
The Effects of Temperature and Pressure on Protein-Ligand Binding in the Presence of Mars-Relevant Salts †

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Samples Preparation
2.3. Steady-State Fluorescence Spectroscopy
2.4. Circular Dichroism Spectroscopy
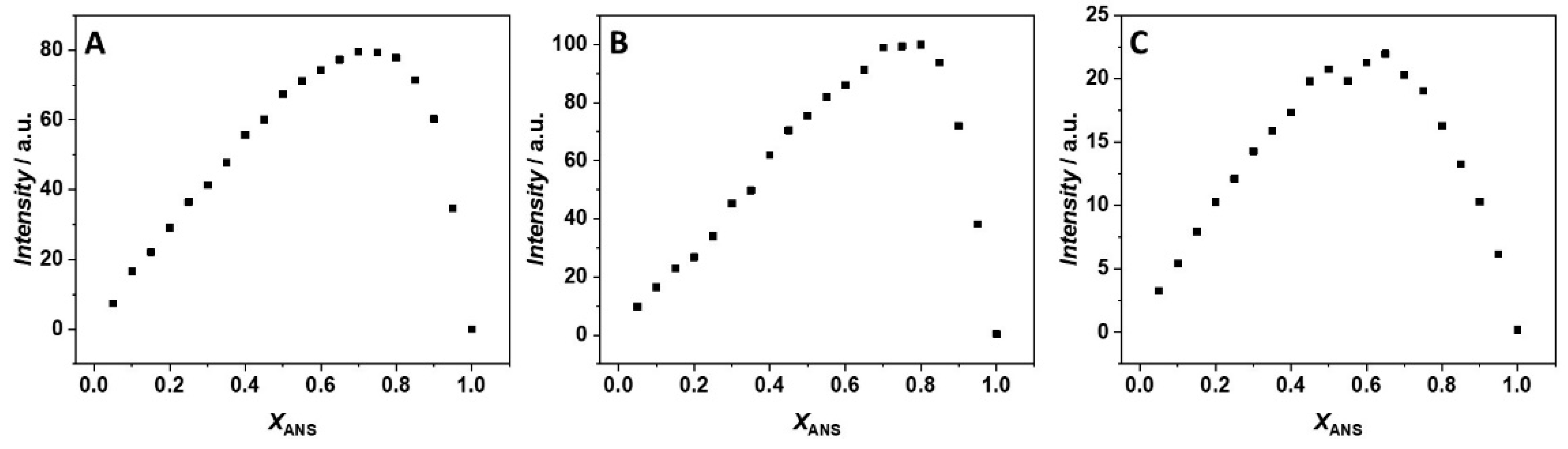
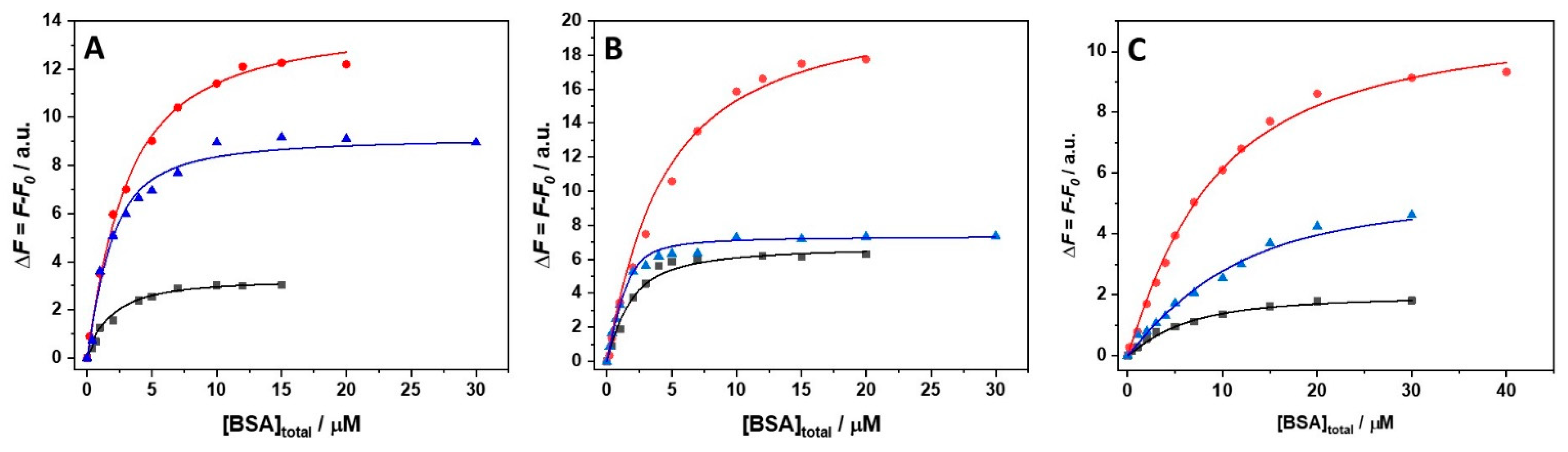
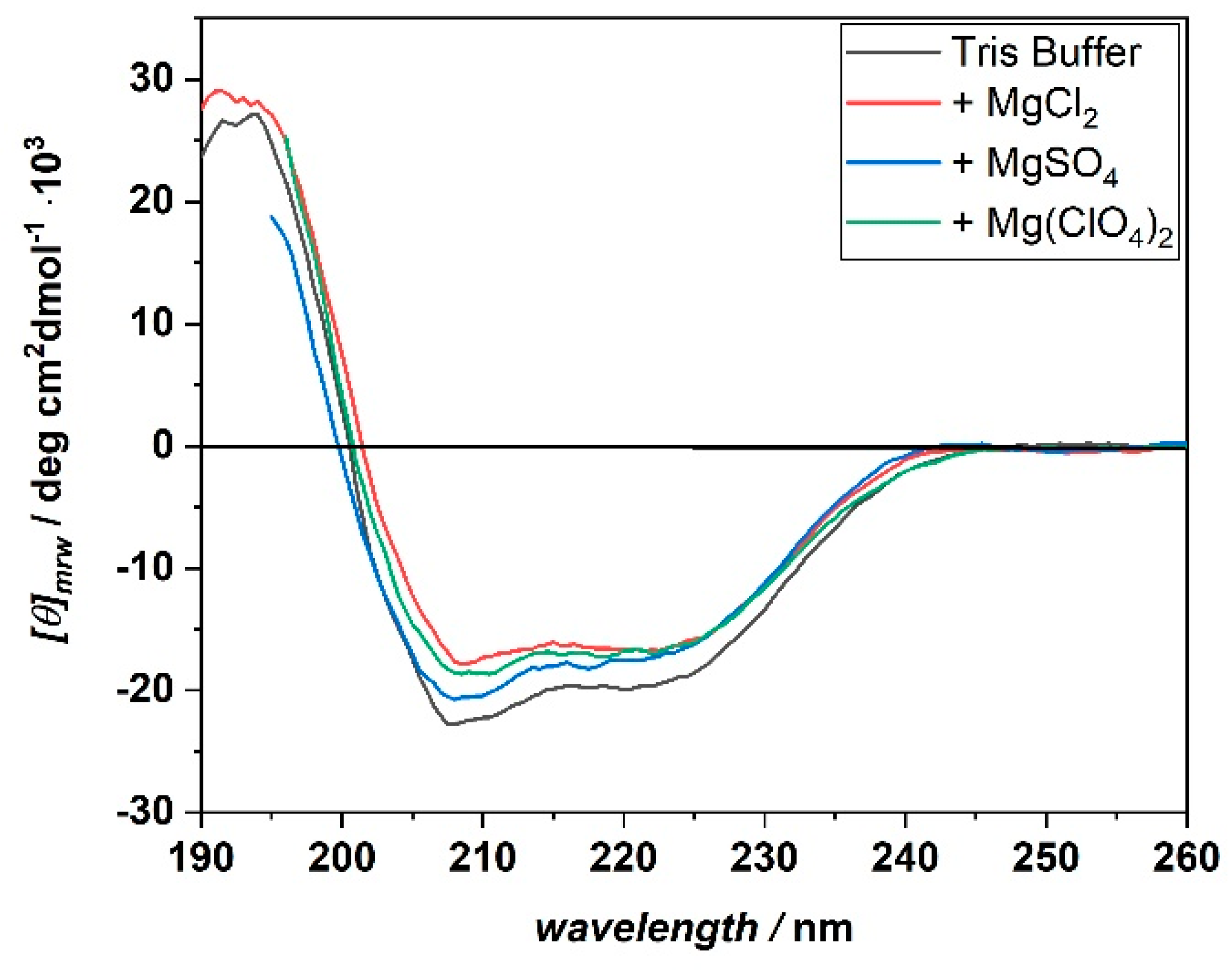
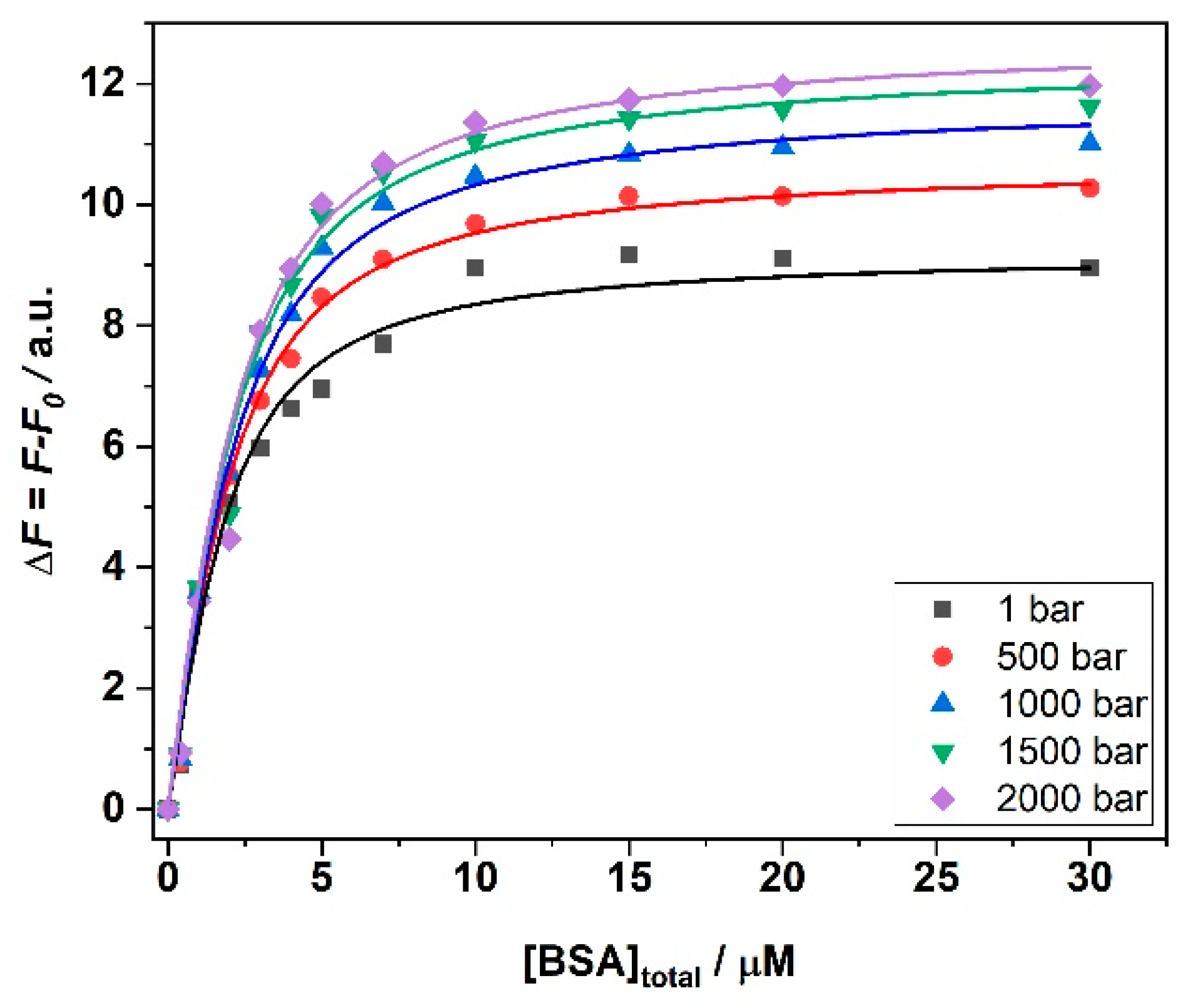
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cera, E.D. Thermodynamic Theory of Site-Specific Binding Processes in Biological Macromolecules, 1st ed.; Cambridge University Press: Cambridge, UK, 1995; ISBN 978-0-521-41659-7. [Google Scholar]
- Protein-Ligand Interactions; Gohlke, H. (Eds.) Methods and Principles in Medicinal Chemistry; Wiley-VCH: Weinheim, Germany, 2012; ISBN 978-3-527-32966-3. [Google Scholar]
- Pang, X.; Zhou, H.-X. Rate Constants and Mechanisms of Protein–Ligand Binding. Annu. Rev. Biophys. 2017, 46, 105–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodbury, C.P., Jr. Introduction to Macromolecular Binding Equilibria; CRC Press: Boca Raton, FL, USA, 2019; ISBN 978-0-367-38832-4. [Google Scholar]
- Du, X.; Li, Y.; Xia, Y.-L.; Ai, S.-M.; Liang, J.; Sang, P.; Ji, X.-L.; Liu, S.-Q. Insights into Protein–Ligand Interactions: Mechanisms, Models, and Methods. Int. J. Mol. Sci. 2016, 17, 144. [Google Scholar] [CrossRef]
- Akasaka, K. (Ed.) High Pressure Bioscience: Basic Concepts, Applications and Frontiers; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2015; ISBN 978-94-017-9917-1. [Google Scholar]
- Daniel, I.; Oger, P.; Winter, R. Origins of Life and Biochemistry under High-Pressure Conditions. Chem. Soc. Rev. 2006, 35, 858. [Google Scholar] [CrossRef] [PubMed]
- Meersman, F.; Daniel, I.; Bartlett, D.H.; Winter, R.; Hazael, R.; McMillan, P.F. High-Pressure Biochemistry and Biophysics. Rev. Miner. Geochem. 2013, 75, 607–648. [Google Scholar] [CrossRef] [Green Version]
- Roche, J.; Caro, J.A.; Norberto, D.R.; Barthe, P.; Roumestand, C.; Schlessman, J.L.; Garcia, A.E.; Garcia-Moreno, E.B.; Royer, C.A. Cavities Determine the Pressure Unfolding of Proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 6945–6950. [Google Scholar] [CrossRef] [Green Version]
- Winter, R. Interrogating the Structural Dynamics and Energetics of Biomolecular Systems with Pressure Modulation. Annu. Rev. Biophys. 2019, 48, 441–463. [Google Scholar] [CrossRef]
- Osterloo, M.M.; Hamilton, V.E.; Bandfield, J.L.; Glotch, T.D.; Baldridge, A.M.; Christensen, P.R.; Tornabene, L.L.; Anderson, F.S. Chloride-Bearing Materials in the Southern Highlands of Mars. Science 2008, 319, 1651–1654. [Google Scholar] [CrossRef] [Green Version]
- Gendrin, A. Sulfates in Martian Layered Terrains: The OMEGA/Mars Express View. Science 2005, 307, 1587–1591. [Google Scholar] [CrossRef] [Green Version]
- Hecht, M.H.; Kounaves, S.P.; Quinn, R.C.; West, S.J.; Young, S.M.M.; Ming, D.W.; Catling, D.C.; Clark, B.C.; Boynton, W.V.; Hoffman, J.; et al. Detection of Perchlorate and the Soluble Chemistry of Martian Soil at the Phoenix Lander Site. Science 2009, 325, 64–67. [Google Scholar] [CrossRef] [Green Version]
- Clifford, S.M.; Lasue, J.; Heggy, E.; Boisson, J.; McGovern, P.; Max, M.D. Depth of the Martian Cryosphere: Revised Estimates and Implications for the Existence and Detection of Subpermafrost Groundwater. J. Geophys. Res. 2010, 115, E07001. [Google Scholar] [CrossRef]
- Orosei, R.; Lauro, S.E.; Pettinelli, E.; Cicchetti, A.; Coradini, M.; Cosciotti, B.; Di Paolo, F.; Flamini, E.; Mattei, E.; Pajola, M.; et al. Radar Evidence of Subglacial Liquid Water on Mars. Science 2018, 361, 490–493. [Google Scholar] [CrossRef] [Green Version]
- Lauro, S.E.; Pettinelli, E.; Caprarelli, G.; Guallini, L.; Rossi, A.P.; Mattei, E.; Cosciotti, B.; Cicchetti, A.; Soldovieri, F.; Cartacci, M.; et al. Multiple Subglacial Water Bodies below the South Pole of Mars Unveiled by New MARSIS Data. Nat. Astron. 2021, 5, 63–70. [Google Scholar] [CrossRef]
- Oliva, R.; Banerjee, S.; Cinar, H.; Ehrt, C.; Winter, R. Alteration of Protein Binding Affinities by Aqueous Two-Phase Systems Revealed by Pressure Perturbation. Sci. Rep. 2020, 10, 8074. [Google Scholar] [CrossRef]
- Connors, K.A. Binding Constants: The Measurement of Molecular Complex Stability; Wiley: New York, NY, USA, 1987; ISBN 978-0-471-83083-2. [Google Scholar]
- Ulatowski, F.; Dąbrowa, K.; Bałakier, T.; Jurczak, J. Recognizing the Limited Applicability of Job Plots in Studying Host–Guest Interactions in Supramolecular Chemistry. J. Org. Chem. 2016, 81, 1746–1756. [Google Scholar] [CrossRef]
- Oliva, R.; Mukherjee, S.; Winter, R. Unraveling the Binding Characteristics of Small Ligands to Telomeric DNA by Pressure Modulation. Sci. Rep. 2021, 11, 9714. [Google Scholar] [CrossRef]
- Cattoni, D.I.; Kaufman, S.B.; Flecha, F.L.G. Kinetics and Thermodynamics of the Interaction of 1-Anilino-Naphthalene-8-Sulfonate with Proteins. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2009, 1794, 1700–1708. [Google Scholar] [CrossRef]
- Kelly, S.M.; Jess, T.J.; Price, N.C. How to Study Proteins by Circular Dichroism. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2005, 1751, 119–139. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, E.; Oliva, R.; Morra, R.; Bosso, A.; Ragucci, S.; Petraccone, L.; Del Vecchio, P.; Di Maro, A. Binding of a Type 1 RIP and of Its Chimeric Variant to Phospholipid Bilayers: Evidence for a Link between Cytotoxicity and Protein/Membrane Interactions. Biochim. Biophys. Acta (BBA) Biomembr. 2017, 1859, 2106–2112. [Google Scholar] [CrossRef] [PubMed]
- Majorek, K.A.; Porebski, P.J.; Dayal, A.; Zimmerman, M.D.; Jablonska, K.; Stewart, A.J.; Chruszcz, M.; Minor, W. Structural and Immunologic Characterization of Bovine, Horse, and Rabbit Serum Albumins. Mol. Immunol. 2012, 52, 174–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayakawa, I.; Kajihara, J.; Morikawa, K.; Oda, M.; Fujio, Y. Denaturation of Bovine Serum Albumin (BSA) and Ovalbumin by High Pressure, Heat and Chemicals. J. Food Sci. 1992, 57, 288–292. [Google Scholar] [CrossRef]
- Masson, P.; Reybaud, J. Hydrophobic Interaction Electrophoresis under High Hydrostatic Pressure: Study of the Effects of Pressure upon the Interaction of Serum Albumin with a Long-Chain Aliphatic Ligand. Electrophoresis 1988, 9, 157–161. [Google Scholar] [CrossRef]
- Aswal, V.K.; Chodankar, S.; Kohlbrecher, J.; Vavrin, R.; Wagh, A.G. Small-Angle Neutron Scattering Study of Protein Unfolding and Refolding. Phys. Rev. E 2009, 80, 011924. [Google Scholar] [CrossRef] [Green Version]
- Oliva, R.; Jahmidi-Azizi, N.; Mukherjee, S.; Winter, R. Harnessing Pressure Modulation for Exploring Ligand Binding Reactions in Cosolvent Solutions. J. Phys. Chem. B 2021, 125, 539–546. [Google Scholar] [CrossRef]
- Bye, J.W.; Falconer, R.J. Thermal Stability of Lysozyme as a Function of Ion Concentration: A Reappraisal of the Relationship between the Hofmeister Series and Protein Stability: Thermal Stability of Lysozyme. Protein Sci. 2013, 22, 1563–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medda, L.; Monduzzi, M.; Salis, A. The Molecular Motion of Bovine Serum Albumin under Physiological Conditions is Ion Specific. Chem. Commun. 2015, 51, 6663–6666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levi, V.; González Flecha, F.L. Labeling of Proteins with Fluorescent Probes: Photophysical Characterization of Dansylated Bovine Serum Albumin. Biochem. Mol. Biol. Educ. 2003, 31, 333–336. [Google Scholar] [CrossRef]
- Fox, J.M.; Kang, K.; Sherman, W.; Héroux, A.; Sastry, G.M.; Baghbanzadeh, M.; Lockett, M.R.; Whitesides, G.M. Interactions between Hofmeister Anions and the Binding Pocket of a Protein. J. Am. Chem. Soc. 2015, 137, 3859–3866. [Google Scholar] [CrossRef] [PubMed]
- Ory, J.J.; Banaszak, L.J. Studies of the Ligand Binding Reaction of Adipocyte Lipid Binding Protein Using the Fluorescent Probe 1,8-Anilinonaphthalene-8-Sulfonate. Biophys. J. 1999, 77, 1107–1116. [Google Scholar] [CrossRef] [Green Version]
- Schonbrunn, E.; Eschenburg, S.; Luger, K.; Kabsch, W.; Amrhein, N. Structural Basis for the Interaction of the Fluorescence Probe 8-Anilino-1-Naphthalene Sulfonate (ANS) with the Antibiotic Target MurA. Proc. Natl. Acad. Sci. USA 2000, 97, 6345–6349. [Google Scholar] [CrossRef] [Green Version]
- Matulis, D.; Lovrien, R. 1-Anilino-8-Naphthalene Sulfonate Anion-Protein Binding Depends Primarily on Ion Pair Formation. Biophys. J. 1998, 74, 422–429. [Google Scholar] [CrossRef] [Green Version]
- Zhukov, A.; Karlsson, R. Statistical Aspects of van’t Hoff Analysis: A Simulation Study. J. Mol. Recognit. 2007, 20, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Chalikian, T.V.; Macgregor, R.B. On Empirical Decomposition of Volumetric Data. Biophys. Chem. 2019, 246, 8–15. [Google Scholar] [CrossRef]
- Mozhaev, V.V.; Heremans, K.; Frank, J.; Masson, P.; Balny, C. High Pressure Effects on Protein Structure and Function. Proteins 1996, 24, 81–91. [Google Scholar] [CrossRef]
- Boonyaratanakornkit, B.B.; Park, C.B.; Clark, D.S. Pressure Effects on Intra- and Intermolecular Interactions within Proteins. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 2002, 1595, 235–249. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
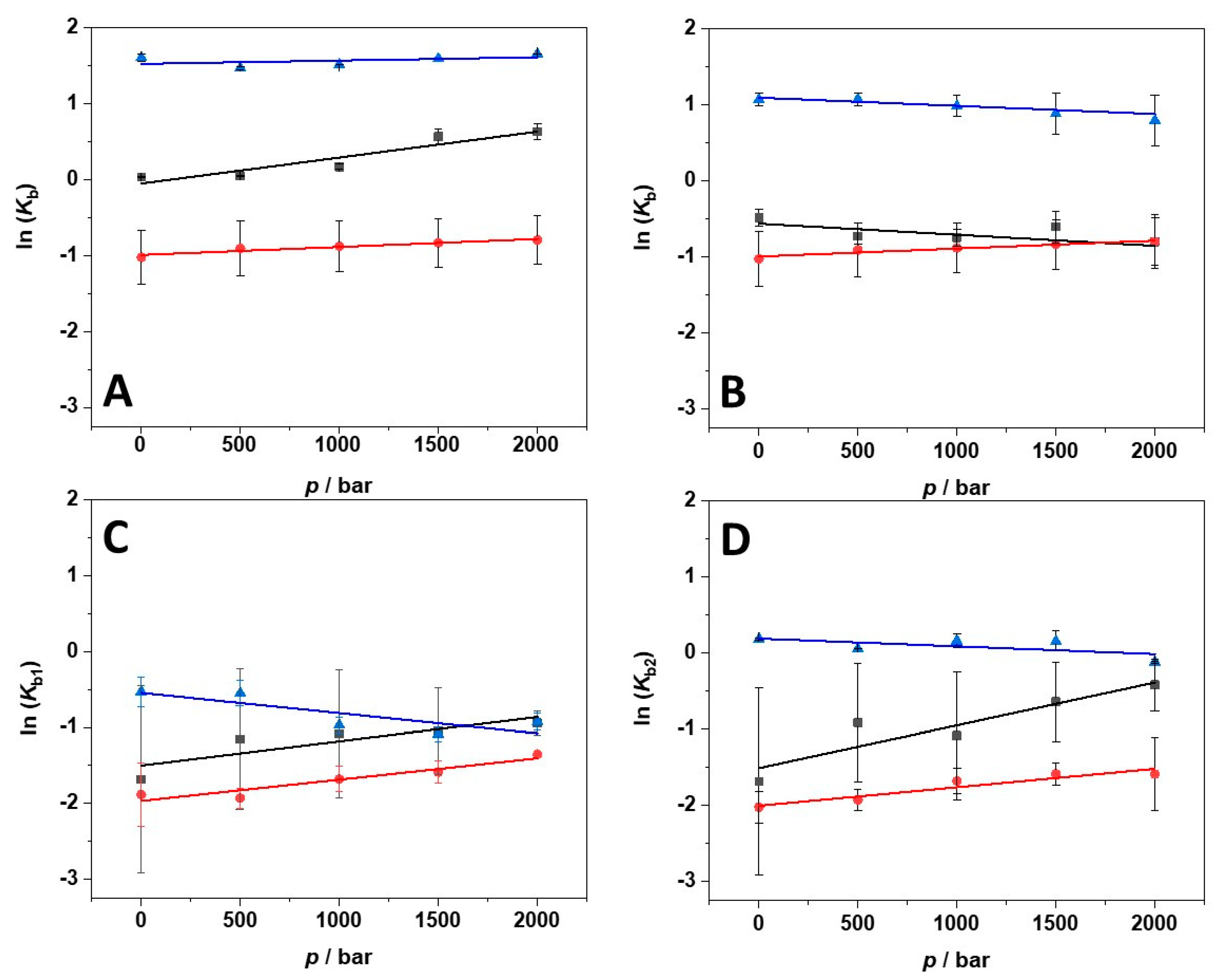
| Solution Conditions | T/°C | Kb1/M−1 × 106 | Kb2/M−1 × 106 | 1n |
|---|---|---|---|---|
| 250 mM MgCl2 | 5 | 1.0 ± 0.1 | - | 1:3 |
| 15 | 0.36 ± 0.13 | - | 1:3 | |
| 25 | 4.9 ± 0.2 | - | 1:3 | |
| 250 mM MgSO4 | 5 | 0.62 ± 0.15 | - | 1:3 |
| 15 | 0.39 ± 0.14 | - | 1:3 | |
| 25 | 2.9 ± 0.3 | - | 1:3 | |
| 250 mM Mg(ClO4)2 | 5 | 0.19± 0.13 | 0.19 ± 0.13 | 1:2 |
| 15 | 0.15 ± 0.04 | 0.13 ± 0.02 | 1:2 | |
| 25 | 0.59 ± 0.22 | 1.2 ± 0.1 | 1:2 |
| Solution Conditions | T/°C | p/bar | Kb1/M−1 ∙106 | Kb2/M−1 ∙106 | 1n |
|---|---|---|---|---|---|
| 250 mM MgCl2 | 5 | 1 | 1.0 ± 0.1 | - | 1:3 |
| 5 | 500 | 1.0 ± 0.2 | - | 1:3 | |
| 5 | 1000 | 1.2 ± 0.3 | - | 1:3 | |
| 5 | 1500 | 1.8 ± 0.3 | - | 1:3 | |
| 5 | 2000 | 1.9 ± 0.3 | - | 1:3 | |
| 250 mM MgCl2 | 15 | 1 | 0.36 ± 0.13 | - | 1:3 |
| 15 | 500 | 0.40 ± 0.16 | - | 1:3 | |
| 15 | 1000 | 0.41 ± 0.16 | - | 1:3 | |
| 15 | 1500 | 0.43 ± 0.17 | - | 1:3 | |
| 15 | 2000 | 0.45 ± 0.18 | - | 1:3 | |
| 250 mM MgCl2 | 25 | 1 | 4.9 ± 0.2 | - | 1:3 |
| 25 | 500 | 4.3 ± 0.1 | - | 1:3 | |
| 25 | 1000 | 4.5 ± 0.1 | - | 1:3 | |
| 25 | 1500 | 4.9 ± 0.2 | - | 1:3 | |
| 25 | 2000 | 5.2 ± 0.1 | - | 1:3 | |
| 5 | 1 | 0.62 ± 0.15 | - | 1:3 | |
| 5 | 500 | 0.48 ± 0.07 | - | 1:3 | |
| 250 mM MgSO4 | 5 | 1000 | 0.47 ± 0.07 | - | 1:3 |
| 5 | 1500 | 0.55 ± 0.20 | - | 1:3 | |
| 5 | 2000 | 0.44 ± 0.20 | - | 1:3 | |
| 15 | 1 | 0.39 ± 0.14 | - | 1:3 | |
| 15 | 500 | 0.40 ± 0.16 | - | 1:3 | |
| 250 mM MgSO4 | 15 | 1000 | 0.42 ± 0.15 | - | 1:3 |
| 15 | 1500 | 0.43 ± 0.17 | - | 1:3 | |
| 15 | 2000 | 0.45 ± 0.18 | - | 1:3 | |
| 25 | 1 | 2.9 ± 0.3 | - | 1:3 | |
| 25 | 500 | 2.9 ± 0.3 | - | 1:3 | |
| 250 mM MgSO4 | 25 | 1000 | 2.7 ± 0.4 | - | 1:3 |
| 25 | 1500 | 2.4 ± 0.7 | - | 1:3 | |
| 25 | 2000 | 2.2 ± 0.9 | - | 1:3 | |
| 5 | 1 | 0.19± 0.13 | 0.19 ± 0.13 | 1:2 | |
| 5 | 500 | 0.32 ± 0.25 | 0.40 ± 0.33 | 1:2 | |
| 250 mM Mg(ClO4)2 | 5 | 1000 | 0.34 ± 0.26 | 0.34 ± 0.26 | 1:2 |
| 5 | 1500 | 0.35 ± 0.19 | 0.52 ± 0.43 | 1:2 | |
| 5 | 2000 | 0.39 ± 0.07 | 0.66 ± 0.53 | 1:2 | |
| 15 | 1 | 0.15± 0.04 | 0.13 ± 0.13 | 1:2 | |
| 15 | 500 | 0.14 ± 0.01 | 0.15 ± 0.10 | 1:2 | |
| 250 mM Mg(ClO4)2 | 15 | 1000 | 0.19 ± 0.02 | 0.19 ± 0.18 | 1:2 |
| 15 | 1500 | 0.21 ± 0.02 | 0.21 ± 0.21 | 1:2 | |
| 15 | 2000 | 0.26 ± 0.01 | 0.24 ± 0.20 | 1:2 | |
| 25 | 1 | 0.59 ± 0.22 | 1.2 ± 0.1 | 1:2 | |
| 25 | 500 | 0.59 ± 0.17 | 1.1 ± 0.1 | 1:2 | |
| 250 mM Mg(ClO4)2 | 25 | 1000 | 0.38 ± 0.10 | 1.2 ± 0.2 | 1:2 |
| 25 | 1500 | 0.34 ± 0.03 | 1.2 ± 0.3 | 1:2 | |
| 25 | 2000 | 0.40 ± 0.05 | 0.89 ± 0.05 | 1:2 |
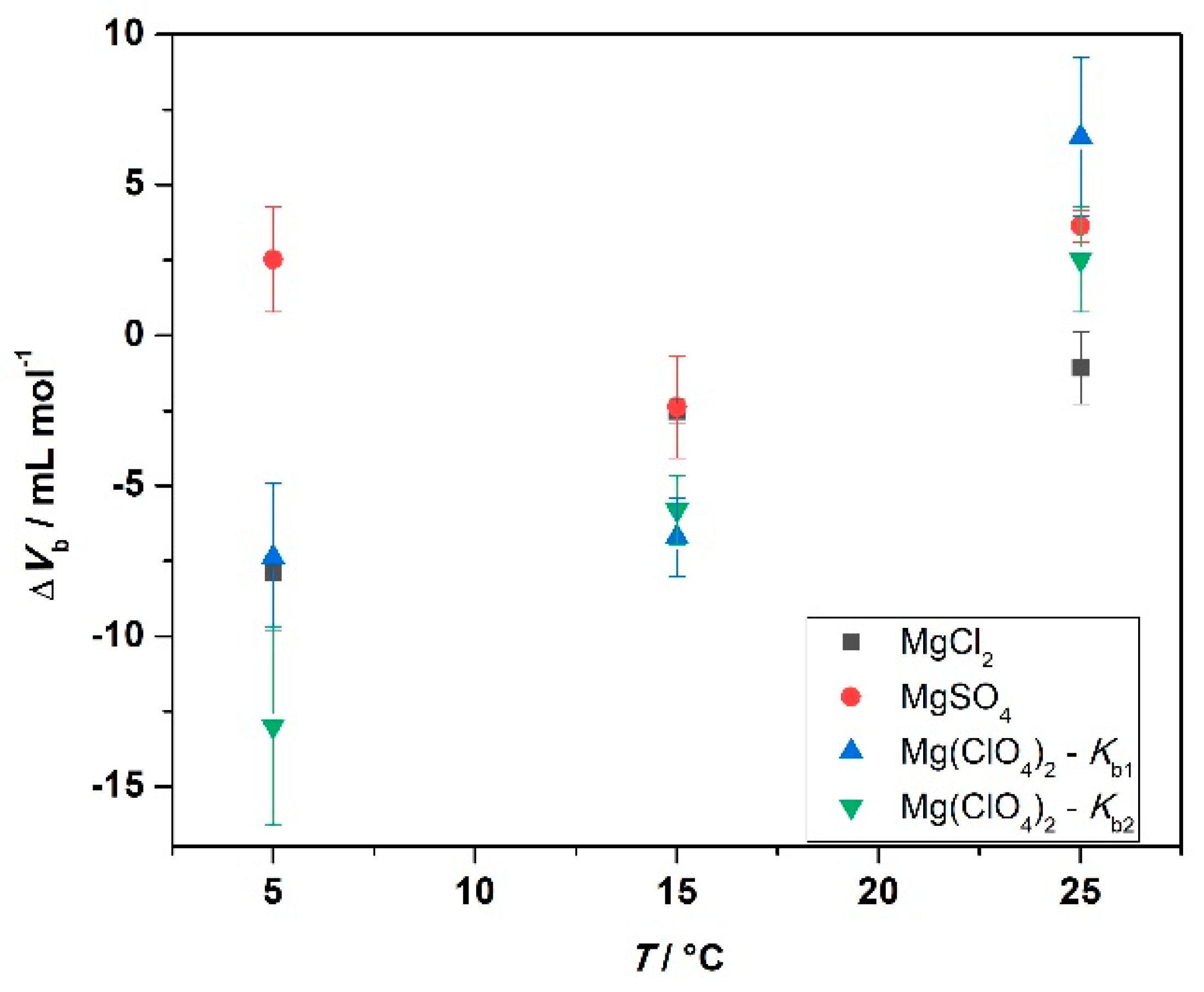
| MgCl2 | MgSO4 | Mg(ClO4)2 | Mg(ClO4)2 | |
|---|---|---|---|---|
| T/°C | ΔVb/mL mol−1 | ΔVb/mL mol−1 | ΔVb1/mL mol−1 | ΔVb2/mL mol−1 |
| 5 | −7.9 ± 0.1 | 2.0 ± 1.0 | −7.4 ± 2.4 | −12.9 ± 3.2 |
| 15 | −2.5 ± 0.4 | −2.0 ± 1.0 | −6.0 ± 1.0 | −5.0 ± 1.0 |
| 25 | −1.0 ± 1.0 | 3.6 ± 0.5 | 6.0 ± 2.0 | 2.0 ± 1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jahmidi-Azizi, N.; Oliva, R.; Gault, S.; Cockell, C.S.; Winter, R. The Effects of Temperature and Pressure on Protein-Ligand Binding in the Presence of Mars-Relevant Salts. Biology 2021, 10, 687. https://doi.org/10.3390/biology10070687
Jahmidi-Azizi N, Oliva R, Gault S, Cockell CS, Winter R. The Effects of Temperature and Pressure on Protein-Ligand Binding in the Presence of Mars-Relevant Salts. Biology. 2021; 10(7):687. https://doi.org/10.3390/biology10070687
Chicago/Turabian StyleJahmidi-Azizi, Nisrine, Rosario Oliva, Stewart Gault, Charles S. Cockell, and Roland Winter. 2021. "The Effects of Temperature and Pressure on Protein-Ligand Binding in the Presence of Mars-Relevant Salts" Biology 10, no. 7: 687. https://doi.org/10.3390/biology10070687
APA StyleJahmidi-Azizi, N., Oliva, R., Gault, S., Cockell, C. S., & Winter, R. (2021). The Effects of Temperature and Pressure on Protein-Ligand Binding in the Presence of Mars-Relevant Salts. Biology, 10(7), 687. https://doi.org/10.3390/biology10070687