
Manipulation of Meiotic Recombination to Hasten Crop Improvement

 ,
, {kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
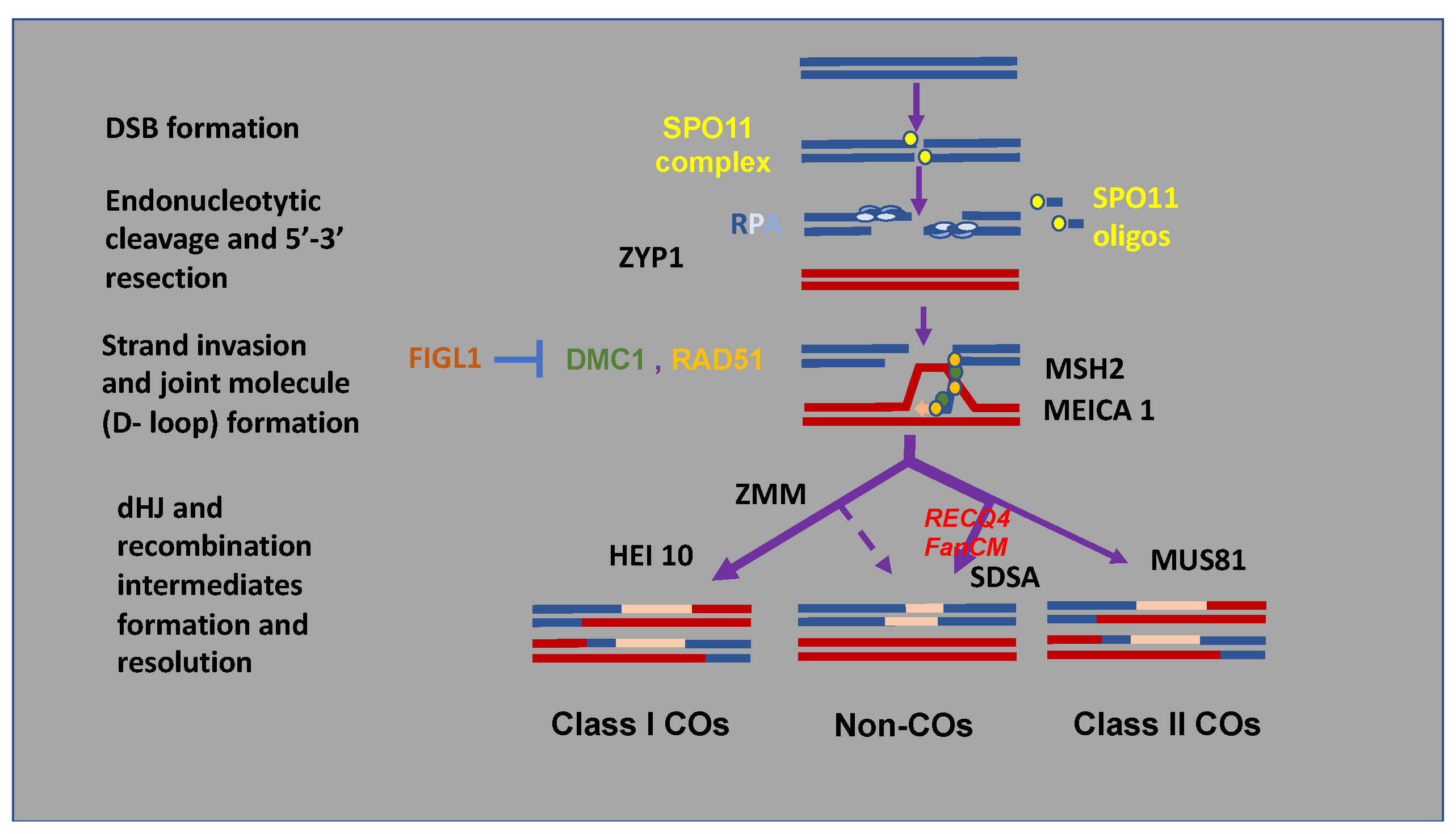
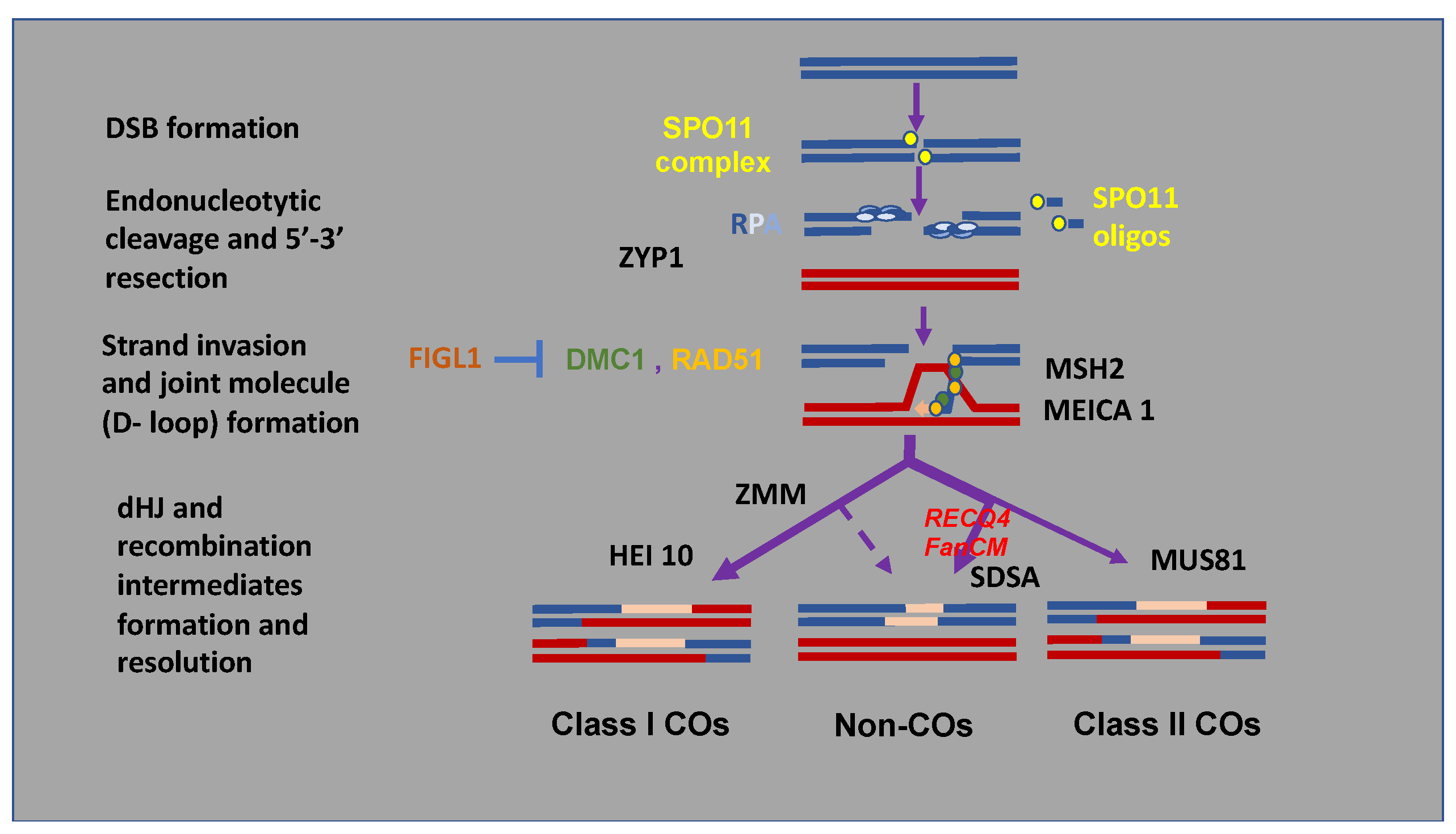
2. Control of Meiotic Recombination
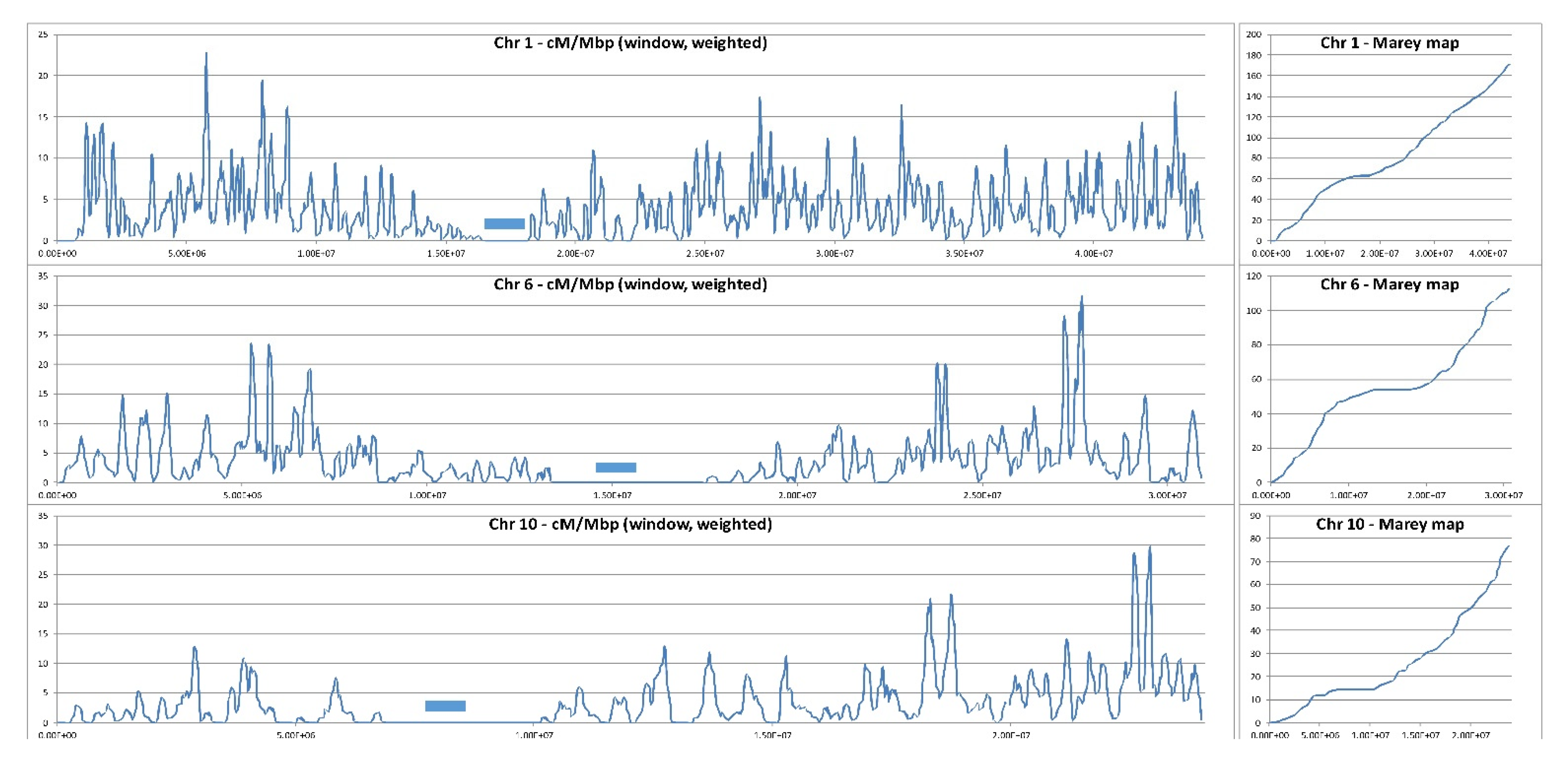
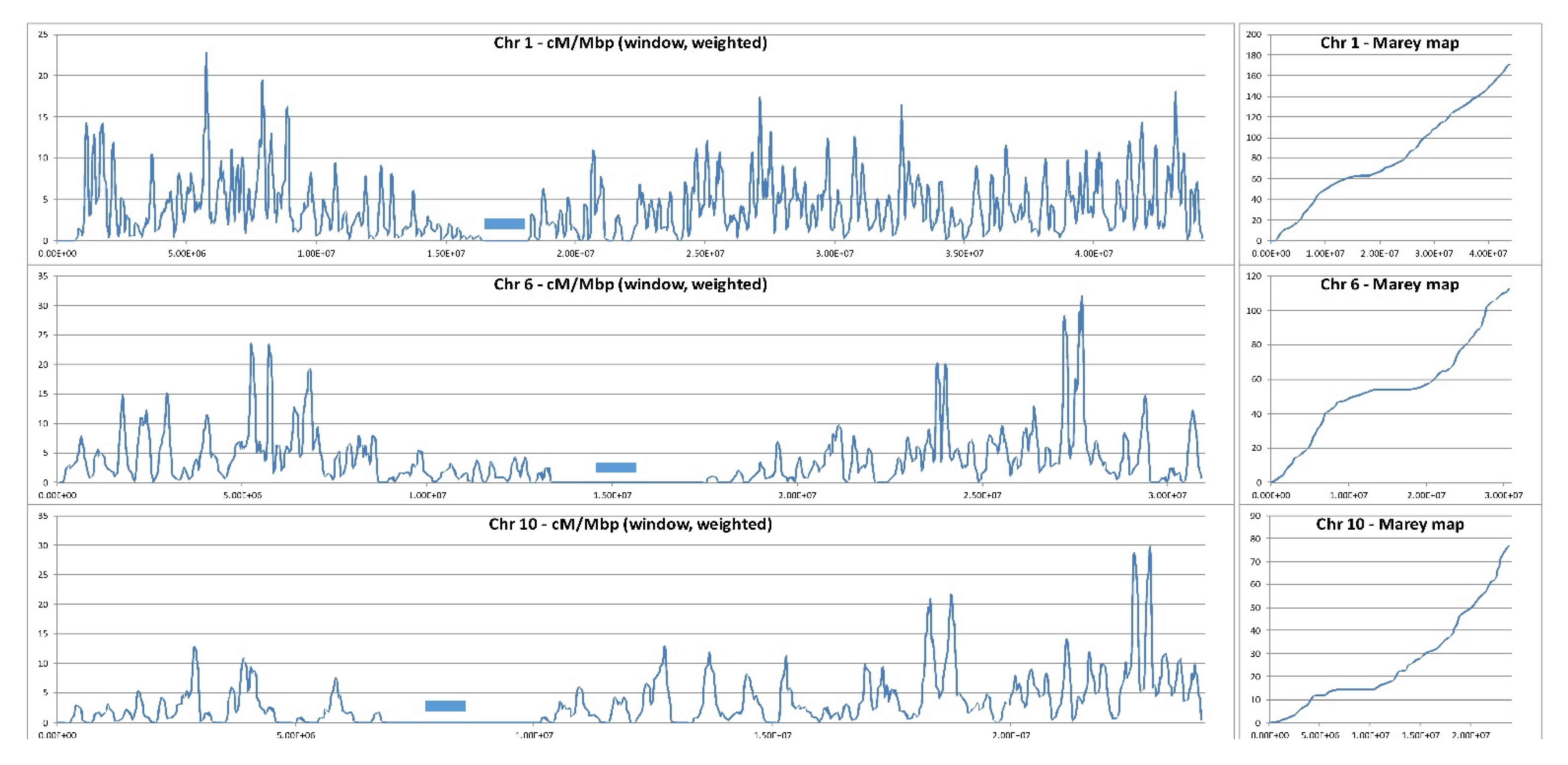
3. Stimulation and Redistribution of Meiotic Recombination
3.1. Modulation by Sex
3.2. Modulation by Environmental Conditions
3.3. Modulation by Novel Genomic Situation
3.4. Modulation by Epigenetic Factors
3.5. Modulation by Altering the Expression of Genes Involved in Homolog Synapsis and DNA Repair
3.6. Interest in Stimulating and Redistributing Recombination in Breeding
4. Targeting of Meiotic Recombination
4.1. Targeting Somatic COs with CRISPR/Cas9
4.2. Targeting Meiotic COs with Partners of the DSB Catalytic Complex
4.3. Interest in Targeting Recombination in Breeding
5. Conclusions and Prospects
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, Z.; Wang, W.; Wu, Z.; Sun, C.; Li, M.; Lu, J.; Fu, B.; Shi, J.; Xu, J.; Ruan, J.; et al. Novel sequences, structural variations and gene presence variations of Asian cultivated rice. Sci. Data 2018, 5, 180079. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Lu, H.; Du, H.; Wang, H.; Chen, W.; Chen, Z.; He, Q.; Ou, S.; Zhang, H.; Li, X.; et al. Pan-genome analysis of 33 genetically diverse rice accessions reveals hidden genomic variations. Cell 2021, 184, 3542–3558.e16. [Google Scholar] [CrossRef]
- He, Y.; Wang, M.; Dukowic-Schulze, S.; Zhou, A.; Tiang, C.-L.; Shilo, S.; Sidhu, G.K.; Eichten, S.; Bradbury, P.; Springer, N.M.; et al. Genomic features shaping the landscape of meiotic double-strand-break hotspots in maize. Proc. Natl. Acad. Sci. USA 2017, 114, 12231–12236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.; Zhao, X.; Kelly, K.A.; Venn, O.; Higgins, J.; Yelina, N.E.; Hardcastle, T.J.; Ziolkowski, P.; Copenhaver, G.; Franklin, F.C.H.; et al. Arabidopsis meiotic crossover hot spots overlap with H2A.Z nucleosomes at gene promoters. Nat. Genet. 2013, 45, 1327–1336. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.; Henderson, I. Meiotic recombination hotspots - a comparative view. Plant J. 2015, 83, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Mizuno, H.; Tsugane, M.; Ito, Y.; Chiden, Y.; Fujisawa, M.; Katagiri, S.; Saji, S.; Yoshiki, S.; Karasawa, W.; et al. Physical maps and recombination frequency of six rice chromosomes. Plant J. 2003, 36, 720–730. [Google Scholar] [CrossRef] [PubMed]
- Choulet, F.; Alberti, A.; Theil, S.; Glover, N.; Barbe, V.; Daron, J.; Pingault, L.; Sourdille, P.; Couloux, A.; Paux, E.; et al. Structural and functional partitioning of bread wheat chromosome 3B. Science 2014, 345, 1249721. [Google Scholar] [CrossRef] [PubMed]
- Mercier, R.; Mézard, C.; Jenczewski, E.; Macaisne, N.; Grelon, M. The Molecular Biology of Meiosis in Plants. Annu. Rev. Plant Biol. 2015, 66, 297–327. [Google Scholar] [CrossRef]
- Yadav, V.K.; Bouuaert, C.C. Mechanism and Control of Meiotic DNA Double-Strand Break Formation in S. cerevisiae. Front. Cell Dev. Biol. 2021, 9, 287. [Google Scholar] [CrossRef]
- Crismani, W.; Girard, C.; Froger, N.; Pradillo, M.; Santos, J.L.; Chelysheva, L.; Copenhaver, G.P.; Horlow, C.; Mercier, R. FANCM Limits Meiotic Crossovers. Science 2012, 336, 1588–1590. [Google Scholar] [CrossRef] [Green Version]
- Séguéla-Arnaud, M.; Crismani, W.; Larchevêque, C.; Mazel, J.; Froger, N.; Choinard, S.; Lemhemdi, A.; Macaisne, N.; Van Leene, J.; Gevaert, K.; et al. Multiple mechanisms limit meiotic crossovers: TOP3α and two BLM homologs antagonize crossovers in parallel to FANCM. Proc. Natl. Acad. Sci. USA 2015, 112, 4713–4718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Copenhaver, G.P. Meiotic Recombination: Mixing It Up in Plants. Annu. Rev. Plant Biol. 2018, 69, 577–609. [Google Scholar] [CrossRef] [PubMed]
- Berchowitz, L.E.; Copenhaver, G. Genetic Interference: Dont Stand So Close to Me. Curr. Genom. 2010, 11, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Pinzón, Y.G.; Kise, J.K.G.; Rueda, P.; Ronceret, A. The Formation of Bivalents and the Control of Plant Meiotic Recombination. Front. Plant Sci. 2021, 12, 1919. [Google Scholar] [CrossRef]
- Choi, K.; Zhao, X.; Tock, A.J.; Lambing, C.; Underwood, C.J.; Hardcastle, T.J.; Serra, H.; Kim, J.; Cho, H.S.; Kim, J.; et al. Nucleosomes and DNA methylation shape meiotic DSB frequency in Arabidopsis thaliana transposons and gene regulatory regions. Genome Res. 2018, 28, 532–546. [Google Scholar] [CrossRef] [Green Version]
- Wijnker, E.; James, G.V.; Ding, J.; Becker, F.; Klasen, J.R.; Rawat, V.; Rowan, B.; De Jong, D.F.; De Snoo, C.B.; Zapata, L.; et al. The genomic landscape of meiotic crossovers and gene conversions in Arabidopsis thaliana. eLife 2013, 2, e01426. [Google Scholar] [CrossRef] [PubMed]
- Shilo, S.; Melamed-Bessudo, C.; Dorone, Y.; Barkai, N.; Levy, A.A. DNA Crossover Motifs Associated with Epigenetic Modifications Delineate Open Chromatin Regions in Arabidopsis. Plant Cell 2015, 27, 2427–2436. [Google Scholar] [CrossRef] [Green Version]
- Si, W.; Yuan, Y.; Huang, J.; Zhang, X.; Zhang, Y.; Zhang, Y.; Tian, D.; Wang, C.; Yang, Y.; Yang, S. Widely distributed hot and cold spots in meiotic recombination as shown by the sequencing of rice F 2 plants. New Phytol. 2015, 206, 1491–1502. [Google Scholar] [CrossRef]
- Demirci, S.; van Dijk, A.-J.; Perez, G.S.; Aflitos, S.; De Ridder, D.; Peters, S.A. Distribution, position and genomic characteristics of crossovers in tomato recombinant inbred lines derived from an interspecific cross betweenSolanum lycopersicumandSolanum pimpinellifolium. Plant J. 2017, 89, 554–564. [Google Scholar] [CrossRef] [Green Version]
- Golicz, A.A.; Bhalla, P.L.; Edwards, D.; Singh, M.B. Rice 3D chromatin structure correlates with sequence variation and meiotic recombination rate. Commun. Biol. 2020, 3, 1–9. [Google Scholar] [CrossRef]
- Marand, A.P.; Zhao, H.; Zhang, W.; Zeng, Z.; Fang, C.; Jiang, J. Historical Meiotic Crossover Hotspots Fueled Patterns of Evolutionary Divergence in Rice. Plant Cell 2019, 31, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Farmer, A.D.; Langley, R.J.; Mudge, J.; Crow, J.A.; May, G.D.; Huntley, J.; Smith, A.G.; Retzel, E.F. Meiosis-specific gene discovery in plants: RNA-Seq applied to isolated Arabidopsis male meiocytes. BMC Plant Biol. 2010, 10, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, J.; Gao, H.; Zhang, J.; Aldridge, B.; Vickers, M.; Higgins, J.D.; Feng, X. Sexual-lineage-specific DNA methylation regulates meiosis in Arabidopsis. Nat. Genet. 2017, 50, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Tock, A.J.; Henderson, I.R. Hotspots for Initiation of Meiotic Recombination. Front. Genet. 2018, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Lambing, C.; Franklin, F.C.H.; Wang, C.-J.R. Understanding and Manipulating Meiotic Recombination in Plants. Plant Physiol. 2017, 173, 1530–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, Y.; Liu, Y.-G.; Zhang, Q. Hybrid sterility in plant: Stories from rice. Curr. Opin. Plant Biol. 2010, 13, 186–192. [Google Scholar] [CrossRef]
- Drouaud, J.; Mercier, R.; Chelysheva, L.; Bérard, A.; Falque, M.; Martin, O.; Zanni, V.; Brunel, D.; Mézard, C. Sex-Specific Crossover Distributions and Variations in Interference Level along Arabidopsis thaliana Chromosome 4. PLoS Genet. 2007, 3, e106. [Google Scholar] [CrossRef]
- Giraut, L.; Falque, M.; Drouaud, J.; Pereira, L.; Martin, O.C.; Mézard, C. Genome-Wide Crossover Distribution in Arabidopsis thaliana Meiosis Reveals Sex-Specific Patterns along Chromosomes. PLoS Genet. 2011, 7, e1002354. [Google Scholar] [CrossRef] [Green Version]
- Phillips, D.; Jenkins, G.; Macaulay, M.; Nibau, C.; Wnetrzak, J.; Fallding, D.; Colas, I.; Oakey, H.; Waugh, R.; Ramsay, L. The effect of temperature on the male and female recombination landscape of barley. New Phytol. 2015, 208, 421–429. [Google Scholar] [CrossRef]
- Luo, C.; Li, X.; Zhang, Q.; Yan, J. Single gametophyte sequencing reveals that crossover events differ between sexes in maize. Nat. Commun. 2019, 10, 1–8. [Google Scholar] [CrossRef]
- Kianian, P.M.A.; Wang, M.; Simons, K.; Ghavami, F.; He, Y.; Dukowic-Schulze, S.; Sundararajan, A.; Sun, Q.; Pillardy, J.; Mudge, J.; et al. High-resolution crossover mapping reveals similarities and differences of male and female recombination in maize. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Modliszewski, J.L.; Copenhaver, G. Meiotic recombination gets stressed out: CO frequency is plastic under pressure. Curr. Opin. Plant Biol. 2017, 36, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.; Perry, R.M.; Barakate, A.; Ramsay, L.; Waugh, R.; Halpin, C.; Armstrong, S.J.; Franklin, F.C.H. Spatiotemporal Asymmetry of the Meiotic Program Underlies the Predominantly Distal Distribution of Meiotic Crossovers in Barley. Plant Cell 2012, 24, 4096–4109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, A.; Morgan, C.; Franklin, F.C.H.; Bomblies, K. Plasticity of Meiotic Recombination Rates in Response to Temperature in Arabidopsis. Genetics 2018, 208, 1409–1420. [Google Scholar] [CrossRef] [Green Version]
- Modliszewski, J.L.; Wang, H.; Albright, A.R.; Lewis, S.M.; Bennett, A.R.; Huang, J.; Ma, H.; Wang, Y.; Copenhaver, G.P. Elevated temperature increases meiotic crossover frequency via the interfering (Type I) pathway in Arabidopsis thaliana. PLoS Genet. 2018, 14, e1007384. [Google Scholar] [CrossRef] [PubMed]
- Capilla-Pérez, L.; Durand, S.; Hurel, A.; Lian, Q.; Chambon, A.; Taochy, C.; Solier, V.; Grelon, M.; Mercier, R. The synaptonemal complex imposes crossover interference and heterochiasmy inArabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, 2023613118. [Google Scholar] [CrossRef]
- Leflon, M.; Grandont, L.; Eber, F.; Huteau, V.; Coriton, O.; Chelysheva, L.; Jenczewski, E.; Chèvre, A.-M. Crossovers Get a Boost inBrassicaAllotriploid and Allotetraploid Hybrids. Plant Cell 2010, 22, 2253–2264. [Google Scholar] [CrossRef] [Green Version]
- Pelé, A.; Falque, M.; Trotoux, G.; Eber, F.; Negre, S.; Gilet, M.; Huteau, V.; Lodé, M.; Jousseaume, T.; Dechaumet, S.; et al. Amplifying recombination genome-wide and reshaping crossover landscapes in Brassicas. PLoS Genet. 2017, 13, e1006794. [Google Scholar] [CrossRef]
- Mirouze, M.; Lieberman-Lazarovich, M.; Aversano, R.; Bucher, E.; Nicolet, J.; Reinders, J.; Paszkowski, J. Loss of DNA methylation affects the recombination landscape in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 5880–5885. [Google Scholar] [CrossRef] [Green Version]
- Yelina, N.E.; Choi, K.; Chelysheva, L.; Macaulay, M.; de Snoo, B.; Wijnker, E.; Miller, N.; Drouaud, J.; Grelon, M.; Copenhaver, G.P.; et al. Epigenetic Remodeling of Meiotic Crossover Frequency in Arabidopsis thaliana DNA Methyltransferase Mutants. PLoS Genet. 2012, 8, e1002844. [Google Scholar] [CrossRef] [Green Version]
- Yelina, N.; Diaz, P.; Lambing, C.; Henderson, I.R. Epigenetic control of meiotic recombination in plants. Sci. China Life Sci. 2015, 58, 223–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underwood, C.J.; Choi, K.; Lambing, C.; Zhao, X.; Serra, H.; Borges, F.; Simorowski, J.; Ernst, E.; Jacob, Y.; Henderson, I.R.; et al. Epigenetic activation of meiotic recombination near Arabidopsis thaliana centromeres via loss of H3K9me2 and non-CG DNA methylation. Genome Res. 2018, 28, 519–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yelina, N.E.; Lambing, C.; Hardcastle, T.J.; Zhao, X.; Santos, B.; Henderson, I.R. DNA methylation epigenetically silences crossover hot spots and controls chromosomal domains of meiotic recombination in Arabidopsis. Genes Dev. 2015, 29, 2183–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Wang, C.; Wang, H.; Lu, P.; Zheng, B.; Ma, H.; Copenhaver, G.P.; Wang, Y. Meiocyte-Specific and AtSPO11-1–Dependent Small RNAs and Their Association with Meiotic Gene Expression and Recombination. Plant Cell 2019, 31, 444–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Wang, C.; Liu, Q.; Liu, W.; Fu, Y. Increasing the Genetic Recombination Frequency by Partial Loss of Function of the Synaptonemal Complex in Rice. Mol. Plant 2015, 8, 1295–1298. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Cao, Y.; Hua, Y.; Du, G.; Liu, Q.; Wei, X.; Sun, T.; Lin, J.; Wu, M.; Cheng, Z.; et al. Concurrent Disruption of Genetic Interference and Increase of Genetic Recombination Frequency in Hybrid Rice Using CRISPR/Cas9. Front. Plant Sci. 2021, 12, 2182. [Google Scholar] [CrossRef]
- Girard, C.; Chelysheva, L.; Choinard, S.; Froger, N.; Macaisne, N.; Lehmemdi, A.; Mazel, J.; Crismani, W.; Mercier, R. AAA-ATPase FIDGETIN-LIKE 1 and Helicase FANCM Antagonize Meiotic Crossovers by Distinct Mechanisms. PLoS Genet. 2015, 11, e1005369. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.B.; Séguéla-Arnaud, M.; Larchevêque, C.; Lloyd, A.H.; Mercier, R. Unleashing meiotic crossovers in hybrid plants. Proc. Natl. Acad. Sci. USA 2018, 115, 2431–2436. [Google Scholar] [CrossRef] [Green Version]
- Mieulet, D.; Aubert, G.; Bres, C.; Klein, A.; Droc, G.; Vieille, E.; Rond-Coissieux, C.; Signor, C.L.; Dalmais, M.; Mauxion, J.-P.; et al. Unleashing meiotic crossovers in crops. Nat. Plants 2018, 4, 1010–1016. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Zhang, Y.; Sun, L.; Sinumporn, S.; Yang, Z.; Sun, B.; Xuan, D.; Li, Z.; Yu, P.; Wu, W.; et al. The Rice AAA-ATPase OsFIGNL1 Is Essential for Male Meiosis. Front. Plant Sci. 2017, 8, 1639. [Google Scholar] [CrossRef]
- Ziolkowski, P.A.; Underwood, C.J.; Lambing, C.; Martinez-Garcia, M.; Lawrence, E.J.; Ziolkowska, L.; Griffin, C.; Choi, K.; Franklin, F.C.H.; Martienssen, R.A.; et al. Natural variation and dosage of the HEI10 meiotic E3 ligase control Arabidopsis crossover recombination. Genes Dev. 2017, 31, 306–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, H.; Lambing, C.; Griffin, C.H.; Topp, S.D.; Nageswaran, D.C.; Underwood, C.J.; Ziolkowski, P.A.; Séguéla-Arnaud, M.; Fernandes, J.B.; Mercier, R.; et al. Massive crossover elevation via combination of HEI10 and recq4a recq4b during Arabidopsis meiosis. Proc. Natl. Acad. Sci. USA 2018, 115, 2437–2442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blary, A.; Gonzalo, A.; Eber, F.; Bérard, A.; Bergès, H.; Bessoltane, N.; Charif, D.; Charpentier, C.; Cromer, L.; Fourment, J.; et al. FANCM Limits Meiotic Crossovers in Brassica Crops. Front. Plant Sci. 2018, 9, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Yu, M.; Bolaños-Villegas, P.; Zhang, J.; Ni, D.; Ma, H.; Wang, Y. Fanconi anemia ortholog FANCM regulates meiotic crossover distribution in plants. Plant Physiol. 2021, 186, 344–360. [Google Scholar] [CrossRef]
- De Maagd, R.A.; Loonen, A.; Chouaref, J.; Pelé, A.; Meijer-Dekens, F.; Fransz, P.; Bai, Y. CRISPR/Cas inactivation of RECQ4 increases homeologous crossovers in an interspecific tomato hybrid. Plant Biotechnol. J. 2019, 18, 805–813. [Google Scholar] [CrossRef] [Green Version]
- Arrieta, M.; Macaulay, M.; Colas, I.; Schreiber, M.; Shaw, P.D.; Waugh, R.; Ramsay, L. An Induced Mutation in HvRECQL4 Increases the Overall Recombination and Restores Fertility in a Barley HvMLH3 Mutant Background. Front. Plant Sci. 2021, 12, 2358. [Google Scholar] [CrossRef]
- Blackwell, A.R.; Dluzewska, J.; Szymanska-Lejman, M.; Desjardins, S.; Tock, A.J.; Kbiri, N.; Lambing, C.; Lawrence, E.J.; Bieluszewski, T.; Rowan, B.; et al. MSH 2 shapes the meiotic crossover landscape in relation to interhomolog polymorphism in Arabidopsis. EMBO J. 2020, 39, e104858. [Google Scholar] [CrossRef]
- Kunkel, T.A.; Erie, D.A. Eukaryotic Mismatch Repair in Relation to DNA Replication. Annu. Rev. Genet. 2015, 49, 291–313. [Google Scholar] [CrossRef] [Green Version]
- Myung, K.; Datta, A.; Chen, C.; Kolodner, R.D. SGS1, the Saccharomyces cerevisiae homologue of BLM and WRN, suppresses genome instability and homeologous recombination. Nat. Genet. 2001, 27, 113–116. [Google Scholar] [CrossRef]
- Mazina, O.M.; Mazin, A.V.; Nakagawa, T.; Kolodner, R.D.; Kowalczykowski, S.C. Saccharomyces cerevisiae Mer3 Helicase Stimulates 3′–5′ Heteroduplex Extension by Rad51: Implications for Crossover Control in Meiotic Recombination. Cell 2004, 117, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Emmanuel, E.; Yehuda, E.; Melamed-Bessudo, C.; Avivi-Ragolsky, N.; Levy, A.A. The role ofAtMSH2in homologous recombination inArabidopsis thaliana. EMBO Rep. 2006, 7, 100–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Li, Y.; Wang, H.; Shen, Y.; Zhang, C.; Du, G.; Tang, D.; Cheng, Z. Meiotic Chromosome Association 1 Interacts with TOP3α and Regulates Meiotic Recombination in Rice. Plant Cell 2017, 29, 1697–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, H.; Svačina, R.; Baumann, U.; Whitford, R.; Sutton, T.; Bartoš, J.; Sourdille, P. Ph2 encodes the mismatch repair protein MSH7-3D that inhibits wheat homoeologous recombination. Nat. Commun. 2021, 12, 1–10. [Google Scholar] [CrossRef]
- Miao, Y.; Shi, W.; Wang, H.; Xue, Z.; You, H.; Zhang, F.; Du, G.; Tang, D.; Li, Y.; Shen, Y.; et al. Replication protein A large subunit (RPA1a) limits chiasma formation during rice meiosis. Plant Physiol. 2021, 187, 1605–1618. [Google Scholar] [CrossRef]
- Tourrette, E.; Bernardo, R.; Falque, M.; Martin, O.C. Assessing by Modeling the Consequences of Increased Recombination in Recurrent Selection of Oryza sativa and Brassica rapa. G3 Genes|Genomes|Genetics 2019, 9, 4169–4181. [Google Scholar] [CrossRef] [Green Version]
- Boideau, F.; Pelé, A.; Tanguy, C.; Trotoux, G.; Eber, F.; Maillet, L.; Gilet, M.; Lodé-Taburel, M.; Huteau, V.; Morice, J.; et al. A Modified Meiotic Recombination in Brassica napus Largely Improves Its Breeding Efficiency. Biology 2021, 10, 771. [Google Scholar] [CrossRef]
- Sadhu, M.J.; Bloom, J.S.; Day, L.; Kruglyak, L. CRISPR-directed mitotic recombination enables genetic mapping without crosses. Science 2016, 352, 1113–1116. [Google Scholar] [CrossRef] [Green Version]
- Filler-Hayut, S.; Bessudo, C.M.; Levy, A.A. Targeted recombination between homologous chromosomes for precise breeding in tomato. Nat. Commun. 2017, 8, 15605. [Google Scholar] [CrossRef]
- Filler-Hayut, S.; Kniazev, K.; Melamed-Bessudo, C.; Levy, A.A. Targeted Inter-Homologs Recombination in Arabidopsis Euchromatin and Heterochromatin. Int. J. Mol. Sci. 2021, 22, 12096. [Google Scholar] [CrossRef]
- Kouranov, A.; Armstrong, C.; Shrawat, A.; Sidorov, V.; Huesgen, S.; Lemke, B.; Boyle, T.; Gasper, M.; Lawrence, R.; Yang, S. Demonstration of targeted crossovers in hybrid maize using CRISPR technology. Commun. Biol. 2022, 5, 1–11. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, Z.; Feng, Z.; Wei, P.; Zhang, H.; Botella, J.; Zhu, J.-K. Development of germ-line-specific CRISPR-Cas9 systems to improve the production of heritable gene modifications inArabidopsis. Plant Biotechnol. J. 2016, 14, 519–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eid, A.; Ali, Z.; Mahfouz, M.M. High efficiency of targeted mutagenesis in arabidopsis via meiotic promoter-driven expression of Cas9 endonuclease. Plant Cell Rep. 2016, 35, 1555–1558. [Google Scholar] [CrossRef] [PubMed]
- Bergerat, A.; De Massy, B.; Gadelle, D.; Varoutas, P.-C.; Nicolas, A.; Forterre, P. An atypical topoisomerase II from archaea with implications for meiotic recombination. Nature 1997, 386, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Neale, M.J.; Keeney, S. Clarifying the mechanics of DNA strand exchange in meiotic recombination. Nature 2006, 442, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Bouuaert, C.C.; Tischfield, S.E.; Pu, S.; Mimitou, E.P.; Arias-Palomo, E.; Berger, J.M.; Keeney, S. Structural and functional characterization of the Spo11 core complex. Nat. Struct. Mol. Biol. 2021, 28, 92–102. [Google Scholar] [CrossRef]
- Sprink, T.; Hartung, F. The splicing fate of plant SPO11 genes. Front. Plant Sci. 2014, 5, 214. [Google Scholar] [CrossRef] [Green Version]
- Grelon, M.; Vezon, D.; Gendrot, G.; Pelletier, G. AtSPO11-1 is necessary for efficient meiotic recombination in plants. EMBO J. 2001, 20, 589–600. [Google Scholar] [CrossRef] [Green Version]
- Hartung, F. Molecular characterisation of two paralogous SPO11 homologues in Arabidopsis thaliana. Nucleic Acids Res. 2000, 28, 1548–1554. [Google Scholar] [CrossRef] [Green Version]
- Hartung, F.; Angelis, K.; Meister, A.; Schubert, I.; Melzer, M.; Puchta, H. An Archaebacterial Topoisomerase Homolog Not Present in Other Eukaryotes Is Indispensable for Cell Proliferation of Plants. Curr. Biol. 2002, 12, 1787–1791. [Google Scholar] [CrossRef] [Green Version]
- Vrielynck, N.; Chambon, A.; Vezon, D.; Pereira, L.; Chelysheva, L.; De Muyt, A.; Mézard, C.; Mayer, C.; Grelon, M. A DNA topoisomerase VI–like complex initiates meiotic recombination. Science 2016, 351, 939–943. [Google Scholar] [CrossRef]
- Robine, N.; Uematsu, N.; Amiot, F.; Gidrol, X.; Barillot, E.; Nicolas, A.; Borde, V. Genome-Wide Redistribution of Meiotic Double-Strand Breaks in Saccharomyces cerevisiae. Mol. Cell. Biol. 2007, 27, 1868–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peciña, A.; Smith, K.N.; Mézard, C.; Murakami, H.; Ohta, K.; Nicolas, A. Targeted Stimulation of Meiotic Recombination. Cell 2002, 111, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Murakami, H.; Nicolas, A. Locally, Meiotic Double-Strand Breaks Targeted by Gal4BD-Spo11 Occur at Discrete Sites with a Sequence Preference. Mol. Cell. Biol. 2009, 29, 3500–3516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarno, R.; Vicq, Y.; Uematsu, N.; Luka, M.; Lapierre, C.; Carroll, D.; Bastianelli, G.; Serero, A.; Nicolas, A. Programming sites of meiotic crossovers using Spo11 fusion proteins. Nucleic Acids Res. 2017, 45, e164. [Google Scholar] [CrossRef] [Green Version]
- Yelina, N.E.; Gonzalez-Jorge, S.; Hirsz, D.; Yang, Z.; Henderson, I.R. CRISPR Targeting of MEIOTIC-TOPOISOMERASE VIB-DCas9 to a Recombination Hotspot Is Insufficient to Increase Crossover Frequency in Arabidopsis. bioRxiv 2021, 2021, 429210. [Google Scholar] [CrossRef]
- Vrielynck, N.; Schneider, K.; Rodriguez, M.; Sims, J.; Chambon, A.; Hurel, A.; De Muyt, A.; Ronceret, A.; Krsicka, O.; Mézard, C.; et al. Conservation and divergence of meiotic DNA double strand break forming mechanisms in Arabidopsis thaliana. Nucleic Acids Res. 2021, 49, 9821–9835. [Google Scholar] [CrossRef]
- Pan, C.; Sretenovic, S.; Qi, Y. CRISPR/dCas-mediated transcriptional and epigenetic regulation in plants. Curr. Opin. Plant Biol. 2021, 60, 101980. [Google Scholar] [CrossRef]
- Vikram, P.; Swamy, B.P.M.; Dixit, S.; Singh, R.; Singh, B.P.; Miro, B.; Kohli, A.; Henry, A.; Singh, N.K.; Kumar, A. Drought susceptibility of modern rice varieties: An effect of linkage of drought tolerance with undesirable traits. Sci. Rep. 2015, 5, 14799. [Google Scholar] [CrossRef] [Green Version]
- Voss-Fels, K.P.; Qian, L.; Parra-Londono, S.; Uptmoor, R.; Frisch, M.; Keeble-Gagnère, G.; Appels, R.; Snowdon, R.J. Linkage drag constrains the roots of modern wheat. Plant Cell Environ. 2016, 40, 717–725. [Google Scholar] [CrossRef]
- Luo, X.; Ji, S.-D.; Yuan, P.-R.; Lee, H.-S.; Kim, D.-M.; Balkunde, S.; Kang, J.-W.; Ahn, S.-N. QTL mapping reveals a tight linkage between QTLs for grain weight and panicle spikelet number in rice. Rice 2013, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.-Q.; Fan, Y.-Y.; Chen, J.; Shi, Y.-F.; Wu, J.-L. Avoidance of Linkage Drag Between Blast Resistance Gene and the QTL Conditioning Spikelet Fertility Based on Genotype Selection Against Heading Date in Rice. Rice Sci. 2009, 16, 21–26. [Google Scholar] [CrossRef]
- Bernardo, R. Prospective Targeted Recombination and Genetic Gains for Quantitative Traits in Maize. Plant Genome 2017, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandariz, S.P.; Bernardo, R. Predicted Genetic Gains from Targeted Recombination in Elite Biparental Maize Populations. Plant Genome 2019, 12, 180062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ru, S.; Bernardo, R. Targeted recombination to increase genetic gain in self-pollinated species. Theor. Appl. Genet. 2018, 132, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Taagen, E.; Bogdanove, A.J.; Sorrells, M.E. Counting on Crossovers: Controlled Recombination for Plant Breeding. Trends Plant Sci. 2020, 25, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Morgan, C.; White, M.A.; Franklin, F.C.H.; Zickler, D.; Kleckner, N.; Bomblies, K. Evolution of crossover interference enables stable autopolyploidy by ensuring pairwise partner connections in Arabidopsis arenosa. Curr. Biol. 2021, 31, 4713–4726. [Google Scholar] [CrossRef]
- Schmidt, C.; Fransz, P.; Rönspies, M.; Dreissig, S.; Fuchs, J.; Heckmann, S.; Houben, A.; Puchta, H. Changing local recombination patterns in Arabidopsis by CRISPR/Cas mediated chromosome engineering. Nat. Commun. 2020, 11, 1–8. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fayos, I.; Frouin, J.; Meynard, D.; Vernet, A.; Herbert, L.; Guiderdoni, E. Manipulation of Meiotic Recombination to Hasten Crop Improvement. Biology 2022, 11, 369. https://doi.org/10.3390/biology11030369
Fayos I, Frouin J, Meynard D, Vernet A, Herbert L, Guiderdoni E. Manipulation of Meiotic Recombination to Hasten Crop Improvement. Biology. 2022; 11(3):369. https://doi.org/10.3390/biology11030369
Chicago/Turabian StyleFayos, Ian, Julien Frouin, Donaldo Meynard, Aurore Vernet, Léo Herbert, and Emmanuel Guiderdoni. 2022. "Manipulation of Meiotic Recombination to Hasten Crop Improvement" Biology 11, no. 3: 369. https://doi.org/10.3390/biology11030369
APA StyleFayos, I., Frouin, J., Meynard, D., Vernet, A., Herbert, L., & Guiderdoni, E. (2022). Manipulation of Meiotic Recombination to Hasten Crop Improvement. Biology, 11(3), 369. https://doi.org/10.3390/biology11030369