
The Genetic Network of Forkhead Gene Family in Development of Brown Planthoppers

 and
and
Abstract
:Simple Summary
Abstract
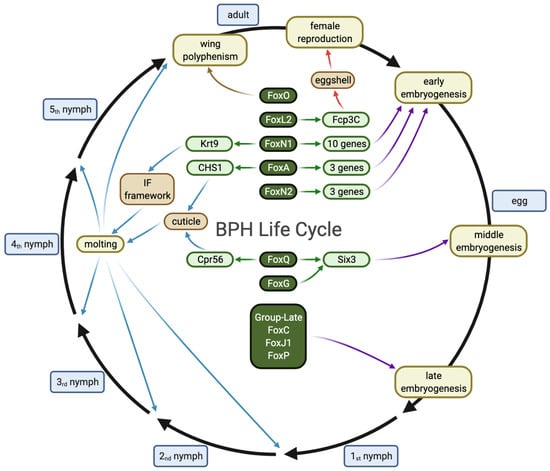
{kind=link}
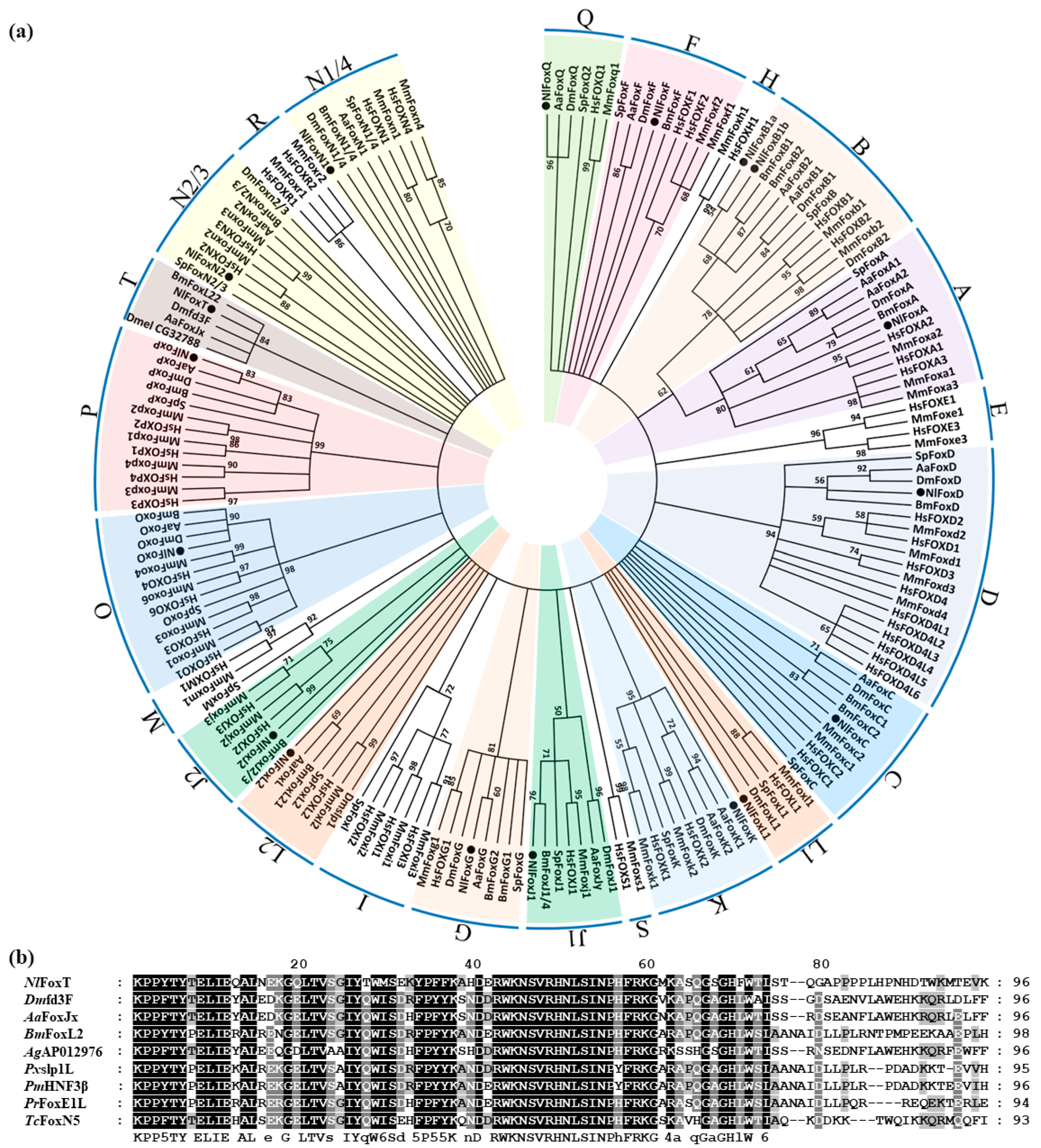{kind=link}
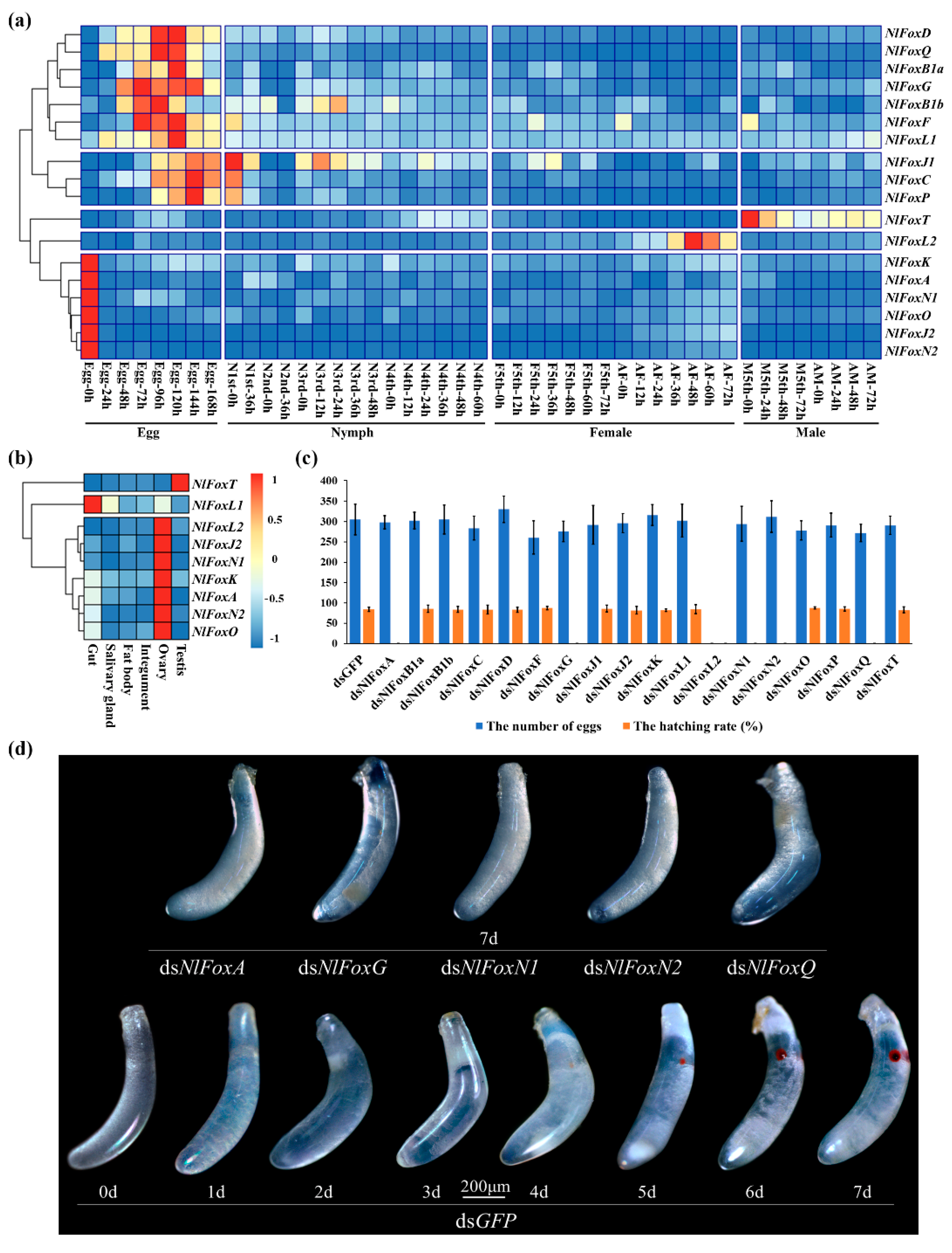{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Gene Identification
2.2. Sequence Analysis
2.3. Real-Time Quantitative PCR (qRT-PCR) Analysis
2.4. Expression Pattern Analysis
2.5. RNAi Effects
2.6. RNA-Seq
3. Results
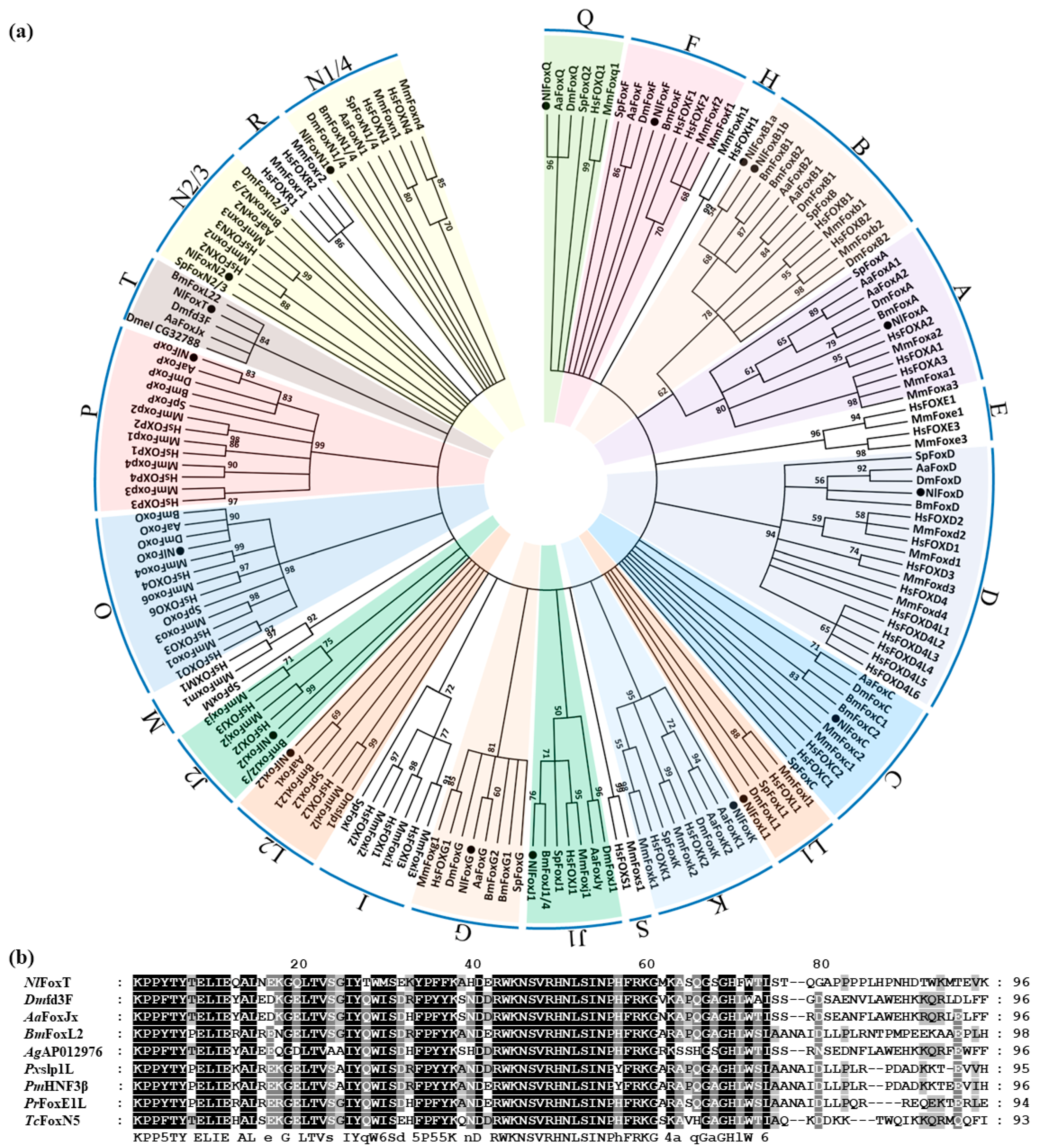
3.1. Identification and Sequence Analyses of Forkhead Genes in the Brown Planthopper
3.2. Temporal and Spatial Expression Patterns
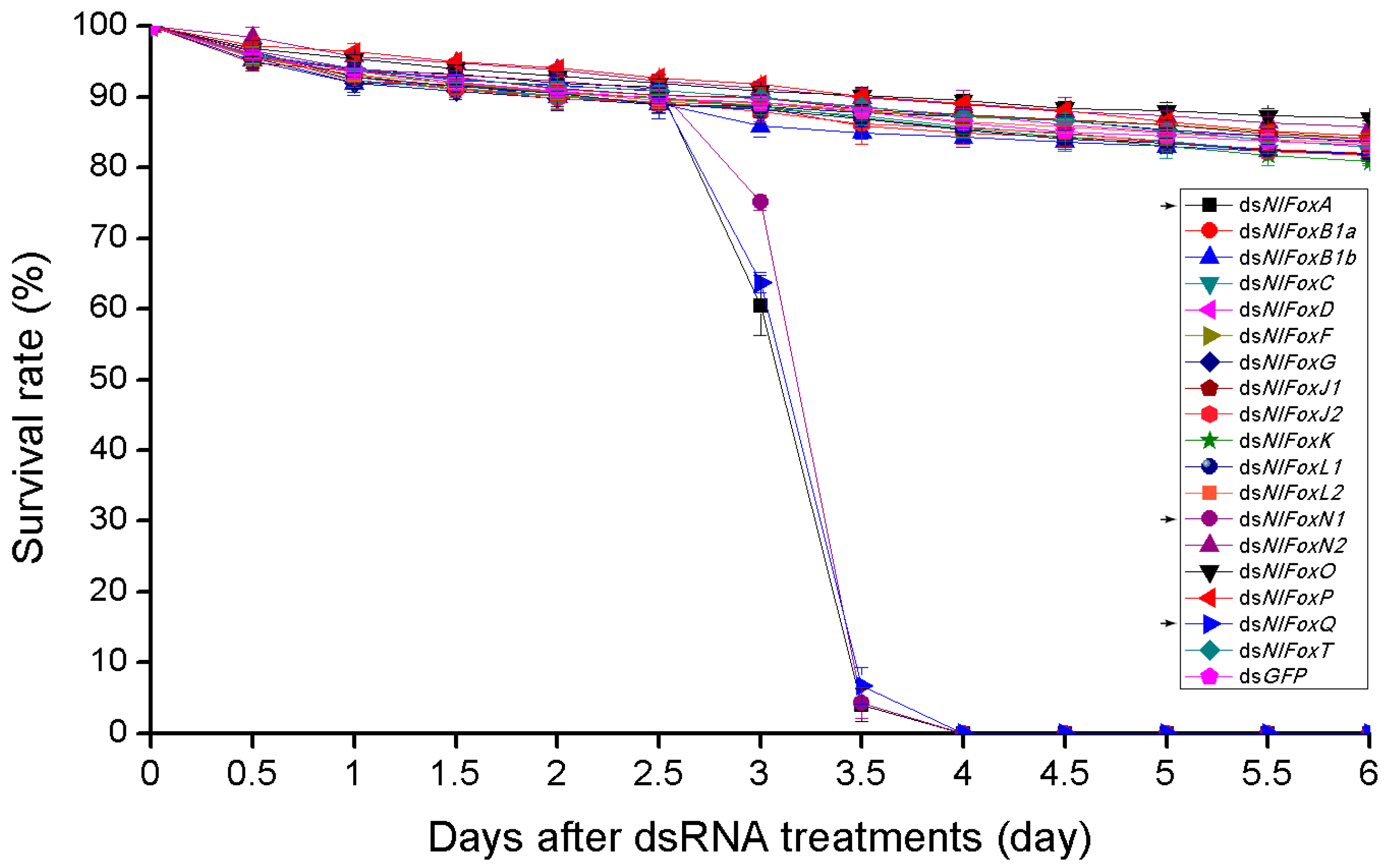
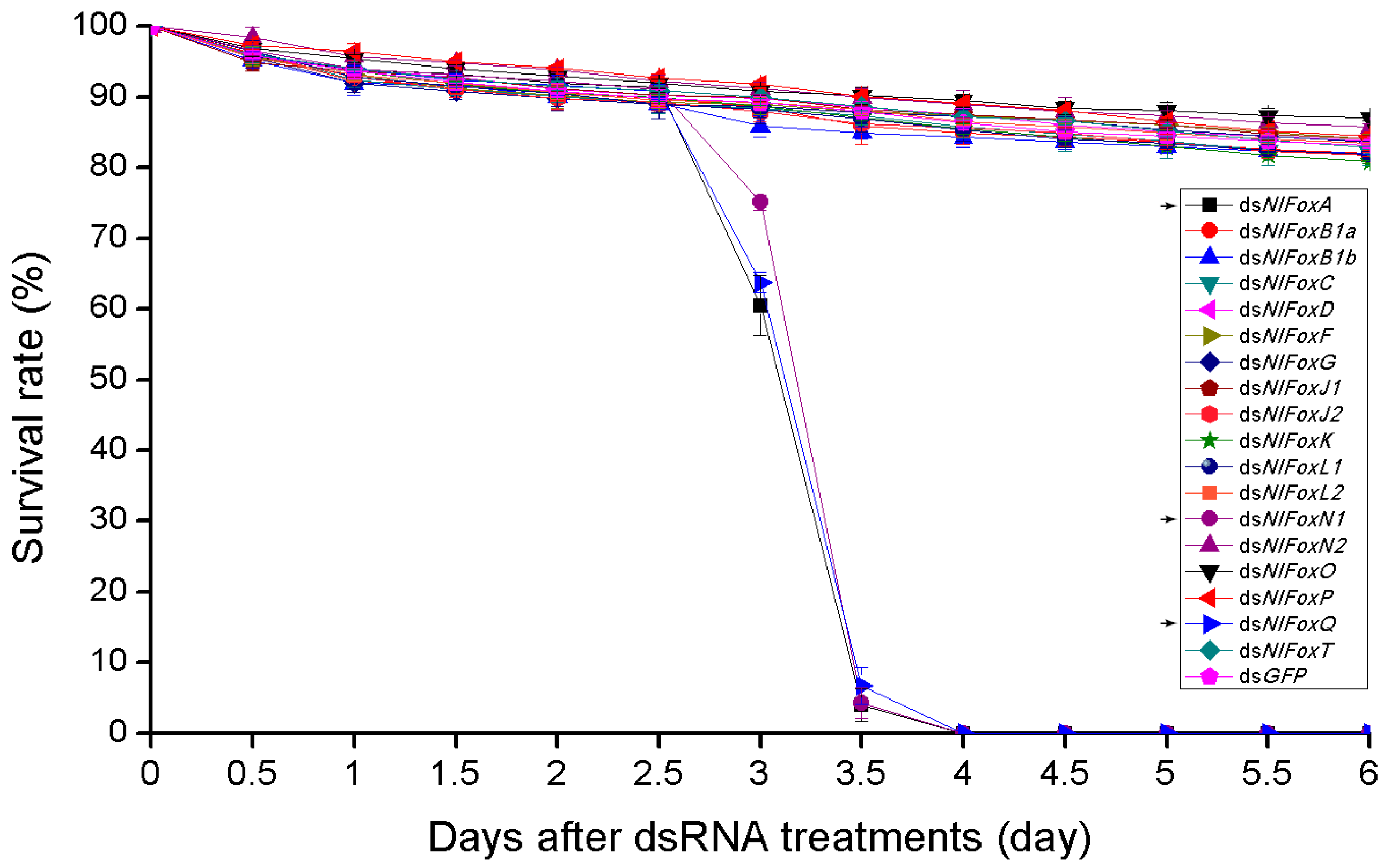
3.3. NlFoxG and NlFoxN2 Are Indispensable for Embryogenesis
3.4. Pleiotropic Functions of Fox Genes (A/N1/Q) and Their Potential Targets in the Brown Planthopper
3.5. Complementary Functions of NlFoxC, NlFoxJ1 and NlFoxP in the Late Embryonic Stage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaestner, K.H.; Knochel, W.; Martinez, D.E. Unified nomenclature for the winged helix/forkhead transcription factors. Genes Dev. 2000, 14, 142–146. [Google Scholar] [PubMed]
- Weigel, D.; Jürgens, G.; Küttner, F.; Seifert, E.; Jäckle, H. The homeotic gene fork head encodes a nuclear protein and is expressed in the terminal regions of the Drosophila embryo. Cell 1989, 57, 645–658. [Google Scholar] [CrossRef] [Green Version]
- Weigel, D.; Jäckle, H. The fork head domain: A novel DNA binding motif of eukaryotic transcription factors? Cell 1990, 63, 455. [Google Scholar] [CrossRef]
- Carlsson, P.; Mahlapuu, M. Forkhead Transcription Factors: Key Players in Development and Metabolism. Dev. Biol. 2002, 250, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Lam, E.W.-F.; Brosens, J.; Gomes, A.R.; Koo, C.Y. Forkhead box proteins: Tuning forks for transcriptional harmony. Nat. Rev. Cancer 2013, 13, 482–495. [Google Scholar] [CrossRef]
- Jackson, B.C.; Carpenter, C.; Nebert, D.W.; Vasiliou, V. Update of human and mouse forkhead box (FOX) gene families. Hum. Genom. 2010, 4, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Hacker, U.; Grossniklaus, U.; Gehring, W.J.; Jackle, H. Developmentally Regulated Drosophila Gene Family Encoding the Fork Head Domain. Proc. Natl. Acad. Sci. USA 1992, 89, 8754–8758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fetting, J.L.; Guay, J.A.; Karolak, M.J.; Iozzo, R.; Adams, D.C.; Maridas, D.E.; Brown, A.C.; Oxburgh, L. FOXD1 promotes nephron progenitor differentiation by repressing decorin in the embryonic kidney. Development 2014, 141, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Vaidya, H.J.; Leon, A.B.; Blackburn, C.C. FOXN1 in thymus organogenesis and development. Eur. J. Immunol. 2016, 46, 1826–1837. [Google Scholar] [CrossRef]
- Kume, T.; Deng, K.; Hogan, B.L. Murine forkhead/winged helix genes Foxc1 (Mf1) and Foxc2 (Mfh1) are required for the early organogenesis of the kidney and urinary tract. Development 2000, 127, 1387. [Google Scholar] [CrossRef] [PubMed]
- Hannenhalli, S.; Kaestner, K.H. The evolution of Fox genes and their role in development and disease. Nat. Rev. Genet. 2009, 10, 233–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritzenwanker, J.H.; Gerhart, J.; Freeman, R.M.; Lowe, C.J. The Fox/Forkhead transcription factor family of the hemichordate Saccoglossus kowalevskii. EvoDevo 2014, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazet, F.; Yu, J.-K.; Liberles, D.A.; Holland, L.Z.; Shimeld, S.M. Phylogenetic relationships of the Fox (Forkhead) gene family in the Bilateria. Gene 2003, 316, 79–89. [Google Scholar] [CrossRef]
- Song, J.; Li, Z.; Tong, X.; Chen, C.; Chen, M.; Meng, G.; Chen, P.; Li, C.; Xin, Y.; Gai, T.; et al. Genome-wide identification and characterization of Fox genes in the silkworm, Bombyx mori. Funct. Integr. Genom. 2015, 15, 511–522. [Google Scholar] [CrossRef]
- Hansen, I.A.; Sieglaff, D.H.; Munro, J.; Shiao, S.-H.; Cruz, J.; Lee, I.W.; Heraty, J.M.; Raikhel, A.S. Forkhead transcription factors regulate mosquito reproduction. Insect Biochem. Mol. Biol. 2007, 37, 985–997. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Zhang, H.; Li, D.; Zhuo, J.; Shen, Y.; Hu, Q.; Zhang, C. Chromosome-level assembly of the brown planthopper genome with a characterized Y chromosome. Mol. Ecol. Resour. 2021, 21, 1287–1298. [Google Scholar] [CrossRef]
- Paul, F.; Arkin, Y.; Giladi, A.; Jaitin, D.A.; Kenigsberg, E.; Keren-Shaul, H.; Winter, D.; Lara-Astiaso, D.; Gury, M.; Weiner, A.; et al. Transcriptional Heterogeneity and Lineage Commitment in Myeloid Progenitors. Cell 2015, 163, 1663–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, M.; Blackshields, G.; Brown, N.; Chenna, R.; McGettigan, P.; McWilliam, H.; Valentin, F.; Wallace, I.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Xu, H.-J.; Xue, J.; Lu, B.; Zhang, X.-C.; Zhuo, J.-C.; He, S.-F.; Ma, X.-F.; Jiang, Y.-Q.; Fan, H.-W.; Xu, J.-Y.; et al. Two insulin receptors determine alternative wing morphs in planthoppers. Nature 2015, 519, 464–467. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, M.P.D.K.G.M.P.S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S.J.G.B. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.; Tindall, D.J. FOXOs, cancer and regulation of apoptosis. Oncogene 2008, 27, 2312–2319. [Google Scholar] [CrossRef] [Green Version]
- Pan, P.-L.; Ye, Y.-X.; Lou, Y.-H.; Lu, J.-B.; Cheng, C.; Shen, Y.; Moussian, B.; Zhang, C.-X. A comprehensive omics analysis and functional survey of cuticular proteins in the brown planthopper. Proc. Natl. Acad. Sci. USA 2018, 115, 5175–5180. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Ye, Y.-X.; Zhang, C.-X. Pleiotropic Functions of FoxN1: Regulating Different Target Genes during Embryogenesis and Nymph Molting in the Brown Planthopper. Int. J. Mol. Sci. 2020, 21, 4222. [Google Scholar] [CrossRef]
- Merzendorfer, H. Insect chitin synthases: A review. J. Comp. Physiol. B 2006, 176, 1–15. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, H.-W.; Huang, H.; Xue, J.; Wu, W.-J.; Bao, Y.-Y.; Xu, H.-J.; Zhu, Z.-R.; Cheng, J.-A.; Zhang, C.-X. Chitin synthase 1 gene and its two alternative splicing variants from two sap-sucking insects, Nilaparvata lugens and Laodelphax striatellus (Hemiptera: Delphacidae). Insect Biochem. Mol. Biol. 2012, 42, 637–646. [Google Scholar] [CrossRef]
- Zinovyeva, M.V.; Kuzmich, A.I.; Monastyrskaya, G.S.; Sverdlov, E.D. The role of FOXA subfamily factors in embryonic development and carcinogenesis of the pancreas. Mol. Genet. Microbiol. Virol. 2016, 31, 135–142. [Google Scholar] [CrossRef]
- Kumamoto, T.; Hanashima, C. Evolutionary conservation and conversion of Foxg1 function in brain development. Dev. Growth Differ. 2017, 59, 258–269. [Google Scholar] [CrossRef] [Green Version]
- Grossniklaus, U.; Cadigan, K.M.; Gehring, W.J. Three maternal coordinate systems cooperate in the patterning of the Drosophila head. Development 1994, 120, 3155–3171. [Google Scholar] [CrossRef] [PubMed]
- Kitzmann, P.; Weißkopf, M.; Schacht, M.I.; Bucher, G. A key role for foxQ2 in anterior head and central brain patterning in insects. Development 2017, 144, 2969–2981. [Google Scholar] [PubMed] [Green Version]
- Posnien, N.; Koniszewski, N.D.B.; Hein, H.J.; Bucher, G. Candidate Gene Screen in the Red Flour Beetle Tribolium Reveals Six3 as Ancient Regulator of Anterior Median Head and Central Complex Development. PLoS Genet. 2011, 7, e1002416. [Google Scholar] [CrossRef]
- Dong, X.; Zhai, Y.; Zhang, J.; Sun, Z.; Chen, J.; Chen, J.; Zhang, W. Fork head transcription factor is required for ovarian mature in the brown planthopper, Nilaparvata lugens (Stål). BMC Mol. Biol. 2011, 12, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Wei, P.; Guo, H.; Wang, S.; Tang, B. Suppressing the expression of a forkhead transcription factor disrupts the chitin biosynthesis pathway in Spodoptera exigua. Arch. Insect Biochem. Physiol. 2014, 86, 4–18. [Google Scholar]
- Hong, H.-K.; Noveroske, J.K.; Headon, D.J.; Liu, T.; Sy, M.-S.; Justice, M.J.; Chakravarti, A. The winged helix/forkhead transcription factor Foxq1 regulates differentiation of hair in satin mice. Genesis 2001, 29, 163–171. [Google Scholar] [CrossRef]
- Potter, C.S.; Peterson, R.L.; Barth, J.L.; Pruett, N.D.; Jacobs, D.F.; Kern, M.J.; Argraves, W.S.; Sundberg, J.P.; Awgulewitsch, A. Evidence That the Satin Hair Mutant Gene Foxq1 Is among Multiple and Functionally Diverse Regulatory Targets for Hoxc13 during Hair Follicle Differentiation. J. Biol. Chem. 2006, 281, 29245–29255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, S.O.; Hojrup, P.; Roepstorff, P. Insect cuticular proteins. Insect Biochem. Mol. Biol. 1995, 25, 153–176. [Google Scholar] [CrossRef]
- Hamodrakas, S.J.; Willis, J.H.; Iconomidou, V.A. A structural model of the chitin-binding domain of cuticle proteins. Insect Biochem. Mol. Biol. 2002, 32, 1577–1583. [Google Scholar] [CrossRef]
- Ye, Y.-X.; Pan, P.-L.; Xu, J.-Y.; Shen, Z.-F.; Kang, D.; Lu, J.-B.; Hu, Q.-L.; Huang, H.; Lou, Y.-H.; Zhou, N.-M.; et al. Forkhead box transcription factor L2 activates Fcp3C to regulate insect chorion formation. Open Biol. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, H.-Y.; Zhu, C.-Q.; Zhang, H.-H.; Shen, Z.-C.; Zhang, C.-X.; Ye, Y.-X. The Genetic Network of Forkhead Gene Family in Development of Brown Planthoppers. Biology 2021, 10, 867. https://doi.org/10.3390/biology10090867
Lin H-Y, Zhu C-Q, Zhang H-H, Shen Z-C, Zhang C-X, Ye Y-X. The Genetic Network of Forkhead Gene Family in Development of Brown Planthoppers. Biology. 2021; 10(9):867. https://doi.org/10.3390/biology10090867
Chicago/Turabian StyleLin, Hai-Yan, Cheng-Qi Zhu, Hou-Hong Zhang, Zhi-Cheng Shen, Chuan-Xi Zhang, and Yu-Xuan Ye. 2021. "The Genetic Network of Forkhead Gene Family in Development of Brown Planthoppers" Biology 10, no. 9: 867. https://doi.org/10.3390/biology10090867
APA StyleLin, H.-Y., Zhu, C.-Q., Zhang, H.-H., Shen, Z.-C., Zhang, C.-X., & Ye, Y.-X. (2021). The Genetic Network of Forkhead Gene Family in Development of Brown Planthoppers. Biology, 10(9), 867. https://doi.org/10.3390/biology10090867