
Meta-Analysis of Mitochondrial DNA Control Region Diversity to Shed Light on Phylogenetic Relationship and Demographic History of African Sheep (Ovis aries) Breeds


Abstract
:Simple Summary
Abstract
1. Introduction
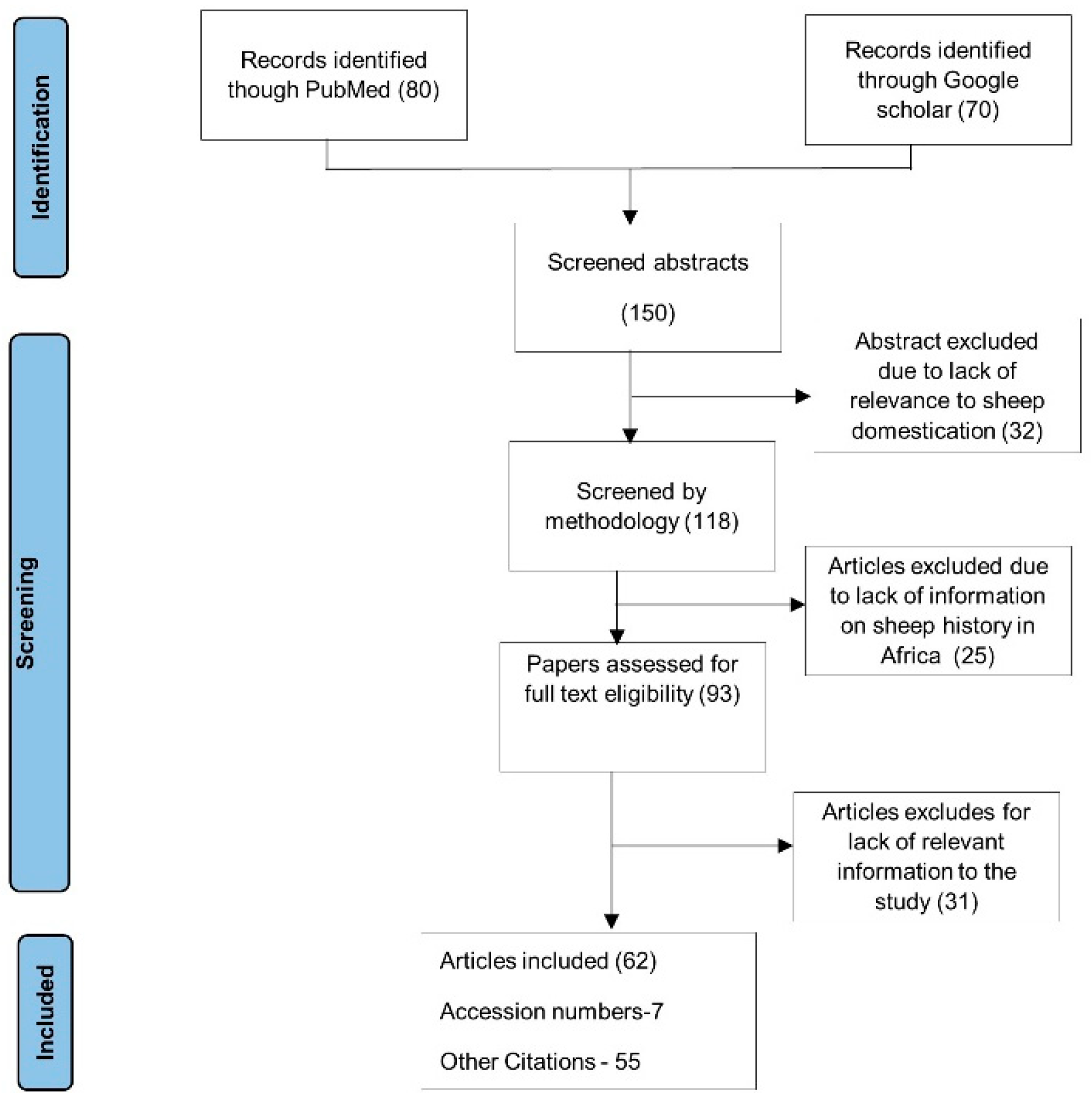
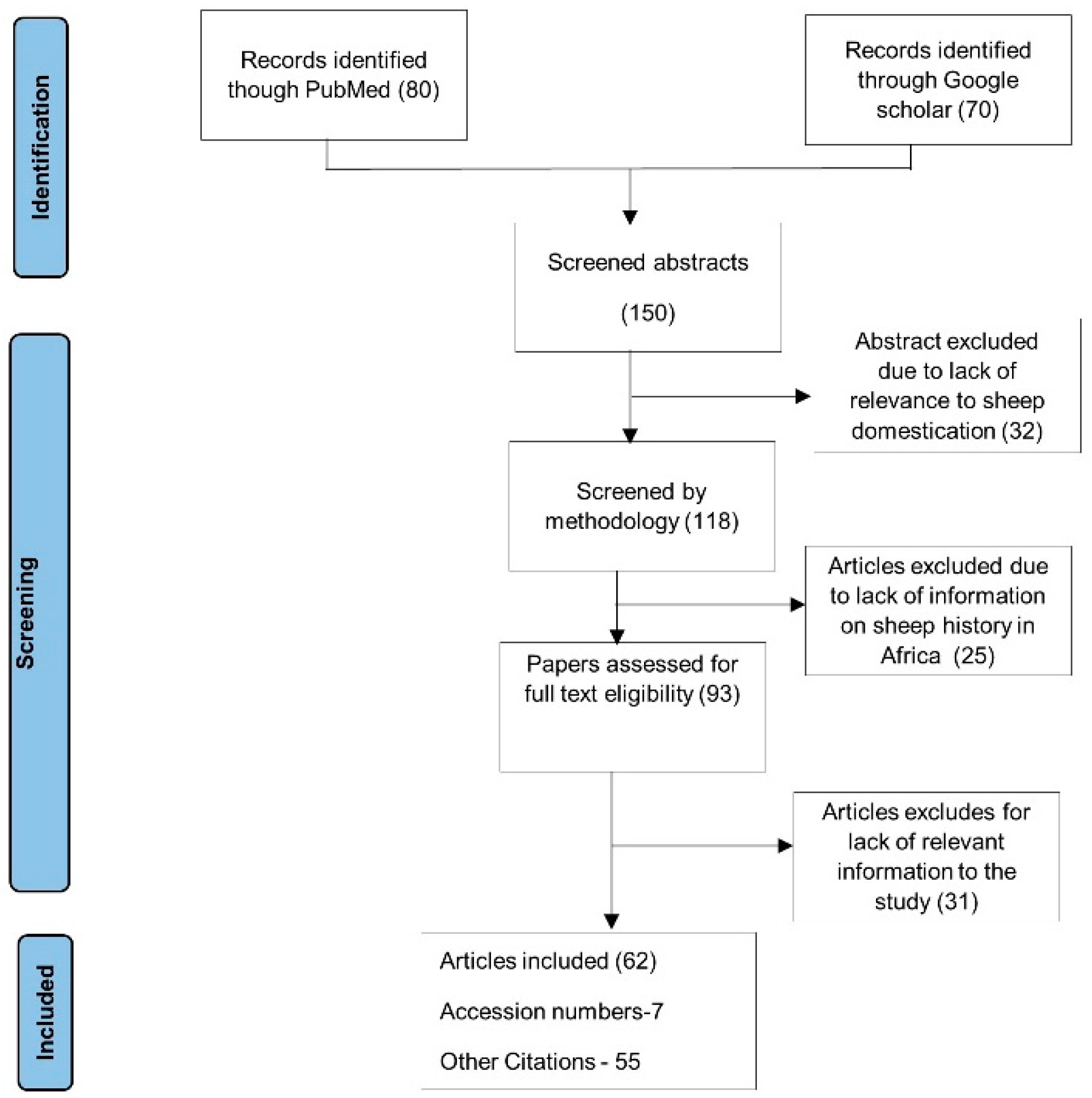
2. Methods
Data Source
3. Results
3.1. mtDNA CR Polymorphism Indices
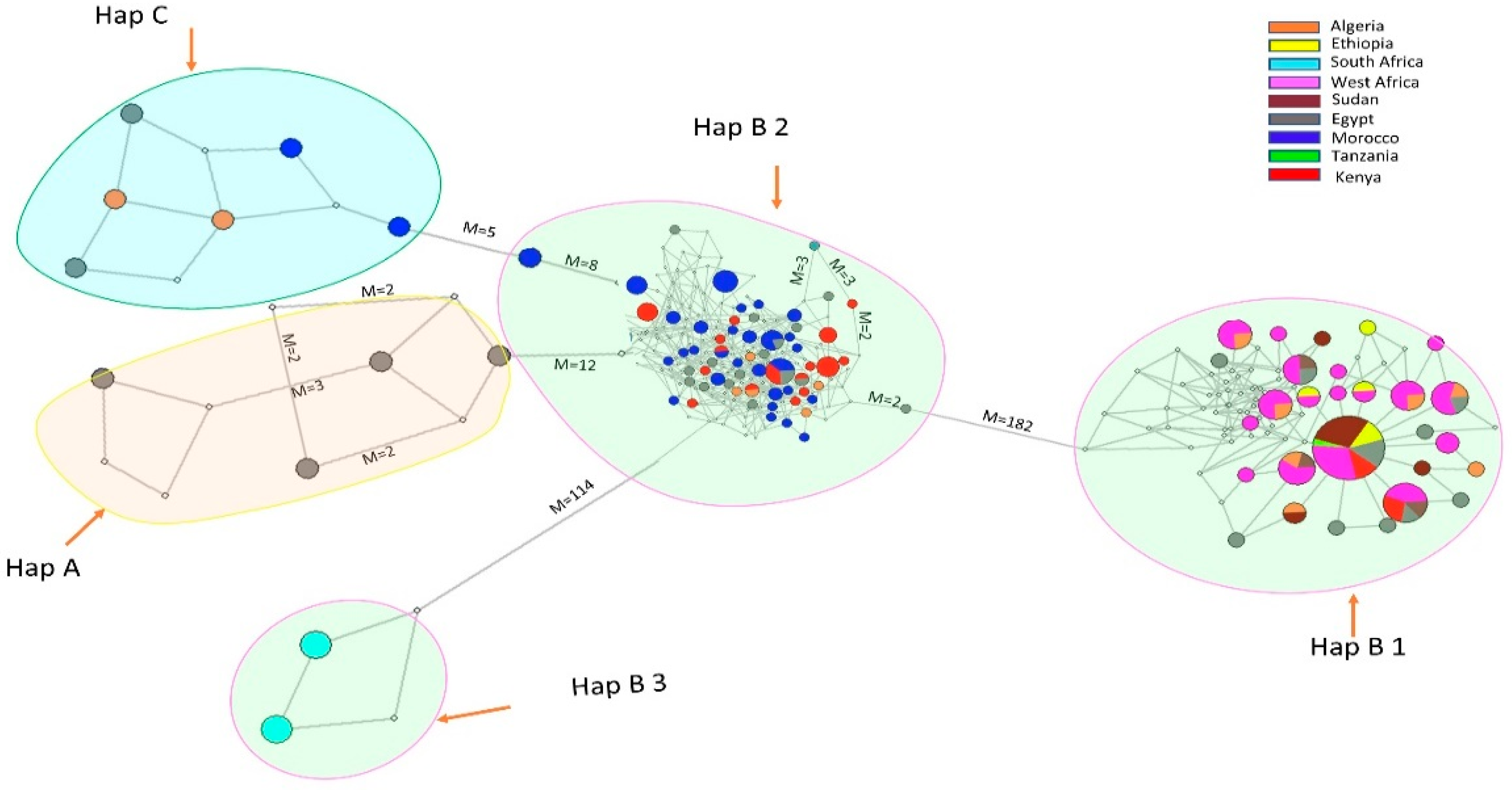
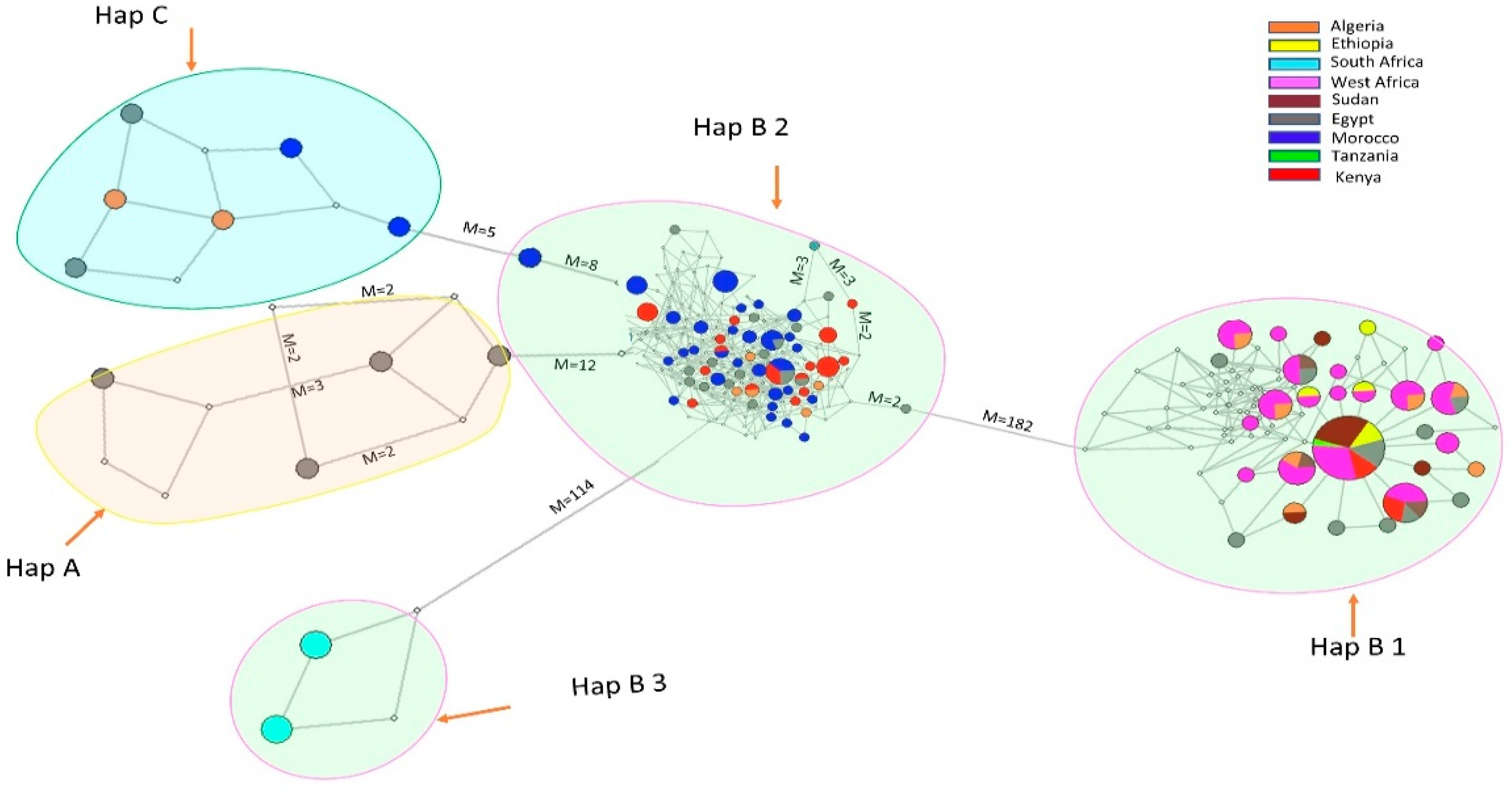
3.2. Phylogenetic Relationship
3.3. Neutrality Test
3.4. Population Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mollier, L.; Seyler, F.; Chotte, J.L.; Ringler, C. End hunger, achieve food security and improved nutrition and promote sustainable agriculture: SDG 2. In A Guide to SDG Interactions: From Science to Implementation; ICSU: Paris, France, 2017. [Google Scholar]
- FAO. Faostat-Production, “Livestock Primary”. Available online: http://www.fao.org/faostat/en/#data/QA (accessed on 5 May 2021).
- Molina, T.; Abadal, E. The Evolution of Communicating the Uncertainty of Climate Change to Policymakers: A Study of IPCC Synthesis Reports. Sustainability 2021, 13, 2466. [Google Scholar] [CrossRef]
- Resende, A.; Gonçalves, J.; Muigai, A.W.; Pereira, F. Mitochondrial DNA variation of domestic sheep (Ovis aries) in Kenya. Anim. Genet. 2016, 47, 377–381. [Google Scholar] [CrossRef]
- Othman, O.E.; Pariset, L.; Balabel, E.A.; Marioti, M. Genetic characterization of Egyptian and Italian sheep breeds using mitochondrial DNA. J. Genet. Eng. Biotechnol. 2015, 13, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Tapio, M.; Marzanov, N.; Ozerov, M.; Ćinkulov, M.; Gonzarenko, G.; Kiselyova, T.; Murawski, M.; Viinalass, H.; Kantanen, J. Sheep mitochondrial DNA variation in European, Caucasian, and Central Asian areas. Mol. Biol. Evol. 2006, 23, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Revelo, H.A.; López-Alvarez, D.; Landi, V.; Rizzo, L.; Alvarez, L.A. Mitochondrial DNA Variations in Colombian Creole Sheep Confirm an Iberian Origin and Shed Light on the Dynamics of Introduction Events of African Genotypes. Animals 2020, 10, 1594. [Google Scholar] [CrossRef]
- Ressaissi, Y.; Amills, M.; Noce, A.; Hamouda, M.B. Characterizing the Mitochondrial Diversity of Arbi Goats from Tunisia. Biochem. Genet. 2021, 1–8. [Google Scholar] [CrossRef]
- Al-Araimi, N.A.; Al-Atiyat, R.M.; Gaafar, O.M.; Vasconcelos, R.; Luzuriaga-Neira, A.; Eisa, M.O.; Amir, N.; Benaissa, M.H.; Alfaris, A.A.; Aljumaah, R.S.; et al. Maternal genetic diversity and phylogeography of native Arabian goats. Livest. Sci. 2017, 206, 88–94. [Google Scholar] [CrossRef]
- Ofori, S.A.; Hagan, J.K.; Kyei, F. Morphometric characterization and differentiation of West African Dwarf goat populations in Ghana. Trop. Anim. Health Prod. 2021, 53, 1–14. [Google Scholar]
- King, F.J.; Banga, C.B.; Visser, C. Genetic diversity and population structure of three native cattle populations in Mozambique. Trop. Anim. Health Prod. 2021, 53, 117. [Google Scholar] [CrossRef]
- Boettcher, P.J.; Tixier-Boichard, M.; Toro, M.A.; Simianer, H.; Eding, H.; Gandini, G.; Joost, S.; Garcia, D.; Colli, L.; Ajmone-Marsan, P.; et al. Objectives, criteria and methods for using molecular genetic data in priority setting for conservation of animal genetic resources. Anim. Genet. 2010, 41, 64–77. [Google Scholar] [CrossRef] [Green Version]
- Eydivandi, S.; Sahana, G.; Momen, M.; Moradi, M.H.; Schönherz, A.A. Genetic diversity in Iranian indigenous sheep vis-à-vis selected exogenous sheep breeds and wild mouflon. Anim. Genet. 2020, 51, 772–787. [Google Scholar] [CrossRef]
- Pedrosa, S.; Uzun, M.; Arranz, J.J.; Gutiérrez-Gil, B.; San Primitivo, F.; Bayón, Y. Evidence of three maternal lineages in Near Eastern sheep supporting multiple domestication events. Proc. R. Soc. B Biol. Sci. 2005, 272, 2211–2217. [Google Scholar] [CrossRef] [Green Version]
- Meadows, J.R.S.; Hanotte, O.; Drögemüller, C.; Calvo, J.; Godfrey, R.; Coltman, D.; Maddox, J.F.; Marzanov, N.; Kantanen, J.; Kijas, J.W. Globally dispersed Y chromosomal haplotypes in wild and domestic sheep. Anim. Genet. 2006, 37, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Muigai, A.W.; Hanotte, O. The origin of African sheep: Archaeological and genetic perspectives. Afr. Archaeol. Rev. 2013, 30, 39–50. [Google Scholar] [CrossRef]
- Ghernouti, N.; Bodinier, M.; Ranebi, D.; Maftah, A.; Petit, D.; Gaouar, S.B.S. Control Region of mtDNA identifies three migration events of sheep breeds in Algeria. Small Rumin. Res. 2017, 155, 66–71. [Google Scholar] [CrossRef]
- Nigussie, H.; Mwacharo, J.M.; Osama, S.; Agaba, M.; Mekasha, Y.; Kebede, K.; Abegaz, S.; Pal, S.K. Genetic diversity and matrilineal genetic origin of fat-rumped sheep in Ethiopia. Trop. Anim. Health Prod. 2019, 51, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Selepe, M.M. Genetic Variation among Zulu Sheep Sub-Populations of South Africa Assessed by Microsatellites and Mitochondrial DNA (mtDNA). Master’s Thesis, University of Zululand, Richards Bay, South Africa, 2018. [Google Scholar]
- Othman, O.E.; Balabel, E.A.; Abdel-Samad, M.F. Mitochondrial DNA diversity in five Egyptian sheep breeds. Glob. Vet. 2014, 12, 369–375. [Google Scholar]
- Kandoussi, A.; Boujenane, I.; Auger, C.; Serranito, B.; Germot, A.; Piro, M.; Maftah, A.; Badaoui, B.; Petit, D. The origin of sheep settlement in Western Mediterranean. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Nosrati, M.; Nanaei, H.A.; Javanmard, A.; Esmailizadeh, A. The pattern of runs of homozygosity and genomic inbreeding in world-wide sheep populations. Genomics 2021, 113, 1407–1415. [Google Scholar] [CrossRef]
- Machová, K.; Milerski, M.; Rychtářová, J.; Hofmanová, B.; Vostrá-Vydrová, H.; Moravčíková, N.; Kasarda, R.; Vostrý, L. Assessment of the genetic diversity of Two Czech autochthonous sheep breeds. Small Rumin. Res. 2021, 195, 106301. [Google Scholar] [CrossRef]
- Ocampo, R.J.; Martínez, J.F.; Martínez, R. Assessment of genetic diversity and population structure of Colombian Creole cattle using microsatellites. Trop. Anim. Health Prod. 2021, 53, 124. [Google Scholar] [CrossRef]
- Menezes, M.P.C.; Martinez, A.M.; Pimenta Filho, E.C.; Vega-Pla, J.L.; Delgado, J.V.; Arandas, J.K.G.; Rocha, L.L.D.; Ribeiro, M.N. Diversity Analysis and Genetic Relationships among Local Brazilian Goat Breeds Using SSR Markers. Animals 2020, 10, 1842. [Google Scholar] [CrossRef] [PubMed]
- Chokoe, T.C.; Mdladla-Hadebe, K.; Muchadeyi, F.; Dzomba, E.; Matelele, T.; Mphahlele, T.; Mpofu, T.J.; Nephawe, K.; Mtileni, B. Genetic Diversity of South African Indigenous Goat Population from Four Provinces Using Genome-Wide SNP Data. Sustainability 2020, 12, 10361. [Google Scholar] [CrossRef]
- Monau, P.I.; Visser, C.; Muchadeyi, F.C.; Okpeku, M.; Nsoso, S.J.; Van Marle-Köster, E. Population structure of indigenous southern African goats based on the Illumina Goat50K SNP panel. Trop. Anim. Health Prod. 2020, 52, 1795–1802. [Google Scholar] [CrossRef] [PubMed]
- Dotsev, A.V.; Kunz, E.; Kharzinova, V.R.; Okhlopkov, I.M.; Lv, F.H.; Li, M.H.; Rodionov, A.N.; Shakhin, A.V.; Sipko, T.P.; Medvedev, D.G.; et al. Mitochondrial DNA Analysis Clarifies Taxonomic Status of the Northernmost Snow Sheep (Ovis nivicola) Population. Life 2021, 11, 252. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ding, X.; Zeng, Y.; Yue, Y.; Guo, X.; Guo, T.; Chu, M.; Wang, F.; Han, J.; Feng, R.; et al. Genetic diversity and phylogenetic evolution of Tibetan sheep based on mtDNA D-Loop sequences. PLoS ONE 2016, 11, e0159308. [Google Scholar] [CrossRef]
- Murunga, P.; Kennedy, G.M.; Imboma, T.; Malaki, P.; Kariuki, D.; Ndiema, E.; Obanda, V.; Agwanda, B.; Lichoti, J.K.; Ommeh, S.C. Mitochondrial DNA D-Loop diversity of the helmeted guinea fowls in Kenya and its implications on HSP70 gene functional polymorphism. BioMed Res. Int. 2018, 2018, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, D.A.D.; Martínez, A.M.; Carolino, I.; Barros, M.C.; Vallejo, M.E.C.; Santos-Silva, F.; Almeida, M.J.D.O.; Carolino, N.; Bermejo, J.V.D.; Sarmento, J.L.R. Diversity and Genetic Relationship of Free-Range Chickens from the Northeast Region of Brazil. Animals 2020, 10, 1857. [Google Scholar] [CrossRef]
- Nguluma, A. Mitochondrial DNA D-Loop Sequence Variability Reveals High Haplotype Diversity and Multiple Maternal Origins in Twelve Indigenous Goat Populations from Tanzania. 2021. Available online: https://www.authorea.com/users/399326/articles/511867-mitochondrial-dna-d-loop-sequence-variability-reveals-high-haplotype-diversity-and-multiple-maternal-origins-in-twelve-indigenous-goat-populations-from-tanzania (accessed on 10 May 2021).
- Kosgey, I.S.; Rowlands, G.J.; van Arendonk, J.A.; Baker, R.L. Small ruminant production in smallholder and pastoral/extensive farming systems in Kenya. Small Rumin. Res. 2008, 77, 11–24. [Google Scholar] [CrossRef]
- Moradi, M.H.; Nejati-Javaremi, A.; Moradi-Shahrbabak, M.; Dodds, K.G.; McEwan, J.C. Genomic scan of selective sweeps in thin and fat tail sheep breeds for identifying of candidate regions associated with fat deposition. BMC Genet. 2012, 13, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rege, J.E.O.; Yapi-Gnoare, C.V.; Tawah, C.L. The Indigenous Domestic Ruminant Genetic Resource. 1996. Available online: https://agris.fao.org/agris-search/search.do?recordID=QT2016100541 (accessed on 10 May 2021).
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2012, 41, D36–D42. [Google Scholar] [CrossRef] [Green Version]
- Opoku, A. Biodiversity and the built environment: Implications for the Sustainable Development Goals (SDGs). Resour. Conserv. Recycl. 2019, 141, 1–7. [Google Scholar] [CrossRef]
- Page, M.J.; Moher, D.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. PRISMA 2020 explanation and elaboration: Updated guidance and exemplars for reporting systematic reviews. BMJ 2021, 372, 1–36. [Google Scholar] [CrossRef]
- Li, K.B. ClustalW-MPI: ClustalW analysis using distributed and parallel computing. Bioinformatics 2003, 19, 1585–1586. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Tajima, F. Evolutionary relationship of DNA sequences in finite populations. Genetics 1983, 105, 437–460. [Google Scholar] [CrossRef]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Onsins, S.E.; Rozas, J. Statistical properties of new neutrality tests against population growth. Mol. Biol. Evol. 2002, 19, 2092–2100. [Google Scholar] [CrossRef] [Green Version]
- Kusza, S.; Zakar, E.; Budai, C.; Cziszter, L.T.; Padeanu, I.; Gavojdian, D. Mitochondrial DNA variability in Gyimesi Racka and Turcana sheep breeds. Acta Biochim. Pol. 2015, 62, 273–280. [Google Scholar] [CrossRef]
- De Jong, M.A.; Wahlberg, N.; Van Eijk, M.; Brakefield, P.M.; Zwaan, B.J. Mitochondrial DNA signature for range-wide populations of Bicyclus anynana suggests a rapid expansion from recent refugia. PLoS ONE 2011, 6, e21385. [Google Scholar] [CrossRef] [Green Version]
- Mengoni, A.; Bazzicalupo, M. The statistical treatment of data and the analysis of molecular variance (AMOVA) in molecular microbial ecology. Ann. Microbiol. 2002, 52, 95–102. [Google Scholar]
- Bradley, D.G.; MacHugh, D.E.; Cunningham, P.; Loftus, R.T. Mitochondrial diversity and the origins of African and European cattle. Proc. Natl. Acad. Sci. USA 1996, 93, 5131–5135. [Google Scholar] [CrossRef] [Green Version]
- Pereira, F.; Queirós, S.; Gusmão, L.; Nijman, I.J.; Cuppen, E.; Lenstra, J.A.; Consortium, E.; Davis, S.J.; Nejmeddine, F.; Amorim, A. Tracing the history of goat pastoralism: New clues from mitochondrial and Y chromosome DNA in North Africa. Mol. Biol. Evol. 2009, 26, 2765–2773. [Google Scholar] [CrossRef] [Green Version]
- Agaviezor, B.O.; Adefenwa, M.A.; Peters, S.O.; Yakubu, A.; Adebambo, O.A.; Ozoje, M.O.; Ikeobi, C.O.N.; Ilori, B.M.; Wheto, M.; Ajayi, O.O.; et al. Genetic diversity analysis of the mitochondrial D-loop of Nigerian indigenous sheep. Anim. Genet. Res. 2012, 50, 13–20. [Google Scholar] [CrossRef]
- Mauki, D.H.; Adeola, A.C. Genetic variation of Nigerian cattle inferred from maternal and paternal genetic markers. PeerJ 2021, 9, e10607. [Google Scholar] [CrossRef]
- Hoban, S.; Bruford, M.; Jackson, J.D.U.; Lopes-Fernandes, M.; Heuertz, M.; Hohenlohe, P.A.; Paz-Vinas, I.; Sjögren-Gulve, P.; Segelbacher, G.; Vernesi, C.; et al. Genetic diversity targets and indicators in the CBD post-2020 Global Biodiversity Framework must be improved. Biol. Conserv. 2020, 248, 108654. [Google Scholar] [CrossRef]
- Kijas, J.W.; Lenstra, J.A.; Hayes, B.; Boitard, S.; Neto, L.R.P.; San Cristobal, M.; Servin, B.; McCulloch, R.; Whan, V.; Gietzen, K.; et al. Genome-wide analysis of the world’s sheep breeds reveals high levels of historic mixture and strong recent selection. PLoS Biol. 2012, 10, e1001258. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, I.; Capote, J.; Traoré, A.; Fonseca, N.; Pérez, K.; Cuervo, M.; Fernandez, I.; Goyache, F. Mitochondrial analysis sheds light on the origin of hair sheep. Anim. Genet. 2013, 44, 344–347. [Google Scholar] [CrossRef]
- Tarekegn, G.M.; Tesfaye, K.; Mwai, O.A.; Djikeng, A.; Dessie, T.; Birungi, J.; Osama, S.; Zergaw, N.; Alemu, A.; Achieng, G.; et al. Mitochondrial DNA variation reveals maternal origins and demographic dynamics of Ethiopian indigenous goats. Ecol. Evol. 2018, 8, 1543–1553. [Google Scholar] [CrossRef] [PubMed]
- Horsburgh, K.A.; Rhines, A. Genetic characterization of an archaeological sheep assemblage from South Africa’s Western Cape. J. Archaeol. Sci. 2010, 37, 2906–2910. [Google Scholar] [CrossRef]
- Kijas, J.W.; Townley, D.; Dalrymple, B.P.; Heaton, M.P.; Maddox, J.F.; McGrath, A.; Wilson, P.; Ingersoll, R.G.; McCulloch, R.; McWilliam, S.; et al. A genome wide survey of SNP variation reveals the genetic structure of sheep breeds. PLoS ONE 2009, 4, e4668. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.; Huang, G.; Wang, Z.; Sun, J.; Wu, Z.; Chen, N.; Lei, C.; Hanif, Q. Mitogenome diversity and maternal origins of Guangxi cattle breeds. Animals 2020, 10, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenstra, J.A.; Ajmone-Marsan, P.; Beja-Pereira, A.; Bollongino, R.; Bradley, D.G.; Colli, L.; De Gaetano, A.; Edwards, C.J.; Felius, M.; Ferretti, L.; et al. Meta-analysis of mitochondrial DNA reveals several population bottlenecks during worldwide migrations of cattle. Diversity 2014, 6, 178–187. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| N | P | M | Pi | K | H | HD | G + C | |
|---|---|---|---|---|---|---|---|---|
| 1 | 244 | 278 | 351 | 0.131 ± 0.017 | 47.436 | 205 | 0.998 ± 0.001 | 0.304 |
| 2 | 44 | 22 | 23 | 0.008 ± 0.001 | 3.615 | 29 | 0.977 ± 0.010 | 0.418 |
| 3 | 101 | 290 | 320 | 0.263 ± 0.015 | 116.994 | 71 | 0.977 ± 0.008 | 0.337 |
| 4 | 10 | 12 | 12 | 0.009 ± 0.002 | 3.578 | 8 | 0.956 ± 0.059 | 0.405 |
| Total | 399 | 284 | 401 | 0.254 ± 0.012 | 84.170 | 272 | 0.993 ± 0.002 | 0.321 |
| Test | Statistics | 1 | 2 | 3 | 4 | Mean | S.D. |
|---|---|---|---|---|---|---|---|
| Tajima’s D test | Tajima’s D | 0.029 | −0.921 | 3.715 | −0.707 | 0.529 | 2.163 |
| p-value | 0.621 | 0.197 | 0.998 | 0.262 | 0.515 | 0.368 | |
| Fu’s FS test | FS | −24.003 | −10.426 | 5.659 | −1.303 | −7.518 | 12.812 |
| p-value | 0.000 | 0.000 | 0.938 | 0.217 | 0.229 | 0.465 |
| Scheme | D.F. | Sum Squares | Variance Components | Percentage Variation |
|---|---|---|---|---|
| Among populations | 3 | 9923.905 | 44.785 | 54.696 |
| Within populations | 395 | 14,652.210 | 37.095 | 45.304 |
| Total | 398 | 24,576.110 | 81.880 |
| Source of Variations | Fixation | Index | FST | Percentage of Variations |
|---|---|---|---|---|
| Among populations | 3 | 3.195 | 0.008 | 1.570 |
| Within populations | 395 | 194.402 | 0.492 | 98.430 |
| Total | 398 | 197.596 | 0.500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wanjala, G.; Bagi, Z.; Kusza, S. Meta-Analysis of Mitochondrial DNA Control Region Diversity to Shed Light on Phylogenetic Relationship and Demographic History of African Sheep (Ovis aries) Breeds. Biology 2021, 10, 762. https://doi.org/10.3390/biology10080762
Wanjala G, Bagi Z, Kusza S. Meta-Analysis of Mitochondrial DNA Control Region Diversity to Shed Light on Phylogenetic Relationship and Demographic History of African Sheep (Ovis aries) Breeds. Biology. 2021; 10(8):762. https://doi.org/10.3390/biology10080762
Chicago/Turabian StyleWanjala, George, Zoltán Bagi, and Szilvia Kusza. 2021. "Meta-Analysis of Mitochondrial DNA Control Region Diversity to Shed Light on Phylogenetic Relationship and Demographic History of African Sheep (Ovis aries) Breeds" Biology 10, no. 8: 762. https://doi.org/10.3390/biology10080762
APA StyleWanjala, G., Bagi, Z., & Kusza, S. (2021). Meta-Analysis of Mitochondrial DNA Control Region Diversity to Shed Light on Phylogenetic Relationship and Demographic History of African Sheep (Ovis aries) Breeds. Biology, 10(8), 762. https://doi.org/10.3390/biology10080762