
Assessment of Paracetamol Toxic Effects under Varying Seawater pH Conditions on the Marine Polychaete Hediste diversicolor Using Biochemical Endpoints


 ,
,  and
and 
Abstract
:Simple Summary
Abstract
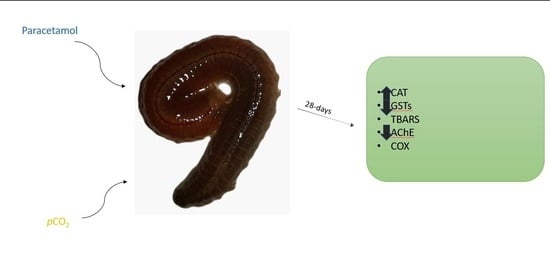
1. Introduction
2. Materials and Methods
2.1. Biological Sampling
2.2. Chemicals and Test Concentrations
2.3. SPE and LC-MS/MS Procedures
2.4. Experimental Design
2.5. Tissue Processing
2.6. Biomarker Quantification
2.7. Statistical Analysis
3. Results
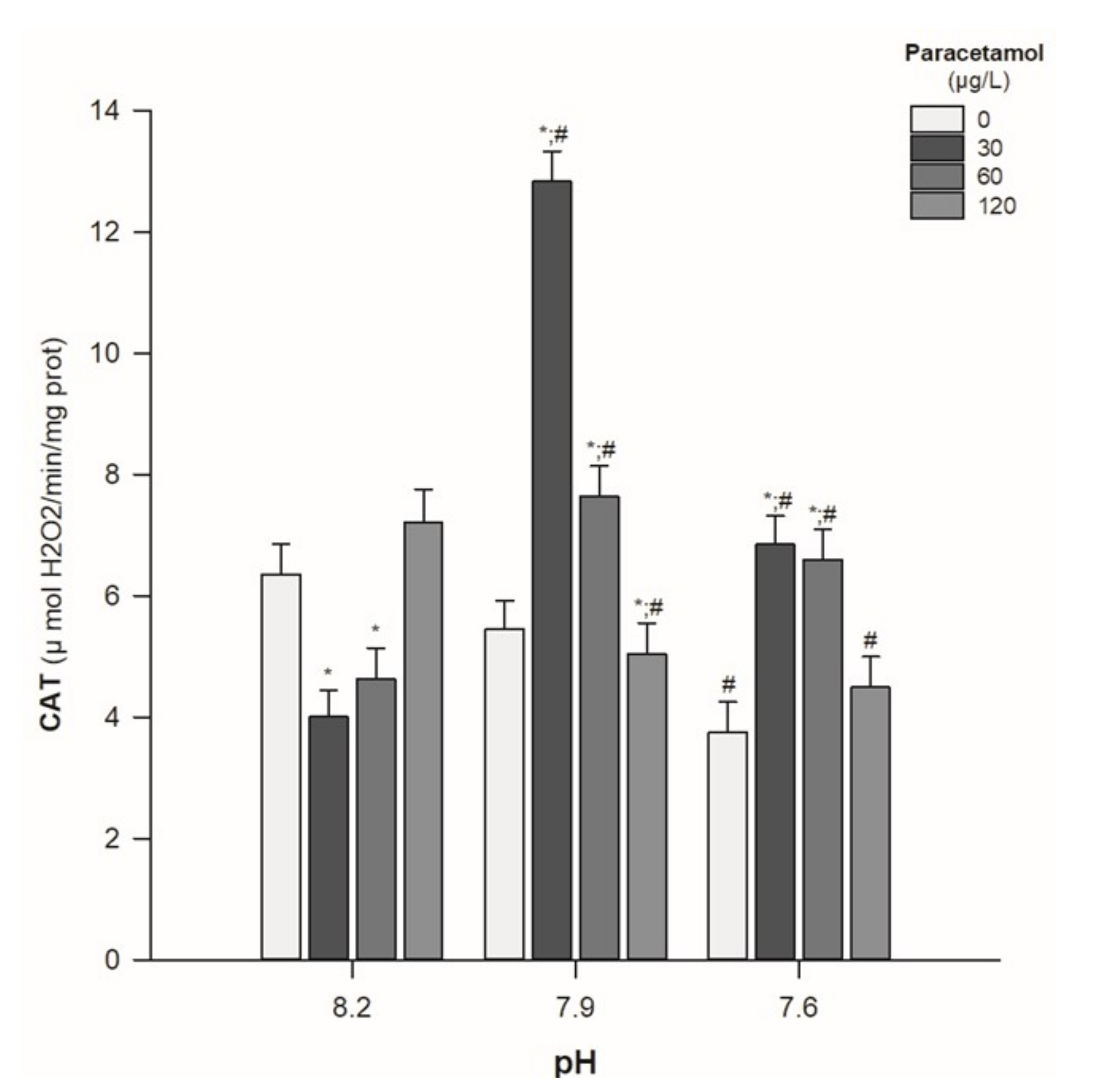
- Catalase activity
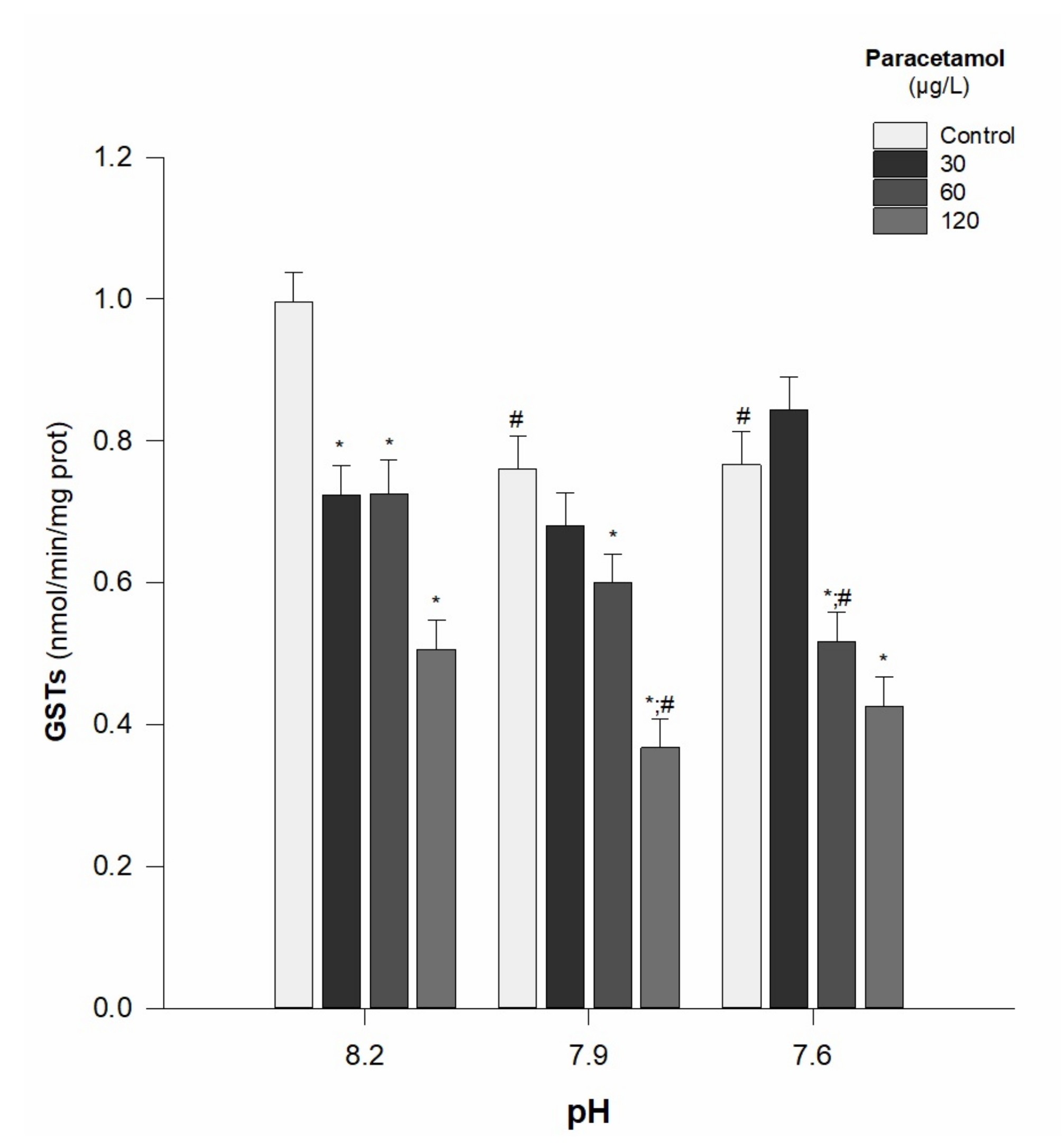
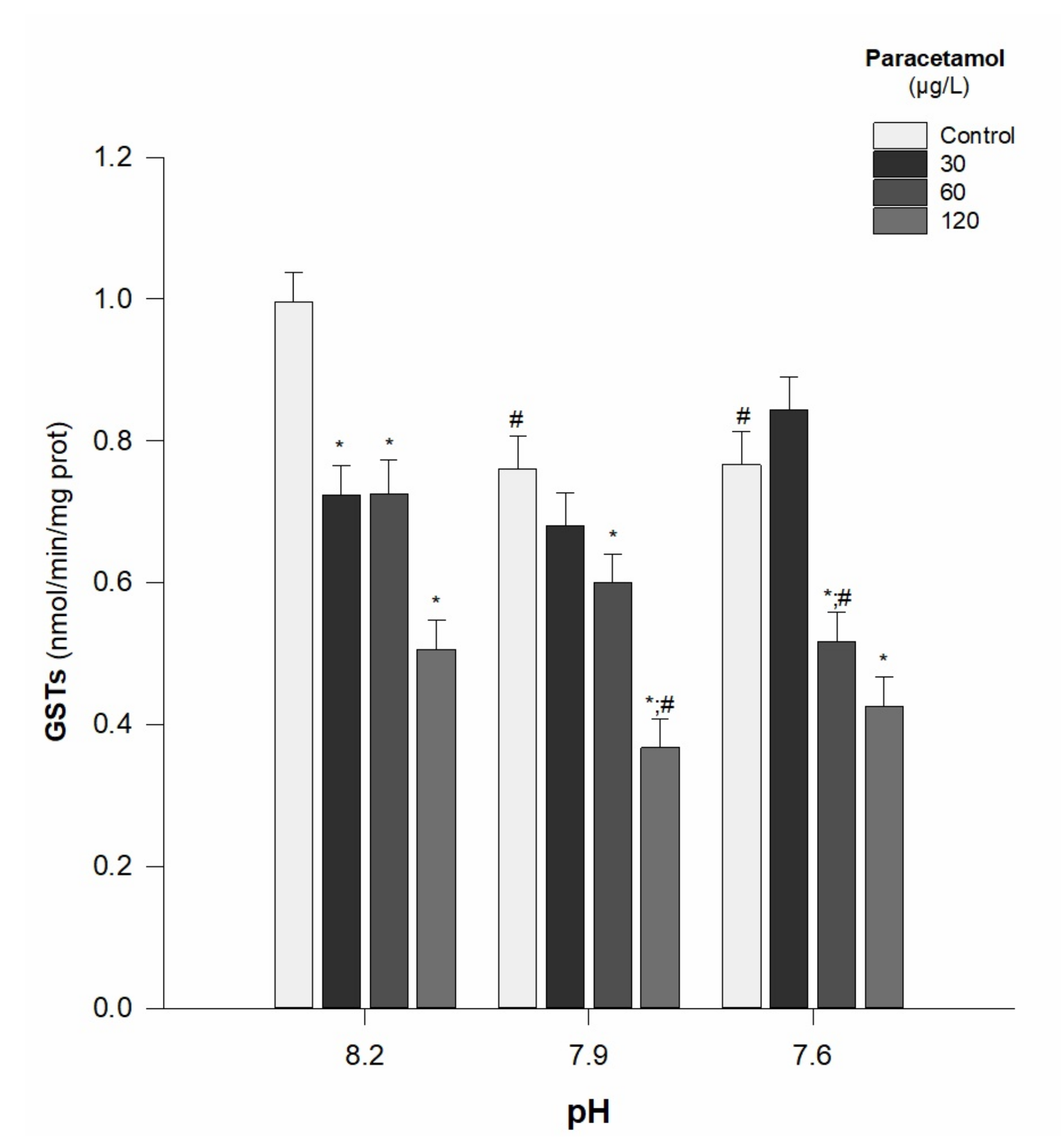
- GSTs activity
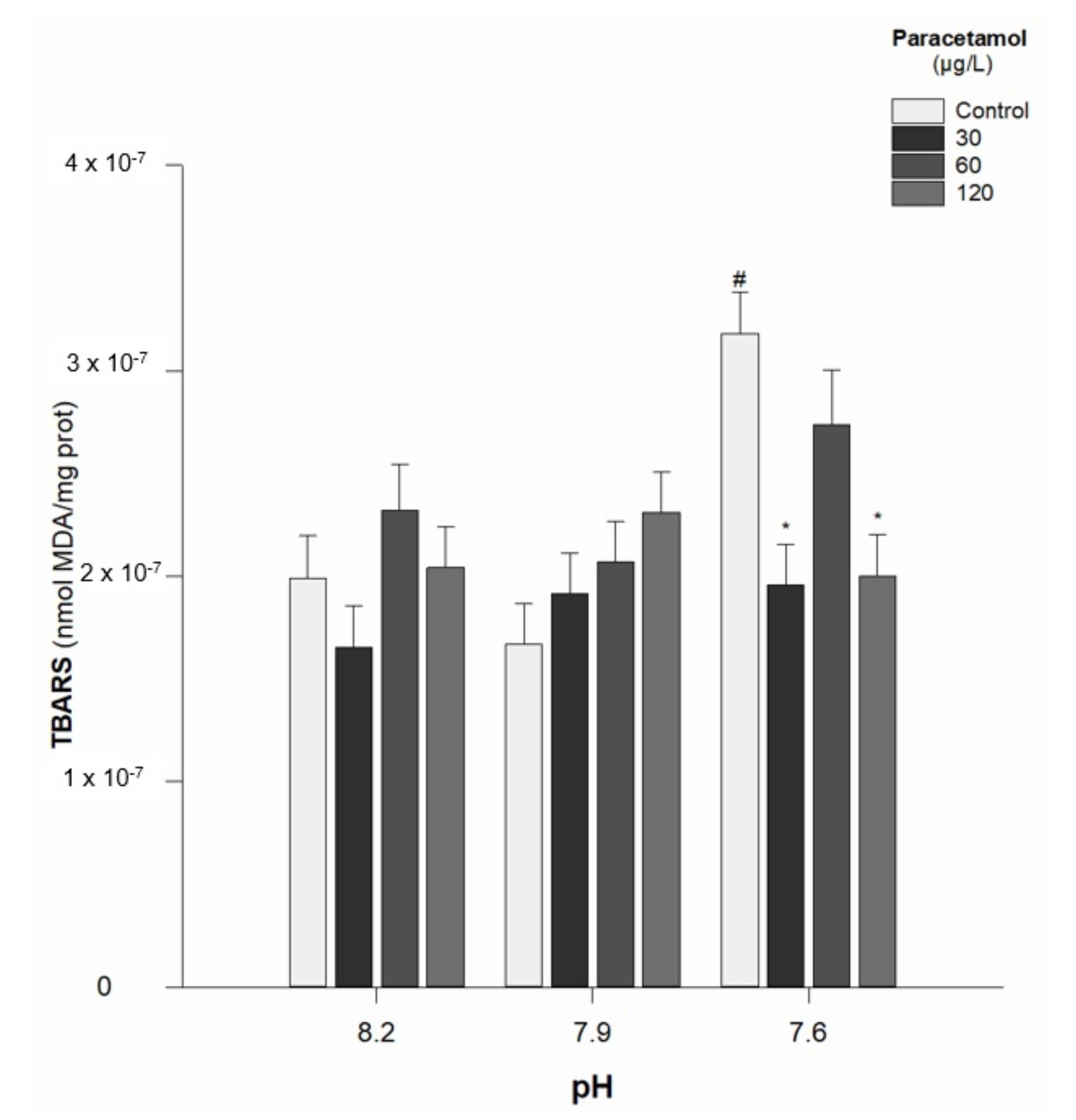
- Lipid peroxidation
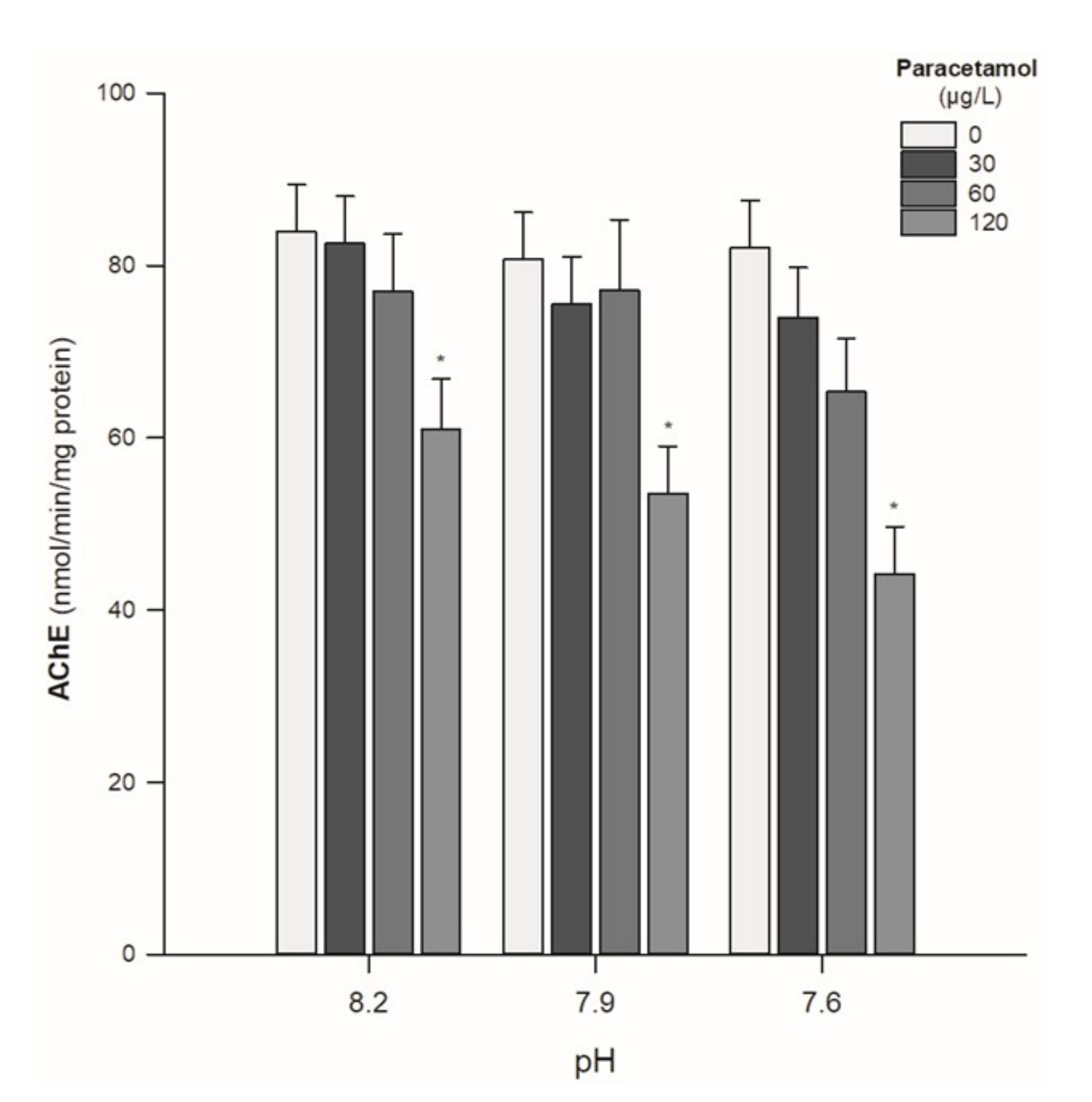
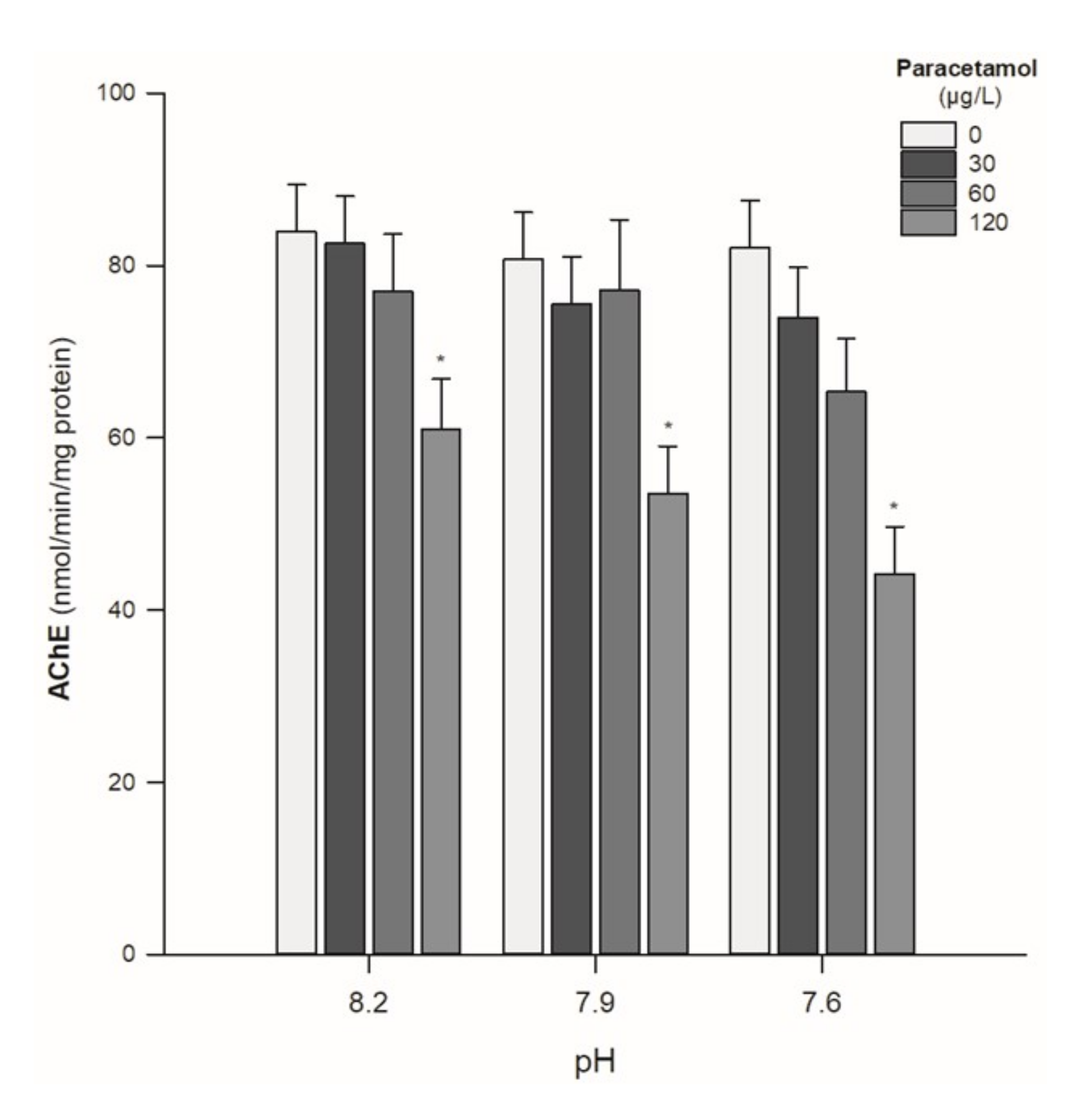
- AChE activity
- COX activity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andrew, S.B.; Michael, J.K. Impacts of Climate Change on Marine Organisms and Ecosystems. Curr. Biol. 2009, 19, 602–614. [Google Scholar]
- Le Quéré, C.; Raupach, M.R.; Canadell, J.G.; Marland, G.; Bopp, L.; Ciais, P.; Conway, T.J.; Doney, S.C.; Feely, R.A.; Foster, P.; et al. Trends in the sources and sinks of carbon dioxide. Nat. Geosci. 2009, 2, 831–836. [Google Scholar] [CrossRef]
- Mishra, S.; Nehal, A.S. A Review on Environmental and Health Impacts of Cement Manufacturing Emissions. J. Geol. Agric. Environ. Sci. 2014, 2, 27–31. [Google Scholar]
- IPCC. Climate change 2007: The physical science basis. Intergov. Panel Clim. Chang. 2007, 7137, 727–728. [Google Scholar]
- Knorr, W. Is the airborne fraction of anthropogenic CO2 emissions increasing? Geophys. Res. Lett. 2009, 36, 1–5. [Google Scholar] [CrossRef]
- Jackson, J.B.C. The future of the oceans past. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3765–3778. [Google Scholar] [CrossRef] [Green Version]
- Elderfield, H.; Hoegh-Guldberg, O.; Liss, P.; Riebesell, U.; Shepherd, J.; Turley, C.; Watson, A. Ocean acidification due to increasing atmospheric carbon dioxide. In The Royal Society; The Clyvedon Press Ltd.: Cardiff, UK, 2005. [Google Scholar]
- Caldeira, K.; Wickett, M.E. Anthropogenic carbon and ocean pH. Nature 2003, 425, 365. [Google Scholar] [CrossRef]
- Key, R.M.; Kozyr, A.; Sabine, C.L.; Lee, K.; Wanninkhof, R.; Bullister, J.L.; Feely, R.A.; Millero, F.J.; Mordy, C.; Peng, T.H. A global ocean carbon climatology: Results from Global Data Analysis Project (GLODAP). Glob. Biogeochem. Cycles 2004, 18, 1–23. [Google Scholar] [CrossRef]
- Jacobson, M.Z. Studying ocean acidification with conservative, stable numerical schemes for non equilibrium air-ocean exchange and ocean equilibrium chemistry. J. Geophys. Res. Atmos. 2005, 110, 1–17. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2013: The Physical Science Basis Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; Volume 10, pp. 1–30. [Google Scholar]
- Hartin, C.A.; Bond-Lamberty, B.; Patel, P.; Mundra, A. Ocean acidification over the next three centuries using a simple global climate carbon-cycle model: Projections and sensitivities. Biogeosciences 2016, 13, 4329–4342. [Google Scholar] [CrossRef] [Green Version]
- Moiseenko, T.I. Effects of acidification on aquatic ecosystems. Russ. J. Ecol. 2005, 36, 93–102. [Google Scholar] [CrossRef]
- Skjelkvåle, B.L.; Aherne, J.; Bergman, T.; Bishop, K.; Forsius, M.; Forsström, L.; Gashinka, N.A.; Hettelingh, J.P.; Jeffries, D.S.; Kaste, O.; et al. Effects on freshwater ecosystems. In AMAP Assessment 2006: Acidifying Pollutants, Arctic Haze, and Acidification; AMAP: Oslo, Norway, 2006; Volume 13, pp. 64–90. [Google Scholar]
- Sauvé, S.; Desrosiers, M. A review of what is an emerging contaminant. Chem. Cent. J. 2014, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapworth, D.J.; Baran, N.; Stuart, M.E.; Ward, R.S. Emerging organic contaminants in groundwater: A review of sources, fate and occurrence. Environ. Pollut. 2012, 163, 287–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huerta, B.; Rodríguez-Mozaz, S.; Barceló, D. Pharmaceuticals in biota in the aquatic environment: Analytical methods and environmental implications. Anal. Bioanal. Chem. 2012, 404, 2611–2624. [Google Scholar] [CrossRef] [PubMed]
- Deziel, N. Pharmaceuticals in Wastewater Treatment Plant Effluent Waters. Sch. Horizons Univ. Minn. Morris Undergrad. J. 2014, 1, 12. [Google Scholar]
- Ebele, A.J.; Abou-Elwafa Abdallah, M.; Harrad, S. Pharmaceuticals and personal care products (PPCPs) in the freshwater aquatic environment. Emerg. Contam. 2016, 3, 1–16. [Google Scholar] [CrossRef]
- Fent, K.; Weston, A.A.; Caminada, D. Ecotoxicology of human pharmaceuticals. Aquat. Toxicol. 2006, 76, 122–159. [Google Scholar] [CrossRef]
- Buchberger, W. Current approaches to trace analysis of pharmaceuticals and personal care products in the environment. J. Chromatogr. A 2011, 1218, 603–618. [Google Scholar] [CrossRef]
- Gaw, S.; Thomas, K.V.; Hutchinson, T.H. Sources, impacts and trends of pharmaceuticals in the marine and coastal environment. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130572. [Google Scholar] [CrossRef] [Green Version]
- Küster, A.; Adler, N. Pharmaceuticals in the environment: Scientific evidence of risks and its regulation. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2014, 369, 20130587. [Google Scholar] [CrossRef] [Green Version]
- Meek, I.L.; van de Laar, M.A.F.J.; Vonkeman, H.E. Non-steroidal anti-inflammatory drugs: An overview of cardiovascular risks. Pharmaceuticals 2010, 3, 2146–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Rey, M.; Bebianno, M.J. Effects of non-steroidal anti-inflammatory drug (NSAID) diclofenac exposure in mussel Mytilus galloprovincialis. Aquat. Toxicol. 2014, 148, 818–831. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.M.; McGuire, T.M.; van Driel, M.L. Consumer concerns about paracetamol: A retrospective analysis of a medicines call centre. BMJ Open 2016, 6, e010860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jóźwiak-Bebenista, M.; Nowak, J.Z. Paracetamol: Mechanism of action, applications and safety concern. Acta Pol. Pharm.-Drug Res. 2014, 71, 11–23. [Google Scholar]
- Jaeschke, H. Acetaminophen: Dose-Dependent Drug Hepatotoxicity and Acute Liver Failure in Patients. Dig. Dis. 2015, 33, 464–471. [Google Scholar] [CrossRef] [Green Version]
- Roberts, P.H.; Thomas, K.V. The occurrence of selected pharmaceuticals in wastewater effluent and surface waters of the lower Tyne catchment. Sci. Total Environ. 2006, 356, 143–153. [Google Scholar] [CrossRef]
- Togola, A.; Budzinski, H. Multi-residue analysis of pharmaceutical compounds in aqueous samples. J. Chromatogr. A 2008, 1177, 150–158. [Google Scholar] [CrossRef]
- Al-kaf, A.G.; Naji, K.M.; Yusuf, Q.M.A.; Edrees, W.H.A. Occurrence of Paracetamol in Aquatic Environments and Transformation by Microorganisms: A Review. Chronicles Pharm. Sci. 2017, 1, 341–355. [Google Scholar]
- K’oreje, K.O.; Kandie, F.J.; Vergeynst, L.; Abira, M.A.; Van Langenhove, H.; Okoth, M.; Demeestere, K. Occurrence, fate and removal of pharmaceuticals, personal care products and pesticides in wastewater stabilization ponds and receiving rivers in the Nzoia Basin, Kenya. Sci. Total Environ. 2018, 6637, 336–348. [Google Scholar] [CrossRef]
- Ramos, A.S.; Correia, A.T.; Antunes, S.C.; Gonçalves, F.; Nunes, B. Effect of acetaminophen exposure in Oncorhynchus mykiss gills and liver: Detoxification mechanisms, oxidative defence system and peroxidative damage. Environ. Toxicol. Pharmacol. 2014, 37, 1221–1228. [Google Scholar] [CrossRef]
- Nunes, B.; Verde, M.F.; Soares, A.M.V.M. Biochemical effects of the pharmaceutical drug paracetamol on Anguilla anguilla. Environ. Sci. Pollut. Res. 2015, 22, 11574–11584. [Google Scholar] [CrossRef] [PubMed]
- Daniel, D.; Dionísio, R.; Alkimin, G.D.; Nunes, B. Acute and chronic effects of paracetamol exposure on Daphnia magna: How oxidative effects may modulate responses at distinct levels of organization in a model species. Environ. Sci. Pollut. Res. 2018, 26, 3320–3329. [Google Scholar] [CrossRef]
- Oliveira, L.L.D.; Antunes, S.C.; Gonçalves, F.; Rocha, O.; Nunes, B. Evaluation of ecotoxicological effects of drugs on Daphnia magna using different enzymatic biomarkers. Ecotoxicol. Environ. Saf. 2015, 119, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Nunes, B.; Nunes, J.; Soares, A.M.V.M.; Figueira, E.; Freitas, R. Toxicological effects of paracetamol on the clam Ruditapes philippinarum: Exposure vs recovery. Aquat. Toxicol. 2017, 192, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.; Coelho, D.; Pires, A.; Soares, A.M.V.M.; Figueira, E.; Nunes, B. Preliminary evaluation of Diopatra neapolitana regenerative capacity as a biomarker for paracetamol exposure. Environ. Sci. Pollut. Res. 2015, 22, 13382–13392. [Google Scholar] [CrossRef] [PubMed]
- Nunes, B.; Antunes, S.C.; Santos, J.; Martins, L.; Castro, B.B. Toxic potential of paracetamol to freshwater organisms: A headache to environmental regulators? Ecotoxicol. Environ. Saf. 2014, 107, 178–185. [Google Scholar] [CrossRef]
- Crouthamel, W.G.; Tan, G.H.; Dittert, L.W.; Doluisio, J.T. Drug absorption IV: Influence of pH on absorption kinetics of weakly acidic drugs. J. Pharm. Sci. 1971, 60, 1160–1163. [Google Scholar] [CrossRef]
- Martinez, M.N.; Amidon, G.L. A Mechanistic Approach to Understanding the Factors Affecting Drug Absorption: A Review of Fundamentals. J. Clin. Pharmacol. 2002, 42, 620–643. [Google Scholar] [CrossRef] [Green Version]
- Mitra, A.; Kesisoglou, F. Impaired drug absorption due to high stomach pH: A review of strategies for mitigation of such effect to enable pharmaceutical product development. Mol. Pharm. 2013, 10, 3970–3979. [Google Scholar] [CrossRef]
- Hu, M.; Lin, D.; Shang, Y.; Hu, Y.; Lu, W.; Huang, X.; Ning, K.; Chen, Y.; Wang, Y. CO2-induced pH reduction increases physiological toxicity of nano-TiO2 in the mussel Mytilus coruscus. Sci. Rep. 2017, 7, 40015. [Google Scholar] [CrossRef] [Green Version]
- Dionísio, R.; Daniel, D.; Arenas, F.; Campos, J.C.; Costa, P.J.C.; Nunes, B.; Correia, A.T. Effects of pH on salicylic acid toxicity in terms of biomarkers determined in the marine gastropod Gibbula umbilicalis. Mar. Environ. Res. 2020, 158, 104995. [Google Scholar] [CrossRef] [PubMed]
- Dastmalchi, S.; Rashidi, M.; Rassi, M. Simultaneous determination of the pka and octanol/water partition coefficient (pm) of acetaminophen. J. Sch. Pharm. Med. Sci. 1995, 4, 7–14. [Google Scholar]
- Costa, P.F.E.; Gil, J.; Passos, A.M.; Pereira, P.; Melo, P.; Batista, F.; da Fonseca, L.C. The market features of imported non-indigenous polychaetes in Portugal and consequent ecological concerns. Sci. Mar. 2006, 70, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Thit, A.; Dybowska, A.; Købler, C.; Kennaway, G.; Selck, H. Influence of copper oxide nanoparticle shape on bioaccumulation, cellular internalization and effects in the estuarine sediment-dwelling polychaete, Nereis diversicolor. Mar. Environ. Res. 2015, 111, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Catalano, B.; Moltedo, G.; Martuccio, G.; Gastaldi, L.; Virno-Lamberti, C.; Lauria, A.; Ausili, A. Can Hediste diversicolor (Nereidae, Polychaete) be considered a good candidate in evaluating PAH contamination? A multimarker approach. Chemosphere 2012, 86, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Scaps, P.; Borot, O. Acetylcholinesterase activity of the polychaete Nereis diversicolor: Effects of temperature and salinity. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 2000, 125, 377–383. [Google Scholar] [CrossRef]
- Pook, C.; Lewis, C.; Galloway, T. The metabolic and fitness costs associated with metal resistance in Nereis diversicolor. Mar. Pollut. Bull. 2009, 58, 1063–1071. [Google Scholar] [CrossRef]
- Buffet, P.E.; Poirier, L.; Zalouk-Vergnoux, A.; Lopes, C.; Amiard, J.C.; Gaudin, P.; Risso-de Faverney, C.; Guibbolini, M.; Gilliland, D.; Perrein-Ettajani, H.; et al. Biochemical and behavioural responses of the marine polychaete Hediste diversicolor to cadmium sulfide quantum dots (CdS QDs): Waterborne and dietary exposure. Chemosphere 2014, 100, 63–70. [Google Scholar] [CrossRef]
- Maranho, L.A.; André, C.; DelValls, T.A.; Gagné, F.; Martín-Díaz, M.L. Toxicological evaluation of sediment samples spiked with human pharmaceutical products: Energy status and neuroendocrine effects in marine polychaetes Hediste diversicolor. Ecotoxicol. Environ. Saf. 2015, 118, 27–36. [Google Scholar] [CrossRef]
- Nunes, B. Acute ecotoxicological effects of salicylic acid on the Polychaeta species Hediste diversicolor: Evidences of low to moderate pro-oxidative effects. Environ. Sci. Pollut. Res. 2019, 26, 7873–7882. [Google Scholar] [CrossRef]
- Nunes, B.; Costa, M. Study of the effects of zinc pyrithione in biochemical parameters of the Polychaeta Hediste diversicolor: Evidences of neurotoxicity at ecologically relevant concentrations. Environ. Sci. Pollut. Res. 2019, 26, 13551–13559. [Google Scholar] [CrossRef] [PubMed]
- Hamza-Chaffai, A. Usefulness of Bioindicators and Biomarkers in Pollution Biomonitoring. Int. J. Biotechnol. Wellness Ind. 2014, 3, 19–26. [Google Scholar] [CrossRef]
- Tomanek, L.; Zuzow, M.J.; Ivanina, A.V.; Beniash, E.; Sokolova, I.M. Proteomic response to elevated pCO2 level in eastern oysters, Crassostrea virginica: Evidence for oxidative stress. J. Exp. Biol. 2011, 214, 1836–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, R.; Pires, A.; Moreira, A.; Wrona, F.J.; Figueira, E.; Soares, A.M.V.M. Biochemical alterations induced in Hediste diversicolor under seawater acidification conditions. Mar. Environ. Res. 2016, 117, 75–84. [Google Scholar] [CrossRef]
- Zhang, D.; Guo, D.; Wang, G.; Li, S. Response of antioxidant defense system in copepod Calanus sinicus Brodsky exposed to CO2-acidified seawater. Acta Oceanol. Sin. 2016, 35, 82–88. [Google Scholar] [CrossRef]
- Brandão, F.P.; Pereira, J.L.; Gonçalves, F.; Nunes, B. The impact of paracetamol on selected biomarkers of the mollusc species Corbicula fluminea. Environ. Toxicol. 2014, 29, 74–83. [Google Scholar] [CrossRef]
- Delwing-de Lima, D.; Wollinger, L.F.; Casagrande, A.C.M.; Delwing, F.; da Cruz, J.G.P.; Wyse, A.T.S.; Delwing-Dal Magro, D. Guanidino compounds inhibit acetylcholinesterase and butyrylcholinesterase activities: Effect neuroprotector of vitamins E plus C. Int. J. Dev. Neurosci. 2010, 28, 456–473. [Google Scholar] [CrossRef]
- SFT. Guidelines for classification of environmental quality in fjords and coastal areas. In Revision of Classification of Metals and Organic Contaminants in Water and Sediment; Norwegian Pollution Control Authority SFT TA-2229/2007; SFT: San Francisco, CA, USA, 2007. [Google Scholar]
- Ghribi, R.; Correia, A.T.; Elleuch, B.; Nunes, B. Toxicity assessment of impacted sediments from southeast coast of Tunisia using a biomarker approach with the polychaete Hediste diversicolor. Arch. Environ. Contam. Toxicol. 2019, 76, 678–691. [Google Scholar] [CrossRef]
- ASTM E1562-00; Standard Guide for Conducting Acute, Chronic, and Life-Cycle Aquatic Toxicity Tests with Polychaetous Annelids; ASTM International: West Conshohocken, PA, USA, 2013.
- Bagarrão, R.M.E. Reprodução e Crescimento do Poliqueta Hediste diversicolor (O.F. Müller, 1776) sob Diferentes Condições Ambientais. Master’s Thesis, University of Aveiro, Aveiro, Portugal, 2013. Volume 44. [Google Scholar]
- Durou, C.; Mouneyrac, C.; Amiard-Triquet, C. Environmental quality assessment in estuarine ecosystems: Use of biometric measurements and fecundity of the ragworm Nereis diversicolor (Polychaeta, Nereididae). Water Res. 2008, 42, 2157–2165. [Google Scholar] [CrossRef]
- Żur, J.; Piński, A.; Marchlewicz, A.; Hupert-Kocurek, K.; Wojcieszyńska, D.; Guzik, U. Organic micropollutants paracetamol and ibuprofen—Toxicity, biodegradation, and genetic background of their utilization by bacteria. Environ. Sci. Pollut. Res. Int. 2018, 25, 21498. [Google Scholar] [CrossRef] [Green Version]
- Olaitan, J.O.; Chimezie, A.; Tolulope, B.; Aminat, T.T. Determination of pharmaceutical compounds in surface and underground water by solid phase extraction-liquid chromatography. J. Environ. Chem. Ecotoxicol. 2014, 6, 20–26. [Google Scholar]
- Afonso-Olivares, C.; Torres-Padrón, M.E.; Sosa-Ferrera, Z.; Santana-Rodríguez, J.J. Assessment of the Presence of Pharmaceutical Compounds in Seawater Samples from Coastal Area of Gran Canaria Island (Spain). Antibiotics 2013, 2, 274–287. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, P.G.; Loganimoce, E.M.; Neuparth, T.; Rocha, M.J.; Rocha, E.; Arenas, F. Interactive effects of increased temperature, pCO2 and the synthetic progestin levonorgestrel on the fitness and breeding of the amphipod Gammarus locusta. Environ. Pollut. 2018, 236, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Nunes, B.; Carvalho, F.; Guilhermino, L. Behaviour and biomarkers of oxidative stress in Gambusia holbrooki after acute exposure to widely used pharmaceuticals and a detergent. Ecotoxicol. Environ. Saf. 2008, 71, 341–354. [Google Scholar] [CrossRef]
- Nunes, B.; Carvalho, F.; Guilhermino, L. Characterization and use of the total head soluble cholinesterases from mosquitofish (Gambusia holbrooki) for screening of anticholinesterasic activity. J. Enzym. Inhib. Med. Chem. 2005, 20, 369–376. [Google Scholar] [CrossRef]
- Petrovic, N.; Murray, M. Using N,N,N’,N’-tetramethyl-p-phenylenediamine (TMPD) to Assay Cyclooxygenase Activity In Vitro. Adv. Protoc. Oxidative Stress III 2009, 594, 1–477. [Google Scholar]
- Aebi, H. Oxygen Radicals in Biological Systems. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases the first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7140. [Google Scholar] [CrossRef]
- Büege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–331. [Google Scholar]
- Scaps, P.; Demuynck, S.; Descamps, M.; Dhainaut, A. Biochemical and enzymatic characterization of an acetylcholinesterase from Nereis diversicolor(Annelida, Polychaeta): Comparison with the cholinesterases of Eisenia fetida (Anelida, Oligochaeta). Biol. Bull. Mar. Biol. Lab. 1996, 190, 396–402. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef] [Green Version]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2017, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.J.; Hendriks, B.S.; Zhao, J.; de Graaf, D. Multiple effects of acetaminophen and p38 inhibitors: Towards pathway toxicology. FEBS Lett. 2008, 582, 1276–1282. [Google Scholar] [CrossRef] [Green Version]
- Parolini, M.; Binelli, A.; Cogni, D.; Provini, A. Multi-biomarker approach for the evaluation of the cytogenotoxicity of paracetamol on the zebra mussel (Dreissena polymorpha). Chemosphere 2010, 79, 489–498. [Google Scholar] [CrossRef]
- Sun, T.; Tang, X.; Jiang, Y.; Wang, Y. Seawater acidification induced immune function changes of haemocytes in Mytilus edulis: A comparative study of CO2 and HCl enrichment. Sci. Rep. 2017, 7, 41488. [Google Scholar] [CrossRef]
- Lores Arnaiz, S.; Llesuy, S.; Cutrín, J.C.; Boveris, A. Oxidative stress by acute acetaminophen administration in mouse liver. Free Radic. Biol. Med. 1995, 19, 303–310. [Google Scholar] [CrossRef]
- Kennedy, G.Y.; Nicol, J.A.C. Pigments of Chaetopterus variopedatus (Polychaeta). Proc. R. Soc. Biol. Sci. 1959, 150, 509–538. [Google Scholar]
- Abele-Oeschger, D.; Oeschger, R.; Theede, H. Biochemical adaptations of Nereis diversicolor(Polychaeta) to temporarily increased hydrogen peroxide levels in intertidal sandflats. Mar. Ecol. Prog. Ser. 1994, 106, 101–110. [Google Scholar] [CrossRef]
- Costa, P.M.; Carrapiço, F.; Alves de Matos, A.P.; Costa, M.H. A microscopical study of the “chlorophylloid” pigment cells of the marine polychaete Eulalia viridis (L.). Microsc. Microanal. 2013, 19, 15–16. [Google Scholar] [CrossRef] [Green Version]
- Pörtner, H.O.; Langenbuch, M.; Reipschläger, A. Biological Impact of Elevated Ocean CO2 Concentrations: Lessons from Animal Physiology and Earth History. J. Oceanogr. 2004, 60, 705–718. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Mei, C.F.; Ying, G.G.; Xu, M.Y. Toxicity Thresholds for Diclofenac, Acetaminophen and Ibuprofen in the Water Flea Daphnia magna. Bull. Environ. Contam. Toxicol. 2016, 97, 84–90. [Google Scholar] [CrossRef]
- Manimaran, A.; Nath Sarkar, S.; Sankar, P. Toxicodynamics of subacute co-exposure to groundwater contaminant arsenic and analgesic-antipyretic drug acetaminophen in rats. Ecotoxicol. Environ. Saf. 2010, 73, 94–100. [Google Scholar] [CrossRef]
- Ben-Shachar, R.; Chen, Y.; Luo, S.; Hartman, C.; Reed, M.; Nijhout, H.F. The biochemistry of acetaminophen hepatotoxicity and rescue: A mathematical model. Theor. Biol. Med. Model. 2012, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Li, S.; Wang, G.; Guo, D.; Xing, K.; Zhang, S. Biochemical responses of the copepod Centropages tenuiremis to CO2-driven acidified seawater. Water Sci. Technol. 2012, 65, 30–37. [Google Scholar] [CrossRef]
- Todgham, A.E.; Hofmann, G.E. Transcriptomic response of sea urchin larvae Strongylocentrotus purpuratus to CO2 driven seawater acidification. J. Exp. Biol. 2009, 212, 2579–2594. [Google Scholar] [CrossRef] [Green Version]
- Hinson, J.A.; Roberts, D.W.; James, L.P. Mechanisms of Acetaminophen-Induced Liver Necrosis. Handb. Exp. Pharmacol. 2010, 196, 369–405. [Google Scholar]
- Thijssen, H.H.W.; Soute, B.A.; Vervoort, L.M.; Claessens, J.G. Paracetamol (acetaminophen) warfarin interaction: NAPQI, the toxic metabolite of paracetamol, is an inhibitor of enzymes in the vitamin K cycle. Thromb. Haemost. 2004, 92, 797–802. [Google Scholar]
- Attri, P.; Choi, E.H. Influence of Reactive Oxygen Species on the Enzyme Stability and Activity in the Presence of Ionic Liquids. PLoS ONE 2013, 8, 142–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxico 2005, 45, 51–88. [Google Scholar] [CrossRef] [PubMed]
- Giménez, V.; Nunes, B. Effects of commonly used therapeutic drugs, paracetamol, and acetylsalicylic acid, on key physiological traits of the sea snail Gibbula umbilicalis. Environ. Sci. Pollut. Res. 2019, 26, 21858–21870. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Yang, Z.; Dou, Z.; Sun, F.; Kou, S.; Zhang, Z.; Huang, X.; Bao, Z. Impact of ocean acidification on the energy metabolism and antioxidant responses of the Yesso scallop (I). Front. Physiol. 2019, 9, 1967. [Google Scholar] [CrossRef] [Green Version]
- Aguirre-Martínez, G.V.; DelValls, T.A.; Martín-Díaz, M.L. General stress, detoxification pathways, neurotoxicity and genotoxicity evaluated in Ruditapes philippinarum exposed to human pharmaceuticals. Ecotoxicol. Environ. Saf. 2016, 124, 18–31. [Google Scholar] [CrossRef]
- Solé, M.; Shaw, J.P.; Frickers, P.E.; Readman, J.W.; Hutchinson, T.H. Effects on feeding rate and biomarker responses of marine mussels experimentally exposed to propranolol and acetaminophen. Anal. Bioanal. Chem. 2010, 396, 649–656. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostangladins Arter. Thromb. Vasc. Boil. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Cashman, J.N. The Mechanisms of Action of NSAIDs in Analgesia. Drugs 1996, 52, 13–23. [Google Scholar] [CrossRef]
- Hinz, B.; Cheremina, O.; Brune, K. Acetaminophen (paracetamol) is a selective cyclooxygenase-2 inhibitor in man. FASEB J. 2008, 22, 383–390. [Google Scholar] [CrossRef]
- Ghanem, C.I.; Pérez, M.J.; Manautou, J.E.; Mottino, A.D. Acetaminophen from liver to brain: New insights into drug pharmacological action and toxicity. Pharmacol. Res. 2016, 109, 119–131. [Google Scholar] [CrossRef] [Green Version]





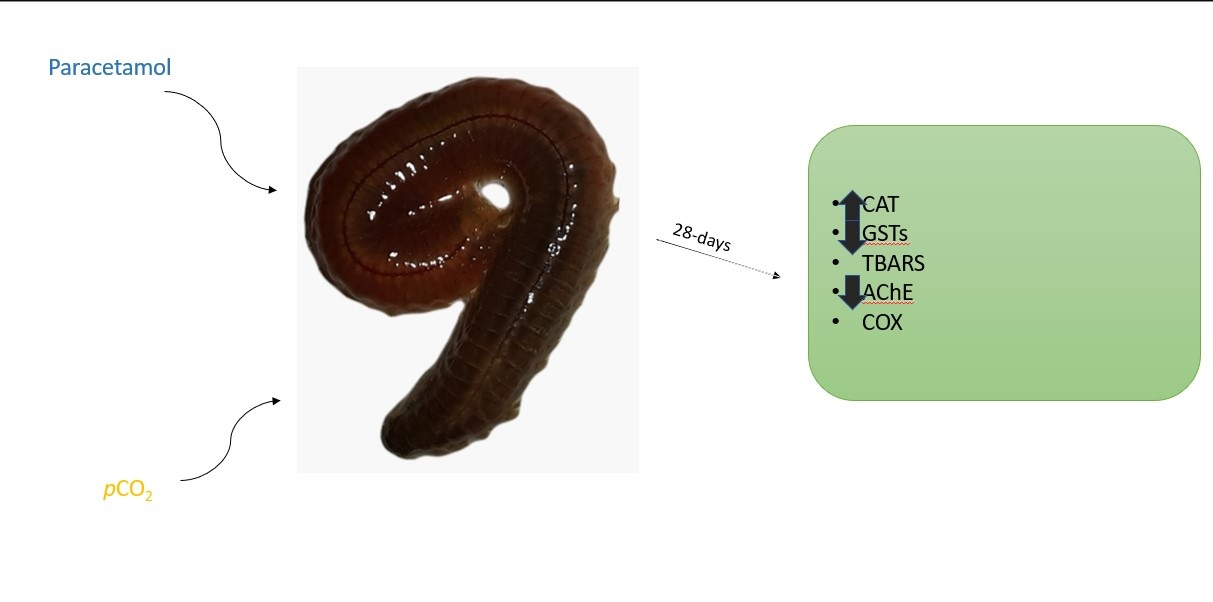{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Retention Time | Mass Transition | Cone Voltage | Collision Energy |
|---|---|---|---|---|
| Acetaminophen | 2.08 ± 0.05 | 152.2 > 65 a | 25 | 25 |
| 152.2 > 110.3 b | 25 | 15 | ||
| Acetaminophen-D4 | 2.08 ± 0.05 | 156.2 > 114.2 | 25 | 15 |
| Concentration | pH | pH × Concentration | Total | ||
|---|---|---|---|---|---|
| DF | 3 | 2 | 6 | 97 | |
| GSTs | F | 50.174 | 10.487 | 3.663 | |
| p-value | <0.0001 | <0.0001 | <0.0001 | ||
| DF | 3 | 2 | 6 | 86 | |
| CAT | F | 12.474 | 23.228 | 27.406 | |
| p-value | <0.0001 | <0.0001 | <0.0001 | ||
| DF | 3 | 2 | 6 | 100 | |
| TBARs | F | 3.795 | 6.622 | 4.246 | |
| p-value | 0.013 | 0.002 | <0.0001 | ||
| DF | 3 | 2 | 6 | 95 | |
| AChE | F | 16.166 | 2.287 | 0.437 | |
| p-value | <0.0001 | 0.065 | 0.952 | ||
| DF | 3 | 2 | 6 | 58 | |
| COX | F | 7.705 | 4.42 | 2.912 | |
| p-value | <0.0001 | 0.020 | 0.017 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daniel, D.; Nunes, B.; Pinto, E.; Ferreira, I.M.P.L.V.O.; Correia, A.T. Assessment of Paracetamol Toxic Effects under Varying Seawater pH Conditions on the Marine Polychaete Hediste diversicolor Using Biochemical Endpoints. Biology 2022, 11, 581. https://doi.org/10.3390/biology11040581
Daniel D, Nunes B, Pinto E, Ferreira IMPLVO, Correia AT. Assessment of Paracetamol Toxic Effects under Varying Seawater pH Conditions on the Marine Polychaete Hediste diversicolor Using Biochemical Endpoints. Biology. 2022; 11(4):581. https://doi.org/10.3390/biology11040581
Chicago/Turabian StyleDaniel, David, Bruno Nunes, Edgar Pinto, Isabel M. P. L. V. O. Ferreira, and Alberto Teodorico Correia. 2022. "Assessment of Paracetamol Toxic Effects under Varying Seawater pH Conditions on the Marine Polychaete Hediste diversicolor Using Biochemical Endpoints" Biology 11, no. 4: 581. https://doi.org/10.3390/biology11040581
APA StyleDaniel, D., Nunes, B., Pinto, E., Ferreira, I. M. P. L. V. O., & Correia, A. T. (2022). Assessment of Paracetamol Toxic Effects under Varying Seawater pH Conditions on the Marine Polychaete Hediste diversicolor Using Biochemical Endpoints. Biology, 11(4), 581. https://doi.org/10.3390/biology11040581