Selenium Nanoparticles Biosynthesized by Pantoea agglomerans and Their Effects on Cellular and Physiological Parameters in the Rainbow Trout Oncorhynchus mykiss

 ,
,  , ,
, ,
Simple Summary
Abstract
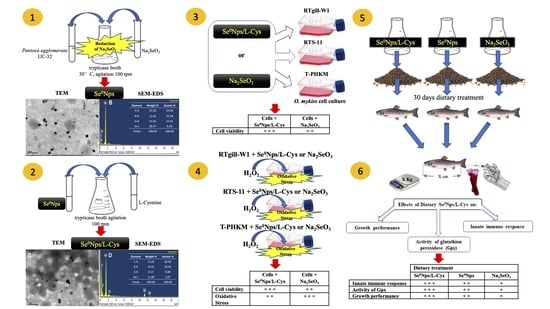
1. Introduction
2. Materials and Methods
2.1. Biosynthesis, Purification, and Functionalization of Se0Nps
2.2. Characterization of Se0Nps Biosynthesized by P. agglomerans UC-32 Strain
2.3. Antioxidant Capacity of Se0Nps/L-Cys
2.4. Effect of Se0Nps/L-Cys in Rainbow Trout’s Cells Culture (In Vitro Model)
2.4.1. Oncorhynchus Mykiss Cell Lines and Primary Head Kidney Monocyte-like Cells Culture
2.4.2. In Vitro Analysis of the Toxicity of Se0Nps/L-Cys
2.4.3. In Vitro Effect of Se0Nps/L-Cys against H2O2-Induced Oxidative Stress on Rainbow Trout Cell Cultures
2.5. Effect of Se0Nps/L-Cys Supplemented Food in Rainbow Trout’s (In Vivo Model)
2.5.1. Feeding Trial Design
2.5.2. Fish Sampling
2.5.3. Innate Immune Responses
2.5.4. Activity of the Antioxidant Enzyme Glutathione Peroxidase (Gpx)
2.5.5. Effect of Se0Nps/L-Cys on Trout Growth Performance and Survival Rate
2.6. Statistics
3. Results
3.1. Characterization of Se0Nps Biosynthesized by P. agglomerans UC-32 without and after Functionalization
3.2. Antioxidant Capacity of Se0Nps/L-Cys
3.3. Toxicity of Se0Nps/L-Cys for Cell Lines RTgill-W1 and RTS-11 and Primary Culture T-PHKM
3.4. Effect of Se0Nps/L-Cys on H2O2-Induced Oxidative Stress in Cell Lines RTgill-W1 and RTS-11 and T-PHKM Primary Cell Culture
3.5. In Vitro Effect of Se0Nps/L-Cys on ROS Concentration in Cell Lines RTgill-W1 and RTS-11 and Primary Culture T-PHKM
3.6. Effect of Se0Nps/L-Cys Supplemented Food in Rainbow Trout (In Vivo Model)
3.6.1. Innate Immune Responses
3.6.2. Activity of the Antioxidant Enzyme Gpx
3.6.3. Growth Performance and Survival
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fouré, J.; Bénassy-Quérét, A.; Fontané, L. Modelling the world economy at the 2050 horizon. Econ. Transit. 2013, 21, 617–654. [Google Scholar] [CrossRef]
- Ellis, T.; Turnbull, J.; Knowles, T.G.; Lines, J.A.; Auchterlonie, N.A. Trends during development of Scottish salmon farming: An example of sustainable intensification? Aquaculture 2016, 458, 82–99. [Google Scholar] [CrossRef]
- Iversen, A.; Asche, F.; Øystein, H.; Nystøyl, R. Production cost and competitiveness in major salmon farming countries 2003–2018. Aquaculture 2020, 522, 735089. [Google Scholar] [CrossRef]
- Yarahmadi, P.; Miandare, H.K.; Fayaz, S.; Caipang, C.M.A. Increased stocking density causes changes in expression of selected stress- and immune-related genes, humoral innate immune parameters and stress responses of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2015, 48, 43–53. [Google Scholar] [CrossRef]
- Trenzado, C.E.; Morales, A.E.; Palma, J.M.; de la Higuera, M. Blood antioxidant defenses and hematological adjustments in crowded/uncrowded rainbow trout (Oncorhynchus mykiss) fed on diets with different levels of antioxidant vitamins and HUFA. Comp. Biochem. Physiol. Part C 2009, 149, 440–447. [Google Scholar] [CrossRef]
- Küçükbay, F.Z.; Yazlak, H.; Karaca, I.; Sahin, N.; Tuzcu, M.; Cakmak, M.N.; Sahin, K. The effects of dietary organic or inorganic selenium in rainbow trout (Oncorhynchus mykiss) under crowding conditions. Aquac. Nutr. 2009, 15, 569–576. [Google Scholar] [CrossRef]
- Takahashi, K.; Suzuki, N.; Ogra, Y. Effect of gut microflora on nutritional availability of selenium. Food Chem. 2020, 319, 126537. [Google Scholar] [CrossRef]
- Mahdi Naderi, M.; Keyvanshokooh, S.; Salati, A.P.; Ghaedi, A. Combined or individual effects of dietary vitamin E and selenium nanoparticles on humoral immune status and serum parameters of rainbow trout (Oncorhynchus mykiss) under high stocking density. Aquaculture 2017, 474, 40–47. [Google Scholar] [CrossRef]
- Kousha, M.; Yeganeh, S.; Amirkolaie, A.K. Synergistic effect of sodium selenite and Pediococcus acidilactici on growth, intestinal bacterial counts, selenium bioavailability, hepatic enzymes and non-specific immune response in rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2019, 26, 74–87. [Google Scholar] [CrossRef]
- Baeverfjord, G.; Prabhu, P.A.; Fjelldal, P.G.; Albrektsen, S.; Hatlen, B.; Denstadli, V.; Ytteborg, E.; Takle, H.; Lock, E.-J.; Berntssen, M.H.G.; et al. Mineral nutrition and bone health in salmonids. Rev. Aquac. 2018, 11, 740–765. [Google Scholar] [CrossRef]
- Kohshahi, A.J.; Sourinejad, I.; Sarkheil, M.; Johari, S.A. Dietary cosupplementation with curcumin and different selenium sources (nanoparticulate, organic, and inorganic selenium): Influence on growth performance, body composition, immune responses, and glutathione peroxidase activity of rainbow trout (Oncorhynchus mykiss). Fish Physiol. Biochem. 2019, 45, 793–804. [Google Scholar] [PubMed]
- Hilton, J.W.; Hodson, P.V.; Slinger, S.J. The requirement and toxicity of selenium in rainbow trout (Salmo gairdneri). J. Nutr. 1980, 110, 2527–2535. [Google Scholar] [CrossRef] [PubMed]
- Rider, S.A.; Davies, S.J.; Jha, A.N.; Fisher, A.A.; Knight, J.; Sweetman, J.W. Supra-nutritional dietary intake of selenite and selenium yeast in normal and stressed rainbow trout (Oncorhynchus mykiss): Implications on selenium status and health responses. Aquaculture 2009, 295, 282–291. [Google Scholar] [CrossRef]
- Ytrestøyl, T.; Aas, T.S.; Åsgård, T. Utilisation of feed resources in production of Atlantic salmon (Salmo salar) in Norway. Aquaculture 2015, 448, 365–374. [Google Scholar] [CrossRef]
- Sanden, M.; Hemre, G.-I.; Måge, A.; Lunestad, B.T.; Espe, M.; Lundebye, A.-K.; Ørnsrud, R. Program for Overvåking av Fiskefôr. Nasjonalt Institutt for Ernærings-og Sjømatforskning (NIFES). 2013. Available online: https://www.mattilsynet.no/dyr_og_dyrehold/for/overvaakingsprogram_fiskefor_2013.16731/binary/Overvåkingsprogram%20fiskefôr%202013 (accessed on 20 September 2021).
- Sanden, M.; Hemre, G.-I.; Måge, A.; Lunestad, B.T.; Espe, M.; Lie, K.K.; Lundebye, A.-K.; Amlund, H.; Waagbø, R.; Ørnsrud, R. Program for Overvåking av Fiskefôr. Nasjonalt Institutt for Ernærings-og Sjømatforskning (NIFES). 2017. Available online: https://www.hi.no/resources/publikasjoner/rapporter-nifes/2017/forrapport2017.pdf (accessed on 20 September 2021).
- Betancor, M.B.; Dam, T.M.C.; Walton, J.; Morken, T.; Campbell, P.J.; Tocher, D.R. Modulation of selenium tissue distribution and selenoprotein expression in Atlantic salmon (Salmo salar L.) fed diets with graded levels of plant ingredients. Br. J. Nutr. 2016, 115, 1325–1338. [Google Scholar] [CrossRef] [PubMed]
- Marta, S.; Silva, M.S.; Kröckel, S.; Prabhu, P.A.J.; Koppe, W.; Ørnsrud, R.; Waagbø, R.; Araujo, P.; Amlund, H. Apparent availability of zinc, selenium and manganese as inorganic metal salts or organic forms in plant-based diets for Atlantic salmon (Salmo salar). Aquaculture 2019, 503, 562–570. [Google Scholar]
- Committee on the Nutrient Requirements of Fish and Shrimp & National Research Council (NRC). Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Pacitti, D.; Wang, T.; Page, M.M.; Martin, S.A.M.; Sweetman, J.; Feldmann, J.; Secombes, C.J. Characterization of cytosolic glutathione phospholipid-hydroperoxide peroxidase and glutathione peroxidase genes in rainbow trout (Oncorhynchus mykiss) and their modulation by in vitro selenium exposure. Aquat. Toxicol. 2013, 130–131, 97–111. [Google Scholar] [CrossRef]
- Fontagné-Dicharry, S.; Godin, S.; Liu, H.; Prabhu, P.A.J.; Bouyssière, B.; Bueno, M.; Tacon, P.; Médale, F.; Kaushik, S. Influence of the forms and levels of dietary selenium on antioxidant status and oxidative stress-related parameters in rainbow trout (Oncorhynchus mykiss) fry. Br. J. Nutr. 2015, 113, 1876–1887. [Google Scholar] [CrossRef]
- Petros, R.A.; DeSimone, J.M. Strategies in the design of nanoparticles for therapeutic applications. Nat. Rev. Drug Discov. 2010, 9, 615–627. [Google Scholar] [CrossRef]
- Cholewinska, E.; Ognik, K.; Fotschki, B.; Zdunczyk, Z.; Juskiewicz, J. Comparison of the effect of dietary copper nanoparticles and one copper (II) salt on the copper biodistribution and gastrointestinal and hepatic morphology and function in a rat model. PLoS ONE 2018, 13, e0197083. [Google Scholar] [CrossRef]
- Adegbeye, M.J.; Elghandour, M.M.; Barbabosa-Pliego, A.; Monroy, J.C.; Mellado, M.; Salem, A.Z. Nanoparticles in Equine Nutrition: Mechanism of Action and Application as Feed Additives. J. Equine Vet. Sci. 2019, 78, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wong, Y.S.; Zheng, W.; Bai, Y.; Huang, L. Selenium nanoparticles fabricated in Undaria pinnatifida polysaccharide solutions induce mitochondria-mediated apoptosis in A375 human melanoma cells. Colloids Surf. B Biointerfaces 2008, 67, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Naderi, M.; Keyvanshokooh, S.; Salati, A.P.; Ghaedi, A. Proteomic analysis of liver tissue from rainbow trout (Oncorhynchus mykiss) under high rearing density after administration of dietary vitamin E and selenium nanoparticles. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 22, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Dobias, J.; Suvorova, E.I.; Bernier-Latmani, R. Role of proteins in controlling selenium nanoparticle size. Nanotechnology 2011, 22, 195605. [Google Scholar] [CrossRef] [PubMed]
- Arshad, A. Bacterial Synthesis and Applications of Nanoparticles. Nano Sci. Nano Technol. 2017, 11, 119. [Google Scholar]
- Subbiah, R.; Veerapandian, M.; Yun, K.S. Nanoparticles: Functionalization and Multifunctional Applications in Biomedical Sciences. Curr. Med. Chem. 2010, 17, 4559–4577. [Google Scholar] [CrossRef] [PubMed]
- Torres, S.K.; Campos, V.L.; León, C.G.; Rodríguez-Llamazares, S.M.; Rojas, S.M.; González, M.; Smith, C.T.; Mondaca, M.A. Biosynthesis of selenium nanoparticles by Pantoea agglomerans and their antioxidant activity. J. Nanoparticle Res. 2012, 14, 1236. [Google Scholar] [CrossRef]
- Tarrahi, R.; Khataee, A.; Movafeghi, A.; Rezanejad, F.; Gohari, G. Toxicological implications of selenium nanoparticles with different coatings along with Se4+ on Lemna minor. Chemosphere 2017, 181, 655–665. [Google Scholar] [CrossRef]
- Dhanjal, S.; Cameotra, S.S. Aerobic biogenesis of selenium nanospheres by Bacillus cereus isolated from coalmine soil. Microb. Cell Factories 2010, 9, 52. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Bensaci, C.; Ghiaba, Z.; Dakmouche, M.; Belfar, A.; Belguidoum, M.; Bentebba, F.Z.; Saidi, M.; Hadjadj, M. In Vitro Evaluation of Antioxidant Potential of Date Palm Collected in Algeria using Electrochemical and Spectrophotometrical Techniques. Korean Chem. Eng. Res. 2021, 59, 153–158. [Google Scholar]
- Dudonné, S.; Vitrac, X.; Coutière, P.; Woillez, M.; Mérillon, J. Comparative study of antioxidant properties and total phenolic content of 30 plant extracts of industrial interest using DPPH, ABTS, FRAP, SOD, and ORAC assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Romay, C.; Pascual, C.E.L.; Lissi, E.A. The reaction between ABTS radical cation and antioxidants and its use to evaluate the antioxidant status of serum samples. Braz. J. Med. Biol. Res. 1996, 29, 175–183. [Google Scholar] [PubMed]
- Abarca, A.; Bethke, J.; Narváez, E.; Flores, R.; Mercado, L. Parameters to evaluate the immunostimulant effect of Zymosan A in head kidney leucocytes (HKL) of salmonids. Lat. Am. J. Aquat. Res. 2012, 40, 545–552. [Google Scholar] [CrossRef]
- Singh, S.P.; Rastogi, R.P.; Hader, D.P.; Sinha, R.P. Temporal dynamics of ROS biogenesis under simulated solar radiation in the cyanobacterium Anabaena variabilis PCC 7937. Protoplasma 2014, 251, 1223–1230. [Google Scholar] [CrossRef]
- Holmström, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef]
- Kling, P.; Olsson, P.-E. Involvement of differential metallothionein expression in free radical sensitivity of RTG-2 and CHSE-214 Cells. Free Radic. Biol. Med. 2000, 28, 1628–1637. [Google Scholar] [CrossRef]
- Chen, L.-D.; Liu, Z.-H.; Zhang, L.-F.; Yao, J.-N.; Wa, C.-F. Sanggenon C induces apoptosis of colon cancer cells via inhibition of NO production, iNOS expression and ROS activation of the mitochondrial pathway. Oncol. Rep. 2017, 38, 2123–2131. [Google Scholar] [CrossRef]
- Valenzuela, A.; Campos, V.; Yañez, F.; Alveal, K.; Gutiérrez, P.; Rivas, M.; Contreras, N.; Klempau, A.; Fernandez, I.; Oyarzun, C. Application of artificial photoperiod in fish: A factor that increases susceptibility to infectious diseases? Fish Physiol. Biochem. 2012, 38, 943–950. [Google Scholar] [CrossRef]
- Vera, B. Bio-Obtención de Nanopartículas de Selenio y su Potencial Aplicación Como Suplemento Alimentario Inmunoestimulante en Trucha Arcoíris (Oncorhynchus mykiss). Bachelor’s Thesis, University of Concepcion, Concepción, Chile, March 2016. [Google Scholar]
- Takemura, A.; Takano, K. Lysozyme in the ovary of tilapia (Oreochromis mossambicus): Its purification and some biological properties. Fish Physiol. Biochem. 1995, 14, 415–421. [Google Scholar] [CrossRef]
- Anderson, D.; Siwicki, A. Measuring the effects of contaminants on fish by haematological and serological methods. In Modulators of Fish Immune Responses; Stolen, J., Anderson, D., Zelikoff, S., Twerdok, L., Kaattari, S., Eds.; SOS Publications: Fair Haven, NJ, USA, 1993; pp. 95–118. [Google Scholar]
- Lawrence, R.; Burk, R. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem. Biophys. Res. Commun. 1976, 71, 952–958. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Lugert, V.; Thaller, G.; Tetens, J.; Schulz, C.; Krieter, J. A review on fish growth calculation: Multiple functions in fish production and their specific application. Rev. Aquac. 2014, 8, 30–42. [Google Scholar] [CrossRef]
- Mansour, A.T.-E.; Goda, A.A.; Omar, E.A.; SaberKhalil, H.; Esteban, M.-A. Dietary supplementation of organic selenium improves growth, survival, antioxidantand immune status of meagre, Argyrosomus regius, juveniles. Fish Shellfish Immunol. 2017, 68, 516–524. [Google Scholar] [CrossRef]
- Khan, K.U.; Zuberi, A.; Nazir, S.; Fernandes, J.B.K.; Jamil, Z.; Sarwar, H. Effects of dietary selenium nanoparticles on physiological and biochemical aspects of juvenile Tor putitora. Turk. J. Zool. 2016, 40, 704–712. [Google Scholar] [CrossRef]
- Baldissera, M.D.; Souza, C.F.; Parmeggiani, B.; Leipnitz, G.; Verdi, C.M.; Santos, R.V.; Stefani, L.M.; Baldisserotto, B. The disturbance of antioxidant/oxidant balance in fish experimentally infected by Aeromonas caviae: Relationship with disease pathophysiology. Microb. Pathog. 2018, 122, 53–57. [Google Scholar] [CrossRef]
- Farzad, R.; Kuhn, D.; Smith, S.; O’Keefe, S.; Hines, I.; Bushman, T.; Galagarza, O.; Stevens, A. Effects of selenium-enriched prebiotic on the growth performance, innate immune response, oxidative enzyme activity and microbiome of rainbow trout (Oncorhynchus mykiss). Aquaculture 2021, 735980. [Google Scholar] [CrossRef]
- Saffari, S.; Saeed Keyvanshokooh, S.; Zakeri, M.; Johari, S.A.; Pasha-Zanoosi, H.; Mozanzadeh, M.T. Effects of dietary organic, inorganic, and nanoparticulate selenium sources on growth, hemato-immunological, and serum biochemical parameters of common carp (Cyprinus carpio). Fish Physiol. Biochem. 2018, 44, 1087–1097. [Google Scholar] [CrossRef]
- Daza, C.; Campos, V.; Rojas, C.; Rodríguez-Llamazares, S.; Smith, C.; Mondaca, M. Reduction of selenite to elemental Selenium by Pantoea agglomerans. Gayana 2016, 80, 67–74. [Google Scholar] [CrossRef]
- Prasanth, S.; Sudarsanakumar, C. Elucidating the interaction of L-cysteine-capped selenium nanoparticles and human serum albumin: Spectroscopic and thermodynamic analysis. New J. Chem. 2017, 41, 9521–9530. [Google Scholar] [CrossRef]
- Perni, S.; Hakala, V.; Prokopovich, P. Biogenic synthesis of antimicrobial silver nanoparticles capped with L-cysteine. Colloids Surf. A 2013, 460, 219–224. [Google Scholar] [CrossRef]
- Prasanth, S.; Rithesh Raj, D.; Vineeshkumar, T.V.; Thomas, R.K.; Sudarsanakumar, C. Exploring the interaction of l-cysteine capped CuS nanoparticles with bovine serum albumin (BSA): A spectroscopic study. RSC Adv. 2016, 6, 58288–58295. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Q.; Ding, Y.; Cai, X.; Gu, S.; Cao, Z. Application of l-cysteine capped core–shell CdTe/ZnS nanoparticles as a fluorescence probe for cephalexin. Anal. Methods 2014, 6, 2715–2721. [Google Scholar] [CrossRef]
- Wojnicki, M.; Luty-Błocho, M.; Kotańska, M.; Wytrwal, M.; Tokarski, T.; Krupa, A.; Kołaczkowski, M.; Bucki, A.; Kobielusz, M. Novel and effective synthesis protocol of AgNPs functionalized using L-cysteine as a potential drug carrier. Naunyn-Schmiedebergs Arch. Pharmacol. 2017, 391, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Majzik, A.; Patakfalvi, R.; Hornok, V.; Dékány, I. Growing and stability of gold nanoparticles and their functionalization by cysteine. Gold Bull. 2009, 42, 113–123. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, L.; Sun, Y. Size-controlled green synthesis of silver nanoparticles assisted by L-cysteine. Front. Chem. Sci. Eng. 2015, 9, 494–500. [Google Scholar] [CrossRef]
- Chatterjee, A.; Priyam, A.; Das, S.K.; Saha, A. Size tunable synthesis of cysteine-capped CdS nanoparticles by γ-irradiation. J. Colloid Interface Sci. 2006, 294, 334–342. [Google Scholar] [CrossRef]
- Huang, B.; Zhang, J.; Hou, J.; Chen, C. Free radical scavenging efficiency of nano-se in vitro. Free Radic. Biol. Med. 2003, 35, 805–813. [Google Scholar] [CrossRef]
- Matsuura, S.; Katsumi, H.; Suzuki, H.; Hirai, N.; Takashima, R.; Morishita, M.; Sakane, T.; Yamamoto, A. l-Cysteine and l-Serine Modified Dendrimer with Multiple Reduced Thiols as a Kidney-Targeting Reactive Oxygen Species Scavenger to Prevent Renal Ischemia/Reperfusion Injury. Pharmaceutics 2018, 10, 251. [Google Scholar] [CrossRef]
- Bai, K.; Hong, B.; Huang, W.; He, J. Selenium-Nanoparticles-Loaded Chitosan/Chitooligosaccharide Microparticles and Their Antioxidant Potential: A Chemical and in vivo Investigation. Pharmaceutics 2020, 12, 43. [Google Scholar] [CrossRef]
- Forootanfar, H.; Adeli-Sardou, M.; Nikkhoo, M.; Mehrabani, M.; Amir-Heidari, B.; Shahverdi, A.R.; Shakibaie, M. Antioxidant and cytotoxic effect of biologically synthesized selenium nanoparticles in comparison to selenium dioxide. J. Trace Elem. Med. Biol. 2014, 28, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Qiao, L.; Guo, Y.; Ma, L.; Cheng, Y. Preparation, characteristics and antioxidant activity of polysaccharides and proteins-capped selenium nanoparticles synthesized by Lactobacillus casei ATCC 393. Carbohydr. Polym. 2018, 195, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Bettaib, J.; Talarmin, H.; Kalai, F.Z.; Giroux-Metges, M.-A.; Ksouri, R. Limoniastrum guyonianum prevents H2O2-induced oxidative damage in IEC-6 cells by enhancing enzyamtic defense, reducing glutathione depletion and JNK phosphorylation. Biomed. Pharmacother. 2017, 95, 1404–1411. [Google Scholar] [CrossRef] [PubMed]
- Mou, K.; Pan, W.; Han, D.; Wen, X.; Cao, F.; Miao, Y.; Li, P. Glycyrrhizin protects human melanocytes from H2O2-induced oxidative damage via the Nrf2-dependent induction of HO-1. Int. J. Mol. Med. 2019, 44, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Franco, M.E.; Sutherland, G.E.; Lavado, R. Xenobiotic metabolism in the fish hepatic cell lines Hepa-E1 and RTH-149, and the gill cell lines RTgill-W1 and G1B: Biomarkers of CYP450 activity and oxidative stress. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2018, 206–207, 32–40. [Google Scholar] [CrossRef]
- Bopp, S.K.; Abicht, H.K.; Knauer, K. Copper-induced oxidative stress in rainbow trout gill cells. Aquat. Toxicol. 2008, 86, 197–204. [Google Scholar] [CrossRef]
- Uçar, A.; Parlak, V.; Özgeriş, F.B.; Yeltekin, A.Ç.; Alak, G.; Atamanalp, M. Determination of Fipronil toxicity by different biomarkers in gill and liver tissue of rainbow trout (Oncorhynchus mykiss). Vitr. Cell. Dev. Biol.–Anim. 2020, 56, 543–549. [Google Scholar] [CrossRef]
- Banaee, M. Chapter 4. Physiological Dysfunction in Fish after Insecticides Exposure. In Insecticides Development of Safer and More Effective Technologies, 1st ed.; Trdan, S., Ed.; IntechOpen: London, UK, 2013; pp. 103–142. ISBN 978-953-51-5348-1. [Google Scholar]
- Tkachenko, H.; Kurhaluk, N.; Grudniewska, J.; Andriichuk, A. Tissue-specific responses of oxidative stress biomarkers and antioxidant defenses in rainbow trout Oncorhynchus mykiss during a vaccination against furunculosis. Fish Physiol. Biochem. 2014, 40, 1289–1300. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Zommara, M.; Eweedah, N.M.; Helal, A.I.; Aboel-Darag, M.A. The potential role of nano-selenium and vitamin C on the performances of Nile tilapia (Oreochromis niloticus). Environ. Sci. Pollut. Res. 2020, 27, 9843–9852. [Google Scholar] [CrossRef]
- Harsij, M.; Kanani, H.G.; Adineh, H. Effects of antioxidant supplementation (nano-selenium, vitamin C and E) on growth performance, blood biochemistry, immune status and body composition of rainbow trout (Oncorhynchus mykiss) under sub-lethal ammonia exposure. Aquaculture 2020, 521, 734942. [Google Scholar] [CrossRef]
- Neamat-Allah, A.N.F.; Mahmoud, E.A.; Abd El Hakim, Y. Efficacy of dietary Nano-selenium on growth, immune response, antioxidant, transcriptomic profile and resistance of Nile tilapia, Oreochromis niloticus against Streptococcus iniae infection. Fish Shellfish Immunol. 2019, 94, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Dalmo, R.A.; Ingebrigtsen, K.; Bøgwald, J. Non-specific defence mechanisms in fish, with particular reference to the reticuloendothelial system (RES). J. Fish Dis. 1997, 20, 241–273. [Google Scholar] [CrossRef]
- Rodríguez, A.; Esteban, M.Á.; Meseguer, J. Phagocytosis and peroxidase release by seabream (Sparus aurata L.) leucocytes in response to yeast cells. Anat. Rec. A Discov. Mol. Cell Evol. Biol. 2003, 272A, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Zommara, M.; Eweedah, N.M.; Helal, A.I. Synergistic Effects of Selenium Nanoparticles and Vitamin E on Growth, Immune-Related Gene Expression, and Regulation of Antioxidant Status of Nile Tilapia (Oreochromis niloticus). Biol. Trace Elem. Res. 2019, 195, 624–635. [Google Scholar] [CrossRef] [PubMed]
- Xia, I.F.; Cheung, J.S.; Wu, M.; Wong, K.-S.; Kong, H.; Zheng, X.; Ka-Hing Wong, K.-H.; Kwok, K.W. Dietary chitosan-selenium nanoparticle (CTS-SeNP) enhance immunity and disease resistance in zebrafish. Fish Shellfish Immunol. 2019, 87, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Longbaf Dezfouli, M.; Ghaedtaheri, A.; Keyvanshokooh, S.; Salati, A.P.; Mousavi, S.M.; Pasha-Zanoosi, H. Combined or individual effects of dietary magnesium and selenium nanoparticles on growth performance, immunity, blood biochemistry and antioxidant status of Asian seabass (Lates calcarifer) reared in freshwater. Aquac. Nutr. 2019, 25, 1422–1430. [Google Scholar] [CrossRef]
- Sattin, G.; Bakiu, R.; Tolomeo, A.M.; Carraro, A.; Coppola, D.; Ferro, D.; Patarnello, T.; Santovito, G. Characterization and expression of a new cytoplasmic glutathione peroxidase 1 gene in the Antarctic fish Trematomus bernacchii. Hydrobiologia 2015, 761, 363–372. [Google Scholar] [CrossRef]
- Tollerz Bratteby, U. Factors Explaining Variation in the Fecundity of Female Baltic Salmon (Salmo salar)–The Influence of Length, Body Condition and Growth Rate at Sea. Master’s Thesis, Swedish University of Agricultural Sciences, Öregrund, Sweden, 2019. [Google Scholar]
- Rautela, A.; Rani, J.; Das, M.D. Green synthesis of silver nanoparticles from Tectona grandis seeds extract: Characterization and mechanism of antimicrobial action on different microorganisms. J. Anal. Sci. Technol. 2019, 10, 5. [Google Scholar] [CrossRef]
- Fang, X.; Wang, Y.; Wang, Z.; Jiang, Z.; Dong, M. Microorganism Assisted Synthesized Nanoparticles for Catalytic Applications. Energies 2019, 12, 190. [Google Scholar] [CrossRef]

 ) of cell lines RTS-11 (A), RTgill-W1 (B), and of the primary culture T-PHKM (C) treated with Se0Nps/L-Cys
) of cell lines RTS-11 (A), RTgill-W1 (B), and of the primary culture T-PHKM (C) treated with Se0Nps/L-Cys  or Na2SeO3
or Na2SeO3  and then subjected to H2O2 as a ROS inducing agent. All data is given as mean ± SD. Positive controls
and then subjected to H2O2 as a ROS inducing agent. All data is given as mean ± SD. Positive controls  . Different letters on top of bars indicate significant differences among groups. * Statistically different from the negative control.
) of cell lines RTS-11 (A), RTgill-W1 (B), and of the primary culture T-PHKM (C) treated with Se0Nps/L-Cys or Na2SeO3 and then subjected to H2O2 as a ROS inducing agent. All data is given as mean ± SD. Positive controls . Different letters on top of bars indicate significant differences among groups. * Statistically different from the negative control.
. Different letters on top of bars indicate significant differences among groups. * Statistically different from the negative control.
) of cell lines RTS-11 (A), RTgill-W1 (B), and of the primary culture T-PHKM (C) treated with Se0Nps/L-Cys or Na2SeO3 and then subjected to H2O2 as a ROS inducing agent. All data is given as mean ± SD. Positive controls . Different letters on top of bars indicate significant differences among groups. * Statistically different from the negative control.
 or Na2SeO3
or Na2SeO3  and then subjected to H2O2 as a ROS inducing agent. All data is given as mean ± SD. Different letters on top of bars indicate significant differences among groups. * Significant reduction of cellular ROS concentration compared to the positive controls
and then subjected to H2O2 as a ROS inducing agent. All data is given as mean ± SD. Different letters on top of bars indicate significant differences among groups. * Significant reduction of cellular ROS concentration compared to the positive controls  . Negative control
. Negative control  .
or Na2SeO3 and then subjected to H2O2 as a ROS inducing agent. All data is given as mean ± SD. Different letters on top of bars indicate significant differences among groups. * Significant reduction of cellular ROS concentration compared to the positive controls . Negative control .
.
or Na2SeO3 and then subjected to H2O2 as a ROS inducing agent. All data is given as mean ± SD. Different letters on top of bars indicate significant differences among groups. * Significant reduction of cellular ROS concentration compared to the positive controls . Negative control .

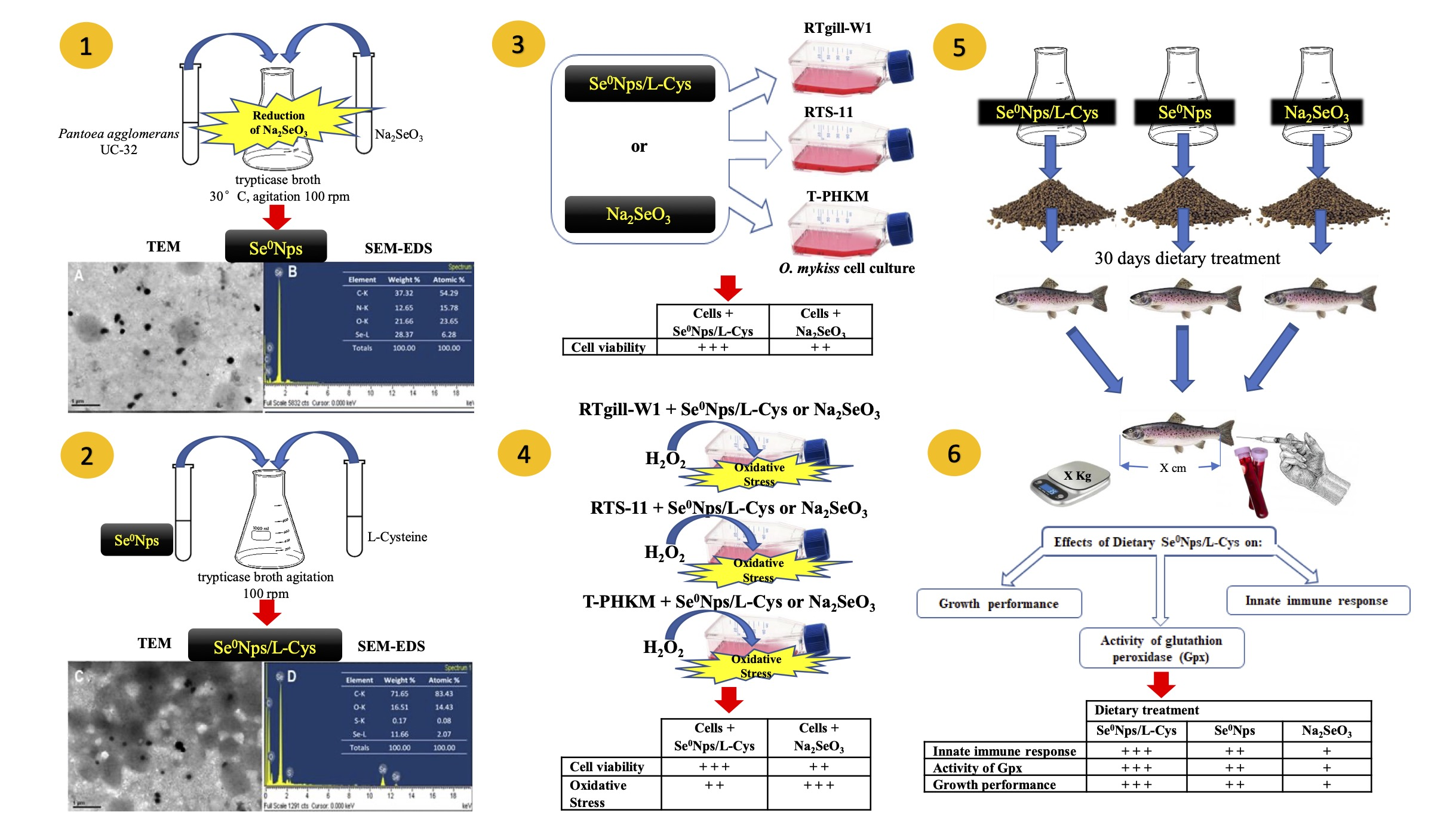{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antioxidant | DPPH IC50 (mg mL−1) ± SD | FRAP (TEAC mM) ± SD | TRAP (TEAC mM) ± SD |
|---|---|---|---|
| Se0Nps/L-Cys | 1.96 ± 0.71 | 0.10 ± 0.03 | 0.19 ± 0.04 |
| Se0Nps | 2.53 ± 0.91 | 0.09 ± 0.01 | 0.15 ± 0.02 |
| Na2SeO3 | 3.47 ± 0.49 | 0.06 ± 0,01 | 0.08 ± 0.01 |
| Vit C | 0.77 ± 0.08 | 0.26 ± 0.06 | 0.85 ± 0.01 |
| Trolox | 1.14 ± 0.06 | n.a | n.a |
| NAC | 1.42 ± 0.19 | 0.05 ± 0.02 | 0.16 ± 0.01 |
| Cells | Se0Nps/L-Cys (nM) | Na2SeO3 (nM) | ||||
|---|---|---|---|---|---|---|
| 160 | 320 | 640 | 160 | 320 | 640 | |
| RTgill-W1 | 95.64 ± 1.83 | 94.47 ± 2.22 | 92.66 ± 1.97 | 93.43 ± 2.35 | 92.05 ± 2.23 | 90.25 ± 1.67 |
| RTS-11 | 96.39 ± 1.13 | 96.92 ± 1.45 | 95.67 ± 1.20 | 95.35 ± 1.65 | 95.05 ± 1.94 | 93.74 ± 2.21 |
| T-PHKM | 96.52 ± 0.43 | 96.05 ± 1.25 | 94.02 ± 1.13 | 96.13 ± 1.25 | 94.23 ± 1.30 | 93.33± 1.30 |
| Dietary Treatment | ||||
|---|---|---|---|---|
| Day | Control | Se0Nps | Se0Nps/L-Cys | Na2SeO3 |
| 0 | 36.37 ± 3.9 | 35.11 ± 4.1 | 35.25 ± 3.3 | 36.56 ± 3.8 |
| 15 | 37.14 ± 4.1 | 39.13 ± 4.7 | 41.47 ± 2.8 | 38.21 ± 3.6 |
| 30 | 37.53 ± 2.1 | 43.34 ± 2.6 | 46.40 ± 2.5 | 41.07 ± 3.3 |
| Dietary Treatment | ||||
|---|---|---|---|---|
| Day | Control | Se0Nps | Se0Nps/L-Cys | Na2SeO3 |
| 0 | 0.46 ± 0.014 | 0.44 ± 0.013 | 0.44 ± 0.11 | 0.45 ± 0.18 |
| 15 | 0.43 ± 0.017 | 0.49 ± 0.020 | 0.46 ± 0.14 | 0.44 ± 0.11 |
| 30 | 0.40 ± 0.021 | 0.45 ± 0.013 | 0.49 ± 0.02 | 0.41 ± 0.24 |
| Gpx Activity | Dietary Treatment | |||
|---|---|---|---|---|
| Control | Se0Nps | Se0Nps/L-Cys | Na2SeO3 | |
| PlasmaLiver | 257.36 ± 4.30 | 274.25 ± 5.86 | 279.39 ± 7.17 | 271.80 ± 8.47 |
| 22.01 ± 2.06 | 28.31 ± 3.31 | 29.46 ± 3.71 | 26.67 ± 3.19 | |
| Muscle | 25.98 ± 2.85 | 36.26 ± 3.64 | 40.06 ± 3.04 | 34.02 ± 1.83 |
| Index | Dietary Treatment | |||
|---|---|---|---|---|
| Control | Se0Nps | Se0Nps/L-Cys | Na2SeO3 | |
| IW (g fish−1) | 104.57 ± 4.69 | 102.26 ± 4.63 | 101.77 ± 4.63 | 103.11 ± 4.64 |
| FW (g fish−1) | 174.45 ± 3.86 | 178.14 ± 3.59 | 179.66 ± 2.71 | 175.76 ± 2.61 |
| WG (g) | 70.54 ± 6.63 | 76.28 ± 6.15 | 78.00 ± 4.25 | 72.69 ± 6.69 |
| SGR (%) | 1.72 ± 0.18 | 1.86 ± 0.17 | 1.89 ± 0.12 | 1.78 ± 0.21 |
| ICF (%) | 1.24 ± 0.11 | 1.26 ± 0.19 | 1.22 ± 0.12 | 1.23 ± 0.10 |
| FCF (%) | 1.27 ± 0.24 | 1.52 ± 0.26 | 1.68 ± 0.35 | 1.45 ± 0.23 |
| Survival rate (%) | 100 | 100 | 100 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yanez-Lemus, F.; Moraga, R.; Mercado, L.; Jara-Gutierrez, C.; Smith, C.T.; Aguayo, P.; Sanchez-Alonzo, K.; García-Cancino, A.; Valenzuela, A.; Campos, V.L. Selenium Nanoparticles Biosynthesized by Pantoea agglomerans and Their Effects on Cellular and Physiological Parameters in the Rainbow Trout Oncorhynchus mykiss. Biology 2022, 11, 463. https://doi.org/10.3390/biology11030463
Yanez-Lemus F, Moraga R, Mercado L, Jara-Gutierrez C, Smith CT, Aguayo P, Sanchez-Alonzo K, García-Cancino A, Valenzuela A, Campos VL. Selenium Nanoparticles Biosynthesized by Pantoea agglomerans and Their Effects on Cellular and Physiological Parameters in the Rainbow Trout Oncorhynchus mykiss. Biology. 2022; 11(3):463. https://doi.org/10.3390/biology11030463
Chicago/Turabian StyleYanez-Lemus, Francisco, Rubén Moraga, Luis Mercado, Carlos Jara-Gutierrez, Carlos T. Smith, Paulina Aguayo, Kimberly Sanchez-Alonzo, Apolinaria García-Cancino, Ariel Valenzuela, and Victor L. Campos. 2022. "Selenium Nanoparticles Biosynthesized by Pantoea agglomerans and Their Effects on Cellular and Physiological Parameters in the Rainbow Trout Oncorhynchus mykiss" Biology 11, no. 3: 463. https://doi.org/10.3390/biology11030463
APA StyleYanez-Lemus, F., Moraga, R., Mercado, L., Jara-Gutierrez, C., Smith, C. T., Aguayo, P., Sanchez-Alonzo, K., García-Cancino, A., Valenzuela, A., & Campos, V. L. (2022). Selenium Nanoparticles Biosynthesized by Pantoea agglomerans and Their Effects on Cellular and Physiological Parameters in the Rainbow Trout Oncorhynchus mykiss. Biology, 11(3), 463. https://doi.org/10.3390/biology11030463