
Anti-Malarial and Anti-Lipid Peroxidation Activities of Deferiprone-Resveratrol Hybrid in Plasmodium berghei-Infected Mice

 and
and
Abstract
Simple Summary
Abstract
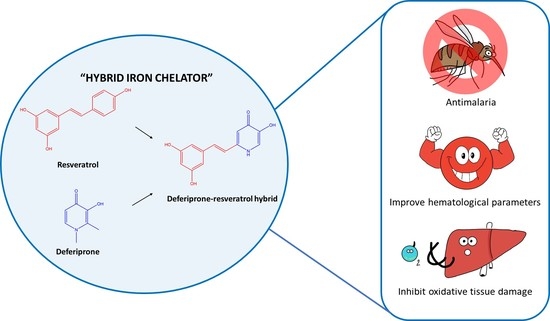
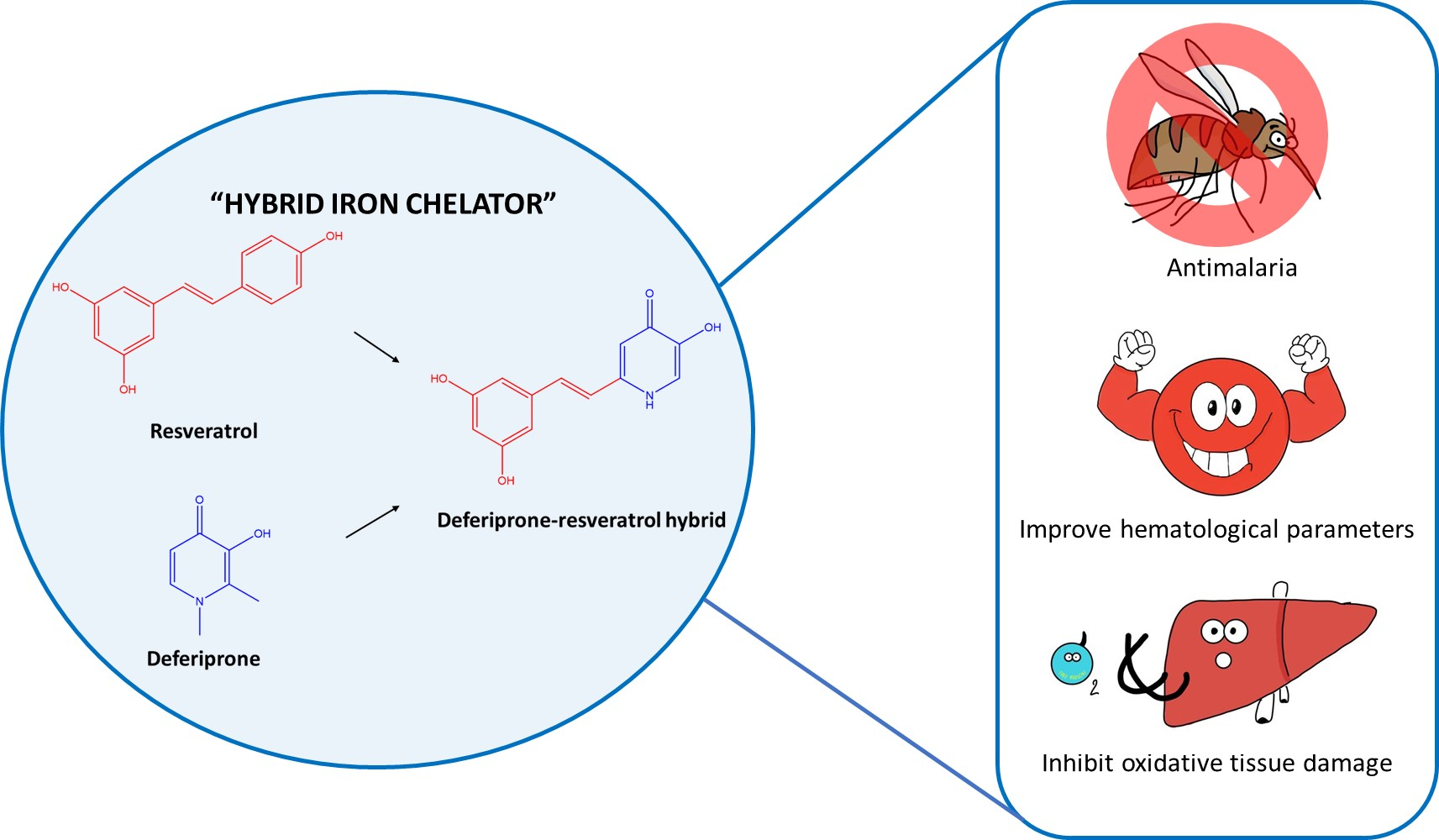
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Methods
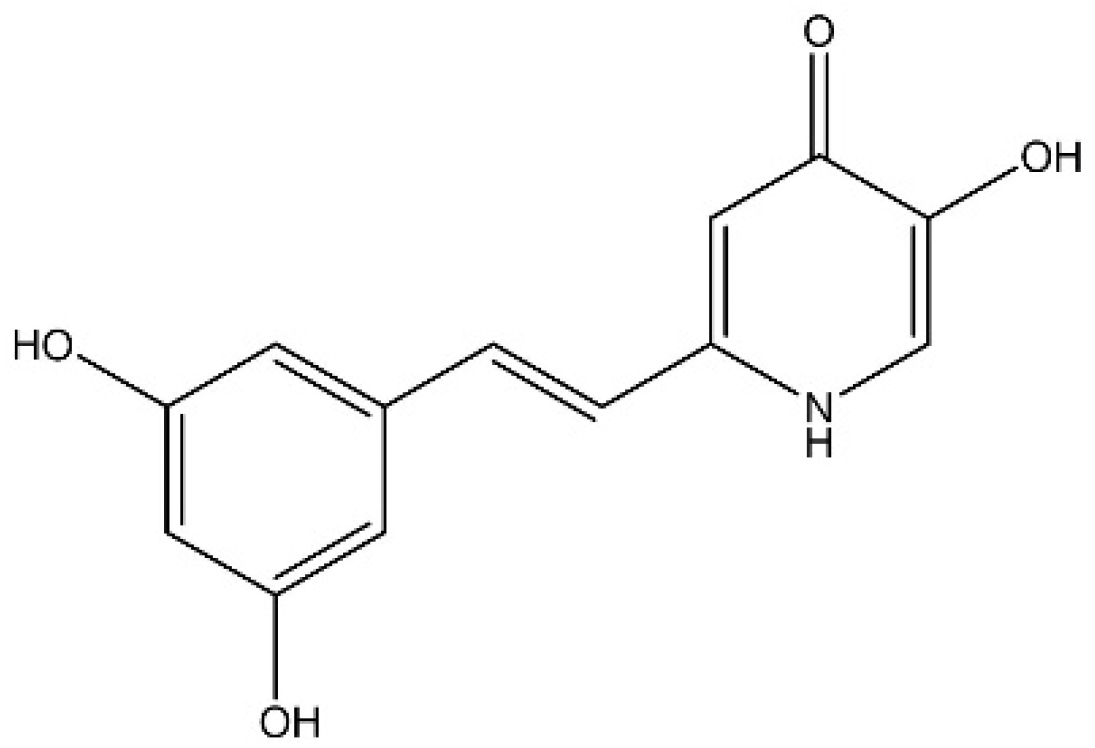
2.2.1. Deferiprone-Resveratrol Hybrid (DFP-RVT) Synthesis
2.2.2. Animal Ethics
2.2.3. Animal Care
2.2.4. Murine P. berghei Infection
2.2.5. Suppressive Activity Test of DFP-RVT in P. berghei-Infected Mice
2.2.6. Hematological Parameter Analysis
2.2.7. Measurement of Lipid-Peroxidation Product
2.2.8. Measurement of AST, ALT and ALP Activities
2.2.9. Statistical Analysis
3. Results
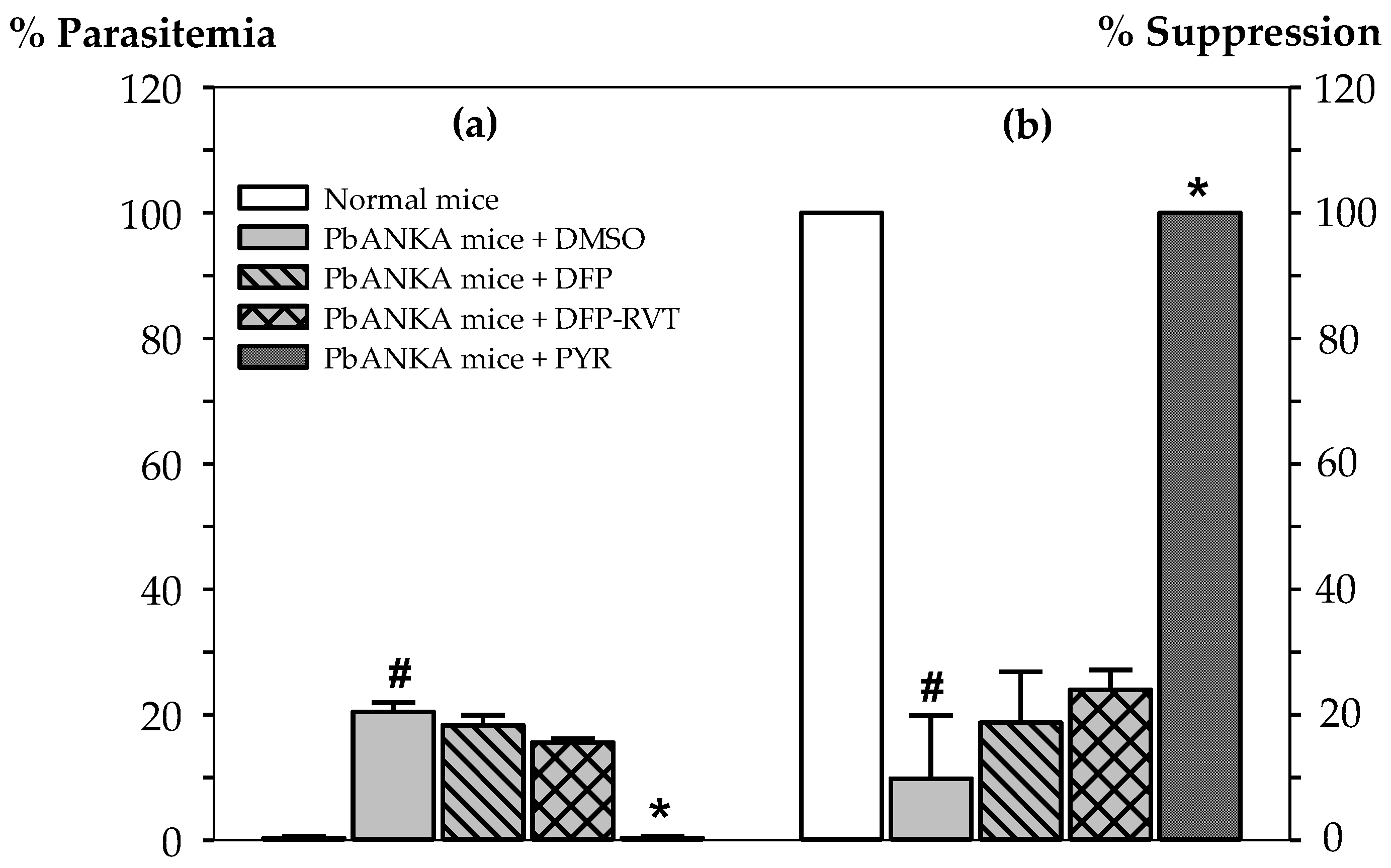
3.1. Malaria Suppressive Activity of DFP-RVT in PbANKA-Infected Mice
3.2. Changes of Hematological Parameters in PbANKA-Infected Mice
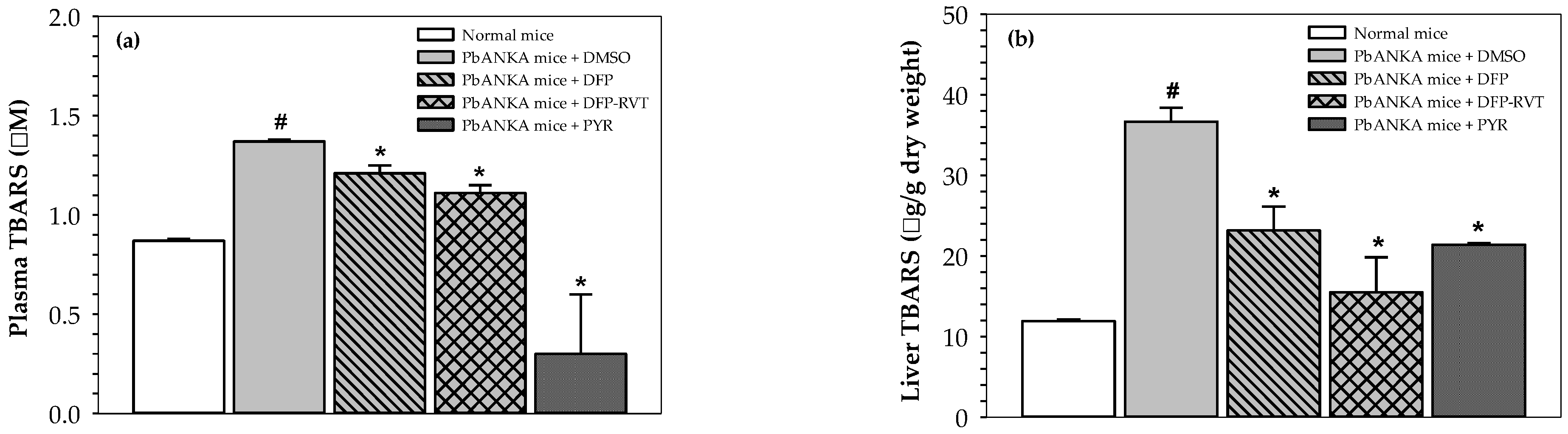
3.3. Inhibitory Effect of Lipid Peroxidation in PbANKA-Infected Mice
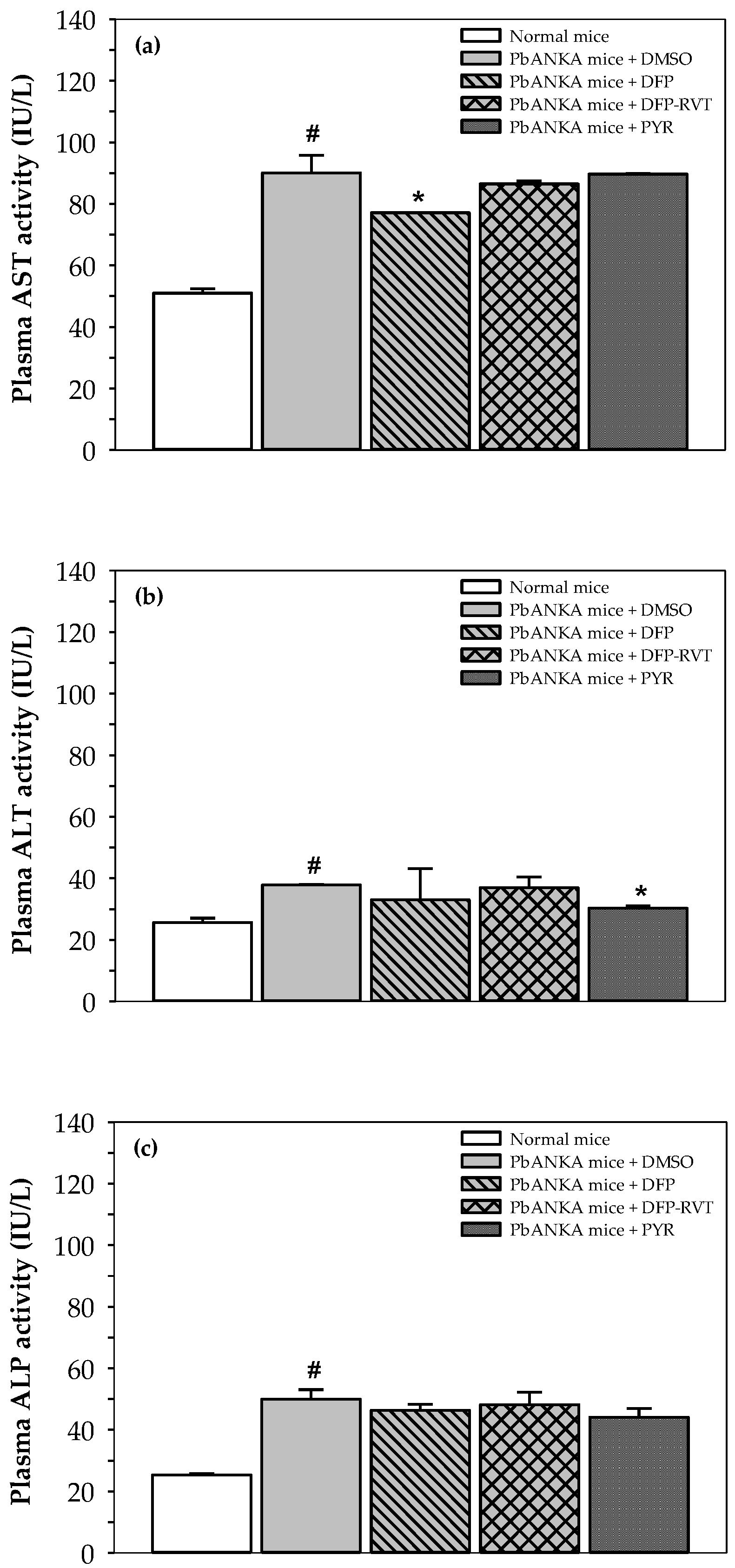
3.4. Liver Function Enzymes in PbANKA-Infected Mice
3.5. Body and Liver Weight Changes in PbANKA-Infected Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Athuman, M.; Kabanywanyi, A.M.; Rohwer, A.C. Intermittent preventive antimalarial treatment for children with anaemia. Cochrane Database Syst. Rev. 2015, 1, Cd010767. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.Q.; Ye, Q.; Huang, H.; Zheng, W.Y. An Overview of Available Antimalarials: Discovery, Mode of Action and Drug Resistance. Curr. Mol. Med. 2020, 20, 583–592. [Google Scholar] [CrossRef]
- Rout, S.; Mahapatra, R.K. Plasmodium falciparum: Multidrug resistance. Chem. Biol. Drug Des. 2019, 93, 737–759. [Google Scholar] [CrossRef] [PubMed]
- Asma, U.E.; Taufiq, F.; Khan, W. Prevalence and clinical manifestations of malaria in Aligarh, India. Korean J. Parasitol. 2014, 52, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Reisinger, E.C.; Horstmann, R.D.; Dietrich, M. Tolerance of mefloquine alone and in combination with sulfadoxine-pyrimethamine in the prophylaxis of malaria. Trans. R. Soc Trop. Med. Hyg. 1989, 83, 474–477. [Google Scholar] [CrossRef]
- Agrawal, A.; Tripathi, L.M.; Puri, S.K.; Pandey, V.C. Studies on ammonia-metabolizing enzymes during Plasmodium yoelii infection and pyrimethamine treatment in mice. Int. J. Parasitol. 1996, 26, 451–455. [Google Scholar] [CrossRef]
- Agrawal, A.; Tripathi, L.M.; Pandey, V.C. Status of urea and related enzymes during Plasmodium yoelii infection and pyrimethamine treatment in mice. Indian J. Exp. Biol. 1997, 35, 1194–1197. [Google Scholar]
- Ali, V.; Nozaki, T. Iron-sulphur clusters, their biosynthesis, and biological functions in protozoan parasites. Adv. Parasitol. 2013, 83, 1–92. [Google Scholar]
- Testi, C.; Boffi, A.; Montemiglio, L.C. Structural analysis of the transferrin receptor multifaceted ligand(s) interface. Biophys. Chem. 2019, 254, 106242. [Google Scholar] [CrossRef]
- Shanzer, A.; Libman, J.; Lytton, S.D.; Glickstein, H.; Cabantchik, Z.I. Reversed siderophores act as antimalarial agents. Proc. Natl. Acad. Sci. USA 1991, 88, 6585–6589. [Google Scholar] [CrossRef]
- Sanchez-Lopez, R.; Haldar, K. A transferrin-independent iron uptake activity in Plasmodium falciparum-infected and uninfected erythrocytes. Mol. Biochem. Parasitol. 1992, 55, 9–20. [Google Scholar] [CrossRef]
- Lytton, S.D.; Cabantchik, Z.I.; Libman, J.; Shanzer, A. Reversed siderophores as antimalarial agents. II. Selective scavenging of Fe(III) from parasitized erythrocytes by a fluorescent derivative of desferal. Mol. Pharmacol. 1991, 40, 584–590. [Google Scholar] [PubMed]
- Hershko, C.; Gordeuk, V.R.; Thuma, P.E.; Theanacho, E.N.; Spira, D.T.; Hider, R.C.; Peto, T.E.; Brittenham, G.M. The antimalarial effect of iron chelators: Studies in animal models and in humans with mild falciparum malaria. J. Inorg. Biochem. 1992, 47, 267–277. [Google Scholar] [CrossRef]
- Smith, H.J.; Meremikwu, M.M. Iron-chelating agents for treating malaria. Cochrane Database Syst. Rev. 2003, 2003, CD001474. [Google Scholar] [CrossRef] [PubMed]
- Pradines, B.; Rolain, J.M.; Ramiandrasoa, F.; Fusai, T.; Mosnier, J.; Rogier, C.; Daries, W.; Baret, E.; Kunesch, G.; Le Bras, J.; et al. Iron chelators as antimalarial agents: In vitro activity of dicatecholate against Plasmodium falciparum. J. Antimicrob. Chemother. 2002, 50, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Maneekesorn, S.; Chuljerm, H.; Koonyosying, P.; Uthaipibull, C.; Ma, Y.; Srichairatanakool, S. Identifying a Deferiprone–Resveratrol Hybrid as an Effective Lipophilic Anti-Plasmodial Agent. Molecules 2021, 26, 4074. [Google Scholar] [CrossRef] [PubMed]
- Scheibel, L.W.; Stanton, G.G. Antimalarial activity of selected aromatic chelators. IV. Cation uptake by Plasmodium falciparum in the presence of oxines and siderochromes. Mol. Pharmacol. 1986, 30, 364–369. [Google Scholar] [PubMed]
- Hammadi, A.; Ramiandrasoa, F.; Sinou, V.; Rogier, C.; Fusai, T.; Le Bras, J.; Parzy, D.; Kunesch, G.; Pradines, B. Cellular uptake of a catechol iron chelator and chloroquine into Plasmodium falciparum infected erythrocytes. Biochem. Pharmacol. 2003, 65, 1351–1360. [Google Scholar] [CrossRef]
- Thipubon, P.; Uthaipibull, C.; Kamchonwongpaisan, S.; Tipsuwan, W.; Srichairatanakool, S. Inhibitory effect of novel iron chelator, 1-(N-acetyl-6-aminohexyl)-3-hydroxy-2-methylpyridin-4-one (CM1) and green tea extract on growth of Plasmodium falciparum. Malar. J. 2015, 14, 382. [Google Scholar] [CrossRef] [PubMed]
- Buffinton, G.D.; Hunt, N.H.; Cowden, W.B.; Clark, I.A. Detection of short-chain carbonyl products of lipid peroxidation from malaria-parasite (Plasmodium vinckei)-infected red blood cells exposed to oxidative stress. Biochem. J. 1988, 249, 63–68. [Google Scholar] [CrossRef]
- Postma, N.S.; Mommers, E.C.; Eling, W.M.; Zuidema, J. Oxidative stress in malaria; Implications for prevention and therapy. Pharm. World Sci. PWS 1996, 18, 121–129. [Google Scholar] [CrossRef]
- Nsiah, K.; Bahaah, B.; Oppong Afranie, B.; Koffie, S.; Akowuah, E.; Donkor, S. Oxidative Stress and Hemoglobin Level of Complicated and Uncomplicated Malaria Cases among Children: A Cross-Sectional Study in Kumasi Metropolis, Ghana. J. Trop. Med. 2019, 2019, 8479076. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Zhang, M.; Sheng, R.; Ma, Y. Synthesis and biological evaluation of deferiprone-resveratrol hybrids as antioxidants, Abeta1-42 aggregation inhibitors and metal-chelating agents for Alzheimer’s disease. Eur. J. Med. Chem. 2017, 127, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Ounjaijean, S.; Romyasamit, C.; Somsak, V. Evaluation of Antimalarial Potential of Aqueous Crude Gymnema Inodorum Leaf Extract against Plasmodium berghei Infection in Mice. Evid. Based Complementary Altern. Med. 2021, 2021, 9932891. [Google Scholar] [CrossRef]
- Somsak, V.; Jaihan, U.; Srichairatanakool, S.; Uthaipibull, C. Protection of renal function by green tea extract during Plasmodium berghei infection. Parasitol. Int. 2013, 62, 548–551. [Google Scholar] [CrossRef][Green Version]
- Tarkang, P.A.; Okalebo, F.A.; Ayong, L.S.; Agbor, G.A.; Guantai, A.N. Anti-malarial activity of a polyherbal product (Nefang) during early and established Plasmodium infection in rodent models. Malar. J. 2014, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Knight, D.J.; Peters, W. The antimalarial activity of N-benzyloxydihydrotriazines. Ann. Trop. Med. Parasitol. 1980, 74, 393–404. [Google Scholar] [CrossRef]
- Thipubon, P.; Tipsuwan, W.; Uthaipibull, C.; Santitherakul, S.; Srichairatanakool, S. Anti-malarial effect of 1-(N-acetyl-6-aminohexyl)-3-hydroxy-2-methylpyridin-4-one and green tea extract on erythrocyte-stage Plasmodium berghei in mice. Asian Pac. J. Trop. Biomed. 2015, 5, 932–936. [Google Scholar] [CrossRef]
- Koonyosying, P.; Kongkarnka, S.; Uthaipibull, C.; Svasti, S.; Fucharoen, S.; Srichairatanakool, S. Green tea extract modulates oxidative tissue injury in beta-thalassemic mice by chelation of redox iron and inhibition of lipid peroxidation. Biomed. Pharm. 2018, 108, 1694–1702. [Google Scholar] [CrossRef]
- Tietz, N.W.; Wekstein, D.R.; Shuey, D.F.; Brauer, G.A. A two-year longitudinal reference range study for selected serum enzymes in a population more than 60 years of age. J. Am. Geriatr. Soc. 1984, 32, 563–570. [Google Scholar] [CrossRef]
- Tietz, N.W.; Rinker, A.D.; Shaw, L.M. IFCC methods for the measurement of catalytic concentration of enzymes Part 5. IFCC method for alkaline phosphatase (orthophosphoric-monoester phosphohydrolase, alkaline optimum, EC 3.1.3.1). J. Clin. Chem. Clin. Biochem. 1983, 21, 731–748. [Google Scholar] [PubMed]
- Cronin, S.J.F.; Woolf, C.J.; Weiss, G.; Penninger, J.M. The Role of Iron Regulation in Immunometabolism and Immune-Related Disease. Front. Mol. Biosci. 2019, 6, 116. [Google Scholar] [CrossRef] [PubMed]
- Tsafack, A.; Libman, J.; Shanzer, A.; Cabantchik, Z.I. Chemical Determinants of antimalarial activity of reversed siderophores. Antimicrob. Agents Chemother. 1996, 40, 2160–2166. [Google Scholar] [CrossRef]
- Węglińska, L.; Bekier, A.; Dzitko, K.; Pacholczyk-Sienicka, B.; Albrecht, Ł.; Plech, T.; Paneth, P.; Paneth, A. 1,3,4-Thiadiazoles Effectively Inhibit Proliferation of Toxoplasma gondii. Cells 2021, 10, 1053. [Google Scholar] [CrossRef]
- Ferrer, P.; Tripathi, A.K.; Clark, M.A.; Hand, C.C.; Rienhoff, H.Y., Jr.; Sullivan, D.J., Jr. Antimalarial Iron Chelator, FBS0701, Shows Asexual and Gametocyte Plasmodium falciparum Activity and Single Oral Dose Cure in a Murine Malaria Model. PLoS ONE 2012, 7, e37171. [Google Scholar] [CrossRef]
- Wilson, S.; Jones, F.M.; Mwatha, J.K.; Kimani, G.; Booth, M.; Kariuki, H.C.; Vennervald, B.J.; Ouma, J.H.; Muchiri, E.; Dunne, D.W. Hepatosplenomegaly associated with chronic malaria exposure: Evidence for a pro-inflammatory mechanism exacerbated by schistosomiasis. Parasite Immunol. 2009, 3, 64–71. [Google Scholar] [CrossRef]
- Walters, J.H.; McGregor, I.A. The mechanism of malarial hepatomegaly and its relationship to hepatic fibrosis. Trans. R. Soc. Trop. Med. Hyg. 1960, 54, 135–145. [Google Scholar] [CrossRef]
- Black, R.H. Enlargement of the liver due to malaria in natives of the South-West pacific area. Trans. R. Soc. Trop. Med. Hyg. 1954, 48, 226–233. [Google Scholar] [CrossRef]
- McGilvray, I.D.; Serghides, L.; Kapus, A.; Rotstein, O.D.; Kain, K.C. Nonopsonic monocyte/macrophage phagocytosis of Plasmodium falciparum–parasitized erythrocytes: A role for CD36 in malarial clearance. Blood 2000, 96, 3231–3240. [Google Scholar] [CrossRef]
- George, E.L.; Panos, A. Does a high WBC count signal infection? Nursing 2005, 2006, 20–21. [Google Scholar] [CrossRef]
- Lamikanra, A.A.; Brown, D.; Potocnik, A.; Casals-Pascual, C.; Langhorne, J.; Roberts, D.J. Malarial anemia: Of mice and men. Blood 2007, 110, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Narsaria, N.; Mohanty, C.; Das, B.K.; Mishra, S.P.; Prasad, R. Oxidative Stress in Children with Severe Malaria. J. Trop. Pediatrics 2012, 58, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Legorreta-Herrera, M.; Retana-Ugalde, R.; Ventura-Gallegos, J.L.; Narvaez, V. Pyrimethamine induces oxidative stress in Plasmodium yoelii 17XL-infected mice: A novel immunomodulatory mechanism of action for an old antimalarial drug? Exp. Parasitol. 2010, 126, 381–388. [Google Scholar] [CrossRef]
- Loyevsky, M.; Sacci, J.B., Jr.; Boehme, P.; Weglicki, W.; John, C.; Gordeuk, V.R. Plasmodium falciparum and Plasmodium yoelii: Effect of the iron chelation prodrug dexrazoxane on in vitro cultures. Exp. Parasitol. 1999, 91, 105–114. [Google Scholar] [CrossRef]
- Glickstein, H.; Breuer, W.; Loyevsky, M.; Konijn, A.M.; Shanzer, A.; Cabantchik, Z.I. Differential cytotoxicity of iron chelators on malaria-infected cells versus mammalian cells. Blood 1996, 87, 4871–4878. [Google Scholar] [CrossRef]
- Abro, A.H.; Ustadi, A.M.; Abro, H.A.; Abdou, A.S.; Younis, N.J.; Akaila, S.I. Jaundice with hepatic dysfunction in P. falciparum malaria. J. Coll. Physicians Surg. Pak. 2009, 19, 363–366. [Google Scholar] [PubMed]
- Srivastava, P.; Pandey, V.C. Studies on hepatic mitochondrial cytochrome P-450 during Plasmodium yoelii infection and pyrimethamine treatment in mice. Ecotoxicol. Environ. Saf. 2000, 46, 19–22. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Total and Differential WBC Coun | |||||||
| Mice/Treatment | WBC Numbers (×103/mL) | Differential WBC (%) | Mice/Treatment | WBC Numbers (×103/mL) | Differential WBC (%) | Mice/Treatment | - |
| Neutrophil | Neutrophil | - | |||||
| Normal/DI | 3.38 ± 0.33 | 22.0 ± 0.6 | 74.0 ± 0.4 | 4.0 ± 0.0 | 0 | 0 | - |
| PbANKA/60% DMSO | 10.18 ± 0.04 # | 70.0 ± 6.5 # | 33.7 ± 8.7 | 3.0 ± 0.0 | 1.0 ± 0.6 | 0.0 ± 0.0 | - |
| PbANKA/DFP (50 mg/kg BW) | 3.25 ± 0.07 * | 17.0 ± 2.4 * | 37.7 ± 9.9 | 25.7 ± 12.2 | 1.0 ± 0.6 | 0.3 ± 0.3 | - |
| PbANKA/DFP-RVT (50 mg/kg BW) | 4.38 ± 0.44 * | 64.5 ± 4.5 | 27.0 ± 5.3 | 2.7 ± 0.3 | 1.0 ± 0.0 | 0.3 ± 0.3 | - |
| PbANKA/PYR (2 mg/kg BW) | 2.81 ± 0.07 * | 29.0 ± 1.6 * | 57.5 ± 2.9 * | 12.0 ± 4.9* | 1.0 ± 0.0 | 0.5 ± 0.4 | - |
| (b) RBC Indices | |||||||
| Mice/Treatment | RBC Numbers (×106/mL) | Hb (g/dL) | Hct (%) | MCV (fL) | MCH (pg) | MCHC (g/dL) | RDW (%) |
| Normal/DI | 9.70 ± 0.01 | 14.70 ± 0.3 | 51.8 ± 0.4 | 53.0 ± 0.0 | 15.0 ± 0.0 | 28.0 ± 0.0 | 15.0 ± 0.0 |
| PbANKA/60% DMSO | 7.03 ± 0.07 # | 12.6 ± 0.0 # | 44.0 ± 0.0 # | 62.7 ± 0.7 # | 18.0 ± 0.0 # | 28.7 ± 0.3 | 15.0 ± 0.6 |
| PbANKA/DFP (50 mg/kg BW) | 7.72 ± 0.49 * | 13.2 ± 3.9 | 46.0 ± 2.4 | 60.3 ± 0.7 | 17.7 ± 0.7 | 29.3 ± 0.0 | 14.0 ± 0.6 |
| PbANKA/DFP-RVT (50 mg/kg BW) | 7.91 ± 0.41 * | 13.6 ± 0.5 | 47.5 ± 0.4 | 60.7 ± 2.6 | 17.3 ± 0.3 | 28.7 ± 1.2 | 14.7 ± 0.3 |
| PbANKA/PYR (2 mg/kg BW) | 9.19 ± 0.91 * | 14.6 ± 1.6 * | 49.5 ± 5.3 | 61.0 ± 0.0 | 18.0 ± 0.0 | 30.0 ± 0.0 | 14.5 ± 1.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuljerm, H.; Maneekesorn, S.; Somsak, V.; Ma, Y.; Srichairatanakool, S.; Koonyosying, P. Anti-Malarial and Anti-Lipid Peroxidation Activities of Deferiprone-Resveratrol Hybrid in Plasmodium berghei-Infected Mice. Biology 2021, 10, 911. https://doi.org/10.3390/biology10090911
Chuljerm H, Maneekesorn S, Somsak V, Ma Y, Srichairatanakool S, Koonyosying P. Anti-Malarial and Anti-Lipid Peroxidation Activities of Deferiprone-Resveratrol Hybrid in Plasmodium berghei-Infected Mice. Biology. 2021; 10(9):911. https://doi.org/10.3390/biology10090911
Chicago/Turabian StyleChuljerm, Hataichanok, Supawadee Maneekesorn, Voravuth Somsak, Yongmin Ma, Somdet Srichairatanakool, and Pimpisid Koonyosying. 2021. "Anti-Malarial and Anti-Lipid Peroxidation Activities of Deferiprone-Resveratrol Hybrid in Plasmodium berghei-Infected Mice" Biology 10, no. 9: 911. https://doi.org/10.3390/biology10090911
APA StyleChuljerm, H., Maneekesorn, S., Somsak, V., Ma, Y., Srichairatanakool, S., & Koonyosying, P. (2021). Anti-Malarial and Anti-Lipid Peroxidation Activities of Deferiprone-Resveratrol Hybrid in Plasmodium berghei-Infected Mice. Biology, 10(9), 911. https://doi.org/10.3390/biology10090911