
From Water to Land: The Structural Construction and Molecular Switches in Lungs during Metamorphosis of Microhyla fissipes

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Daily Culture
2.2. Experimental Design and Sampling
2.3. Micro-CT
2.4. Histological Analysis
2.5. Transmission Electron Microscopic (TEM) Observation
2.6. Transcriptomic Analyses
2.7. Weighted Correlation Network Analysis (WGCNA)
3. Results
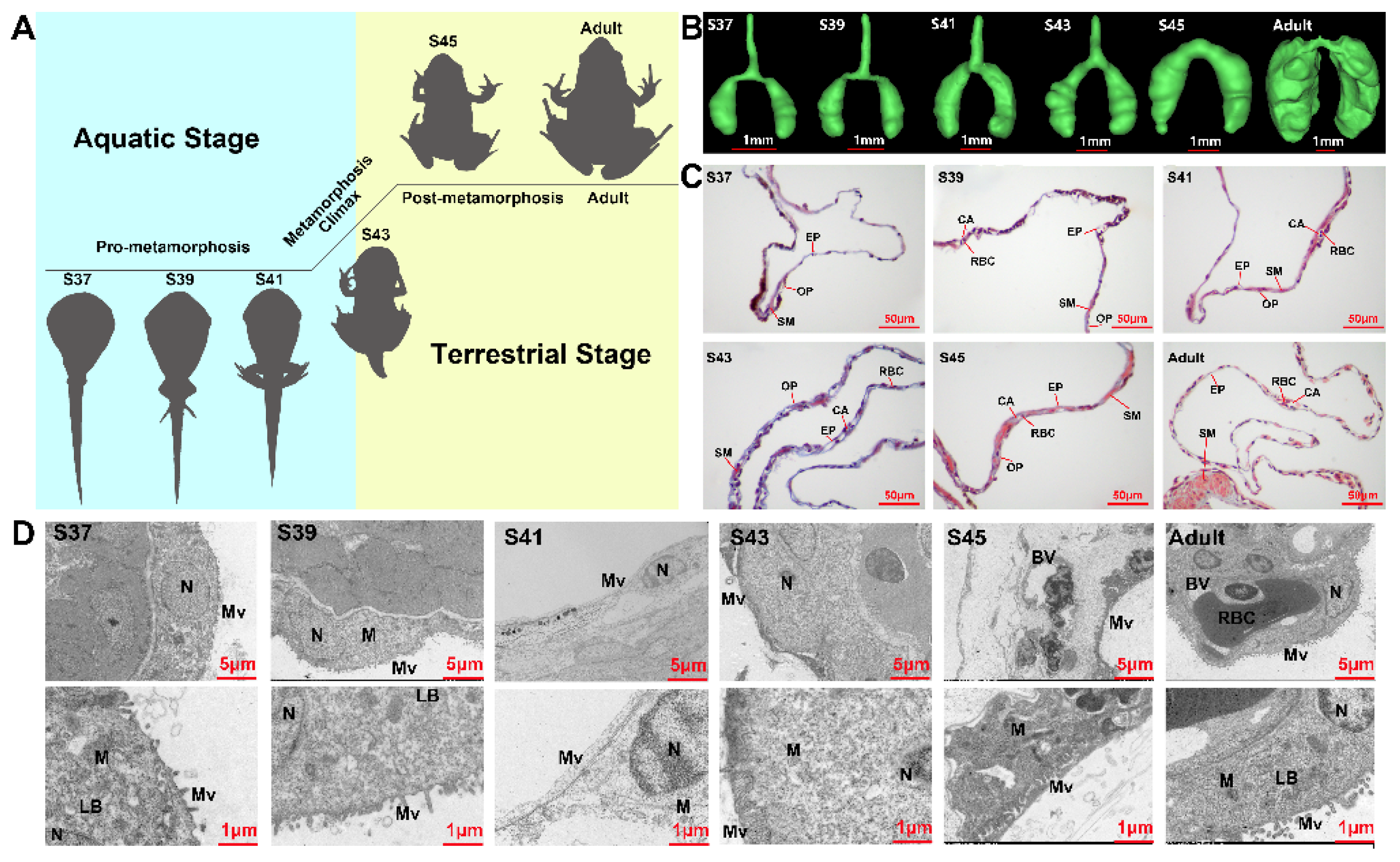
3.1. Structure Observations
3.2. Functional Analysis of Co-Expressed Modules
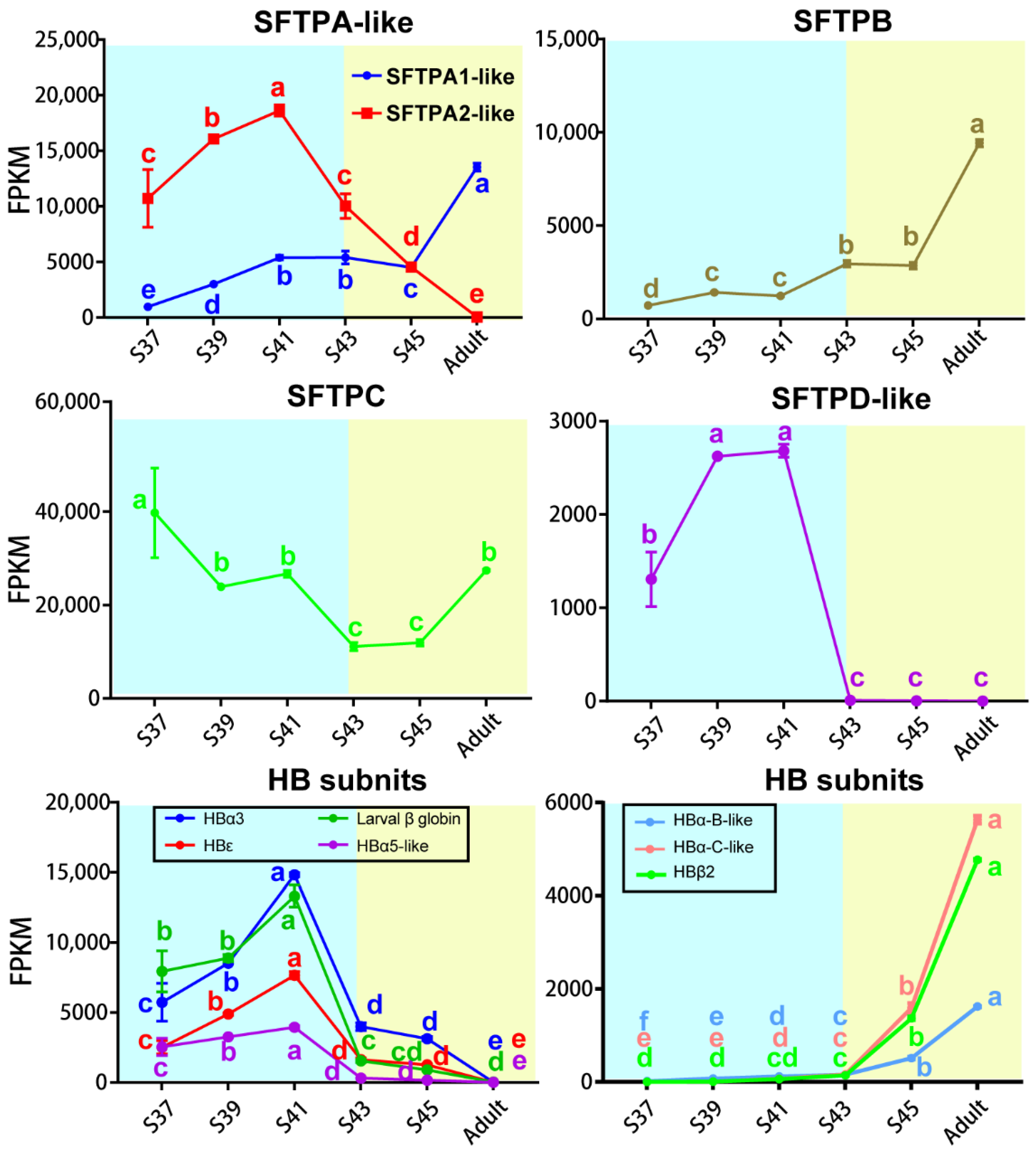
3.3. Transcriptional Switches of Respiration Functional Proteins
4. Discussion
4.1. Rapid Growth and Robust Substrate Metabolism of Lungs during the Pro-Metamorphic Phase
4.2. Construction of Extracellular Matrix and Accelerated Development of Lungs during the Metamorphic Phase
4.3. Reinforcing of Muscle Function in Post-Metamorphic Lungs
4.4. Transcriptional Switches of Respiration Functional Proteins during the Metamorphic Climax Phase
4.5. Completely Functional Maturation in Adult Lungs
4.6. Evolutionary Inspiration from Development of M. fissipes Lung
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burggren, W.W.; Infantino, J.R.L. The Respiratory Transition from Water to Air Breathing During Amphibian Metamorphosis1. Am. Zool. 1994, 34, 238–246. [Google Scholar] [CrossRef]
- Boatright-Horowitz, S.S.; Simmons, A.M. Transient “deafness” accompanies auditory development during metamorphosis from tadpole to frog. Proc. Natl. Acad. Sci. USA 1997, 94, 14877–14882. [Google Scholar] [CrossRef] [PubMed]
- Frieden, E.; Just, J.J. Hormonal responses in amphibian metamorphosis. Biochem. Actions Horm. 1970, 1, 1–52. [Google Scholar]
- Atkinson, B.G. Patterns of macromolecular biosynthesis during amphibian metamorphosis. In Proceedings of the 7th Conference on Endocrinology and Metabolism; University of Missouri Press Columbia: Missouri, MO, USA, 1971; pp. 48–82. [Google Scholar]
- Furlow, J.D.; Neff, E.S. A developmental switch induced by thyroid hormone: Xenopus laevis metamorphosis. Trends Endocrinol. Metab. 2006, 17, 38–45. [Google Scholar] [CrossRef]
- Brown, D.D.; Cai, L. Amphibian metamorphosis. Dev. Biol. 2007, 306, 20–33. [Google Scholar] [CrossRef]
- Wang, S.; Liu, L.; Liu, J.; Zhu, W.; Tanizaki, Y.; Fu, L.; Bao, L.; Shi, Y.; Jiang, J. Gene Expression Program Underlying Tail Resorption During Thyroid Hormone-Dependent Metamorphosis of the Ornamented Pygmy Frog Microhyla fissipes. Front. Endocrinol. 2019, 10, 11. [Google Scholar] [CrossRef]
- Shi, Y.; Ishizuya-Oka, A. Thyroid hormone regulation of apoptotic tissue remodeling: Implications from molecular analysis of amphibian metamorphosis. In Progress in Nucleic Acid Research and Molecular Biology; Moldave, K., Ed.; Elsevier: Amsterdam, The Netherlands, 2001; Volume 65, pp. 53–100. [Google Scholar]
- Burggren, W.W.; Warburton, S. Amphibians as animal models for laboratory research in physiology. ILAR J. 2007, 48, 260–269. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, L.; Wang, S.; Jiang, J. Research proceedings on amphibian model organisms. Zool. Res. 2016, 37, 237–245. [Google Scholar]
- Piiper, J.; Scheid, P. Gas Exchange in Vertebrates Through Lungs, Gills, and Skin. Physiology 1992, 7, 199–203. [Google Scholar] [CrossRef]
- Burggren, W.W. Transition of respiratory processes during amphibian metamorphosis: From egg to adult. In Proceedings of the Respiration and Metabolism of Embryonic Vertebrates; Springer: Dordrecht, The Netherlands, 1984; pp. 31–53. [Google Scholar]
- Wake, M.H. Biology of Amphibians; American Scientist: New York, NY, USA, 1986. [Google Scholar]
- Rollins-Smith, L.A.; Woodhams, D.C. Amphibian Immunity; Oxford University Press New York: New York, NY, USA, 2012. [Google Scholar]
- Menon, J.; Rozman, R. Oxidative stress, tissue remodeling and regression during amphibian metamorphosis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 145, 625–631. [Google Scholar] [CrossRef]
- Prokić, M.D.; Gavrić, J.P.; Petrović, T.G.; Despotović, S.G.; Gavrilović, B.R.; Radovanović, T.B.; Krizmanić, I.I.; Pavlović, S.Z. Oxidative stress in Pelophylax esculentus complex frogs in the wild during transition from aquatic to terrestrial life. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 234, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Weaver, T.E.; Whitsett, J.A. Function and regulation of epression of pulmonary surfactant-associated proteins. Biochem. J. 1991, 273, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Antonini, E.; Brunori, M. Hemoglobin. Annu. Rev. Biochem. 1970, 39, 977–1042. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Ishiko, S.; Daido, S.; Kim, J.; Ikeda, S. Comparative morphology of the lung with special reference to the alveolar epithelial cells. I. Lung of the amphibia. Acta Tuberc. Jpn. 1962, 11, 63–72. [Google Scholar]
- Rankin, S.A.; Hong Thi, T.; Wlizla, M.; Mancini, P.; Shifley, E.T.; Bloor, S.D.; Han, L.; Vleminckx, K.; Wert, S.E.; Zorn, A.M. A Molecular Atlas of Xenopus Respiratory System Development. Dev. Dyn. 2015, 244, 69–85. [Google Scholar] [CrossRef]
- Meban, C. Pneumonocytes in lung of Xenopus laevis. J. Anat. 1973, 114, 235–244. [Google Scholar]
- Fei, L.; Ye, C.; Jiang, J. Colored Atlas of Chinese Amphibians and Their Distributions; Sichuan Publishing House of Science & Technology: Chengdu, China, 2012. [Google Scholar]
- Wang, S.; Zhao, L.; Liu, L.; Yang, D.; Khatiwada, J.R.; Wang, B.; Jiang, J. A Complete Embryonic Developmental Table of Microhyla fissipes (Amphibia, Anura, Microhylidae). Asian Herpetol. Res. 2017, 8, 108–117. [Google Scholar]
- Chang, L.; Wang, B.; Zhang, M.; Liu, J.; Zhao, T.; Zhu, W.; Jiang, J. The effects of corticosterone and background colour on tadpole physiological plasticity. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 39, 100872. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 8. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, A.J.S. Protein Turnover: A Functional Appraisal. Funct. Ecol. 1991, 5, 222–233. [Google Scholar] [CrossRef]
- Houlihan, D.; Hall, S.; Gray, C.; Noble, B. Growth Rates and Protein Turnover in Atlantic Cod, Gadus morhua. Can. J. Fish. Aquat. Sci. Can. J. Fish. Aquat. Sci. 1988, 45, 951–964. [Google Scholar] [CrossRef]
- Zhou, Y.; Horowitz, J.C.; Naba, A.; Ambalavanan, N.; Atabai, K.; Balestrini, J.; Bitterman, P.B.; Corley, R.A.; Ding, B.S.; Engler, A.J.; et al. Extracellular matrix in lung development, homeostasis and disease. Matrix Biol. J. Int. Soc. Matrix Biol. 2018, 73, 77–104. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Hynes, R.O. The Extracellular Matrix: Not Just Pretty Fibrils. Science 2009, 326, 1216–1219. [Google Scholar] [CrossRef]
- Gross, J.; Lapiere, C.M. Collagenolytic activity in amphibian tissues-a tissue culture assay. Proc. Natl. Acad. Sci. USA 1962, 48, 1014. [Google Scholar] [CrossRef]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular matrix degradation and remodeling in development and disease. Cold Spring Harb. Perspect. Biol. 2011, 3, 5058. [Google Scholar] [CrossRef]
- Liu, S.; Young, S.M.; Varisco, B.M. Dynamic expression of chymotrypsin-like elastase 1 over the course of murine lung development. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L1104–L1116. [Google Scholar] [CrossRef]
- Mammoto, T.; Jiang, E.; Jiang, A.; Mammoto, A. Extracellular Matrix Structure and Tissue Stiffness Control Postnatal Lung Development through the Lipoprotein Receptor-Related Protein 5/Tie2 Signaling System. Am. J. Respir. Cell Mol. Biol. 2013, 49, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Cieri, R.L. Pulmonary Smooth Muscle in Vertebrates: A Comparative Review of Structure and Function. Integr. Comp. Biol. 2019, 59, 10–28. [Google Scholar] [CrossRef] [PubMed]
- Pollard, T.D.; Goldman, R.D. Overview of the Cytoskeleton from an Evolutionary Perspective. Cold Spring Harb. Perspect. Biol. 2018, 10, 30288. [Google Scholar] [CrossRef] [PubMed]
- King, R.J. Pulmonary surfactant. J. Appl. Physiol. 1982, 53, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Goerke, J. Pulmonary surfactant: Functions and molecular composition. Biochim. Biophys. Acta Mol. Basis Dis. 1998, 1408, 79–89. [Google Scholar] [CrossRef]
- Chroneos, Z.C.; Sever-Chroneos, Z.; Shepherd, V.L. Pulmonary Surfactant: An Immunological Perspective. Cell. Physiol. Biochem. 2010, 25, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Longo, M.L.; Bisagno, A.M.; Zasadzinski, J.A.N.; Bruni, R.; Waring, A.J. A Function of Lung Surfactant Protein SP-B. Science 1993, 261, 453–456. [Google Scholar] [CrossRef]
- Bourbon, J.R. Pulmonary Surfactant: Biochemical, Functional, Regulatory, and Clinical Concepts; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Weaver, T.E.; Conkright, J.J. Function of Surfactant Proteins B and C. Annu. Rev. Physiol. 2001, 63, 555–578. [Google Scholar] [CrossRef]
- Wright, J.R. Immunoregulatory functions of surfactant proteins. Nat. Rev. Immunol. 2005, 5, 58–68. [Google Scholar] [CrossRef]
- Silveyra, P.; Floros, J. Genetic complexity of the human surfactant-associated proteins SP-A1 and SP-A2. Gene 2013, 531, 126–132. [Google Scholar] [CrossRef]
- Huang, W.; Wang, G.; Phelps, D.S.; Al-Mondhiry, H.; Floros, J. Human SP-A genetic variants and bleomycin-induced cytokine production by THP-1 cells: Effect of ozone-induced SP-A oxidation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 286, L546–L553. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Umstead, T.M.; Phelps, D.S.; Al-Mondhiry, H.; Floros, J. The effect of ozone exposure on the ability of human surfactant protein a variants to stimulate cytokine production. Environ. Health Perspect. 2002, 110, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Bates-Kenney, S.R.; Tao, J.; Phelps, D.S.; Floros, J. Differences in Biochemical Properties and in Biological Function between Human SP-A1 and SP-A2 Variants, and the Impact of Ozone-Induced Oxidation. Biochemistry 2004, 43, 4227–4239. [Google Scholar] [CrossRef] [PubMed]
- Mikerov, A.N.; Wang, G.; Umstead, T.M.; Zacharatos, M.; Thomas, N.J.; Phelps, D.S.; Floros, J. Surfactant Protein A2 (SP-A2) Variants Expressed in CHO Cells Stimulate Phagocytosis of Pseudomonas aeruginosa More than Do SP-A1 Variants. Infect. Immun. 2007, 75, 1403–1412. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mikerov, A.N.; Umstead, T.M.; Gan, X.; Huang, W.; Guo, X.; Wang, G.; Phelps, D.S.; Floros, J. Impact of ozone exposure on the phagocytic activity of human surfactant protein A (SP-A) and SP-A variants. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 294, L121–L130. [Google Scholar] [CrossRef]
- Hellsten, U.; Harland, R.M.; Gilchrist, M.J.; Hendrix, D.; Jurka, J.; Kapitonov, V.; Ovcharenko, I.; Putnam, N.H.; Shu, S.Q.; Taher, L.; et al. The Genome of the Western Clawed Frog Xenopus tropicalis. Science 2010, 328, 633–636. [Google Scholar] [CrossRef]
- Hillier, L.W.; Miller, W.; Birney, E.; Warren, W.; Hardison, R.C.; Ponting, C.P.; Bork, P.; Burt, D.W.; Groenen, M.A.M.; Delany, M.E.; et al. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef]
- Feng, S.; Stiller, J.; Deng, Y.; Armstrong, J.; Fang, Q.; Reeve, A.H.; Xie, D.; Chen, G.; Guo, C.; Faircloth, B.C.; et al. Dense sampling of bird diversity increases power of comparative genomics. Nature 2020, 587, 252–257. [Google Scholar] [CrossRef]
- Wang, K.; Wang, J.; Zhu, C.L.; Yang, L.D.; Ren, Y.D.; Ruan, J.; Fan, G.Y.; Hu, J.; Xu, W.J.; Bi, X.P.; et al. African lungfish genome sheds light on the vertebrate water-to-land transition. Cell 2021, 184, 1362. [Google Scholar] [CrossRef]
- Hawgood, S.; Derrick, M.; Poulain, F. Structure and properties of surfactant protein B. Biochim. Biophys. Acta Mol. Basis Dis. 1998, 1408, 150–160. [Google Scholar] [CrossRef]
- Cochrane, C.G.; Revak, S.D. Pulmonary Surfactant Protein B (SP-B): Structure-Function Relationships. Science 1991, 254, 566–568. [Google Scholar] [CrossRef] [PubMed]
- Pryhuber, G.S. Regulation and Function of Pulmonary Surfactant Protein B. Mol. Genet. Metab. 1998, 64, 217–228. [Google Scholar] [CrossRef]
- Galla, H.J.; Bourdos, N.; von Nahmen, A.; Amrein, M.; Sieber, M. The role of pulmonary surfactant protein C during the breathing cycle. Thin Solid Film. 1998, 327–329, 632–635. [Google Scholar] [CrossRef]
- Arıkan-Ayyıldız, Z.; Caglayan-Sozmen, S.; Isık, S.; Deterding, R.; Dishop, M.K.; Couderc, R.; Epaud, R.; Louha, M.; Uzuner, N. Survival of an infant with homozygous surfactant protein C (SFTPC) mutation. Pediatric Pulmonol. 2014, 49, E112–E115. [Google Scholar] [CrossRef] [PubMed]
- Leth-Larsen, R.; Garred, P.; Jensenius, H.; Meschi, J.; Hartshorn, K.; Madsen, J.; Tornoe, I.; Madsen, H.O.; Sørensen, G.; Crouch, E.; et al. A Common Polymorphism in the SFTPD Gene Influences Assembly, Function, and Concentration of Surfactant Protein D. J. Immunol. 2005, 174, 1532. [Google Scholar] [CrossRef]
- Hardison, R.C. Evolution of Hemoglobin and Its Genes. Cold Spring Harb. Perspect. Med. 2012, 2, 11627. [Google Scholar] [CrossRef] [PubMed]
- Maniatis, T.; Fritsch, E.F.; Lauer, J.; Lawn, R.M. The molecular genetics of human hemoglobins. Annu. Rev. Genet. 1980, 14, 145–178. [Google Scholar] [CrossRef]
- Karlsson, S.; Nienhuis, A.W. Developmental regulation of human globin genes. Annu. Rev. Biochem. 1985, 54, 1071–1108. [Google Scholar] [CrossRef]
- Perutz, M.F. Hemoglobin Structure and Respiratory Transport. Sci. Am. 1978, 239, 92–125. [Google Scholar] [CrossRef]
- Bauer, D.E.; Kamran, S.C.; Orkin, S.H. Reawakening fetal hemoglobin: Prospects for new therapies for the β-globin disorders. Blood 2012, 120, 2945–2953. [Google Scholar] [CrossRef]
- Weber, R.; Blum, B.; Muller, P.R. The switch from larval to adult globin gene expression in Xenopus laevis is mediated by erythroid cells from distinct compartments. Development 1991, 112, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Mukhi, S.; Cai, L.Q.; Brown, D.D. Gene switching at Xenopus laevis metamorphosis. Dev. Biol. 2010, 338, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Session, A.M.; Uno, Y.; Kwon, T.; Hapman, J.A.C.; Toyoda, A.; Takahashi, S.; Fukui, A.; Hikosaka, A.; Suzuki, A.; Kondo, M.; et al. Genome evolution in the allotetraploid frog Xenopus laevis. Nature 2016, 538, 336. [Google Scholar] [CrossRef] [PubMed]
- García de Vinuesa, A.; Abdelilah-Seyfried, S.; Knaus, P.; Zwijsen, A.; Bailly, S. BMP signaling in vascular biology and dysfunction. Cytokine Growth Factor Rev. 2016, 27, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Thiriet, M. Vascular Endothelium. In Tissue Functioning and Remodeling in the Circulatory and Ventilatory Systems; Thiriet, M., Ed.; Springer: New York, NY, USA, 2013; pp. 453–603. [Google Scholar]
- Marconcini, L.; Marchiò, S.; Morbidelli, L.; Cartocci, E.; Albini, A.; Ziche, M.; Bussolino, F.; Oliviero, S. C-fos-induced growth factor/vascular endothelial growth factor D induces angiogenesis in vivo and in vitro. Proc. Natl. Acad. Sci. USA 1999, 96, 9671–9676. [Google Scholar] [CrossRef]
- Masaki, T. Historical review: Endothelin. Trends Pharmacol. Sci. 2004, 25, 219–224. [Google Scholar] [CrossRef]
- Arikan, H.; Cicek, K. Haematology of amphibians and reptiles: A review. North-West. J. Zool. 2014, 10, 190–209. [Google Scholar]
- Rose, C.S.; James, B. Plasticity of lung development in the amphibian, Xenopus laevis. Biol. Open 2013, 2, 1324–1335. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, L.; Zhang, M.; Chen, Q.; Liu, J.; Zhu, W.; Jiang, J. From Water to Land: The Structural Construction and Molecular Switches in Lungs during Metamorphosis of Microhyla fissipes. Biology 2022, 11, 528. https://doi.org/10.3390/biology11040528
Chang L, Zhang M, Chen Q, Liu J, Zhu W, Jiang J. From Water to Land: The Structural Construction and Molecular Switches in Lungs during Metamorphosis of Microhyla fissipes. Biology. 2022; 11(4):528. https://doi.org/10.3390/biology11040528
Chicago/Turabian StyleChang, Liming, Meihua Zhang, Qiheng Chen, Jiongyu Liu, Wei Zhu, and Jianping Jiang. 2022. "From Water to Land: The Structural Construction and Molecular Switches in Lungs during Metamorphosis of Microhyla fissipes" Biology 11, no. 4: 528. https://doi.org/10.3390/biology11040528
APA StyleChang, L., Zhang, M., Chen, Q., Liu, J., Zhu, W., & Jiang, J. (2022). From Water to Land: The Structural Construction and Molecular Switches in Lungs during Metamorphosis of Microhyla fissipes. Biology, 11(4), 528. https://doi.org/10.3390/biology11040528