
Deep Insights into the Specific Evolution of Fungal Hybrid B Heme Peroxidases

 ,
,  , , , , and
, , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Strain and Growth Conditions
2.2. Isolation of Fungal DNA, Total RNA and Synthesis of cDNA
2.3. Investigation of Fungal Secretome and Proteomic Analysis
2.4. Multiple Sequence Alignment and Reconstruction of Evolutionary Relationships
2.5. Ancestral Sequence Reconstruction for Peroxidases
2.6. Modelling of 3D Structures for Ancestral Heme Peroxidases
2.7. Identification of Protein Tunnels and Ligand Docking
3. Results
3.1. Detection of Specific Fungal mRNA for ScepHyBpox1 and Gene Architecture
3.2. Detection of Specific Protein Patterns of ScepHyBpox1 in the Fungal Secretome
3.3. Evolutionary Relationships of Hybrid B Heme Peroxidases
3.4. Peculiar Conserved-Sequence Motifs of Hybrid B Heme Peroxidases
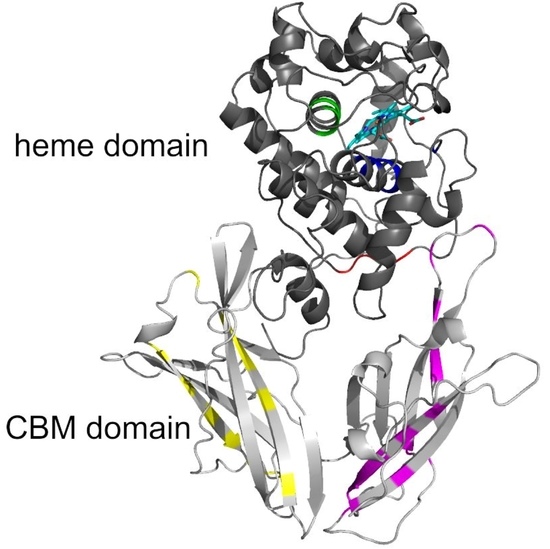
3.5. Typical Structural Folds in Hybrid B Heme Peroxidases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vitale, G.A.; Coppola, D.; Esposito, F.P.; Buonocore, C.; Ausuri, J.; Tortorella, E.; de Pascale, D. Antioxidant Molecules from Marine Fungi: Methodologies and Perspecitves. Antioxidants 2020, 9, 1183. [Google Scholar] [CrossRef]
- Zámocký, M.; Kamlárová, A.; Maresch, D.; Chovanová, K.; Harichová, J.; Furtmüller, P.G. Hybrid Heme Peroxidases from Rice Blast Fungus Magnaporthe oryzae Involved in Defence against Oxidative Stress. Antioxidants 2020, 9, 655. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, S.; Slezak-Prochazka, I.; Bil, P.; Rzeszowska-Wolny, J. Micro RNAs in Regulation of Cellular Redox Homeostasis. Int. J. Mol. Sci. 2021, 22, 6022. [Google Scholar] [CrossRef] [PubMed]
- Galasso, M.; Gambino, S.; Romanelli, M.G.; Donadelli, M.; Scupoli, M.T. Browsing the oldest antioxidant enzyme: Catalase and its multiple regulation in cancer. Free Redic. Biol. Med. 2021, 172, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Andrew, M.; Barua, R.; Short, S.M.; Kohn, L.M. Evidence for a Common Toolbox Based on Necrotrophy in a Fungal Lineage Spanning Necrotrophs, Biotrophs, Endophytes, Host Generalists and Specialists. PLoS ONE 2012, 7, e29943. [Google Scholar] [CrossRef]
- Hovius, M.H.Y.; Mc Donald, M.R. Management of Allium White rot [Sclerotium cepivorum] in Onions on Organic Soil with Soil-Applied Diallyl Disulfide and di-N-propyl Disulfide. Can. J. Plant. Pathol. 2002, 24, 281–286. [Google Scholar] [CrossRef][Green Version]
- Davis, R.M.; Hao, J.J.; Romberg, M.K.; Nunez, J.J.; Smith, R.F. Efficacy of Germination Stimulants of Sclerotia of Sclerotium cepivorum for Management of White Rot of Garlic. Plant Dis. 2007, 91, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Tyson, J.L.; Ridgway, H.J.; Fullerton, R.A.; Stewart, A. Genetic Diversity in New Zealand Populations of Sclerotium cepivorum. N. Z. J. Crop Horticult. Sci. 2002, 30, 37–48. [Google Scholar] [CrossRef]
- Valero-Jiménez, C.A.; Steentjes, M.B.F.; Slot, J.C.; Shi-Kunne, X.; Scholten, O.E.; van Kan, J.A.L. Dynamics of Secondary Metabolite Gene Clusters in Otherwise Highly Syntenic and Stable Genomes in the Fungal Genus Botrytis. Genome Biol. Evol. 2020, 12, 2491–2507. [Google Scholar] [CrossRef] [PubMed]
- Licona-Juárez, K.C.; Acosta-García, G.; Humberto Ramírez-Medina, H.; Wilson Huanca-Mamani, W.; Guevara-Olvera, L. Rapid and Accurate Pcr-based and Boiling DNA Isolation Methodology for Specific Detection of Sclerotium cepivorum in Garlic (Allium sativum) Cloves. Interciencia 2019, 44, 71–74. [Google Scholar]
- Hall, T.A. BioEdit: A User-friendly Biological Sequence Alignment Editor and Analysis Program for Windows. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Koontz, L. TCA Precipitation. Methods Enzymol. 2014, 541, 3–10. [Google Scholar]
- Škodová-Sveráková, I.; Záhonová, K.; Juricová, V.; Danchenko, M.; Moos, M.; Baráth, P.; Prokopchuk, G.; Butenko, A.; Lukáčová, V.; Kohútová, L.; et al. Highly Flexible Metabolism of the Marine Euglenozoan Protist Diplonema papillatum. BMC Biol. 2021, 19, 251. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant Computational Platform for Mass Spectrometry-based Shotgun Proteomics. Nature 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus Computational Platform for Comprehensive Analysis of (Prote)omics Data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Eolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Teufel, F.; Armenteros, J.J.A.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 6.0 Predicts all Five Types of Signal Peptides Using Protein Language Models. Nat. Biotechnol. 2022, 1–3. [Google Scholar] [CrossRef]
- Whelan, S.; Goldman, N. A General Empirical Model of Protein Evolution Derived from Multiple Protein Families Using a Maximum-likelihood Approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef]
- Musil, M.; Khan, R.T.; Beier, A.; Stourac, J.; Konegger, H.; Damborsky, J.; Bednar, D. FireProtASR: A Web Server for Fully Automated Ancestral Sequence Reconstruction. Brief. Bioinform. 2021, 22, bbaa337. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Stamatakis, A. RaxML Version 8: A Tool for Phylogenetic Analysis and Post-analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Hanson-Smith, V.; Kolaczkowski, B.; Thornton, J.W. Robustness of Ancestral Sequence Reconstruction to Phylogenetic Uncertainity. Mol. Biol. Evol. 2010, 27, 1988–1999. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, D.; Nowotny, J.; Cao, R.; Cheng, J. 3Drefine: An Interactive Web Server for Efficient Protein Structure Refinement. Nucleic Acids Res. 2016, 44, W406–W409. [Google Scholar] [CrossRef] [PubMed]
- Chovancová, E.; Pavelka, A.; Beneš, P.; Strnad, O.; Brezovský, J.; Kozlíková, B.; Gora, A.; Šustr, V.; Klvaňa, M.; Medek, P.; et al. CAVER 3.0: A Tool for the Analysis of Transport Pathways in Dynamic Protein Structures. PLoS Comput. Biol. 2012, 8, e1002708. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Phython bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Adak, S.; Datta, A.K. Leishmania major encodes an unusual peroxidase that is a close homologue of plant ascorbate peroxidase: A novel role of the transmembrane domain. Biochem. J. 2005, 390, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Zámocký, M.; Janeček, Š.; Obinger, C. Fungal Hybrid B Heme Peroxidases-Unique Fusions of a Heme Peroxidase Domain with a Carbohydrate-binding Domain. Sci. Rep. 2017, 7, 9393. [Google Scholar] [CrossRef] [PubMed]
- Taylor, T.N.; Krings, M.; Taylor, E.L. Chytridiomycota. In Fossil Fungi; Elsevier: Amsterdam, The Netherlands, 2015; Chapter 4; pp. 41–67. [Google Scholar] [CrossRef]
- Chang, Y.; Wang, S.; Sekimoto, S.; Aerts, A.L.; Choi, C.; Clum, A.; LaButti, K.M.; Lindquist, E.A.; Ngan, C.Y.; Ohm, R.A.; et al. Phylogenomic Analyses Indicate that Early Fungi Evolved Digesting Cell Walls of Algal Ancestors of Land Plants. Genome Biol. Evol. 2015, 7, 1590–1601. [Google Scholar] [CrossRef]
- Shoff, C.J.; Perfect, J.R. Uncommon Yeasts and Molds Causing Human Disease. In Encyclopedia of Mycology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 1, pp. 813–834. [Google Scholar]
- Morin, E.; Kohler, A.; Baker, A.R.; Foulongne-Oriol, M.; Lombard, V.; Nagy, L.G.; Ohm, R.A.; Patyshakuliyeva, A.; Brun, A.; Aerts, A.L.; et al. Genome sequence of the button mushroom Agaricus bisporus reveals mechanisms governing adaptation to a humic-rich ecological niche. Proc. Natl. Acad. Sci. USA 2012, 10, 17501–17506. [Google Scholar] [CrossRef] [PubMed]
- Hess, J.; Skrede, I.; Wolfe, B.E.; LaButti, K.; Ohm, R.A.; Grigoriev, I.V.; Pringle, A. Transposable Element Dynamics among Asymbiotic and Ectomycorrhizal Amanita Fungi. Genome Biol. Evol. 2014, 6, 1564–1578. [Google Scholar] [CrossRef] [PubMed]
- Zámocký, M.; Hofbauer, S.; Schaffner, I.; Gasselhuber, B.; Nicolussi, A.; Soudi, M.; Pirker, K.F.; Furtmüller, P.G.; Obinger, C. Independent Evolution of Four Heme Peroxidase Superfamilies. Arch. Biochem. Biophys. 2015, 574, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Nigro, A.P.; Goodin, D.B. Reaction of N-hydroxyguanidine with the Ferrous-oxy State of a Heme Peroxidase Cavity Mutant: A Model for the Reactions of Nitric Oxide Synthase. Arch. Biochem. Biophys. 2010, 500, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Howes, B.D.; Feist, A.; Raimondi, L.; Indiani, C.; Smulevich, G. The Critical Role of the Proximal Calcium Ion in the Structural Properties of Horseradish Peroxidase. J. Biol. Chem. 2001, 276, 40704–40711. [Google Scholar] [CrossRef] [PubMed]
- Semba, Y.; Ishida, M.; Yokobori, S.; Yamagishi, A. Ancestral Amino Acid Substitution Improves the Thermal Staiblity of Recombinant Lignin Peroxidase from White-Rot Fungi, Phanerochaete chrysosporium Strain UAMH 3641. Protein Eng. Des. Sel. 2015, 28, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Ayuso-Fernández, I.; Martínez, A.T.; Ruiz-Duenas, F.J. Experimental Recreation of the Evolution of Lignin-degrading Enzymes from the Jurassic to Date. Biotechnol. Biofuels 2017, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Verna, J.; Lodder, A.; Lee, K.; Vagts, A.; Ballester, R. A Family of Genes Required for Maintenance of Cell Wall Integrity and for the Stress Response in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1997, 94, 13804–13809. [Google Scholar] [CrossRef] [PubMed]
- Jasion, V.S.; Polanco, V.A.; Meharenna, Y.T.; Li, H.; Poulos, T.L. Crystal Structure of Leishmania major Peroxidase and Characterization of the Compound I Tryptophan Radical. J. Biol. Chem. 2011, 286, 24608–24615. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Ruiz, M.I.; Ayuso-Fernández, I.; Rencoret, J.; González-Ramirez, A.M.; Linde, D.; Davó-Siguero, I.; Romero, A.; Gutiérrez, A.; Martinez, A.T.; Ruiz-Duenas, F.J. Agaricales Mushroom Lignin Peroxidase: From Structure-Function to Degradative Capabilities. Antioxidants 2021, 10, 1446. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Jiménez, V.; Fernández-Fueyo, E.; Medrano, F.J.; Romero, A.; Martínez, A.T.; Ruiz-Duenas, F.J. Improving the pH-stability of Versatile Peroxidase by Comparative Structural Analysis with a Naturally-Stable Manganese Peroxidase. PLoS ONE 2015, 10, e0140984. [Google Scholar] [CrossRef] [PubMed]
- Zámocký, M.; Garcia-Fernandez, Q.; Gasselhuber, B.; Jakopitsch, C.; Furtmüller, P.G.; Loewen, P.C.; Fita, I.; Obinger, C.; Carpena, X. High Conformational Stability of Secreted Eukaryotic Catalase-peroxidases: Answers from First Crytal Structure and Unfolding Studies. J. Biol. Chem. 2012, 287, 32254–32262. [Google Scholar] [CrossRef] [PubMed]
- Ayuso-Fernández, I.; De Lacey, A.L.; Canada, F.J.; Ruiz-Duenas, F.J.; Martínez, A.T. Increase of Redox Potential during the Evolution of Enzymes Degrading Recalcitrant Lignin. Chem. Eur. J. 2019, 25, 2708–2712. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Throughput | Tunnel Opening | Bottleneck Radius | Length | Curvature |
|---|---|---|---|---|---|
| ScepHyBpox1 | 0.58 | 1.4 | 1.2 | 12.6 | 1.2 |
| Node F | 0.56 | 1.5 | 1.1 | 13.7 | 1.2 |
| Node E | 0.62 | 1.9 | 1.2 | 13.0 | 1.2 |
| Node D | 0.76 | 1.8 | 1.5 | 9.2 | 1.1 |
| Node C | 0.60 | 1.9 | 1.0 | 11.6 | 1.1 |
| Node B | 0.64 | 2.2 | 1.1 | 11.3 | 1.1 |
| Node A | 0.68 | 2.5 | 1.1 | 11.7 | 1.2 |
| Protein | Binding Affinity (kcal/mol) | |
|---|---|---|
| Hydrogen Peroxide | Guaiacol | |
| ScepHyBpox1 | −2.8 | 0.8 |
| Node F | −2.4 | −2.2 |
| Node E | −2.4 | −3.8 |
| Node D | −2.3 | −5.0 |
| Node C | −2.6 | −1.6 |
| Node B | −2.9 | −3.5 |
| Node A | −2.2 | −5.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zámocký, M.; Musil, M.; Danchenko, M.; Ferianc, P.; Chovanová, K.; Baráth, P.; Poljovka, A.; Bednář, D. Deep Insights into the Specific Evolution of Fungal Hybrid B Heme Peroxidases. Biology 2022, 11, 459. https://doi.org/10.3390/biology11030459
Zámocký M, Musil M, Danchenko M, Ferianc P, Chovanová K, Baráth P, Poljovka A, Bednář D. Deep Insights into the Specific Evolution of Fungal Hybrid B Heme Peroxidases. Biology. 2022; 11(3):459. https://doi.org/10.3390/biology11030459
Chicago/Turabian StyleZámocký, Marcel, Miloš Musil, Maksym Danchenko, Peter Ferianc, Katarína Chovanová, Peter Baráth, Andrej Poljovka, and David Bednář. 2022. "Deep Insights into the Specific Evolution of Fungal Hybrid B Heme Peroxidases" Biology 11, no. 3: 459. https://doi.org/10.3390/biology11030459
APA StyleZámocký, M., Musil, M., Danchenko, M., Ferianc, P., Chovanová, K., Baráth, P., Poljovka, A., & Bednář, D. (2022). Deep Insights into the Specific Evolution of Fungal Hybrid B Heme Peroxidases. Biology, 11(3), 459. https://doi.org/10.3390/biology11030459