cannons: Explosive spore discharge in the Ascomycota. FEMS Microbiol. Lett. 2007, 276, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Wynns, A.A. Convergent evolution of highly reduced fruiting bodies in Pezizomycotina suggests key adaptations to the bee habitat. BMC Evol. Biol. 2015, 15, 145. [Google Scholar] [CrossRef] [PubMed]
- Calhim, S.; Halme, P.; Petersen, J.H.; Læssøe, T.; Bässler, C.; Heilmann-Clausen, J. Fungal spore diversity reflects substrate-specific deposition challenges. Sci. Rep. 2018, 8, 5356. [Google Scholar] [CrossRef] [PubMed]
- Ingold, C.T. Fungal Spores: Their Liberation and Dispersal; Oxford University Press: New York, NY, USA, 1971; pp. 1–302. [Google Scholar]
- Roper, M.; Pepper, R.E.; Brenner, M.P.; Pringle, A. Explosively launched spores of ascomycete fungi have drag-minimizing shapes. Proc. Natl. Acad. Sci. USA 2008, 105, 20583–20588. [Google Scholar] [CrossRef]
- Pringle, A.; Brenner, M.P.; Fritz, J.A.; Roper, M.; Seminara, A. Reaching the wind: Boundary layer escape as a constraint on ascomycete spore dispersal. In The Fungal Community: Its Organization and Role in the Ecosystem, 4th ed.; Dighton, J., White, J.F., Eds.; CRC Press: Boca Raton, FL, USA; London, UK; New York, NY, USA, 2017; pp. 309–320. [Google Scholar]
- Fritz, J.A.; Seminara, A.; Roper, M.; Pringle, A.; Brenner, M.P. A natural O-ring optimizes the dispersal of fungal spores. J. R. Soc. Interface 2013, 10, 20130187. [Google Scholar] [CrossRef] [PubMed]
- van Brummelen, J. A world-monograph of the genera Ascobolus and Saccobolus (Ascomycetes, Pezizales). Persoonia 1967, 1, 1–260. [Google Scholar]
- Plishka, M.J.R.; Tsuneda, A.; Currah, R.S. Evidence of apothecial ancestry in the cleistothecial ascomata of Pleuroascus nicholsonii. Mycol. Res. 2008, 112, 1319–1326. [Google Scholar] [CrossRef]
- Jaklitsch, W.; Baral, H.-O.; Lücking, R.; Lumbsch, H.T. Ascomycota. In Syllabus of Plant Families-Adolf Engler’s Syllabus der Pflanzenfamilien. Part 1/2 Ascomycota, 13th ed.; Frey, W., Ed.; Borntraeger Science Publishers: Stuttgart, Germany, 2016; pp. 1–322. [Google Scholar]
- Baral, H.O.; Weber, E.; Marson, G. Monograph of Orbiliomycetes (Ascomycota) based on vital taxonomy; National Museum of Natural History: Luxembourg, 2020; Volume Part I and II, pp. 1–1752. [Google Scholar]
- Wyatt, T.T.; Wösten, H.A.B.; Dijksterhuis, J. Fungal spores for dispersion in space and time. Adv. Appl. Microbiol. 2013, 85, 43–91. [Google Scholar]
- Magyar, D.; Vass, M.; Li, D.W. Dispersal strategies of microfungi. In Biology of Microfungi; Li, D.W., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 315–371. [Google Scholar]
- Norros, V.; Rannik, Ü.; Hussein, T.; Petäjä, T.; Vesala, T.; Ovaskainen, O. Do small spores disperse further than large spores? Ecology 2014, 95, 1612–1621. [Google Scholar] [CrossRef]
- Golan, J.J.; Pringle, A. Long-distance dispersal of fungi. Microbiol. Spectr. 2017, 5, 1–24. [Google Scholar] [CrossRef]
- Buller, A.H.R. Researches on Fungi. An Account of the Production, Liberation and Dispersion of the Spores of Hymenomycetes Treated Botanically and Physically. Also Some Observations upon the Discharge and Dispersion of the Spores of Ascomycetes and Pilobolus; Longmans, Green and Co.: London, UK; New York, NY, USA, 1909. [Google Scholar]
- Landvik, S.; Schumacher, T.K.; Eriksson, O.E.; Moss, S.T. Morphology and ultrastructure of Neolecta species. Mycol. Res. 2003, 107, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Norros, V.; Karhu, E.; Nordén, J.; Vähätalo, A.V.; Ovaskainen, O. Spore sensitivity to sunlight and freezing can restrict dispersal in wood-decay fungi. Ecol. Evol. 2015, 5, 3312–3326. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, C.A.; Griffin, D.W. Aerobiology and the global transport of desert dust. Trends Ecol. Evol. 2006, 21, 638–644. [Google Scholar] [CrossRef]
- Pringle, A.; Vellinga, E.; Peay, K. The shape of fungal ecology: Does spore morphology give clues to a species’ niche? Fungal Ecol. 2015, 17, 213–216. [Google Scholar] [CrossRef]
- Zanatta, F.; Patiño, J.; Lebeau, F.; Massinon, M.; Hylander, K.; Hann, M.; Ballings, P.; Degreef, J.; Vanderpoorten, A. Measuring spore settling velocity for an improved assessment of dispersal rates in mosses. Ann. Bot. 2016, 118, 197–206. [Google Scholar] [CrossRef]
- Durrell, L.W. The composition and structure of walls of dark fungus spores. Mycopathol. Mycol. Appl. 1964, 23, 339–345. [Google Scholar] [CrossRef]
- Deveautour, C.; Chieppa, J.; Nielsen, U.N.; Boer, M.M.; Mitchell, C.; Horn, S.; Power, S.A.; Guillen, A.; Bennet, A.; Powell, J.R. Biogeography of arbuscular mycorrhizal fungal spore traits along an aridity gradient, and responses to experimental rainfall manipulation. Fungal Ecol. 2020, 46, 100899. [Google Scholar] [CrossRef]
- Kawamura, C.; Tsujimoto, T.; Tsuge, T. Targeted disruption of a melanin biosynthesis gene affects conidial development and UV tolerance in the Japanese pear pathotype of Alternaria alternata. Mol. Plant-Microbe Interact. 1999, 12, 59–63. [Google Scholar] [CrossRef]
- Robinson, C.H. Cold adaptation in Arctic and Antarctic fungi. New Phytol. 2001, 151, 341–353. [Google Scholar] [CrossRef]
- Gessler, N.N.; Egorova, A.S.; Belozerskaya, T.A. Melanin pigments of fungi under extreme environmental conditions. Appl. Biochem. Microbiol. 2014, 50, 105–113. [Google Scholar] [CrossRef]
- Kejžar, A.; Gobec, S.; Plemenitaš, A.; Lenassi, M. Melanin is crucial for growth of the black yeast Hortaea werneckii in its natural hypersaline environment. Fungal Biol. 2013, 117, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Fogarty, R.V.; Tobin, J.M. Fungal melanins and their interactions with metals. Enzyme Microb. Technol. 1996, 19, 311–317. [Google Scholar] [CrossRef]
- Bloomfield, B.J.; Alexander, M. Melanins and resistance of fungi to lysis. J. Bacteriol. 1967, 93, 1276–1280. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.J.; Alexander, M. Inhibition of the lysis of fungi by melanins. J. Bacteriol. 1967, 94, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Henson, J.M.; Butler, M.J.; Day, A.W. The dark side of the mycelium: Melanins of phytopathogenic fungi. Annu. Rev. Phytopathol. 1999, 37, 447–471. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Casadevall, A. The contribution of melanin to microbial pathogenesis. Cell. Microbiol. 2003, 5, 203–223. [Google Scholar] [CrossRef]
- Berbee, M.L. The phylogeny of plant and animal pathogens in the Ascomycota. Physiol. Mol. Plant Pathol. 2001, 59, 165–187. [Google Scholar] [CrossRef]
- Mueller, G.M.; Foster, M.S.; Bills, G.F. Biodiversity of Fungi: Inventory and Monitoring Methods; Elsevier Academic Press: Burlington, MA, USA, 2004; pp. 1–777. [Google Scholar]
- O’Brien, H.E.; Parrent, J.L.; Jackson, J.A.; Moncalvo, J.M.; Vilgalys, R. Fungal community analysis by large-scale sequencing of environmental samples. Appl. Environ. Microbiol. 2005, 71, 5544–5550. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Sung, G.-H.; Johnson, D.; Hesse, C.; O’Rourke, B.; Serdani, M.; Spotts, R.; Lutzoni, F.; Hofstetter, V.; Miadlikowska, J.; et al. A five-gene phylogeny of Pezizomycotina. Mycologia 2006, 98, 1018–1028. [Google Scholar] [CrossRef]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Prieto, M.; Schultz, M.; Olariaga, I.; Wedin, M. Lichinodium is a new lichenized lineage in the Leotiomycetes. Fungal Divers. 2019, 94, 23–39. [Google Scholar] [CrossRef]
- Suh, S.O.; Blackwell, M. Molecular phylogeny of the cleistothecial fungi placed in Cephalothecaceae and Pseudeurotiaceae. Mycologia 1999, 91, 836–848. [Google Scholar] [CrossRef]
- Gernandt, D.S.; Platt, J.L.; Stone, J.K.; Spatafora, J.W.; Holst-Jensen, A.; Hamelin, R.C.; Kohn, L. Phylogenetics of Helotiales and Rhytismatales based on partial small subunit nuclear ribosomal DNA sequences. Mycologia 2001, 93, 915–933. [Google Scholar] [CrossRef]
- Stajich, J.E.; Berbee, M.L.; Blackwell, M.; Hibbett, D.S.; James, T.Y.; Spatafora, J.W.; Taylor, J.W. The Fungi. Curr. Biol. 2009, 19, R840–R845. [Google Scholar] [CrossRef] [PubMed]
- Untereiner, W.; Yue, Q.; Chen, L.; Li, Y.; Bills, G.; Štěpánek, V.; Réblová, M. Phialophora section Catenulatae disassembled: New genera, species, and combinations and a new family encompassing taxa with cleistothecial ascomata and phialidic asexual states. Mycologia 2019, 111, 998–1027. [Google Scholar] [CrossRef]
- Rodríguez-Andrade, E.; Stchigel, A.M.; Terrab, A.; Guarro, J.; Cano-Lira, J.F. Diversity of xerotolerant and xerophilic fungi in honey. IMA Fungus 2019, 10, 20. [Google Scholar] [CrossRef]
- Haelewaters, D.; Peterson, R.A.; Nevalainen, H.; Aime, M.C. Inopinatum lactosum gen. & comb. nov., the first yeast-like fungus in Leotiomycetes. Int. J. Syst. Evol. Microbiol. 2021, 71, 1–7. [Google Scholar]
- Hoog, G.S.; Göttlich, E.; Platas, G.; Genilloud, O.; Leotta, G.; Brummelen, J. van Evolution, taxonomy and ecology of the genus Thelebolus in Antarctica. Stud. Mycol. 2005, 51, 33–76. [Google Scholar]
- Wang, Z.; Binder, M.; Schoch, C.L.; Johnston, P.R.; Spatafora, J.W.; Hibbett, D.S. Evolution of helotialean fungi (Leotiomycetes, Pezizomycotina): A nuclear rDNA phylogeny. Mol. Phylogenet. Evol. 2006, 41, 295–312. [Google Scholar] [CrossRef]
- LoBuglio, K.F.; Pfister, D.H. Placement of Medeolaria farlowii in the Leotiomycetes, and comments on sampling within the class. Mycol. Prog. 2010, 9, 361–368. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.S.J.; Crane, C.; Barrett, S.; Cano-Lira, J.F.; Le Roux, J.J.; Thangavel, R.; Guarro, J.; et al. Fungal Planet description sheets: 469–557. Persoonia 2016, 37, 218–403. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Norphanphoun, C.; Abreu, V.P.; Bazzicalupo, A.; Chethana, K.W.T.; Clericuzio, M.; Dayarathne, M.C.; Dissanayake, A.J.; Ekanayaka, A.H.; He, M.Q.; et al. Fungal diversity notes 603–708: Taxonomic and phylogenetic notes on genera and species. Fungal Divers. 2017, 87, 1–235. [Google Scholar] [CrossRef]
- Brummelen, J. van Reconsideration of relationships within the Thelebolaceae based on ascus ultrastructure. Persoonia 1998, 16, 119–125. [Google Scholar]
- Kirschner, R. Sex does not sell: The argument for using the terms “anamorph” and “teleomorph” for fungi. Mycol. Prog. 2018, 18, 305–312. [Google Scholar] [CrossRef]
- Baral, H.O. Vital versus herbarium taxonomy: Morphological differences between living and dead cells of ascomycetes, and their taxonomic implications. Mycotaxon 1992, 44, 333–390. [Google Scholar]
- Quijada, L. Estudio de los órdenes Helotiales s.l. y Orbiliales (Ascomycota, Fungi) en la Isla de Tenerife. Ph.D. Thesis, Departamento de Botánica, Ecología y Fisiología Vegetal, Universidad de La Laguna, La Laguna, Spain, 18 September 2015. [Google Scholar]
- Kušan, I. Contribution of “Vital taxonomy” Methods to the Ascomycota Taxonomy. Ph.D. Thesis, Josip Juraj Strossmayer University of Osijek, Osijek, Croatia, 25 September 2015. [Google Scholar]
- Karakehian, J.M.; Quijada, L.; Friebes, G.; Tanney, J.B.; Pfister, D.H. Placement of Triblidiaceae in Rhytismatales and comments on unique ascospore morphologies in Leotiomycetes (Fungi, Ascomycota). MycoKeys 2019, 54, 99–133. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. ISCC-NBS Color-Name Charts Illustrated with Centroid Colors. Inter-Society Color Council; National Bureau of Standards: Washington, DC, USA, 1976. [Google Scholar]
- Quijada, L.; Tanney, J.B.; Popov, E.; Johnston, P.R.; Pfister, D.H. Cones, needles and wood: Micraspis (Micraspidaceae, Micraspidales fam. et ord. nov.) speciation segregates by host plant tissues. Fungal Syst. Evol. 2020, 5, 99–111. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; St.J. Hardy, G.E.; Smith, D.; Summerell, B.A.; Cano-Lira, J.F.; Guarro, J.; Houbraken, J.; et al. Fungal Planet description sheets: 625–715. Persoonia 2017, 39, 270–467. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.70. 2021. Available online: http://www.mesquiteproject.org (accessed on 12 August 2021).
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Batista, T.M.; Hilario, H.O.; De Brito, G.A.M.; Moreira, R.G.; Furtado, C.; De Menezes, G.C.A.; Rosa, C.A.; Rosa, L.H.; Franco, G.R. Whole-genome sequencing of the endemic Antarctic fungus Antarctomyces pellizariae reveals an ice-binding protein, a scarce set of secondary metabolites gene clusters and provides insights on Thelebolales phylogeny. Genomics 2020, 112, 2915–2921. [Google Scholar] [CrossRef]
- Seifert, K.A. A monograph of Stilbella and some allied Hyphomycetes. Stud. Mycol. 1985, 27, 1–225. [Google Scholar]
- Baral, H.O.; Carter, A. Patinella hyalophaea Sacc.–Rediscovered in New Brunswick, Canada. Ascomycete 2013, 5, 91–96. [Google Scholar]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungi-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Zhang, Z.; Dong, C.; Chen, W.; Mou, Q.; Lu, X.; Han, Y.; Huang, J.; Liang, Z. The enigmatic Thelebolaceae (Thelebolales, Leotiomycetes): One new genus Solomyces and five new species. Front. Microbiol. 2020, 11, 572596. [Google Scholar] [CrossRef]
- Korf, R.P.; Abawi, G.S. On Holwaya, Crinula, Claussenomyces, and Corynella. Can. J. Bot. 1971, 49, 1879–1883. [Google Scholar] [CrossRef]
- Krieglsteiner, G.J.; Häffner, J. Über Holwaya mucida (S. Schulzer von Müggenburg 1860) R.P. Korf et G.S. Abawi 1971, subspec. mucida Korf et Abawi 1971 und ihr Vorkommen in Europa. Z. Mykol. 1985, 51, 131–138. [Google Scholar]
- Aronsson, G. Lindskål, Holwaya mucida, i Sverige. Sven. Bot. Tidskr. 1991, 85, 9–18. [Google Scholar]
- Brummelen, J. van Ramgea, a new genus of Pezizales from the Netherlands. Persoonia 1992, 14, 577–582. [Google Scholar]
- GBIF–Global Biodiversity Information Facility. Available online: https://www.gbif.org (accessed on 25 December 2021).
- Matočec, N.; Jukić, N.; Omerović, N.; Kušan, I. Dinaric karst poljes and their importance for mycobiota. In Dinaric Karst Poljes-Nature Conservation and Rural Development; Sackl, P., Ferger, S., Sarajlić, N., Kotrošan, D., Topić, G., Eds.; Ornitološko društvo “Naše ptice”: Sarajevo, Bosnia and Herzegovina, 2019; pp. 27–49. [Google Scholar]
- Held, B.W.; Blanchette, R.A. Deception Island, Antarctica, harbors a diverse assemblage of wood decay fungi. Fungal Biol. 2017, 121, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Ogaki, M.B.; Vieira, R.; Muniz, M.C.; Zani, C.L.; Alves, T.M.; Junior, P.A.; Murta, S.M.; Barbosa, E.C.; Oliveira, J.G.; Ceravolo, I.P.; et al. Diversity, ecology, and bioprospecting of culturable fungi in lakes impacted by anthropogenic activities in Maritime Antarctica. Extremophiles 2020, 24, 637–655. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.B.; Matthiesen, H.; Blanchette, R.A.; Alfredsen, G.; Held, B.W.; Westergaard-Nielsen, A.; Hollesen, J. Fungal attack on archaeological wooden artefacts in the Arctic—Implications in a changing climate. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Bässler, C.; Halbwachs, H.; Karasch, P.; Holzer, H.; Gminder, A.; Krieglsteiner, L.; Gonzalez, R.S.; Müller, J.; Brandl, R. Mean reproductive traits of fungal assemblages are correlated with resource availability. Ecol. Evol. 2016, 6, 582–592. [Google Scholar] [CrossRef]
- Fischer, M.W.; Money, N.P. Why mushrooms form gills: Efficiency of the lamellate morphology. Fungal Biol. 2010, 114, 57–63. [Google Scholar] [CrossRef][Green Version]
- Bässler, C.; Heilmann-Clausen, J.; Karasch, P.; Brandl, R.; Halbwachs, H. Ectomycorrhizal fungi have larger fruit bodies than saprotrophic fungi. Fungal Ecol. 2015, 17, 205–212. [Google Scholar] [CrossRef]
- Halbwachs, H.; Bässler, C. No bull: Dung-dwelling mushrooms show reproductive trait syndromes different from their non-coprophilous allies. Mycol. Prog. 2020, 19, 817–824. [Google Scholar] [CrossRef]
- Moore, D.; Gange, A.C.; Gange, E.G.; Boddy, L. Fruit bodies: Their production and development in relation to environment. In Ecology of Saprotrophic Basidiomycetes; Boddy, L., Frankland, J.C., van West, P., Eds.; Mycological Society Symposia Series; Elsevier Ltd.: London, UK, 2008; Volume 28, pp. 79–103. [Google Scholar]
- Wang, Z.; Johnston, P.R.; Yang, Z.L.; Townsend, J.P. Evolution of reproductive morphology in leaf endophytes. PLoS ONE 2009, 4, e4246. [Google Scholar] [CrossRef] [PubMed]
- Tanney, J.B.; Seifert, K.A. Phacidiaceae endophytes of Picea rubens in Eastern Canada. Botany 2018, 96, 555–588. [Google Scholar] [CrossRef]
- Greif, M.D.; Tsuneda, A.; Currah, R.S. The peridial development and dehiscence mechanism of Cryptendoxyla hypophloia, a cleistothecial ascomycete isolated from the bodies of arthropods. Int. J. Plant Sci. 2004, 165, 957–964. [Google Scholar] [CrossRef]
- Sherwood, M. Convergent evolution in discomycetes from bark and wood. Bot. J. Linn. Soc. 1981, 82, 15–34. [Google Scholar] [CrossRef]
- Rehnstrom, A.L.; Free, S.J. The isolation and characterization of melanin-deficient mutants of Monilinia fructicola. Physiol. Mol. Plant Pathol. 1996, 49, 321–330. [Google Scholar] [CrossRef]
- Wicklow, D.T.; Malloch, D. Studies in the genus Thelebolus: Temperature optima for growth and ascocarp development. Mycologia 1971, 63, 118–131. [Google Scholar] [CrossRef]
- Czymmek, K.; Klomparens, K. The ultrastructure of ascosporogenesis in freeze-substituted Thelebolus crustaceus: Enveloping membrane system and ascospore initial development. Canad. J. Bot. 1992, 70, 1669–1683. [Google Scholar] [CrossRef]
- Torruella, G.; De Mendoza, A.; Grau-Bové, X.; Antó, M.; Chaplin, M.A.; Del Campo, J.; Eme, L.; Pérez-Cordón, G.; Whipps, C.M.; Nichols, K.M.; et al. Phylogenomics reveals convergent evolution of lifestyles in close relatives of animals and fungi. Curr. Biol. 2015, 25, 2404–2410. [Google Scholar] [CrossRef]
- Wicklow, D.T. The coprophilous fungal community: An experimental system. In The Fungal Community, Its Organization and Role in the Ecosystem; Carrol, G.C., Wicklow, D.T., Eds.; Marcel Dekker: New York, NY, USA, 1992; pp. 715–728. [Google Scholar]
- Garnica, S.; Weiss, M.; Walther, G.; Oberwinkler, F. Reconstructing the evolution of agarics from nuclear gene sequences and basidiospore ultrastructure. Mycol. Res. 2007, 111, 1019–1029. [Google Scholar] [CrossRef]
- Halbwachs, H.; Brandl, R.; Bässler, C. Spore wall traits of ectomycorrhizal and saprotrophic agarics may mirror their distinct lifestyles. Fungal Ecol. 2015, 17, 197–204. [Google Scholar] [CrossRef]
- Malloch, D.; Cain, R.F. Four new genera of cleistothecial Ascomycetes with hyaline ascospores. Can. J. Bot. 1971, 49, 847–854. [Google Scholar] [CrossRef]
- Doveri, F.; Sarrocco, S.; Vannacci, G. Studies on three rare coprophilous plectomycetes from Italy. Mycotaxon 2013, 124, 279–300. [Google Scholar] [CrossRef]
- Pivkin, M.; Aleshko, S.; Krasokhin, V.; Khudyakova, Y. Fungal assemblages associated with sponges of the southern coast of Sakhalin Island. Russ. J. Mar. Biol. 2006, 32, 207–213. [Google Scholar] [CrossRef]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi, 2nd ed.; taxonomically revised by W Gams; IHW-Verlag: Eching, Germany, 2007; pp. 1–672. [Google Scholar]
- Bubnova, E. Fungal diversity in bottom sediments of the Kara Sea. Bot. Mar. 2010, 53, 595–600. [Google Scholar] [CrossRef]
- Adhikari, M.; Kim, S.; Yadav, D.; Um, Y.; Kim, H.; Lee, H.; Lee, Y. A new record of Pseudeurotium bakeri from crop field soil in Korea. Korean J. Mycol. 2016, 44, 145–149. [Google Scholar]
- Da Silva, T.H.; Saraiva Silva, D.A.; De Oliveira, F.S.; Goncalves Reynaud Schaefer, C.E.; Rosa, C.A.; Rosa, L.H. Diversity, distribution, and ecology of viable fungi in permafrost and active layer of Maritime Antarctica. Extremophiles 2020, 24, 565–576. [Google Scholar] [CrossRef]
- Bovio, E.; Garzoli, L.; Poli, A.; Prigione, V.; Firsova, D.; McCormack, G.; Varese, G. The culturable mycobiota associated with three Atlantic sponges, including two new species: Thelebolus balaustiformis and T. spongiae. Fungal Syst. Evol. 2018, 1, 141–167. [Google Scholar] [CrossRef]
- Collin, R.; Cipriani, R. Dollo’s law and the re-evolution of shell coiling. Proc. Royal Soc. B Biol. Sci. 2003, 270, 2551–2555. [Google Scholar] [CrossRef]
- Wiens, J.J. Re-evolution of lost mandibular teeth in frogs after more than 200 million years, and re-evaluating Dollo’s law. Evolution 2011, 65, 1283–1296. [Google Scholar] [CrossRef]
- Seher, T.; Ng, C.; Signor, S.; Podlaha, O.; Barmina, O.; Kopp, A. Genetic basis of a violation of Dollo’s Law: Re-evolution of rotating sex combs in Drosophila bipectinata. Genetics 2012, 192, 1465–1475. [Google Scholar] [CrossRef] [PubMed]
- Recknagel, H.; Kamenos, N.; Elmer, K. Common lizards break Dollo’s law of irreversibility: Genome-wide phylogenomics support a single origin of viviparity and re-evolution of oviparity. Mol. Phylogenet. Evol. 2018, 127, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Kleisner K Re-semblance and re-evolution: Paramorphism and semiotic co-option may explain the re-evolution of similar phenotypes. Sign Syst. Stud. 2010, 38, 378–392. [CrossRef]
- Gregory, P.H. Microbiology of the Atmosphere, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1973. [Google Scholar]
- Jennings, D.H.; Lysek, G. Fungal Biology: Understanding the Fungal Lifestyle, 2nd ed.; Bios Scientific Publishers Ltd.: Oxford, UK, 1999; pp. 1–166. [Google Scholar]
- Vanderwolf, K.J.; Malloch, D.; McAlpine, D.F. Ectomycota associated with arthropods from bat hibernacula in Eastern Canada, with particular reference to Pseudogymnoascus destructans. Insects 2016, 7, 16. [Google Scholar] [CrossRef]
- Malloch, D.; Sigler, L.; Hambleton, S.; Vanderwolf, K.; Gibas, C.; McAlpine, D. Fungi associated with hibernating bats in New Brunswick caves: The genus Leuconeurospora. Botany 2016, 94, 1171–1181. [Google Scholar] [CrossRef]
- Pitt, J.I.; Lantz, H.; Pettersson, O.V.; Leong, S.L. Xerochrysium gen. nov. and Bettsia, genera encompassing xerophilic species of Chrysosporium. IMA Fungus 2013, 4, 229–241. [Google Scholar] [CrossRef]
- Seifert, K.; Morgan-Jones, G.; Gams, W.; Kendrick, B. The Genera of Hyphomycetes; CBS Biodiversity Series, no. 9; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2011; pp. 1–997. [Google Scholar]
- Bandoni, R.J.; Koske, R.E. Monolayers and microbial dispersal. Science 1974, 183, 1079–1081. [Google Scholar] [CrossRef]
- Wicklow, D.T. Biogeography and conidial fungi. In Biology of Conidial Fungi; Cole, G.T., Kendrick, B., Eds.; Academic Press: New York, NY, USA, 1981; pp. 417–447. [Google Scholar]
- Arpin, N.; Bouillant, M.L. Light and mycosporines. In The Fungal Spore: Morphogenetic Controls; Turian, G., Hohl, H.R., Eds.; Academic Press: London, UK; New York, NY, USA; Toronto, ON, Canada, 1981; pp. 435–454. [Google Scholar]
- Matočec, N. Cave fungi. Ascomycota, Basidiomycota and Anamorphic (Mitosporic) fungi. An Overview of the Cave and Interstitial Biota of Croatia. Natura Croatica 2002, 11 (Suppl. 1), 21–27. [Google Scholar]
- Hoog, G.S.; Smith, M.T. Hyphozyma, a new genus of yeast-like Hyphomycetes. Antonie van Leeuwenhoek 1981, 47, 339–352. [Google Scholar] [CrossRef]
- Stchigel, A.M.; Cano, J.; Mac Cormack, W.; Guarro, J. Antarctomyces psychrotrophicus gen. et sp. nov., a new ascomycete from Antarctica. Mycol. Res. 2001, 105, 377–382. [Google Scholar] [CrossRef]
- Menezes, G.C.; Godinho, V.M.; Porto, B.A.; Gonçalves, V.N.; Rosa, L.H. Antarctomyces pellizariae sp. nov., a new, endemic, blue, snow resident psychrophilic ascomycete fungus from Antarctica. Extremophiles 2017, 21, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Tanney, J.B.; Quijada, L. Comments on the occurrence of yeast-like morphologies in Leotiomycetes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005141. [Google Scholar] [CrossRef] [PubMed]
- Becher, P.G.; Hagman, A.; Verschut, V.; Chakraborty, A.; Rozpędowska, E.; Lebreton, S.; Bengtsson, M.; Flick, G.; Witzgall, P.; Piškur, J. Chemical signaling and insect attraction is a conserved trait in yeasts. Ecol. Evol. 2018, 8, 2962–2974. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, P.; Vásquez, G.; Gil-Durán, C.; Oliva, V.; Díaz, A.; Henríquez, M.; Alvarez, E.; Laich, F.; Chávez, R.; Vaca, I. Description of the first four species of the genus Pseudogymnoascus from Antarctica. Front. Microbiol. 2021, 12, 713189. [Google Scholar] [CrossRef]
- Sogonov, M.V.; Schroers, H.J.; Gams, W.; Dijksterhuis, J.; Summerbell, R.C. The hyphomycete Teberdinia hygrophila gen. nov., sp. nov. and related anamorphs of Pseudeurotium species. Mycologia 2005, 97, 695–709. [Google Scholar] [CrossRef]
- Sigler, L.; Lumley, T.C.; Currah, R.S. New species and records of saprophytic ascomycetes (Myxotrichaceae) from decaying logs in the boreal forest. Mycoscience 2000, 41, 495–502. [Google Scholar] [CrossRef]
- Smith, G. Polypaecilum gen. nov. Trans. Brit. Mycol. Soc. 1961, 44, 437–440. [Google Scholar] [CrossRef]
- Von Arx, J.A. Notes on Microascaceae with the description of two new species. Persoonia 1978, 10, 23–31. [Google Scholar]
- Rice, A.V.; Currah, R.S. Oidiodendron: A survey of the named species and related anamorphs of Myxotrichum. Stud. Mycol. 2005, 53, 83–120. [Google Scholar] [CrossRef]
- Hayes, M.A. The Geomyces fungi: Ecology and distribution. BioScience 2012, 62, 819–823. [Google Scholar] [CrossRef]
- Vanderwolf, K.J.; McAlpine, D.F.; Malloch, D.; Forbes, G.J. Ectomycota associated with hibernating bats in eastern Canadian caves prior to the emergence of white-nose syndrome. Northeast. Nat. 2013, 20, 115–130. [Google Scholar] [CrossRef]
- Sazanova, K.V.; Senik, S.V.; Kirtsideli, I.Y.; Shavarda, A.L. Metabolomic profiling and lipid composition of Arctic and Antarctic strains of Micromycetes Geomyces pannorum and Thelebolus microsporus grown at different temperatures. Microbiology 2019, 88, 282–291. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Collection Number | ITS | LSU | Asc | Dis | Par | SpMor | Sp | Sp |
|---|---|---|---|---|---|---|---|---|---|
| Col | Orn | ||||||||
| Antarctomyces pellizariae | UFMGCB 12416 | KX576510 | N/A | 2 | 1 | 1 | 1 | 0 | 1 |
| Antarctomyces psychrotrophicus | VKM-4686 | MF375780 | MF375780 | 2 | 1 | 1 | 1 | 0 | 1 |
| Aotearoamyces nothofagi | PDD:55517 | KM677201 | MG807387 | 0 | 0 | 0 | 0 | 0 | 0 |
| Bettsia alvei | CBS 487.91 | MH862266 | MH873948 | 1 | 1 | 1 | 2 | 1 | 0 |
| Claussenomyces prasinulus | CBS 111551 | KX090866 | KX090815 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cleistothelebolus nipigonensis | CBS 778.70 | KC492060 | MH871738 | 1 | 1 | 1 | 1 | 0 | 0 |
| Gymnostellatospora alpina | CBS 620.81 | MH861383 | MH873132 | 1 | 1 | 1 | 1 | 0 | 1 |
| Gymnostellatospora bhattii | CBS 760.71 | MH860337 | MH872092 | 1 | 1 | 1 | 1 | 0 | 0 |
| Gymnostellatospora canadensis | UAMH 8899 | NR111199 | N/A | 1 | 1 | 1 | 1 | 0 | 1 |
| Gymnostellatospora japonica | VKM-4687 | MF375781 | MF375781 | 1 | 1 | 1 | 1 | 0 | 1 |
| Holwaya mucida | CNF 2/8749 | OM282975 | OM282978 | 0 | 0 | 0 | 0 | 0 | 0 |
| Leotia lubrica | KKM 337 | KF836617 | KF836627 | 0 | 0 | 0 | 0 | 0 | 0 |
| Leuconeurospora polypaeciloides | UAMH 11459 | KC884266 | N/A | 1 | 1 | 1 | 1 | 0 | 1 |
| Leuconeurospora pulcherrima | CBS 343.76 | KF049206 | FJ176884 | 1 | 1 | 1 | 1 | 0 | 1 |
| Microglossum olivaceum | FH-DSH97-103 | AY789398 | AY789397 | 0 | 0 | 0 | 0 | 0 | 0 |
| Myriodiscus sparassoides | KKUK2 | JX219380 | JX219382 | 0 | 0 | 0 | 2 | 0 | 0 |
| Patinella hyalophaea | H.B. 9739 | KT876978 | KT876978 | 0 | 0 | 0 | 1 | 0 | 0 |
| Pseudeurotium bakeri | CBS 878.71 | NR145345 | N/A | 1 | 1 | 1 | 2 | 1 | 0 |
| Pseudeurotium desertorum | CBS 986.72 | JX076946 | N/A | 1 | 1 | 1 | 2 | 1 | 0 |
| Pseudeurotium ovale | FMR 13600 | KP686192 | KP686193 | 1 | 1 | 1 | 1 | 1 | 0 |
| Pseudeurotium zonatum | CBS 329.36 | NR111127 | DQ470988 | 1 | 1 | 1 | 2 | 1 | 0 |
| Pseudogymnoascus appendiculatus | 02NH11 | JX270356 | KF017819 | 1 | 1 | 1 | 1 | 0 | 0 |
| Pseudogymnoascus lindneri | 02NH05 | JX270350 | KF017818 | 1 | 1 | 1 | 1 | 0 | 0 |
| Pseudogymnoascus roseus | 05NY06 | JX270385 | KF017824 | 1 | 1 | 1 | 1 | 0 | 0 |
| Pseudogymnoascus turneri | CUP-070715 | MN542214 | N/A | 1 | 1 | 1 | 1 | 0 | 0 |
| Pseudogymnoascus verrucosus | 04NY16 | JX270377 | KF017822 | 1 | 1 | 1 | 1 | 0 | 1 |
| Ramgea ozimecii | CNF 2/9997 | KY368752 | KY368753 | 0 | 0 | 0 | 1 | 0 | 1 |
| Thelebolus balaustiformis | MUT 2357 | NR159056 | MG816492 | 0 | 0 | 1 | 1 | 0 | 0 |
| Thelebolus ellipsoideus | CBS 113937 | AY957550 | FJ176895 | 1 | 1 | 1 | 1 | 0 | 0 |
| Thelebolus globosus | AFTOL-ID 5016 | KM822751 | FJ176905 | 1 | 1 | 1 | 1 | 0 | 0 |
| Thelebolus microcarpus | CBS 137501 | LN609269 | MH877647 | 1 | 1 | 1 | 1 | 0 | 0 |
| Thelebolus microsporus | CBS 115.53 | 16145654 | 16145752 | 0 | 0 | 0 | 1 | 0 | 0 |
| Thelebolus spongiae | MUT 2359 | MG813185 | MG816493 | 1 | 1 | 1 | 1 | 0 | 0 |
| Thelebolus stercoreus | CBS 717.69 | MH859395 | MH871166 | 0 | 0 | 1 | 1 | 0 | 0 |
| Thuemenidium atropurpureum | ILLS:61044 | JQ256427 | JQ256441 | 0 | 0 | 0 | 0 | 0 | 0 |
| Tympanis tsugae | LQH-8 | MH810146 | MH810148 | 0 | 0 | 0 | 2 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quijada, L.; Matočec, N.; Kušan, I.; Tanney, J.B.; Johnston, P.R.; Mešić, A.; Pfister, D.H. Apothecial Ancestry, Evolution, and Re-Evolution in Thelebolales (Leotiomycetes, Fungi). Biology 2022, 11, 583. https://doi.org/10.3390/biology11040583
Quijada L, Matočec N, Kušan I, Tanney JB, Johnston PR, Mešić A, Pfister DH. Apothecial Ancestry, Evolution, and Re-Evolution in Thelebolales (Leotiomycetes, Fungi). Biology. 2022; 11(4):583. https://doi.org/10.3390/biology11040583
Chicago/Turabian StyleQuijada, Luis, Neven Matočec, Ivana Kušan, Joey B. Tanney, Peter R. Johnston, Armin Mešić, and Donald H. Pfister. 2022. "Apothecial Ancestry, Evolution, and Re-Evolution in Thelebolales (Leotiomycetes, Fungi)" Biology 11, no. 4: 583. https://doi.org/10.3390/biology11040583
APA StyleQuijada, L., Matočec, N., Kušan, I., Tanney, J. B., Johnston, P. R., Mešić, A., & Pfister, D. H. (2022). Apothecial Ancestry, Evolution, and Re-Evolution in Thelebolales (Leotiomycetes, Fungi). Biology, 11(4), 583. https://doi.org/10.3390/biology11040583