
Viral and Host Genetic and Epigenetic Biomarkers Related to SARS-CoV-2 Cell Entry, Infection Rate, and Disease Severity


Abstract
:Simple Summary
Abstract
1. Introduction
2. Virus Mutations
2.1. Virus Strains
2.2. Virus Mutations Position and Their Influence on SARS 2 Disease Development
3. Virus–Host Interactions Affecting Viral Replication and Transcription
4. Human Polymorphism May Influence SARS-CoV-2 Viral Entry and Replication
Engineered ACE2 Mutations May Predict Virus–Host Interactions
5. Changes in mRNA Expression—miRNA ‘’Silencing’’ Interference
Changes in DNA Methylation Profile
6. Extracellular Vesicles as Biomarkers
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silhol, F.; Marlinge, M.; Guiol, C.; Chefrour, M.; Mace, P.; Criado, C.; Kipson, N.; Vaisse, B.; Vairo, D.; Sarlon, G.; et al. Characterization of adenosine A2 receptors in peripheral blood mononuclear cells of patients with fibromuscular dysplasia. Hypertens Res. 2020, 43, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef] [PubMed]
- Bestle, D.; Heindl, M.R.; Limburg, H.; Van Lam van, T.; Pilgram, O.; Moulton, H.; Stein, D.A.; Hardes, K.; Eickmann, M.; Dolnik, O.; et al. TMPRSS2, and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life Sci. Alliance 2020, 3, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Pohlmann, S. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Mol. Cell 2020, 78, 779–784.e5. [Google Scholar] [CrossRef]
- Sarker, J.; Das, P.; Sarker, S.; Roy, A.K.; Momen, A. A Review on Expression, Pathological Roles, and Inhibition of TMPRSS2, the Serine Protease Responsible for SARS-CoV-2 Spike Protein Activation. Scientifica 2021, 2021, 2706789. [Google Scholar] [CrossRef]
- Zhang, L.; Richards, A.; Barrasa, M.I.; Hughes, S.H.; Young, R.A.; Jaenisch, R. Reverse-transcribed SARS-CoV-2 RNA can integrate into the genome of cultured human cells and can be expressed in patient-derived tissues. Proc. Natl. Acad. Sci. USA 2021, 118, e2105968118. [Google Scholar] [CrossRef] [PubMed]
- Saraste, J.; Prydz, K. Assembly and Cellular Exit of Coronaviruses: Hijacking an Unconventional Secretory Pathway from the Pre-Golgi Intermediate Compartment via the Golgi Ribbon to the Extracellular Space. Cells 2021, 10, 503. [Google Scholar] [CrossRef]
- Kadam, S.B.; Sukhramani, G.S.; Bishnoi, P.; Pable, A.A.; Barvkar, V.T. SARS-CoV-2, the pandemic coronavirus: Molecular and structural insights. J. Basic. Microbiol. 2021, 61, 180–202. [Google Scholar] [CrossRef]
- Liu, J.; Li, Y.; Liu, Q.; Yao, Q.; Wang, X.; Zhang, H.; Chen, R.; Ren, L.; Min, J.; Deng, F.; et al. SARS-CoV-2 cell tropism and multiorgan infection. Cell Discov. 2021, 7, 17. [Google Scholar] [CrossRef]
- Ricci, D.; Etna, M.P.; Rizzo, F.; Sandini, S.; Severa, M.; Coccia, E.M. Innate Immune Response to SARS-CoV-2 Infection: From Cells to Soluble Mediators. Int. J. Mol. Sci. 2021, 22, 7017. [Google Scholar] [CrossRef]
- NIH. Clinical Spectrum of SARS-CoV-2 Infection. 2021. Available online: https://www.covid19treatmentguidelines.nih.gov/overview/clinical-spectrum/ (accessed on 15 December 2021).
- Tammam, S.N.; El Safy, S.; Ramadan, S.; Arjune, S.; Krakor, E.; Mathur, S. Repurpose but also (nano)-reformulate! The potential role of nanomedicine in the battle against SARS-CoV2. J. Control. Release 2021, 337, 258–284. [Google Scholar] [CrossRef] [PubMed]
- Shakaib, B.; Zohra, T.; Ikram, A.; Shakaib, M.B.; Ali, A.; Bashir, A.; Salman, M.; Khan, M.A.; Ansari, J. A comprehensive review on clinical and mechanistic pathophysiological aspects of COVID-19 Malady: How far have we come? Virol. J. 2021, 18, 120. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, L.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Neopane, P.; Nypaver, J.; Shrestha, R.; Beqaj, S.S. SARS-CoV-2 Variants Detection Using TaqMan SARS-CoV-2 Mutation Panel Molecular Genotyping Assays. Infect. Drug Resist. 2021, 14, 4471–4479. [Google Scholar] [CrossRef]
- Fisman, D.N.; Tuite, A.R. Evaluation of the relative virulence of novel SARS-CoV-2 variants: A retrospective cohort study in Ontario, Canada. CMAJ 2021, 193, E1619–E1625. [Google Scholar] [CrossRef]
- Buchta, C.; Camp, J.V.; Jovanovic, J.; Radler, U.; Benka, B.; Puchhammer-Stockl, E.; Müller, M.M.; Griesmacher, A.; Aberle, S.W.; Görzer, I. Inadequate design of mutation detection panels prevents interpretation of variants of concern: Results of an external quality assessment for SARS-CoV-2 variant detection. Clin. Chem. Lab. Med. 2021, 60, 291–298. [Google Scholar] [CrossRef]
- Aoki, A.; Adachi, H.; Mori, Y.; Ito, M.; Sato, K.; Okuda, K.; Sakakibara, T.; Okamoto, Y.; Jinno, H. A rapid screening assay for L452R and T478K spike mutations in SARS-CoV-2 Delta variant using high-resolution melting analysis. J. Toxicol. Sci. 2021, 46, 471–476. [Google Scholar] [CrossRef]
- Callaway, E. Heavily mutated Omicron variant puts scientists on alert. Nature 2021, 600, 21. [Google Scholar] [CrossRef]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Hozumi, Y.; Yin, C.; Wei, G.W. Mutations on COVID-19 diagnostic targets. Genomics. 2020, 112, 5204–5213. [Google Scholar] [CrossRef]
- Guruprasad, L. Human SARS CoV-2 spike protein mutations. Proteins 2021, 89, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Rodriguez, P.; Frances-Gomez, C.; Chiner-Oms, A.; Lopez, M.G.; Jimenez-Serrano, S.; Cancino-Munoz, I.; Ruiz-Hueso, P.; Torres-Puente, M.; Bracho, M.A.; D’Auria, G.; et al. Evolutionary and Phenotypic Characterization of Two Spike Mutations in European Lineage 20E of SARS-CoV-2. Mbio 2021, 12, e0231521. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, Y.; Liu, C.; Zhang, C.; Han, W.; Hong, X.; Wang, Y.; Hong, Q.; Wang, S.; Zhao, Q.; et al. Conformational dynamics of SARS-CoV-2 trimeric spike glycoprotein in complex with receptor ACE2 revealed by cryo-EM. Sci. Adv. 2021, 7, eabe5575. [Google Scholar] [CrossRef] [PubMed]
- Ozono, S.; Zhang, Y.; Ode, H.; Sano, K.; Tan, T.S.; Imai, K.; Miyoshi, K.; Kishigami, S.; Ueno, T.; Iwatani, Y.; et al. SARS-CoV-2 D614G spike mutation increases entry efficiency with enhanced ACE2-binding affinity. Nat. Commun. 2021, 12, 848. [Google Scholar] [CrossRef] [PubMed]
- Erol, A. Are the emerging SARS-COV-2 mutations friend or foe? Immunol. Lett. 2021, 230, 63–64. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, R.; Wang, M.; Wei, G.W. Mutations Strengthened SARS-CoV-2 Infectivity. J. Mol. Biol. 2020, 432, 5212–5226. [Google Scholar] [CrossRef]
- Suleman, M.; Yousafi, Q.; Ali, J.; Ali, S.S.; Hussain, Z.; Ali, S.; Waseem, M.; Iqbal, A.; Ahmad, S.; Khan, A.; et al. Bioinformatics analysis of the differences in the binding profile of the wild-type and mutants of the SARS-CoV-2 spike protein variants with the ACE2 receptor. Comput. Biol. Med. 2021, 138, 104936. [Google Scholar] [CrossRef]
- Aydemir, M.N.; Aydemir, H.B.; Korkmaz, E.M.; Budak, M.; Cekin, N.; Pinarbasi, E. Computationally predicted SARS-COV-2 encoded microRNAs target NFKB, JAK/STAT and TGFB signaling pathways. Gene Rep. 2021, 22, 101012. [Google Scholar] [CrossRef]
- Nain, Z.; Rana, H.K.; Lio, P.; Islam, S.M.S.; Summers, M.A.; Moni, M.A. Pathogenetic profiling of COVID-19 and SARS-like viruses. Brief. Bioinform. 2021, 22, 1175–1196. [Google Scholar] [CrossRef]
- Pairo-Castineira, E.; Clohisey, S.; Klaric, L.; Bretherick, A.D.; Rawlik, K.; Pasko, D.; Walker, S.; Parkinson, N.; Fourman, M.H.; Russell, C.D.; et al. Genetic mechanisms of critical illness in COVID-19. Nature 2021, 591, 92–98. [Google Scholar] [CrossRef]
- Mohlendick, B.; Schonfelder, K.; Breuckmann, K.; Elsner, C.; Babel, N.; Balfanz, P.; Dahl, E.; Dreher, M.; Fistera, D.; Herbstreit, F.; et al. ACE2 polymorphism and susceptibility for SARS-CoV-2 infection and severity of COVID-19. Pharm. Genom. 2021, 31, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Hamet, P.; Pausova, Z.; Attaoua, R.; Hishmih, C.; Haloui, M.; Shin, J.; Paus, T.; Abrahamowicz, M.; Gaudet, D.; Santucci, L.; et al. SARS-CoV-2 Receptor ACE2 Gene Is Associated with Hypertension and Severity of COVID 19: Interaction with Sex, Obesity, and Smoking. Am. J. Hypertens 2021, 34, 367–376. [Google Scholar] [CrossRef]
- Benetti, E.; Tita, R.; Spiga, O.; Ciolfi, A.; Birolo, G.; Bruselles, A.; Doddato, G.; Giliberti, A.; Marconi, C.; Musacchia, F.; et al. ACE2 gene variants may underlie interindividual variability and susceptibility to COVID-19 in the Italian population. Eur. J. Hum. Genet. 2020, 28, 1602–1614. [Google Scholar] [CrossRef]
- Hou, Y.; Zhao, J.; Martin, W.; Kallianpur, A.; Chung, M.K.; Jehi, L.; Sharifi, N.; Erzurum, S.; Eng, C.; Cheng, F.; et al. New insights into genetic susceptibility of COVID-19: An ACE2 and TMPRSS2 polymorphism analysis. BMC Med. 2020, 18, 216. [Google Scholar] [CrossRef] [PubMed]
- Torre-Fuentes, L.; Matias-Guiu, J.; Hernandez-Lorenzo, L.; Montero-Escribano, P.; Pytel, V.; Porta-Etessam, J.; Gómez-Pinedo, U.; Matías-Guiu, J.A. ACE2, TMPRSS2, and Furin variants and SARS-CoV-2 infection in Madrid, Spain. J. Med. Virol. 2021, 93, 863–869. [Google Scholar] [CrossRef]
- Brest, P.; Refae, S.; Mograbi, B.; Hofman, P.; Milano, G. Host Polymorphisms May Impact SARS-CoV-2 Infectivity. Trends Genet. 2020, 36, 813–815. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.; Blazyte, A.; Yoon, C.; Ryu, H.; Jeon, Y.; Bhak, Y.; Bolser, D.; Manica, A.; Shin, E.S.; Cho, Y.S.; et al. Regional TMPRSS2 V197M Allele Frequencies Are Correlated with COVID-19 Case Fatality Rates. Mol. Cells 2021, 44, 680–687. [Google Scholar] [CrossRef]
- Landrum, M.J.; Lee, J.M.; Benson, M.; Brown, G.; Chao, C.; Chitipiralla, S.; Gu, B.; Hart, J.; Hoffman, D.; Hoover, J.; et al. ClinVar: Public archive of interpretations of clinically relevant variants. Nucleic Acids Res. 2016, 44, D862–D868. [Google Scholar] [CrossRef] [Green Version]
- Arisan, E.D.; Dart, A.; Grant, G.H.; Arisan, S.; Cuhadaroglu, S.; Lange, S.; Uysal-Onganer, P. The Prediction of miRNAs in SARS-CoV-2 Genomes: Hsa-miR Databases Identify 7 Key miRs Linked to Host Responses and Virus Pathogenicity-Related KEGG Pathways Significant for Comorbidities. Viruses 2020, 12, 614. [Google Scholar] [CrossRef]
- van der Made, C.I.; Simons, A.; Schuurs-Hoeijmakers, J.; van den Heuvel, G.; Mantere, T.; Kersten, S.; van Deuren, R.C.; Steehouwer, M.; van Reijmersdal, S.V.; Jaeger, M.; et al. Presence of Genetic Variants Among Young Men With Severe COVID-19. JAMA 2020, 324, 663–673. [Google Scholar] [CrossRef]
- Pati, A.; Padhi, S.; Suvankar, S.; Panda, A.K. Minor Allele of Interferon-Induced Transmembrane Protein 3 Polymorphism (rs12252) Is Covered Against Severe Acute Respiratory Syndrome Coronavirus 2 Infection and Mortality: A Worldwide Epidemiological Investigation. J. Infect. Dis. 2021, 223, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Cuesta-Llavona, E.; Albaiceta, G.M.; Garcia-Clemente, M.; Duarte-Herrera, I.D.; Amado-Rodriguez, L.; Hermida-Valverde, T.; Enríquez-Rodriguez, A.I.; Hernández-González, C.; Melón, S.; Alvarez-Argüelles, M.E.; et al. Association between the interferon-induced transmembrane protein 3 gene (IFITM3) rs34481144 / rs12252 haplotypes and COVID-19. Curr. Res. Virol. Sci. 2021, 2, 100016. [Google Scholar] [CrossRef] [PubMed]
- Hubacek, J.A.; Dusek, L.; Majek, O.; Adamek, V.; Cervinkova, T.; Dlouha, D.; Adamkova, V. ACE I/D polymorphism in Czech first-wave SARS-CoV-2-positive survivors. Clin. Chim. Acta 2021, 519, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, M.; Lahousse, L.; Van Eeckhoutte, H.P.; Wijnant, S.R.A.; Delanghe, J.R.; Brusselle, G.G.; Bracke, K.R. Effect of ACE1 polymorphism rs1799752 on protein levels of ACE2, the SARS-CoV-2 entry receptor, in alveolar lung epithelium. ERJ Open Res. 2021, 7, 206–209. [Google Scholar] [CrossRef]
- Kouhpayeh, H.R.; Tabasi, F.; Dehvari, M.; Naderi, M.; Bahari, G.; Khalili, T.; Clark, C.; Ghavami, S.; Taheri, M. Association between angiotensinogen (AGT), angiotensin-converting enzyme (ACE) and angiotensin-II receptor 1 (AGTR1) polymorphisms and COVID-19 infection in the southeast of Iran: A preliminary case-control study. Transl. Med. Commun. 2021, 6, 26. [Google Scholar] [CrossRef]
- Wang, F.; Huang, S.; Gao, R.; Zhou, Y.; Lai, C.; Li, Z.; Xian, W.; Qian, X.; Zhiyu Li, Z.; Huang, Y.; et al. Initial whole-genome sequencing and analysis of the host genetic contribution to COVID-19 severity and susceptibility. Cell Discov. 2020, 6, 83. [Google Scholar] [CrossRef]
- Ferreira de Araujo, J.L.; Menezes, D.; Saraiva-Duarte, J.M.; de Lima Ferreira, L.; Santana de Aguiar, R.; Pedra de Souza, R. Systematic review of host genetic association with Covid-19 prognosis and susceptibility: What have we learned in 2020? Rev. Med Virol. 2021, 2021, e2283. [Google Scholar]
- Tomita, Y.; Ikeda, T.; Sato, R.; Sakagami, T. Association between HLA gene polymorphisms and mortality of COVID-19: An in silico analysis. Immun. Inflamm. Dis. 2020, 8, 684–694. [Google Scholar] [CrossRef]
- Littera, R.; Campagna, M.; Deidda, S.; Angioni, G.; Cipri, S.; Melis, M.; Firinu, D.; Santus, S.; Lai, A.; Porcella, R.; et al. Human Leukocyte Antigen Complex and Other Immunogenetic and Clinical Factors Influence Susceptibility or Protection to SARS-CoV-2 Infection and Severity of the Disease Course. The Sardinian Experience. Front. Immunol. 2020, 11, 605688. [Google Scholar] [CrossRef]
- Novelli, A.; Andreani, M.; Biancolella, M.; Liberatoscioli, L.; Passarelli, C.; Colona, V.L.; Rogliani, P.; Leonardis, F.; Campana, A.; Carsetti, R.; et al. HLA allele frequencies and susceptibility to COVID-19 in a group of 99 Italian patients. HLA 2020, 96, 610–614. [Google Scholar] [CrossRef]
- Zhang, Q.; Bastard, P.; Liu, Z.; Le Pen, J.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Sabli, I.K.D.; Hodeib, S.; Korol, C.; et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science 2020, 370, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kuznik, A.; Bencina, M.; Svajger, U.; Jeras, M.; Rozman, B.; Jerala, R. Mechanism of endosomal TLR inhibition by antimalarial drugs and imidazoquinolines. J. Immunol. 2011, 186, 4794–4804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Li, C.; Wang, S.; Li, T.; Zhang, H. Genetic variants are identified to increase risk of COVID-19 related mortality from UK Biobank data. Hum. Genom. 2021, 15, 10. [Google Scholar] [CrossRef] [PubMed]
- Taha, S.I.; Shata, A.K.; Baioumy, S.A.; Fouad, S.H.; Anis, S.G.; Mossad, I.M.; Moustafa, N.M.; Abdou, D.M.; Youssef, M.K. Toll-Like Receptor 4 Polymorphisms (896A/G and 1196C/T) as an Indicator of COVID-19 Severity in a Convenience Sample of Egyptian Patients. J. Inflamm. Res. 2021, 14, 6293–6303. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, Y.; Huang, H.; Li, D.; Gu, D.; Lu, X.; Zhang, Z.; Liu, L.; Liu, T.; Liu, Y.K.; et al. Relationship Between the ABO Blood Group and the Coronavirus Disease 2019 (COVID-19) Susceptibility. Clin. Infect. Dis. 2021, 73, 328–331. [Google Scholar] [CrossRef]
- Fan, Q.; Zhang, W.; Li, B.; Li, D.J.; Zhang, J.; Zhao, F. Association Between ABO Blood Group System and COVID-19 Susceptibility in Wuhan. Front. Cell Infect. Microbiol. 2020, 10, 404. [Google Scholar] [CrossRef]
- Zhang, Y.; Garner, R.; Salehi, S.; La Rocca, M.; Duncan, D. Association between ABO blood types and coronavirus disease 2019 (COVID-19), genetic associations, and underlying molecular mechanisms: A literature review of 23 studies. Ann. Hematol. 2021, 100, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.L.; Pilling, L.C.; Atkins, J.L.; Masoli, J.A.H.; Delgado, J.; Kuchel, G.A.; Melzer, D. ApoE e4e4 Genotype and Mortality With COVID-19 in UK Biobank. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 1801–1803. [Google Scholar] [CrossRef]
- Delanghe, J.R.; De Buyzere, M.L.; Speeckaert, M.M. C3 and ACE1 polymorphisms are more important confounders in the spread and outcome of COVID-19 in comparison with ABO polymorphism. Eur. J. Prev. Cardiol. 2020, 27, 1331–1332. [Google Scholar] [CrossRef] [PubMed]
- Delanghe, J.R.; De Buyzere, M.L.; Speeckaert, M.M. Genetic Polymorphisms in the Host and COVID-19 Infection. Adv. Exp. Med. Biol. 2021, 1318, 109–118. [Google Scholar]
- Mehlotra, R.K. Chemokine receptor gene polymorphisms and COVID-19: Could knowledge gained from HIV/AIDS be important? Infect. Genet. Evol. 2020, 85, 104512. [Google Scholar] [CrossRef]
- Ahmed, A.A.; Ad’hiah, A.H. Interleukin-37 gene polymorphism and susceptibility to coronavirus disease 19 among Iraqi patients. Meta. Gene. 2022, 31, 100989. [Google Scholar] [CrossRef] [PubMed]
- Saponi-Cortes, J.M.R.; Rivas, M.D.; Calle-Alonso, F.; Sanchez, J.F.; Costo, A.; Martin, C.; Zamorano, J. IFNL4 genetic variant can predispose to COVID-19. Sci. Rep. 2021, 11, 21185. [Google Scholar] [CrossRef]
- Al-Anouti, F.; Mousa, M.; Karras, S.N.; Grant, W.B.; Alhalwachi, Z.; Abdel-Wareth, L.; Uddin, M.; Alkaabi, N.; Tay, G.K.; Mahboub, B.; et al. Associations between Genetic Variants in the Vitamin D Metabolism Pathway and Severity of COVID-19 among UAE Residents. Nutrients 2021, 13, 3680. [Google Scholar] [CrossRef] [PubMed]
- Pati, A.; Mahto, H.; Padhi, S.; Panda, A.K. ACE deletion allele is associated with susceptibility to SARS-CoV-2 infection and mortality rate: An epidemiological study in the Asian population. Clin. Chim. Acta. 2020, 510, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Avbelj, M.; Horvat, S.; Jerala, R. The role of intermediary domain of MyD88 in cell activation and therapeutic inhibition of TLRs. J. Immunol. 2011, 187, 2394–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidari Nia, M.; Rokni, M.; Mirinejad, S.; Kargar, M.; Rahdar, S.; Sargazi, S.; Sarhadi, M.; Saravani, R. Association of polymorphisms in tumor necrosis factors with SARS-CoV-2 infection and mortality rate: A case-control study and in silico analyses. J. Med. Virol. 2021, 1–11. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, A.C.M.; dos Santos, B.R.C.; Dos Santos, B.B.; de Moura, E.L.; Ferreira, J.M.; Dos Santos, L.K.C.; Oliveira, S.P.; Dias, R.B.F.; Pereira e Silva, A.C.; de Farias, K.F.; et al. Genetic polymorphisms as multi-biomarkers in severe acute respiratory syndrome (SARS) by coronavirus infection: A systematic review of candidate gene association studies. Infect Genet Evol. 2021, 93, 104846. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Zhu, Y.; Lan, J.; Chen, H.; Wang, Y.; Shi, H.; Feng, F.; Chen, D.Y.; Close, B.; Zhao, X.; et al. Susceptibilities of human ACE2 genetic variants in coronavirus infection. J. Virol. 2022, 96, e01492-21. [Google Scholar] [CrossRef]
- Turunen, T.A.; Roberts, T.C.; Laitinen, P.; Vaananen, M.A.; Korhonen, P.; Malm, T.; Ylä-Herttuala, S.; Turunen, M.P. Changes in nuclear and cytoplasmic microRNA distribution in response to hypoxic stress. Sci. Rep. 2019, 9, 10332. [Google Scholar] [CrossRef] [Green Version]
- Banaganapalli, B.; Al-Rayes, N.; Awan, Z.A.; Alsulaimany, F.A.; Alamri, A.S.; Elango, R.; Malik, Z.; Shaik, N.A. Multilevel systems biology analysis of lung transcriptomics data identifies key miRNAs and potential miRNA target genes for SARS-CoV-2 infection. Comput. Biol. Med. 2021, 135, 104570. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Sany, M.R.U.; Islam, M.S.; Islam, A. Epigenetic Regulator miRNA Pattern Differences Among SARS-CoV, SARS-CoV-2, and SARS-CoV-2 World-Wide Isolates Delineated the Mystery Behind the Epic Pathogenicity and Distinct Clinical Characteristics of Pandemic COVID-19. Front. Genet. 2020, 11, 765. [Google Scholar] [CrossRef] [PubMed]
- Sarma, A.; Phukan, H.; Halder, N.; Madanan, M.G. An in-silico approach to study the possible interactions of miRNA between human and SARS-CoV2. Comput. Biol. Chem. 2020, 88, 107352. [Google Scholar] [CrossRef] [PubMed]
- Visacri, M.B.; Nicoletti, A.S.; Pincinato, E.C.; Loren, P.; Saavedra, N.; Saavedra, K.; Salazar, L.A.; Moriel, P. Role of miRNAs as biomarkers of COVID-19: A scoping review of the status and future directions for research in this field. Biomark. Med. 2021, 15, 1785–1795. [Google Scholar] [CrossRef] [PubMed]
- Saksena, N.; Bonam, S.R.; Miranda-Saksena, M. Epigenetic Lens to Visualize the Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2) Infection in COVID-19 Pandemic. Front. Genet. 2021, 12, 581726. [Google Scholar] [CrossRef]
- Castro de Moura, M.; Davalos, V.; Planas-Serra, L.; Alvarez-Errico, D.; Arribas, C.; Ruiz, M.; Aguilera-Albesa, S.; Troya, J.; Valencia-Ramos, J.; Vélez-Santamaria, V.; et al. Epigenome-wide association study of COVID-19 severity with respiratory failure. EBioMedicine 2021, 66, 103339. [Google Scholar] [CrossRef]
- Song, J.W.; Lam, S.M.; Fan, X.; Cao, W.J.; Wang, S.Y.; Tian, H.; Chua, G.H.; Zhang, C.; Meng, F.P.; Xu, Z.; et al. Omics-Driven Systems Interrogation of Metabolic Dysregulation in COVID-19 Pathogenesis. Cell Metab. 2020, 32, 188–202.e5. [Google Scholar] [CrossRef]
- Fujita, Y.; Hoshina, T.; Matsuzaki, J.; Yoshioka, Y.; Kadota, T.; Hosaka, Y.; Fujimoto, S.; Kawamoto, H.; Watanabe, N.; Sawaki, K.; et al. Early prediction of COVID-19 severity using extracellular vesicle COPB2. J. Extracell Vesicles 2021, 10, e12092. [Google Scholar] [CrossRef]
- Mitchell, M.I.; Ben-Dov, I.Z.; Liu, C.; Ye, K.; Chow, K.; Kramer, Y.; Gangadharan, A.; Park, S.; Fitzgerald, S.; Ramnauth, A.; et al. Extracellular Vesicle Capture by AnTibody of CHoice and Enzymatic Release (EV-CATCHER): A customizable purification assay designed for small-RNA biomarker identification and evaluation of circulating small-EVs. J. Extracell Vesicles 2021, 10, e12110. [Google Scholar] [CrossRef]
- Barberis, E.; Vanella, V.V.; Falasca, M.; Caneapero, V.; Cappellano, G.; Raineri, D.; Ghirimoldi, M.; De Giorgis, V.; Puricelli, C.; Vaschetto, R.; et al. Circulating Exosomes Are Strongly Involved in SARS-CoV-2 Infection. Front. Mol. Biosci. 2021, 8, 632290. [Google Scholar] [CrossRef]
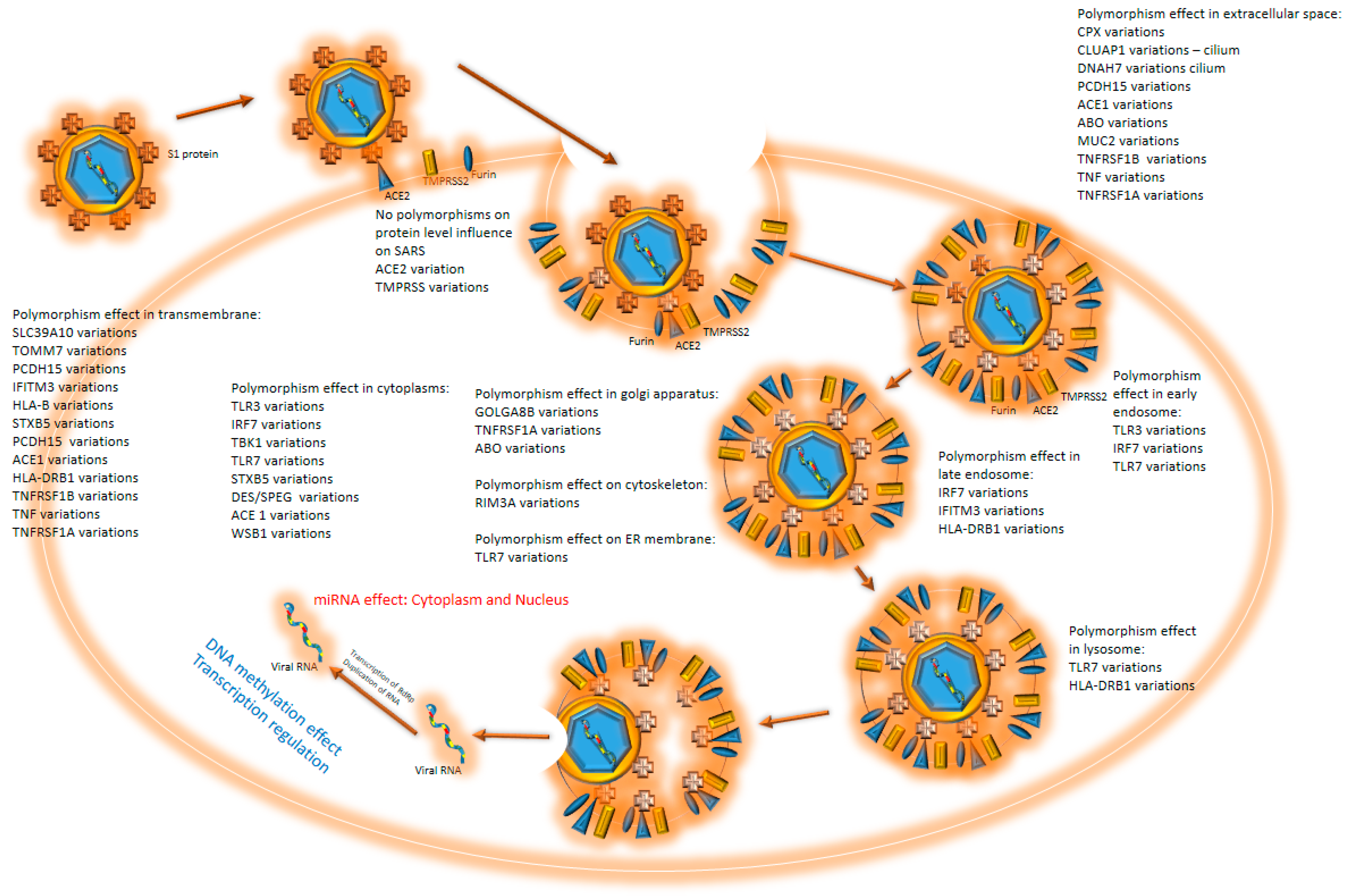
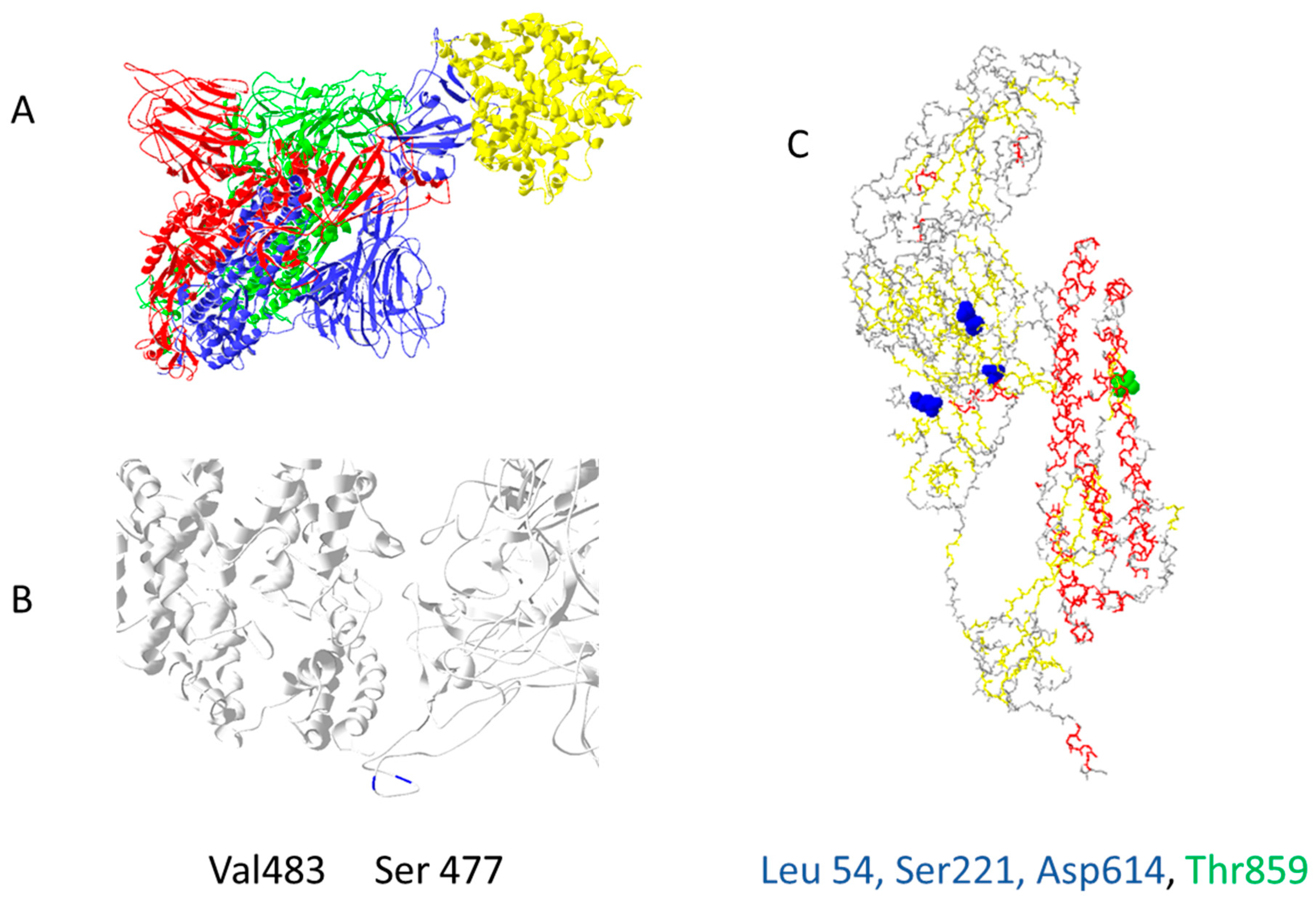
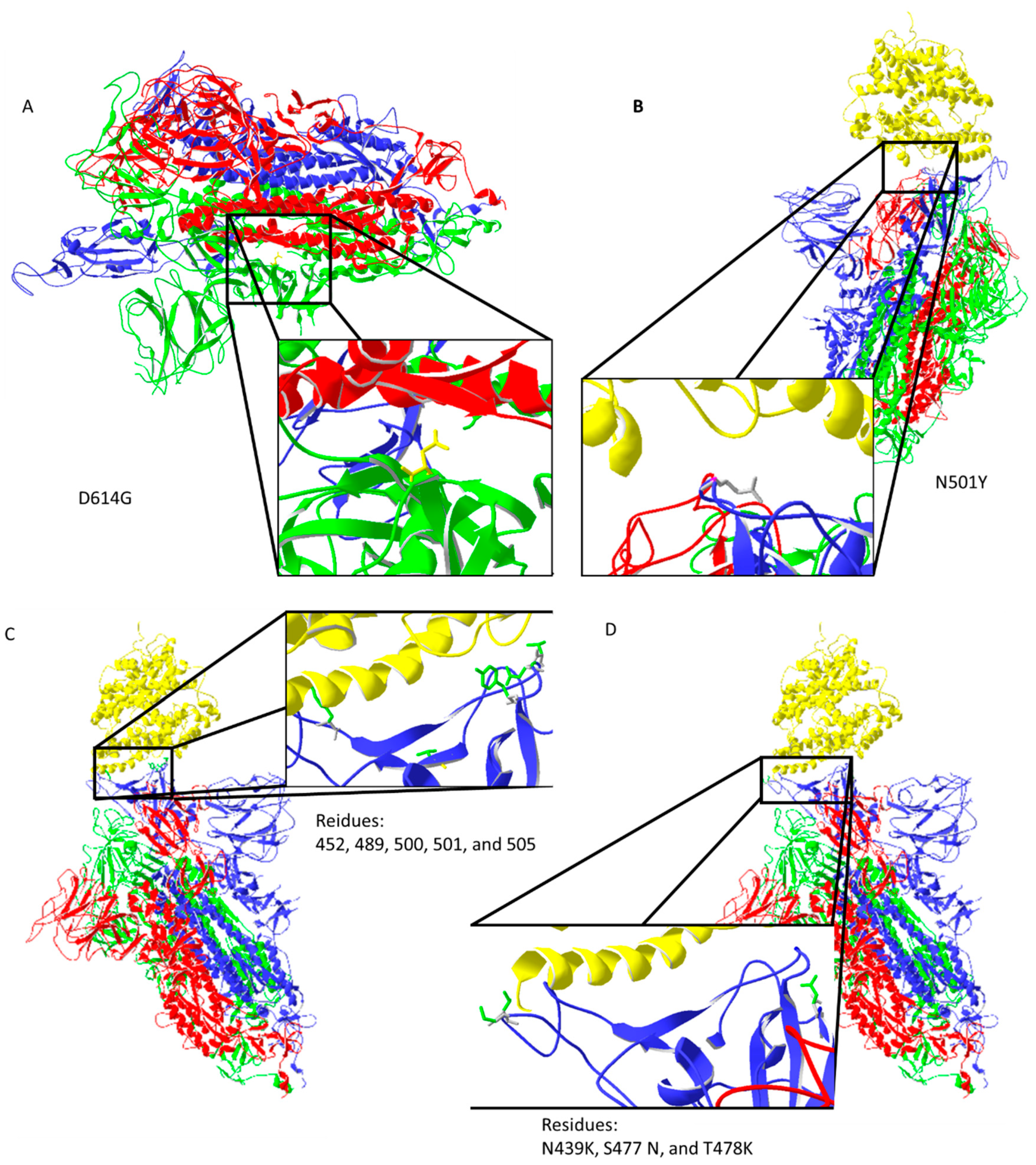
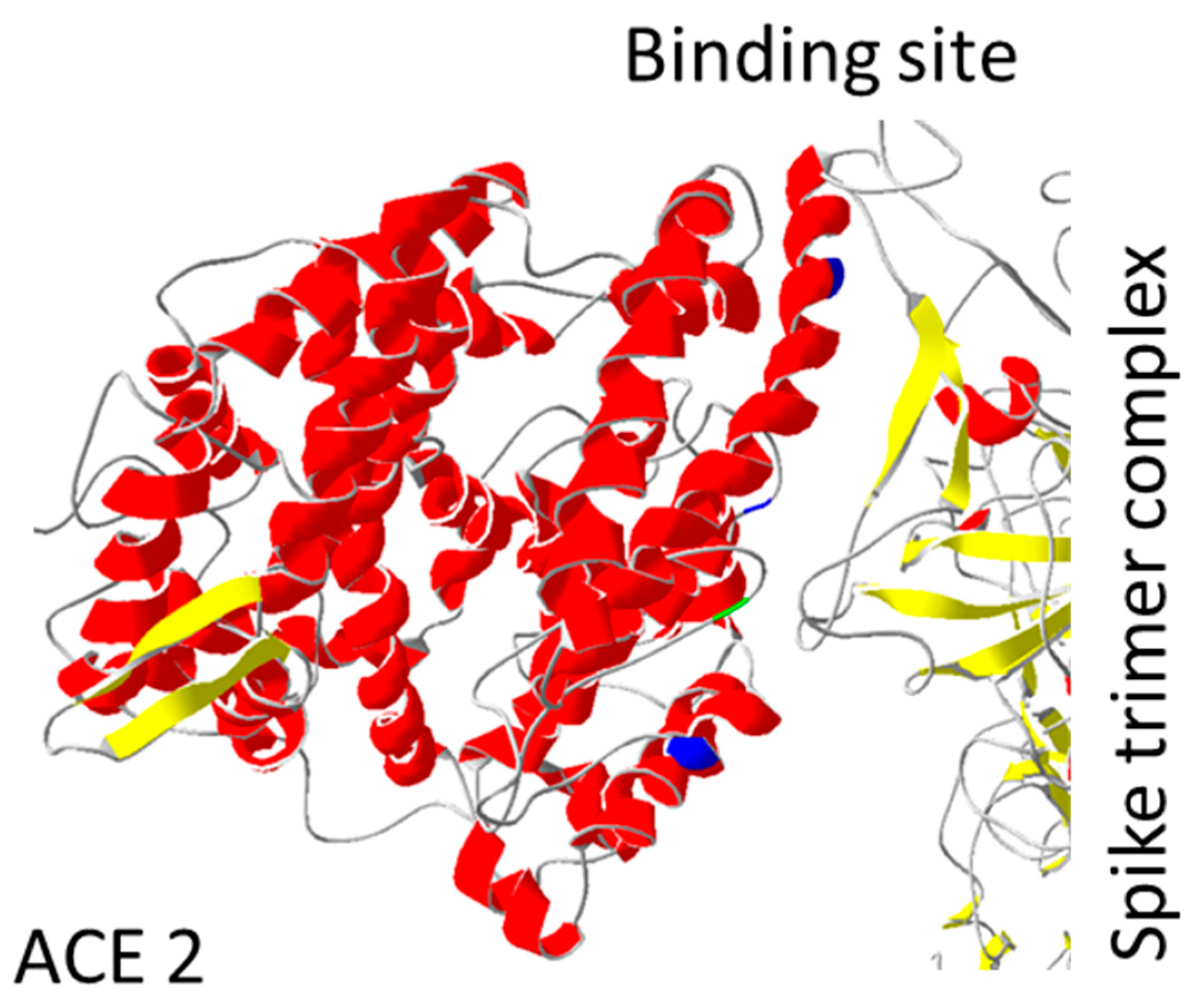

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | rs Code | Protein Mutation | DNA Mutation | Function SARS-CoV-2 | Polyphen-2 | Protein Function | Reference |
|---|---|---|---|---|---|---|---|
| ACE2 | rs2285666 | Intron variant | g.14934G > T | Increased severity of SARS-CoV-2 (GG genotypes or G-allele carriers), 3 times higher risk, fatality risk | Angiotensin-converting enzyme and functional receptor of COVID-19. | [32,33,34,35] | |
| rs2074192 | Intron variant | g.42492G > A | Increased severity of SARS-CoV-2 | ||||
| V749V | c.2247G > A | Possible protective role against SARS-CoV-2 (found in healthy controls) | |||||
| TMPRSS2 | rs12329760 | V197M | c.589G > A | Association with COVID-19 case fatality rate | 0.999- probably damaging | Serine protease also facilitates entry of the COVID-19 virus by proteolytic cleaving and activation. | [36,37,38] |
| rs61735792 | P100P | c.300C > T | Significant association with SARS-CoV-2 infection | ||||
| rs61735794 | G422G | c.1266G > A | Significant association with SARS-CoV-2 infection | ||||
| TNFRSF1B | M196R | c.587T > G | Susceptibility to severe coronavirus disease (COVID-19) | 0-benign | A member of the TNF receptor superfamily; its ligand, cytokine TNF, is involved in regulating a broad spectrum of processes. Mutations in this gene affect susceptibility to several diseases. | [39] | |
| 3’ prime UTR | c.*215C > T | Susceptibility to severe coronavirus disease (COVID-19) | |||||
| TNF | rs1800629 | Upstream variant | c.-488G > A TNFα-311A > G | Susceptibility to severe coronavirus disease (COVID-19) | Cytokine is involved in the regulation of a broad spectrum of processes. Mutations in this gene affect susceptibility to several diseases. | [39] | |
| g.31575324G > A | Susceptibility to severe coronavirus disease (COVID-19) | ||||||
| rs909253 | Upstream variant | TNFβ-252A > G | Is more frequent in COVID-19 patients, conferrers protection against COVID-19 susceptibility | ||||
| TNFRSF1A | Intron variant | c.625 + 10A > G | Susceptibility to severe coronavirus disease (COVID-19) | A member of the TNF receptor superfamily of proteins. Mutations in this gene may be associated with a few diseases. | [39] | ||
| P12P | c.36A > G | Susceptibility to severe coronavirus disease (COVID-19), | |||||
| TBK1 | splice donor variant | c.1760 + 4 1760 + 7del | Severe SARS-CoV-2 infection, susceptibility to | Codes protein, an important kinase for antiviral innate immune response, is similar to IKB. | [39] | ||
| TNFRSF13B | C104R | c.310T > C | Severe SARS-CoV-2 infection, susceptibility to, | 1.000-probably damaging | A member of the TNF receptor superfamily, it plays an important role in humoral immunity. | [39,40] | |
| TLR7 | Q710fs | c.21292132del | Preliminary severe COVID-19-related | Receptor senses ss RNA oligonucleotides with guanosine- and uridine-rich sequences. | [41] | ||
| V795F | c.2383G > T | Immunodeficiency 74, COVID-19-related, X-linked | 0.997-probably damaging | ||||
| IFITM3 | rs12252 | S14S | c.42T > C | Increased severity of SARS-CoV-2 disease in the studied Chinese cohort, the C allele of IFITM3 rs12252 polymorphism was negatively correlated with the SARS-CoV-2 mortality rate per million | Interferon-induced membrane protein helps build up immunity against several viruses. | [42,43] | |
| rs34481144 | 5 Prime UTR Variant | c.-23C > T | Were significantly more frequent in COVID-19 hospitalized patients | ||||
| ACE1 | rs4646994 | Intron variant | c.2306-109_2306-108ins | Might Increase susceptibility to SARS-CoV-2 disease ACE I/D polymorphism (Czech first-wave SARS-CoV-2-positive survivors); decreased risk of infection | An enzyme that catalyzes the conversion of angiotensin I into active angiotensin II regulates homologous ACE2, which may influence the progression of SARS-CoV-2 disease. | [44,45,46] | |
| rs1799752 | Intron variant | g.16471_16472ins | Might facilitate human cell entry of SARS-CoV-2 due to increased level of ACE2 | ||||
| TMEM189–UBE2V1 | rs6020298 | Intron variant | g.48769055G > A | Increased severity of SARS-CoV-2 disease | The function of this read-through mRNA and its protein is unknown. | [47] | |
| HLA | Several variants | Several variants | HLA-B*46:01 increased severity of SARS-CoV-2; HLA DRB1*15:01 and DQB1*06:02 increased severity of SARS-CoV-2 disease; HLA-A 02-01 increased risk for COVID-19; HLA-C*04:01 associated with infection; HLA-A30:02, HLA-B 14:02; HLA-C 08:02 significant association with severe and extremely severe COVID-19 | HLA molecules play a central role in the immune system by presenting antigenic peptides on the cell surface—an important role in the occurrence and outcome of SARS-CoV-2. | [48,49,50,51] | ||
| TLR3 | p.Ser339fs | Increased life-threatening complications in patients with SARS-CoV-2 TLR3 | Recognize pathogen-associated molecular patterns (PAMPs), specifically dsRNA, to combat multiple virus infections. | [48,52,53] | |||
| rs121434431 | p.Pro554Ser | c.1660C > T | Increased life-threatening complications in patients with SARS-CoV-2 TLR3 | 1.000-probably damaging | |||
| p.Trp769* | Increased life-threatening complications in patients with SARS-CoV-2 TLR3 deficiency | ||||||
| p.Met870Val | Increased life-threatening complications in patients with SARS-CoV-2 TLR3 deficiency | 0.995-probably damaging | |||||
| IRF7 | pArg7fs; | g.615095A > C | Increased life-threatening complications in patients with SARS-CoV-2 IRF7 deficiency | Plays a role in the transcriptional activation and innate immune response against DNA and RNA viruses. | [48,52] | ||
| p.Pro364fs | Increased life-threatening complications in patients with SARS-CoV-2 IRF7 deficiency | ||||||
| p.Gln185* | Increased life-threatening complications in patients with SARS-CoV-2 IRF7 deficiency | ||||||
| p.Pro246fs | Increased life-threatening complications in patients with SARS-CoV-2 IRF7 deficiency | ||||||
| STXBP5/STXBP5-AS1 | rs116898161 | Intron variant | g.147538692A > G | Increased mortality in SARS-CoV-2 disease | Protein may be involved in neurotransmitter release by stimulating SNARE complex formation. | [48,54] | |
| rs117928001 | Intron variant | g.147514999C > A | Increased mortality in SARS-CoV-2 disease | ||||
| CPQ | rs1431889 | Intron variant | g.98141643G > C | Increased mortality in SARS-CoV-2 disease | The enzyme catalyzes the stepwise oxidative decarboxylation of coproporphyrinogen III. | [48,54] | |
| rs7817272 | Intron variant | g.98140470T > A | Increased mortality in SARS-CoV-2 disease | ||||
| CLUAP1 | rs2301762 | Upstream variant | g.3550977C > G | Increased mortality in SARS-CoV-2 disease | This gene contains a single coiled-coil region. | [48,54] | |
| WSB1 | rs60811869 | g.25590833T > C | Increased mortality in SARS-CoV-2 disease | This protein has several WD-repeats and a SOCS box in the C-terminus. | [48,54] | ||
| DNAH7/SLC39A10 | rs183712207 | Intron Variant | g.196611282G > A | Increased mortality in SARS-CoV-2 disease | DNAH7 is a part of the inner dynein arm of the ciliary axonemes. SLC39A10 shows the structural characteristics of zinc transporters. | [48,54] | |
| DES/SPEG | rs71040457 | Upstream, downstream variant | g.220294782_220294783insG | Increased mortality in SARS-CoV-2 disease | LCR may control the preferentially muscle-expressed SPEG gene expression downstream of the DES gene. | [48,54] | |
| TOMM7 | rs55986907 | Intron variant | g.22817292C > A | Increased mortality in SARS-CoV-2 disease | This gene encodes a subunit of the translocase of the outer mitochondrial membrane. | [48,54] | |
| PCDH15 | rs9804218 | Intron Variant | g.56495374G > A | Increased mortality in SARS-CoV-2 disease | Plays an essential role in maintenance of normal retinal and cochlear function. | [48,54] | |
| OAS3 | rs10735079 | Intron variant | c.460 + 351G > A | Involved in the critical illness of SARS-CoV-2 disease | This enzyme helps inhibit cellular protein synthesis and helps in viral infection resistance. | [31] | |
| ABO (A, B, and O) | Set of blood type genotypes: rs1556058284; B type: rs8176743; rs8176746; rs8176747 | ABO c.260insG (p.Val87_Thr88fs*); set of variants that contribute to blood type: p.Gly235Arg, p.Leu266Met, p.Gly268Val | Blood type influencing variants: del261- O type- frame shift variant; two amino acid substitutions- B type: c.703G > C, c.796C > A, c.803G > T | Increased susceptibility for SARS-CoV-2 development A type (Fan 2020, Zhao 2020), Decreased susceptibility for SARS-CoV-2 development O-type (Zhao 2020); A > O > B > AB susceptibility (Zhang 2021) | Variations in the ABO gene are the basis of the ABO blood group. Gene has a role in susceptibility and severity of coronavirus disease 2019. | [48,56,57,58] | |
| APOE | rs429358 allele is (T) + rs7412(C) allele | APOE-ε4 (arg112, arg158) | c.466T > C + c.604C > T | Increased susceptibility for SARS-CoV-2 development, a 4-fold increase in mortality | It is essential for the normal catabolism of triglyceride-rich lipoproteins. | [59] | |
| RIMBP3 | E1263A | c.3788A > C | Increased susceptibility for SARS-CoV-2 development | 0.884-possibly damaging | It may be a component of the manchette. | [47,48] | |
| GOLGA8B | rs200975425 | R513P | c.1538G > C | Increased susceptibility for SARS-CoV-2 development | 0.011-benign | May be involved in maintaining Golgi structure. | [47,48] |
| C3 | rs2230199 | R102G | c.304C > G | Determinants for COVID-19 prevalence/mortality | 0.000-benign | Classical and alternative complement activation pathways are regulated through the activation of C3. | [60,61] |
| CCR5 | rs333 | S185fs | c.554_585del | Association with susceptibility to SARS-CoV-2 infection and mortality | A beta chemokine receptor family member can cross the membrane seven times. Frame shifts and gene deletions have been associated with HIV infection resistance. | [62] | |
| IFNAR2 | rs2236757 | Intron variant | c.541-50A > G | Significant associations with critical illness | Activated receptor stimulates Janus protein kinases. | [31] | |
| DPP9 | rs2109069 | Intron variant | c.56 + 420C > T | significant associations with critical illness | Gene encodes a protein that is a member of the serine proteases. | [31] | |
| IFNL4 | rs12979860 | Intron variant | g.39738787C > G(T) | It could be a risk factor for the development of COVID-19 | Interferons are released in response to viral infection. They block replication and propagation to uninfected cells. IFNL4 encodes the interferon (IFN) lambda 4 protein. | [64] | |
| GC | rs59241277, rs113574864, rs182901986, rs60349934, rs113876500 | Intron variants | g.34732T > C, g.48235G > T, g.29797C > T, g.49750A > G, g.9899C > T | Polymorphisms are associated with the critical COVID-19 condition | Gene codes transporter for vitamin D that belongs to albumin family. | [65] | |
| NADSYN1 | rs4944076, rs4944997, rs4944998, rs4944979, rs10898210 | Intron variants | g.71211654G > A, g.71206368A > C, g.71207205C > G, g.71197540T > A, g.71210023G > A | Polymorphisms are associated with the critical COVID-19 condition | NAD synthase catalyzes the final step in the biosynthesis of NAD from nicotinic acid adenine dinucleotide. | [65] | |
| VDR | rs11574018, rs11574024 | Intron variants | g.48297294T > C, g.48296221G > A | Polymorphisms are associated with the critical COVID-19 condition | This gene encodes the vitamin D3 receptor. | [65] | |
| AGT | rs699 | M259T | c.776T > C | Are associated with the risk of COVID-19 infection, increased risk of infection | 1.000-probably damaging | Gene codes angiotensinogen precursor cleaved by the enzyme renin in response to lowered blood pressure. | [46] |
| TLR4 | rs4986790 | Asp299Gly | c.896A/G | Was associated with severe COVID-19 | 0.104-benign | Gene codes Toll-like receptor, which plays a fundamental role in pathogen recognition and activation of innate immunity. | [55] |
| IL37 | rs3811046 | p.Gly31Glu; p.Gly31Ala; p.Gly31Val | c.92G > A; c.92G > C; c.92G > T | Maybe associated with susceptibility to COVID-19 among the Iraqi population | 0.484-possibly damaging | The protein is a member of the interleukin 1 cytokine family, and it may be a ligand for the interleukin 18 receptor. | [63] |
| rs3811047 | p.Thr42Ala; p.Thr42Ser | c.124A > G; c.124A > T | 0.000 benign |
| Protein ACE2 | ||
|---|---|---|
| Position | Protein Mutation | Function SARS-CoV-2 |
| 19 | S into P | increases slightly interaction with RBD |
| 24–26 | QAK into KAE | slightly inhibits interaction |
| 24 | Q into T | slightly increases interaction with RBD |
| 25 | A into V | increases slightly interaction with RBD |
| 27 | T into Y | increases slightly interaction with RBD |
| 29 | L into F | increases slightly the interaction with RBD |
| 31 | K into Y | increases slightly the interaction with RBD |
| 34 | H into A | increases slightly the interaction with RBD |
| 39 | L into R | increases slightly the interaction with RBD |
| 40 | F into D | increases slightly the interaction with RBD |
| 41 | Y into R | increases slightly the interaction with RBD |
| 42 | Q into L | increases slightly the interaction with RBD |
| 69 | W into V | increases slightly the interaction with RBD |
| 72 | F into Y | increases slightly the interaction with RBD |
| 75 | E into K | increases slightly the interaction with RBD |
| 76 | Q into T | increases slightly the interaction with RBD |
| 79 | L into T | increases slightly the interaction with RBD |
| 89 | Q into P | increases slightly the interaction with RBD |
| 90 | N into Q | increases slightly the interaction with RBD |
| 91 | L into P | increases slightly the interaction with RBD |
| 92 | T into Q | increases slightly the interaction with RBD |
| 324 | Q into P | increases slightly the interaction with RBD |
| 330 | N into Y | increases slightly the interaction with RBD, increases interaction with RBD if associated with Y27 and L386 |
| 351 | L into F | increases slightly the interaction with RBD |
| 386 | A into L | increases slightly the interaction with RBD, increases interaction with RBD if associated with Y27 and L330 |
| 389 | P into D | increases slightly the interaction with RBD |
| 393 | R into K | increases very slightly the interaction with RBD |
| 518 | R into G | increases very slightly the interaction with RBD |
| 355 | D into A | Restricts interaction with spike [70] |
| miRNA | Function | Publication |
|---|---|---|
| hsa-miR-342-5p | antiviral SARS-CoV2 gene (ORF1ab) | [71] |
| hsa-miR-432-5p | antiviral SARS-CoV2 gene (ORF1ab) | [71] |
| hsa-miR-98-5p | antiviral SARS-CoV2 gene (ORF1ab) | [71] |
| hsa-miR-17-5p | antiviral SARS-CoV2 gene (ORF1ab) | [71] |
| hsa-miR-17-5p | exhibit experimental evidence of having antiviral roles during infections against SARS1 and SARS2 | [72] |
| hsa-miR-20b-5p | exhibit experimental evidence of having antiviral roles during infections against SARS1 and SARS3 | [72] |
| hsa-miR-323a-5p | exhibit experimental evidence of having antiviral roles during infections against SARS1 and SARS4 | [72] |
| 7c-5p | target COVID-19 genome | [29] |
| miR-27b-3p | target COVID-19 genome | [29] |
| miR-98-5p | target COVID-19 genome | [29] |
| miR-125a-5p | target COVID-19 genome | [29] |
| let-7b-5p | associated with the development of COVID-19 symptoms | [29] |
| miR-155-5p | associated with the development of COVID-19 symptoms | [29] |
| miR-186-5p | associated with the development of COVID-19 symptoms | [29] |
| miR-16-5p | associated with the development of COVID-19 symptoms | [29] |
| miR-27b-3p | associated with the development of COVID-19 symptoms | [29] |
| miR-29a-3p | associated with the development of COVID-19 symptoms | [29] |
| miR-30a-5p | associated with the development of COVID-19 symptoms | [29] |
| miRs 8066, | associated with host response and virus pathogenicity | [73] |
| 5197 | associated with host response and virus pathogenicity | [73] |
| 3611 | associated with host response and virus pathogenicity | [73] |
| 3934-3p | associated with host response and virus pathogenicity | [73] |
| 1307-3p | associated with host response and virus pathogenicity | [73] |
| 3691-3p | associated with host response and virus pathogenicity | [73] |
| 1468-5p | associated with host response and virus pathogenicity | [73] |
| hsa-mir-1267 | were found in all five viral SARS-CoV2 cases | [40] |
| hsa-mir-1-3p | were found in all five viral SARS-CoV2 cases | [40] |
| hsa-mir-5683 | were found in all five viral SARS-CoV2 cases | [40] |
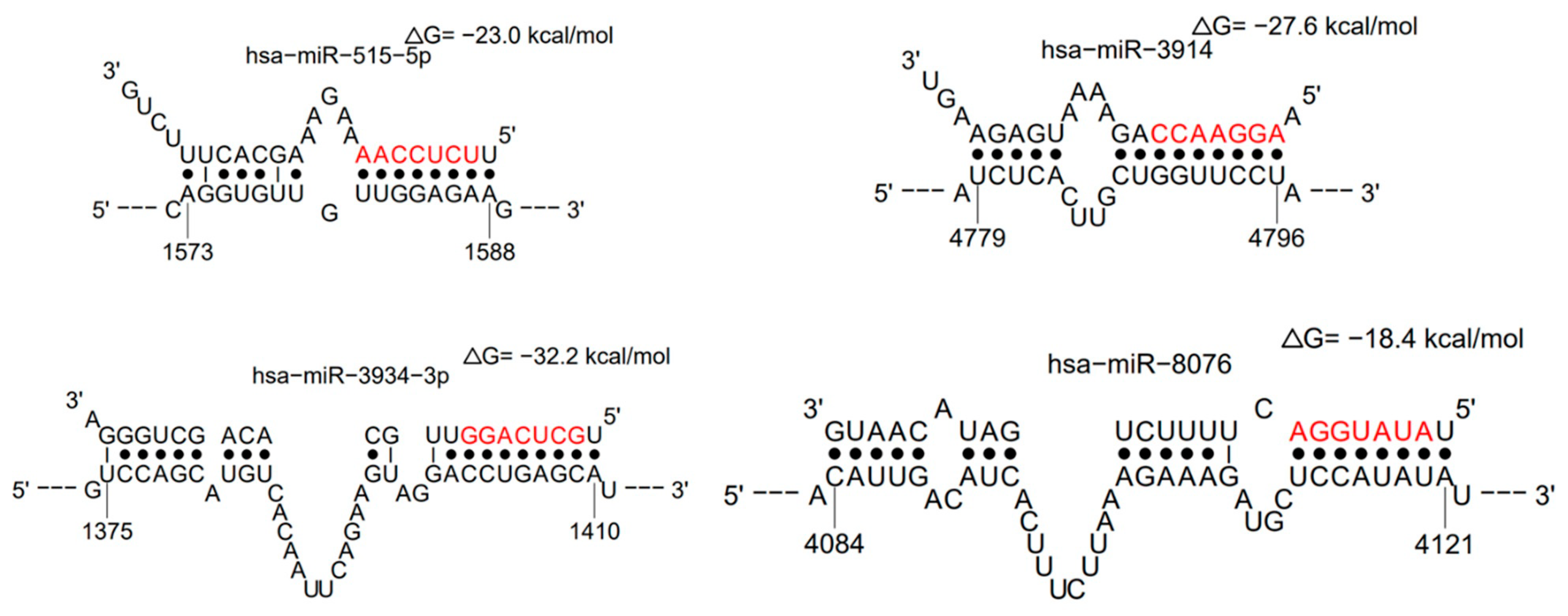
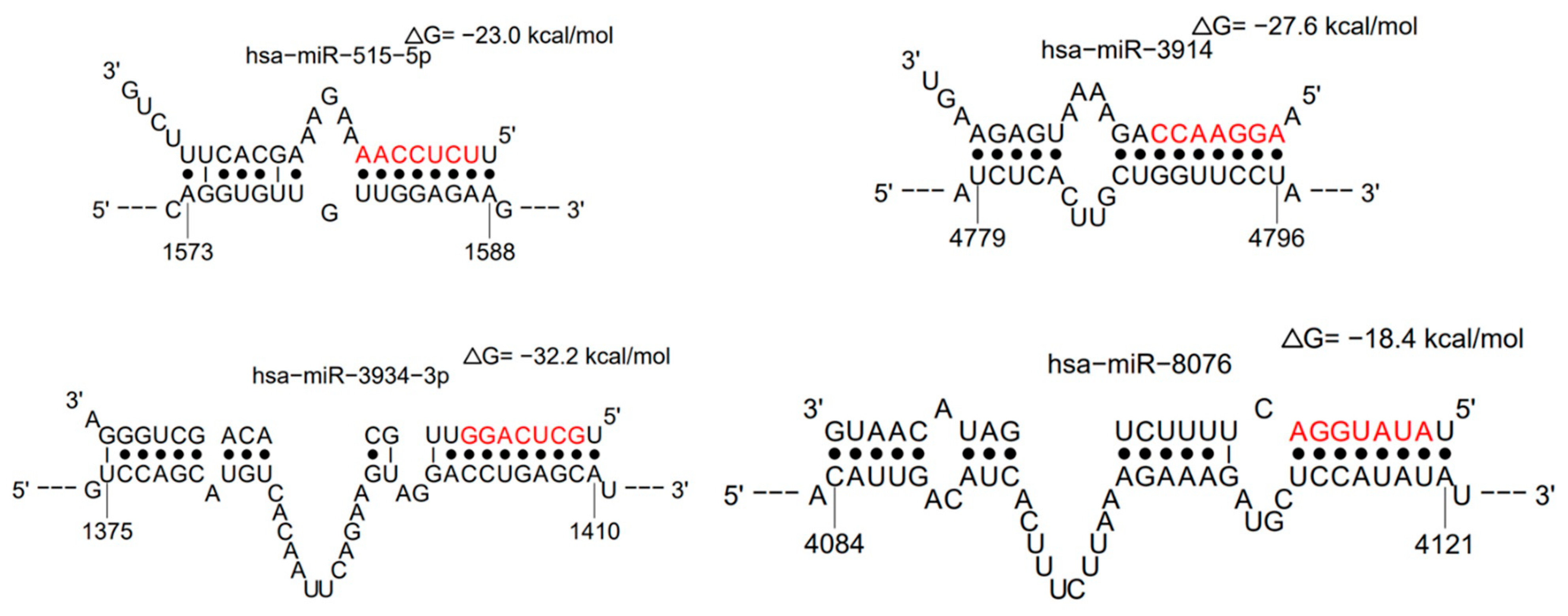
| miRNA | LogitProb | Seed_Position | dG_hybrid | Selected Gene Targets |
|---|---|---|---|---|
| hsa-miR-3914 | 0.945766785 | 4790–4796 | −27.6 | UBA6, FAM86B2, OSBPL9, GREM1, SLC25A28 |
| hsa-miR-515-5p | 0.873907643 | 1581–1587 | −23 | ZNF83, PI15, ZNF195, ZNF84, TRAPPC3L |
| hsa-miR-3934-3p | 0.862152158 | 1403–1409 | −32.2 | C14orf144, BOK, CDR1as, INHBC, NXF1 |
| hsa-miR-8076 | 0.820556086 | 4114–4120 | −18.4 | CRADD, SLC35E3, RP11-664D7.4, POLR2K, FAM13B |
| hsa-miR-4502 | 0.782521425 | 1875–1880 | −20.2 | ZNF23, ZNF616, ZNF225, ZNF544, GSG1L |
| hsa-miR-584-3p | 0.759965431 | 4370–4376 | −22.4 | IRAK4, IL18RAP, IL22RA2, TIRAP, IL12RB2 |
| hsa-miR-8066 | 0.758799201 | 4891–4896 | −13.6 | IL6ST, IL17A, IL1RL1, IRAK4, IRAK3 |
| hsa-miR-5197-3p | 0.751156198 | 4742–4747 | −15.9 | IRAK3, IL1A, TIRAP, IL15, IL7R |
| hsa-miR-1287-5p | 0.744896909 | 1375–1380 | −22.9 | IL7R, IL10RA, IL27RA, IL1RL1, IL12RB2 |
| hsa-miR-3613-5p | 0.731201689 | 3982–3987 | −18 | IL25, IL1RL1, IL17A, ILF3, IL1RAPL1 |
| hsa-miR-3611 | 0.723237424 | 3015–3021 | −18.2 | IL17F, IL6ST, TIRAP, IL1RL1, IL26 |
| hsa-miR-148b-3p | 0.718940862 | 2118–2123 | −15.8 | IL6ST, IL15, IL18BP, SOCS3, DOCK6 |
| hsa-miR-3120-5p | 0.709968406 | 4293–4298 | −30.2 | IRAK1, IL1A, IL13, IL17A, IL11 |
| hsa-miR-3120-5p | 0.703198793 | 3393–3398 | −17.4 | IRAK1, IL1A, IL13, IL17A, IL11 |
| hsa-miR-3611 | 0.696296501 | 2701–2707 | −13.9 | IL17F, IL6ST, TIRAP, IL1RL1, IL26 |
| hsa-miR-148b-3p | 0.690040719 | 1299–1304 | −25.1 | IL6ST, IL15, IL18BP, SOCS3, DOCK6 |
| hsa-miR-3120-5p | 0.673755775 | 3393–3398 | −17.5 | IRAK1, IL1A, IL13, IL17A, IL11 |
| hsa-miR-148b-3p | 0.669885166 | 902–907 | −17.2 | IL6ST, IL15, IL18BP, SOCS3, DOCK6 |
| hsa-miR-3120-5p | 0.665198433 | 2291–2296 | −19.4 | IRAK1, IL1A, IL13, IL17A, IL11 |
| hsa-miR-1287-5p | 0.663470144 | 1176–1181 | −22.6 | IL7R, IL10RA, IL27RA, IL1RL1, IL12RB2 |
| hsa-miR-584-3p | 0.650082156 | 3307–3312 | −17.2 | IRAK4, IL18RAP, IL22RA2, TIRAP, IL12RB2 |
| hsa-miR-3691-3p | 0.633230969 | 4612–4617 | −15.4 | IL1RAP, IL20, IL4R, IL7R, IL5RA |
| hsa-miR-8066 | 0.612297577 | 2022–2027 | -13.5 | IL6ST, IL17A, IL1RL1, IRAK4, IRAK3 |
| hsa-miR−3613-5p | 0.611909431 | 4978–4983 | −15 | IL25, IL1RL1, IL17A, ILF3, IL1RAPL1 |
| hsa-miR-3611 | 0.603752919 | 2021–2026 | −13.4 | IL17F, IL6ST, TIRAP, IL1RL1, IL26 |
| hsa-miR-3914 | 0.601515389 | 1589–1594 | −16 | UBA6, FAM86B2, OSBPL9, GREM1, SLC25A28 |
| hsa-miR-8066 | 0.592544568 | 3017–3022 | −14.7 | IL6ST, IL17A, IL1RL1, IRAK4, IRAK3 |
| hsa-miR-5197-3p | 0.592355306 | 4739–4744 | −15.7 | IRAK3, IL1A, TIRAP, IL15, IL7R |
| hsa-miR-3120-5p | 0.588547281 | 3257–3262 | −23.6 | IRAK1, IL1A, IL13, IL17A, IL11 |
| hsa-miR-3611 | 0.58763097 | 4698–4703 | −19.7 | IL17F, IL6ST, TIRAP, IL1RL1, IL26 |
| hsa-miR-515-5p | 0.587572941 | 1482–1487 | −19.2 | ZNF83, PI15, ZNF195, ZNF84, TRAPPC3L |
| hsa-miR-3914 | 0.583849164 | 1534–1539 | −16.7 | UBA6, FAM86B2, OSBPL9, GREM1, SLC25A28 |
| hsa-miR-5197-3p | 0.580206076 | 3168–3173 | −21.7 | IRAK3, IL1A, TIRAP, IL15, IL7R |
| hsa-miR-148b-3p | 0.56875777 | 714–719 | −18.9 | IL6ST, IL15, IL18BP, SOCS3, DOCK6 |
| hsa-miR-3914 | 0.561173741 | 4006–4011 | −13.8 | UBA6, FAM86B2, OSBPL9, GREM1, SLC25A28 |
| hsa-miR-378c | 0.547705178 | 1374–1379 | −20 | CD226, TFCP2L1, PAPOLG |
| hsa-miR-1287-5p | 0.545606444 | 4056–4061 | −14.8 | IL7R, IL10RA, IL27RA, IL1RL1, IL12RB2 |
| hsa-miR-148b-3p | 0.543191328 | 115–120 | −15.1 | IL6ST, IL15, IL18BP, SOCS3, DOCK6 |
| hsa-miR-3611 | 0.543063193 | 1984–1989 | −13.1 | IL17F, IL6ST, TIRAP, IL1RL1, IL26 |
| hsa-miR-8076 | 0.518454828 | 1100–1105 | −17.8 | CRADD, SLC35E3, RP11-664D7.4, POLR2K, FAM13B |
| hsa-miR-3120-5p | 0.503910403 | 4230–4235 | −20.7 | IRAK1, IL1A, IL13, IL17A, IL11 |
| hsa-miR-3120-5p | 0.502090677 | 1019–1024 | −18.8 | IRAK1, IL1A, IL13, IL17A, IL11 |
| hsa-miR-515-5p | 0.496816506 | 357–363 | −20.5 | ZNF83, PI15, ZNF195, ZNF84, TRAPPC3L |
| hsa-miR-1468-5p | 0.494920764 | 797–802 | −22.4 | IL7, IL12B, TIRAP |
| hsa-miR-1307-3p | 0.49446353 | 676–681 | −26.9 | IL18, IL20RB, IL10RA, IRAK2, IL6R |
| hsa-miR-5197-3p | 0.441353559 | 179–184 | −22 | IRAK3, IL1A, TIRAP, IL15, IL7R |
| hsa-miR-4502 | 0.433056019 | 474–479 | −21.5 | ZNF23, ZNF616, ZNF225, ZNF544, GSG1L |
| hsa-miR-3613-5p | 0.422107209 | 2553–2558 | −13 | IL25, IL1RL1, IL17A, ILF3, IL1RAPL1 |
| hsa-miR-1287-5p | 0.395329021 | 2431–2436 | −14.9 | IL7R, IL10RA, IL27RA, IL1RL1, IL12RB2 |
| hsa-miR-5197-3p | 0.384309463 | 628–633 | −19.4 | IRAK3, IL1A, TIRAP, IL15, IL7R |
| hsa-miR-4502 | 0.38319915 | 221–227 | −26 | ZNF23, ZNF616, ZNF225, ZNF544, GSG1L |
| hsa-miR-3691-3p | 0.37014701 | 574–579 | −20.2 | IL1RAP, IL20, IL4R, IL7R, IL5RA |
| hsa-miR-5197-3p | 0.352983944 | 2484–2489 | −14.6 | IRAK3, IL1A, TIRAP, IL15, IL7R |
| hsa-miR-584-3p | 0.352800085 | 495–500 | −18.8 | IRAK4, IL18RAP, IL22RA2, TIRAP, IL12RB2 |
| hsa-miR-515-5p | 0.329015553 | 267–272 | −18.3 | ZNF83, PI15, ZNF195, ZNF84, TRAPPC3L |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaspersic, J.; Dolzan, V. Viral and Host Genetic and Epigenetic Biomarkers Related to SARS-CoV-2 Cell Entry, Infection Rate, and Disease Severity. Biology 2022, 11, 178. https://doi.org/10.3390/biology11020178
Gaspersic J, Dolzan V. Viral and Host Genetic and Epigenetic Biomarkers Related to SARS-CoV-2 Cell Entry, Infection Rate, and Disease Severity. Biology. 2022; 11(2):178. https://doi.org/10.3390/biology11020178
Chicago/Turabian StyleGaspersic, Jernej, and Vita Dolzan. 2022. "Viral and Host Genetic and Epigenetic Biomarkers Related to SARS-CoV-2 Cell Entry, Infection Rate, and Disease Severity" Biology 11, no. 2: 178. https://doi.org/10.3390/biology11020178
APA StyleGaspersic, J., & Dolzan, V. (2022). Viral and Host Genetic and Epigenetic Biomarkers Related to SARS-CoV-2 Cell Entry, Infection Rate, and Disease Severity. Biology, 11(2), 178. https://doi.org/10.3390/biology11020178