
Distribution Drivers of the Alien Butterfly Geranium Bronze (Cacyreus marshalli) in an Alpine Protected Area and Indications for an Effective Management

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Habitat and Climatic Variables
2.3. Cacyreus marshalli Data
2.3.1. Opportunistic Data
2.3.2. Standardised Sampling Data
2.4. Data Analysis
2.4.1. MaxEnt Model
2.4.2. N-Mixture Models
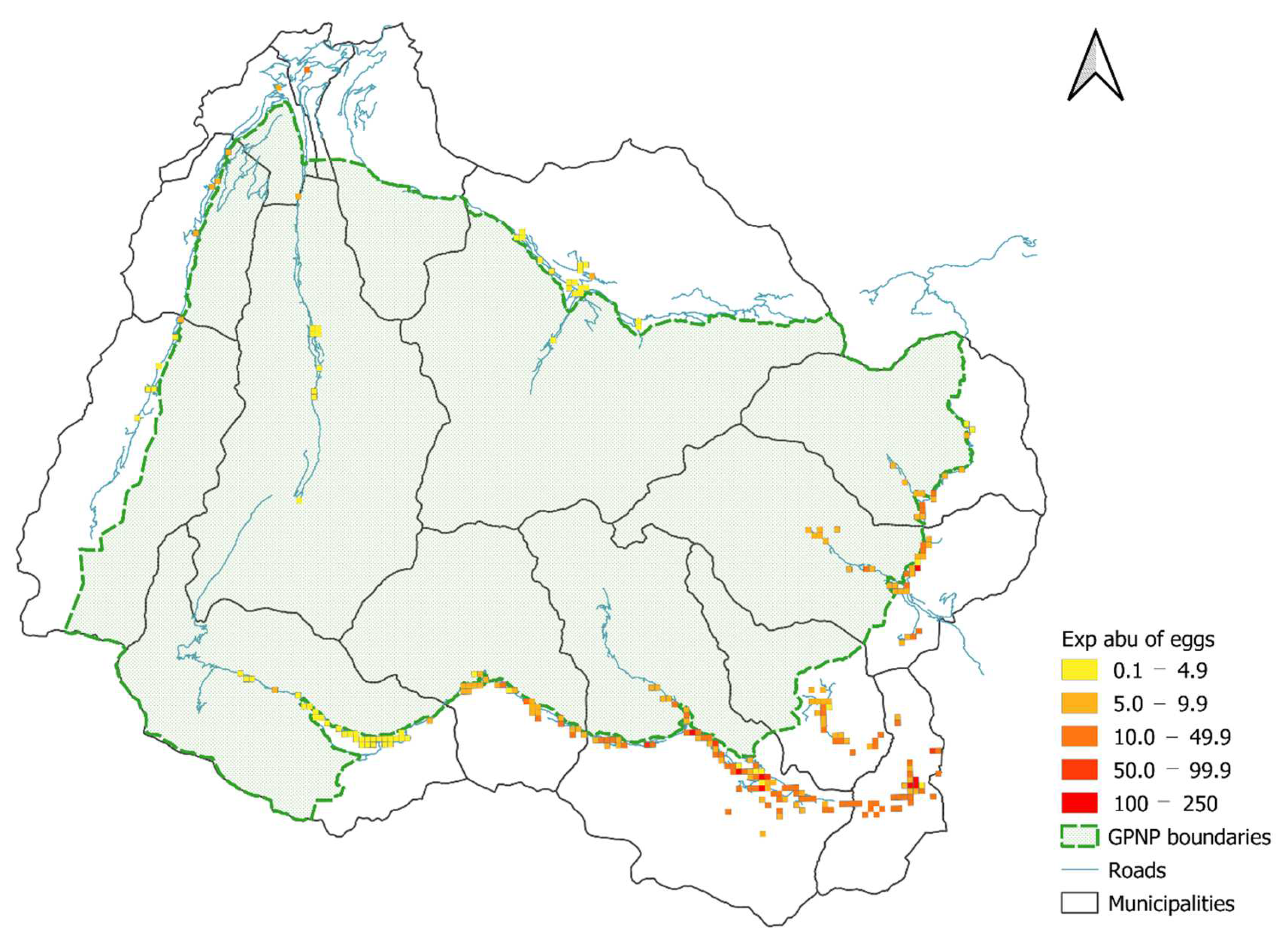
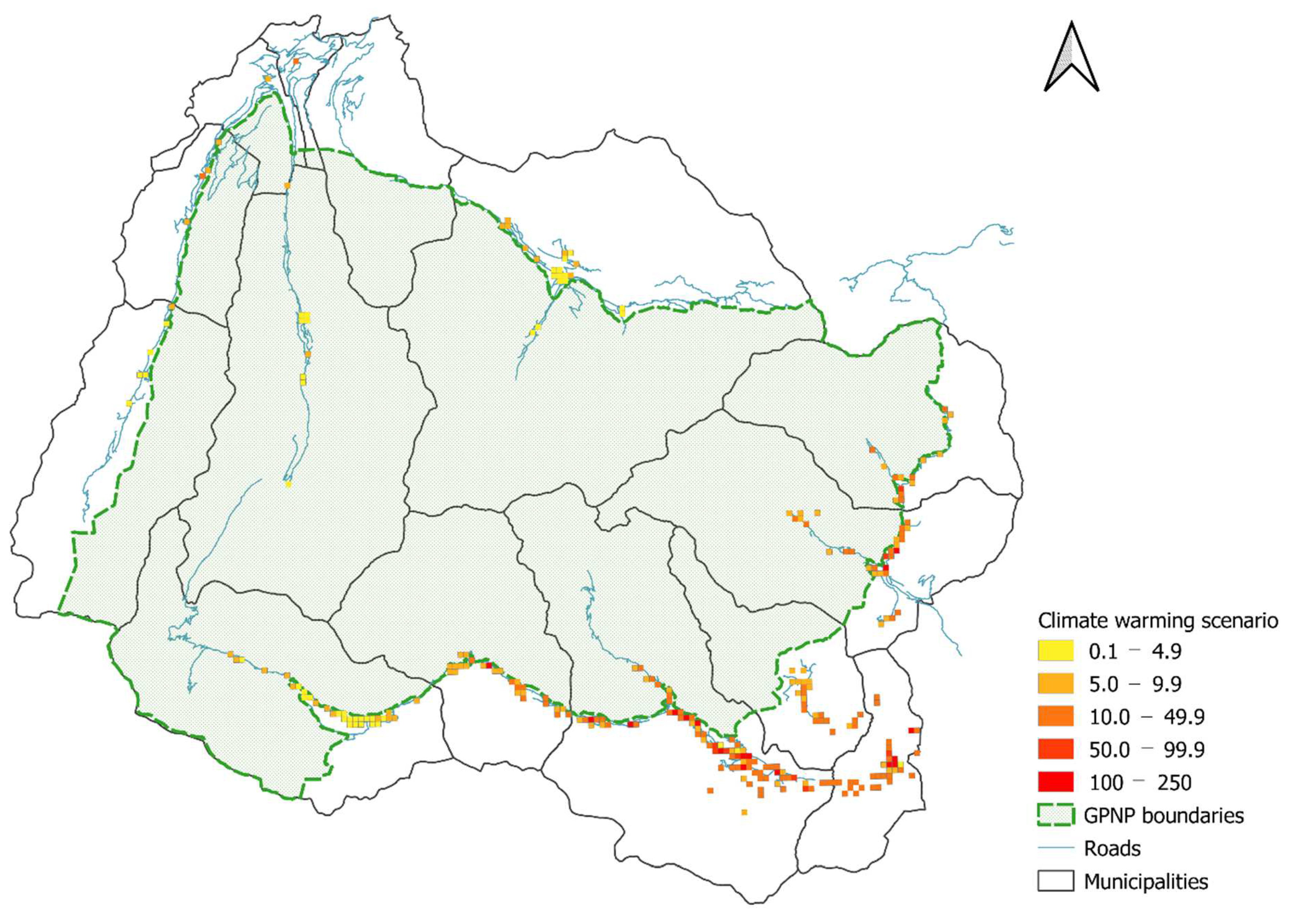
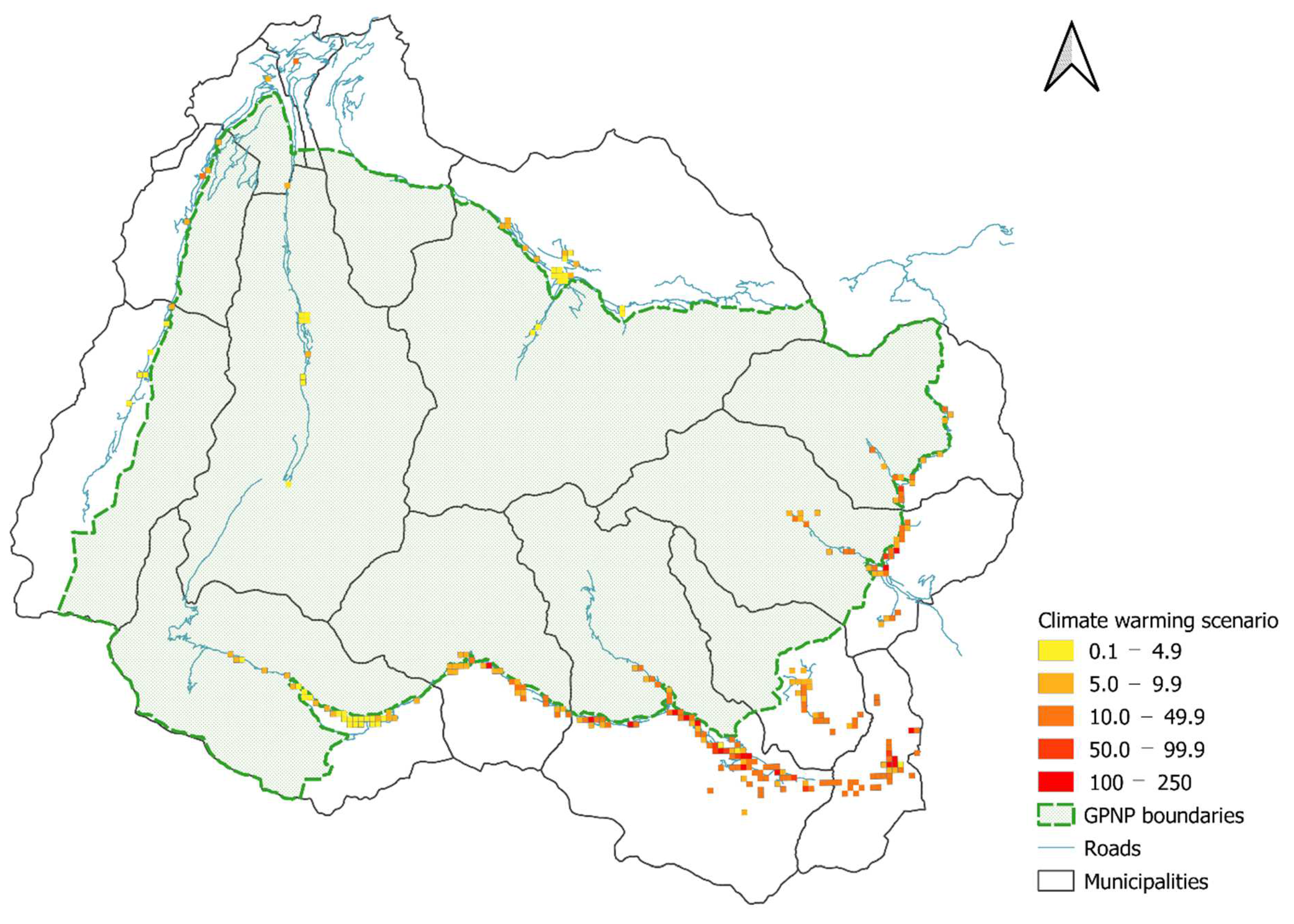
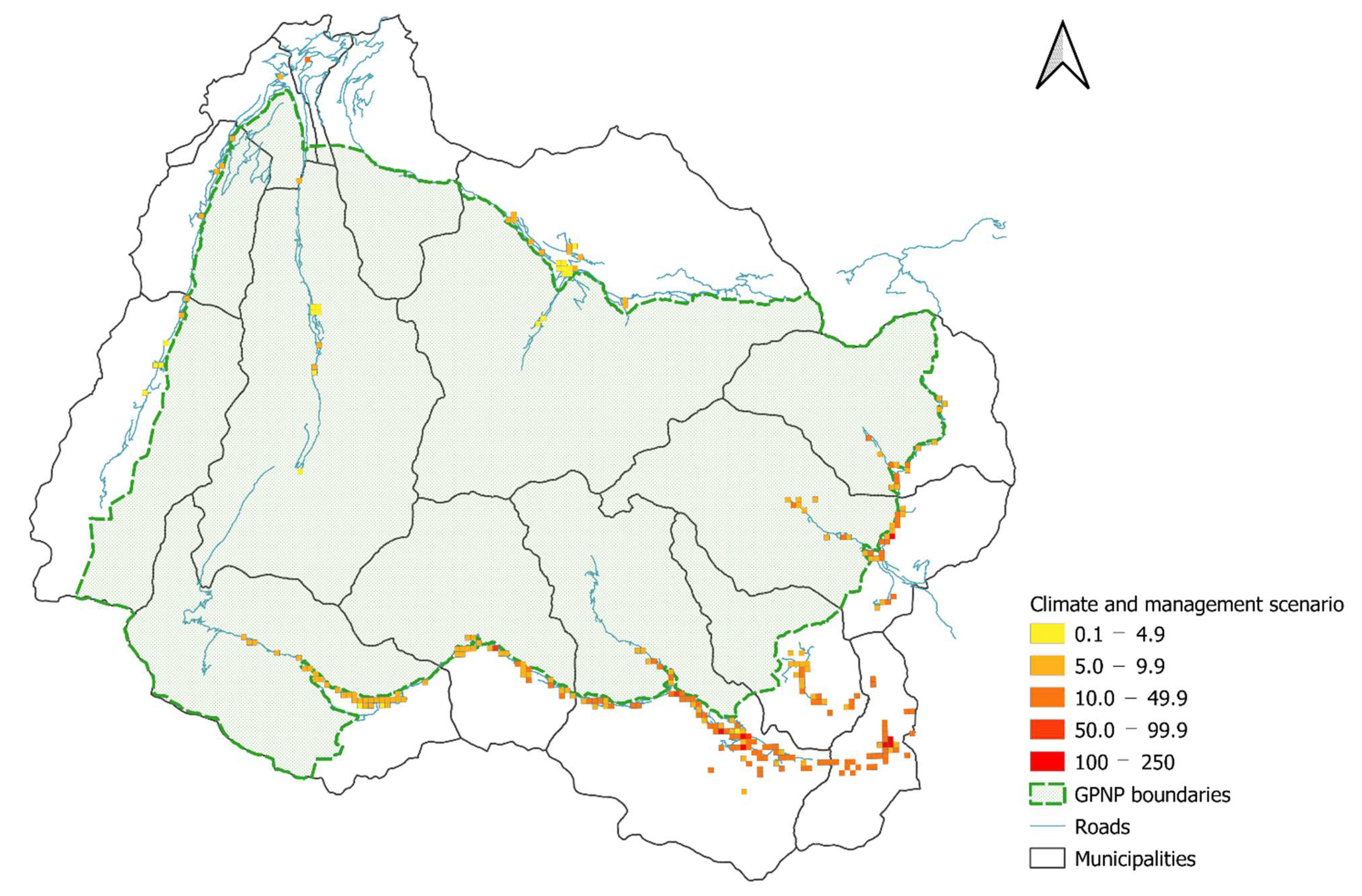
2.4.3. Distribution Maps and Scenarios
3. Results
3.1. Cacyreus marshalli Data
3.2. MaxEnt Model Results
3.3. N-Mixture Model Results
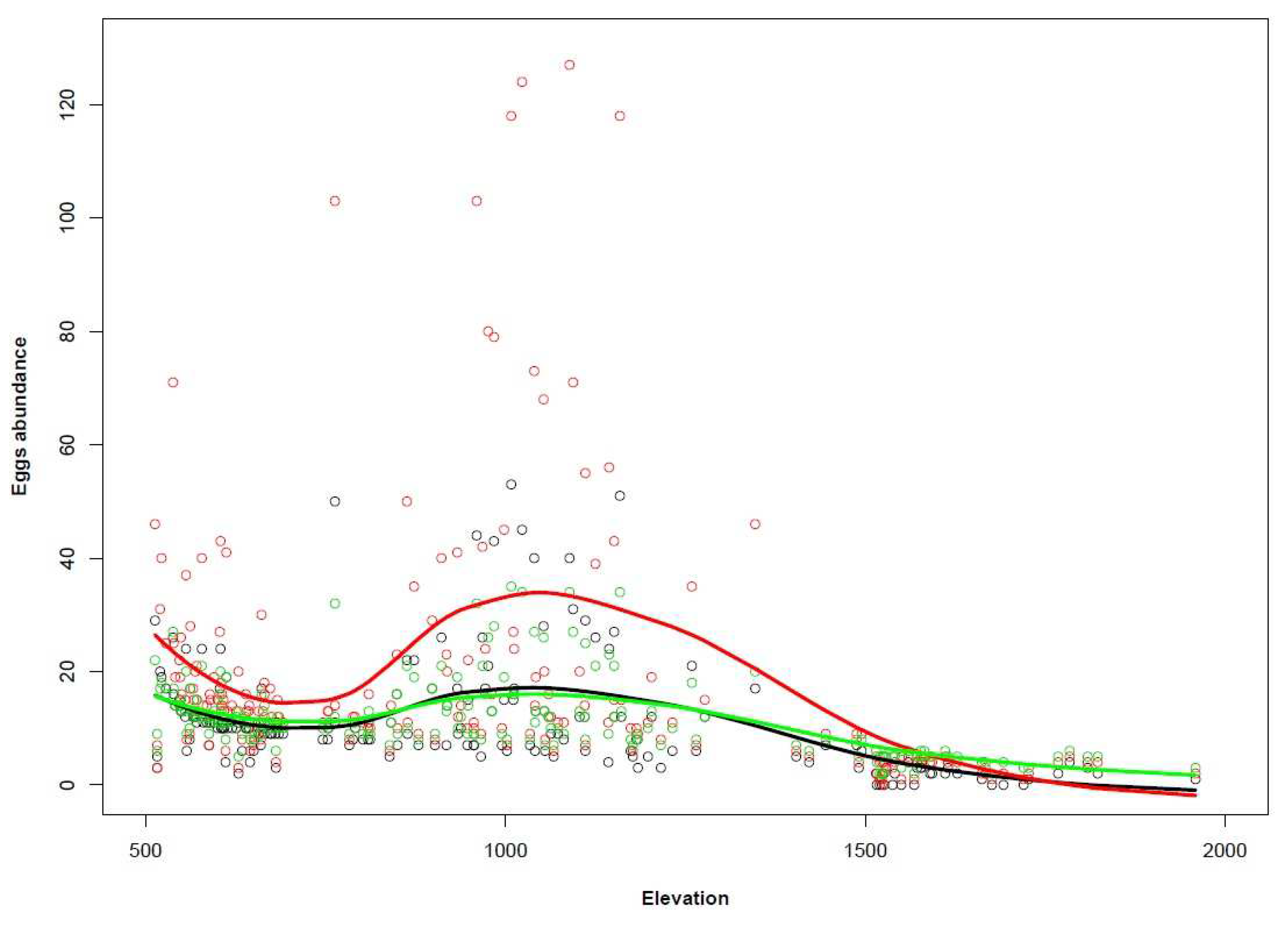
3.4. Distribution Maps and Scenario Results
4. Discussion
4.1. Opportunistic Data and Standardised Sampling
4.2. Drivers of Cacyreus marshalli Distribution
4.3. Distribution Maps, Future Scenarios and a Potential Management Strategy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walther, G.-R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukat, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Clark, G.C.; Dickson, C.G.C. Life Histories of the South African Lycaenid Butterflies; Purnell & Sons: Cape Town, South Africa, 1971. [Google Scholar]
- Dukes, J.S.; Mooney, H.A. Does global change increase the success of biological invaders? Trends Ecol. Evol. 1999, 14, 135–139. [Google Scholar] [CrossRef]
- Van Swaay, C.C.; Cuttelod, A.; Collins, S.; Maes, D.; Munguira, M.L.; Šašić, M.; Settele, J.; Verovnik, R.; Verstrael, T.; Warren, M.; et al. European Red List of Butterflies; Publications Office of the European Communities: Luxembourg, 2010. [Google Scholar]
- Sarto i Monteys, V. Spread of the southern African lycaenid butterfly, Cacyreus marshalli Butler, 1898, (Lep.: Lycaenidae) in the Balearic Archipelago (Spain) and considerations on its likely introduction to continental Europe. J. Res. Lepid. 1992, 31, 24–34. [Google Scholar]
- Quacchia, A.; Ferracini, C.; Bonelli, S.; Balletto, E.; Alma, A. Can the Geranium Bronze, Cacyreus marshalli, become a threat for European biodiversity? Biodivers. Conserv. 2008, 17, 1429–1437. [Google Scholar] [CrossRef]
- Federica, P.; Francesca, M.; Cristiana, C.; Silvia, G.; Ramona, V.; Sara, C.; Chiara, F.; Simona, B. From Africa to the Alps: Risk assessment on an invasion by Cacyreus marshalli (Butler, 1898). J. Insect Conserv. 2019, 23, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Gobiet, A.; Kotlarski, S.; Beniston, M.; Heinrich, G.; Rajczak, J.; Stoffel, M. 21st century climate change in the European Alps—A review. Sci. Total Environ. 2014, 493, 1138–1151. [Google Scholar] [CrossRef]
- Parolo, G.; Rossi, G. Upward migration of vascular plants following a climate warming trend in the Alps. Basic Appl. Ecol. 2008, 9, 100–107. [Google Scholar] [CrossRef]
- Brivio, F.; Zurmühl, M.; Grignolio, S.; von Hardenberg, J.; Apollonio, M.; Ciuti, S. Forecasting the response to global warming in a heat-sensitive species. Sci. Rep. 2019, 9, 3048. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, A.R.; Provenzale, A.; von Hardenberg, A.; Bassano, B.; Festa-Bianchet, M. Climate forcing and density dependence in a mountain ungulate population. Ecology 2004, 85, 1598–1610. [Google Scholar] [CrossRef] [Green Version]
- Rogora, M.; Frate, L.; Carranza, M.; Freppaz, M.; Stanisci, A.; Bertani, I.; Bottarin, R.; Brambilla, A.; Canullo, R.; Carbognani, M.; et al. Assessment of climate change effects on mountain ecosystems through a cross-site analysis in the Alps and Apennines. Sci. Total Environ. 2018, 624, 1429–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Benito Alonso, J.L.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, F.; et al. Continent-wide response of mountain vegetation to climate change. Nat. Clim. Chang. 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Fumy, F.; Löffler, F.; Samways, M.J.; Fartmann, T. Response of Orthoptera assemblages to environmental change in a low-mountain range differs among grassland types. J. Environ. Manag. 2020, 256, 109919. [Google Scholar] [CrossRef] [PubMed]
- Poniatowski, D.; Beckmann, C.; Löffler, F.; Münsch, T.; Helbing, F.; Samways, M.J.; Fartmann, T. Relative impacts of land-use and climate change on grasshopper range shifts have changed over time. Glob. Ecol. Biogeogr. 2020, 29, 2190–2202. [Google Scholar] [CrossRef]
- Cerrato, C.; Rocchia, E.; Brunetti, M.; Bionda, R.; Bassano, B.; Provenzale, A.; Bonelli, S.; Viterbi, R. Butterfly distribution along altitudinal gradients: Temporal changes over a short time period. Nat. Conserv. 2019, 34, 91–118. [Google Scholar] [CrossRef] [Green Version]
- Rödder, D.; Schmitt, T.; Gros, P.; Ulrich, W.; Habel, J.C. Climate change drives mountain butterflies towards the summits. Sci. Rep. 2021, 11, 14382. [Google Scholar] [CrossRef]
- Westwood, R.; Westwood, A.R.; Hooshmandi, M.; Pearson, K.; LaFrance, K.; Murray, C. A field-validated species distribution model to support management of the critically endangered Poweshiek skipperling (Oarisma poweshiek) butterfly in Canada. Conserv. Sci. Pract. 2020, 2, e163. [Google Scholar] [CrossRef] [Green Version]
- Casazza, G.; Abeli, T.; Bacchetta, G.; Dagnino, D.; Fenu, G.; Gargano, D.; Minuto, L.; Montagnani, C.; Orsenigo, S.; Peruzzi, L.; et al. Combining conservation status and species distribution models for planning assisted colonisation under climate change. J. Ecol. 2021, 109, 2284–2295. [Google Scholar] [CrossRef]
- Tiberti, R.; Nelli, L.; Brighenti, S.; Iacobuzio, R.; Rolla, M. Spatial distribution of introduced brook trout Salvelinus fontinalis (Salmonidae) within alpine lakes: Evidences from a fish eradication campaign. Eur. Zool. J. 2017, 84, 73–88. [Google Scholar] [CrossRef] [Green Version]
- Metz, M.; Rocchini, D.; Neteler, M. Surface temperatures at the continental scale: Tracking changes with remote sensing at unprecedented detail. Remote Sens. 2014, 6, 3822–3840. [Google Scholar] [CrossRef] [Green Version]
- Kučinić, M.; Koren, T.; Mihoci, I.; Vuković, M.; Bukovec, D.; Jakovljević, T.; Jenčić, S. Can spreading of the Geranium Bronze Cacyreus marshalli (Butler, 1898)(Insecta, Lepidoptera, Lycaenidae) in Croatia be assigned to climate change? Period. Biol. 2013, 115, 429–433. [Google Scholar]
- Ruffo, S. Checklist e Distribuzione della Fauna Italiana 10,000 Specie Terrestri e Delle Acque Interne; Memorie del Museo Civico di Storia Naturale di Verona: Verona, Italy, 2005; Volume 2. [Google Scholar]
- Balletto, E.; Bonelli, S.; Cassulo, L. Insecta Lepidoptera Papilionoidea. In Checklist and Distribution of the Italian Fauna 10,000 Terrestrial and Inland Water Species; Ruffo, S., Stoch, F., Eds.; Memorie del Museo Civico di Storia Naturale di Verona: Verona, Italy, 2007; Volume 17, pp. 257–261. [Google Scholar]
- Tarquini, S.; Nannipieri, L. The 10 m-resolution TINITALY DEM as a trans-disciplinary basis for the analysis of the Italian territory: Current trends and new perspectives. Geomorphology 2017, 281, 108–115. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F.; Georges, M.D.; Thuiller, C.W. Ensemble Platform for Species Distribution Modeling. Package ‘biomod2’. Species Distribution Modeling Within an Ensemble Forecasting Framework; version 3.3-7. Ecography 2016, 32, 369–373. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping species distributions with MAXENT using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef] [Green Version]
- Joseph, L.N.; Elkin, C.; Martin, T.G.; Possingham, H.P. Modeling abundance using N-mixture models: The importance of considering ecological mechanisms. Ecol. Appl. 2009, 19, 631–642. [Google Scholar] [CrossRef] [Green Version]
- Kéry, M.; Royle, J.A. Applied Hierarchical Modeling in Ecology: Analysis of Distribution, Abundance and Species Richness in R and BUGS; Volume 1: Prelude and Static Models; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Becker, H.; Leon, J. Stability analysis in plant breeding. Plant Breed. 1988, 101, 1–23. [Google Scholar] [CrossRef]
- Fiske, I.; Chandler, R. Unmarked: An R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Porro, Z.; Odicino, M.; Bogliani, G.; Chiatante, G. Intensive forestry and biodiversity: Use of poplar plantations by woodpeckers in a lowland area of Northern Italy. For. Ecol. Manag. 2021, 497, 119490. [Google Scholar] [CrossRef]
- Kroll, A.J.; Fleming, T.L.; Irwin, L.L. Site occupancy dynamics of northern spotted owls in the eastern Cascades, Washington, USA, 1990–2003. J. Wildl. Manag. 2010, 74, 1267–1274. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Wenger, S.J.; Freeman, M.C. Estimating species occurrence, abundance, and detection probability using zero-inflated distributions. Ecology 2008, 89, 2953–2959. [Google Scholar] [CrossRef] [PubMed]
- Kéry, M.; Royle, A.; Meredith, M. AHMbook: Functions and Data for the Book ‘Applied Hierarchical Modeling in Ecology’. R package version 01. 2017. Available online: https://CRAN.R-project.org/package=AHMbook (accessed on 1 September 2021).
- Mazerolle, M. AICcmodavg: Model selection and multimodel inference based on (Q) AIC (c). R package version 2.0–3 [computer program]. 2015. Available online: https://cran.r-project.org/package=AICcmodavg (accessed on 1 September 2021).
- Johnson, D.S.; Laake, J.L.; Ver Hoef, J.M. A model-based approach for making ecological inference from distance sampling data. Biometrics 2010, 66, 310–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kéry, M.; Royle, J.A.; Schmid, H. Modeling avian abundance from replicated counts using binomial mixture models. Ecol. Appl. 2005, 15, 1450–1461. [Google Scholar] [CrossRef]
- Ver Hoef, J.M.; Boveng, P.L. Quasi-Poisson vs. negative binomial regression: How should we model overdispersed count data? Ecology 2007, 88, 2766–2772. [Google Scholar] [CrossRef] [Green Version]
- Rolland, C. Spatial and seasonal variations of air temperature lapse rates in Alpine regions. J. Clim. 2003, 16, 1032–1046. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, J.L.; Zuckerberg, B.; Bonter, D.N. Citizen science as an ecological research tool: Challenges and benefits. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 149–172. [Google Scholar] [CrossRef] [Green Version]
- McKinley, D.C.; Miller-Rushing, A.J.; Ballard, H.L.; Bonney, R.; Brown, H.; Cook-Patton, S.C.; Evans, D.M.; French, R.A.; Parrish, J.K.; Phillips, T.B.; et al. Citizen science can improve conservation science, natural resource management, and environmental protection. Biol. Conserv. 2017, 208, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Martin, V.Y. Citizen science as a means for increasing public engagement in science: Presumption or possibility? Sci. Commun. 2017, 39, 142–168. [Google Scholar] [CrossRef]
- Feldman, M.J.; Imbeau, L.; Marchand, P.; Mazerolle, M.J.; Darveau, M.; Fenton, N.J. Trends and gaps in the use of citizen science derived data as input for species distribution models: A quantitative review. PLoS ONE 2021, 16, e0234587. [Google Scholar]
- Lewandowski, E.J.; Oberhauser, K.S. Butterfly citizen scientists in the United States increase their engagement in conservation. Biol. Conserv. 2017, 208, 106–112. [Google Scholar] [CrossRef]
- Dickinson, J.L.; Shirk, J.; Bonter, D.; Bonney, R.; Crain, R.L.; Martin, J.; Phillips, T.; Purcell, K. The current state of citizen science as a tool for ecological research and public engagement. Front. Ecol. Environ. 2012, 10, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Pacifici, K.; Reich, B.J.; Miller, D.A.; Gardner, B.; Stauffer, G.; Singh, S.; McKerrow, A.; Collazo, J.A. Integrating multiple data sources in species distribution modeling: A framework for data fusion. Ecology 2017, 98, 840–850. [Google Scholar] [CrossRef] [Green Version]
- Dorazio, R.M. Accounting for imperfect detection and survey bias in statistical analysis of presence-only data. Glob. Ecol. Biogeogr. 2014, 23, 1472–1484. [Google Scholar] [CrossRef]
- Manica, M.; Caputo, B.; Screti, A.; Filipponi, F.; Rosà, R.; Solimini, A.; della Torre, A.; Blangiardo, M. Applying the N-mixture model approach to estimate mosquito population absolute abundance from monitoring data. J. Appl. Ecol. 2019, 56, 2225–2235. [Google Scholar] [CrossRef]
- Jarošík, V.; Kenis, M.; Honěk, A.; Skuhrovec, J.; Pyšek, P. Invasive insects differ from non-invasive in their thermal requirements. PLoS ONE 2015, 10, e0131072. [Google Scholar] [CrossRef]
- Essl, F.; Lenzner, B.; Bacher, S.; Bailey, S.; Capinha, C.; Daehler, C.; Dullinger, S.; Genovesi, P.; Hui, C.; Hulme, P.E.; et al. Drivers of future alien species impacts: An expert-based assessment. Glob. Chang. Biol. 2020, 26, 4880–4893. [Google Scholar] [CrossRef]
- Huang, D.; Haack, R.A.; Zhang, R. Does global warming increase establishment rates of invasive alien species? A centurial time series analysis. PLoS ONE 2011, 6, e24733. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Full Model | Test Set Model | ||
|---|---|---|---|---|
| Percent Contribution | Permutation Importance | Percent Contribution | Permutation Importance | |
| pel_abu | 90.7 | 70.7 | 88.1 | 74.8 |
| pel_neigh | 7.2 | 6.3 | 9.7 | 8.6 |
| bio04 | 1.9 | 22.7 | 1.8 | 15.7 |
| bio01 | 0.1 | 0.3 | 0.4 | 0.8 |
| Models | K | AICc | ΔAICc | Wi | Cum.Wi |
|---|---|---|---|---|---|
| ρ pel_ava λ bio01 + pel_abu + pel_abu:bio01 + pel_neigh | 7 | 979.54 | 0.00 | 1 | 1 |
| ρ pel_ava λ bio01 + pel_abu + pel_neigh | 6 | 1099.16 | 119.61 | <0.01 | 1 |
| ρ pel_ava λ bio01 + pel_abu | 5 | 1102.05 | 122.51 | <0.01 | 1 |
| ρ pel_ava λ bio01 + pel_abu + wood | 6 | 1102.58 | 123.04 | <0.01 | 1 |
| ρ pel_ava λ bio01 | 4 | 1135.79 | 156.24 | <0.01 | 1 |
| ρ pel_ava λ bio01 + pel_neigh | 5 | 1136.25 | 156.70 | <0.01 | 1 |
| ρ pel_ava λ bio01 + wood | 5 | 1138.09 | 158.54 | <0.01 | 1 |
| ρ pel_ava λ bio04 | 4 | 1186.08 | 206.53 | <0.01 | 1 |
| ρ pel_ava λ pel_neigh | 4 | 1273.59 | 294.05 | <0.01 | 1 |
| ρ pel_ava λ wood | 4 | 1275.42 | 295.88 | <0.01 | 1 |
| ρ pel_ava λ eco | 4 | 1281.87 | 302.33 | <0.01 | 1 |
| ρ pel_ava λ eco | 4 | 1282.99 | 303.45 | <0.01 | 1 |
| ρ pel_ava λ. | 3 | 1286.70 | 307.15 | <0.01 | 1 |
| ρ pel_ava λ grassland | 4 | 1287.91 | 308.36 | <0.01 | 1 |
| ρ. λ. | 2 | 2375.88 | 1396.33 | <0.01 | 1 |
| Variables | Egg Abundance (SE) | Detectability (SE) |
|---|---|---|
| Intercept | 3.095 (0.249) ** | −2.632 (0.182) ** |
| bio01 | 0.446 (0.209) * | |
| pel_abu | 0.320 (0.058) ** | |
| pel_neigh | −0.343 (0.067) ** | |
| pel_abu:bio01 | 0.592 (0.119) ** | |
| pel_ava | 0.399 (0.049) ** |
| Change Categories | Starting Distribution vs. +1.5 °C Scenario | +1.5 °C Scenario vs. −50% Pelargonium Scenario |
|---|---|---|
| Percentage of cells with egg abundance changes | 98.1 (264) | 74 (199) |
| Percentage of cells with egg increases | 98.1 (264) | 22.7 (61) |
| Percentage of cells with egg decreases | 0 | 51.3 (138) |
| Percentage of cells with new egg infestations | 3.3 (9) | 0 |
| Percentage of cell with no changes | 5.2 (14) | 29.7 (80) |
| Municipality | Elevation | Percentage of Cells with Egg Decreases | Percentage of Cells with Egg Increases |
|---|---|---|---|
| Sparone | 614 | 72.7 (24) | 9.1 (3) |
| Locana | 714 | 70.5 (55) | 15.4 (12) |
| Villeneuve | 966 | 100 (1) | 0 |
| Ronco Canavese | 1050 | 56.7 (17) | 3.3 (1) |
| Introd | 1063 | 0 | 0 |
| Noasca | 1071 | 53.8 (14) | 11.5 (3) |
| Ribordone | 1127 | 52.2 (12) | 8.7 (2) |
| Rhêmes-Saint-Georges | 1249 | 75 (3) | 0 |
| Valprato Soana | 1268 | 66.7(10) | 13.3 (2) |
| Ceresole Reale | 1581 | 0 | 40.6 (13) |
| Cogne | 1586 | 9.5 (2) | 66.7 (14) |
| Valsavarenche | 1635 | 0 | 75 (6) |
| Rhêmes-Notre-Dame | 1699 | 0 | 83.3 (5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocchia, E.; Luppi, M.; Paradiso, F.; Ghidotti, S.; Martelli, F.; Cerrato, C.; Viterbi, R.; Bonelli, S. Distribution Drivers of the Alien Butterfly Geranium Bronze (Cacyreus marshalli) in an Alpine Protected Area and Indications for an Effective Management. Biology 2022, 11, 563. https://doi.org/10.3390/biology11040563
Rocchia E, Luppi M, Paradiso F, Ghidotti S, Martelli F, Cerrato C, Viterbi R, Bonelli S. Distribution Drivers of the Alien Butterfly Geranium Bronze (Cacyreus marshalli) in an Alpine Protected Area and Indications for an Effective Management. Biology. 2022; 11(4):563. https://doi.org/10.3390/biology11040563
Chicago/Turabian StyleRocchia, Emanuel, Massimiliano Luppi, Federica Paradiso, Silvia Ghidotti, Francesca Martelli, Cristiana Cerrato, Ramona Viterbi, and Simona Bonelli. 2022. "Distribution Drivers of the Alien Butterfly Geranium Bronze (Cacyreus marshalli) in an Alpine Protected Area and Indications for an Effective Management" Biology 11, no. 4: 563. https://doi.org/10.3390/biology11040563
APA StyleRocchia, E., Luppi, M., Paradiso, F., Ghidotti, S., Martelli, F., Cerrato, C., Viterbi, R., & Bonelli, S. (2022). Distribution Drivers of the Alien Butterfly Geranium Bronze (Cacyreus marshalli) in an Alpine Protected Area and Indications for an Effective Management. Biology, 11(4), 563. https://doi.org/10.3390/biology11040563