
Assessment of Various Toxicity Endpoints in Duckweed (Lemna minor) at the Physiological, Biochemical, and Molecular Levels as a Measure of Diuron Stress

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture Conditions
2.2. Diuron Toxicity Testing
2.2.1. Measuring Total Frond Area and Fresh Weight
2.2.2. Pigment Contents
2.2.3. Chlorophyll a Fluorescence
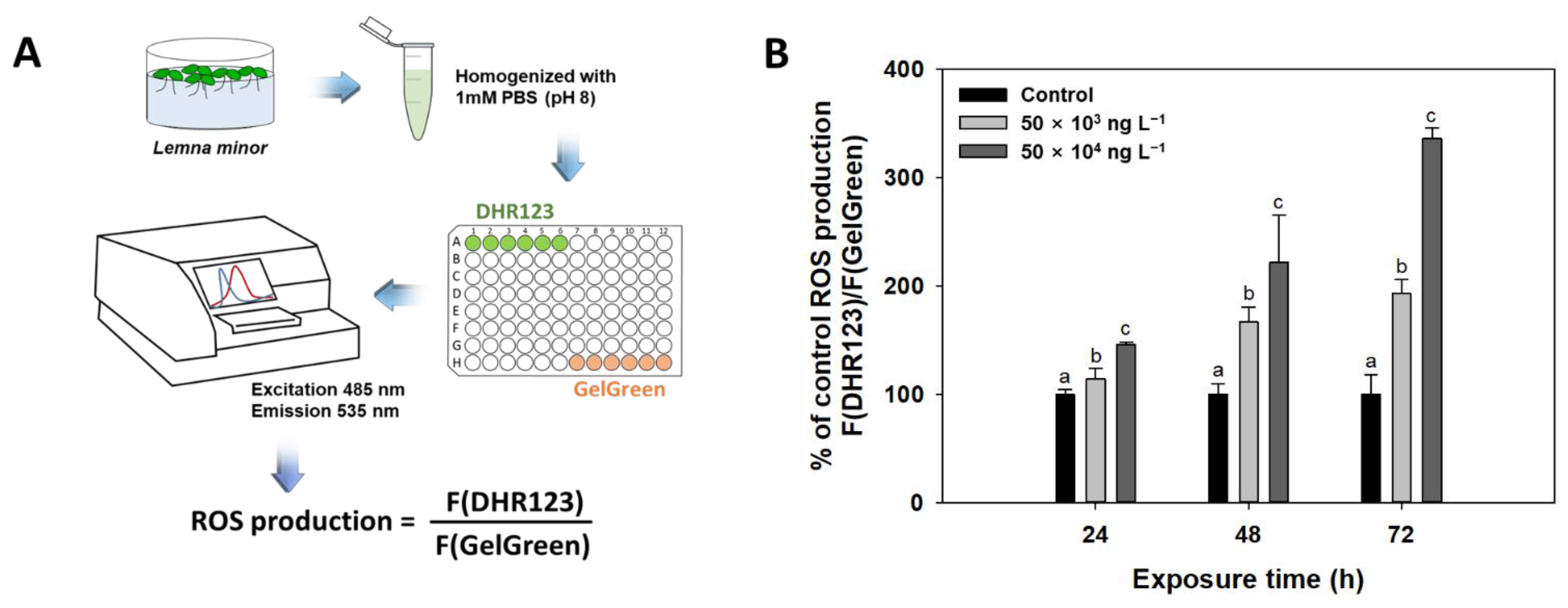
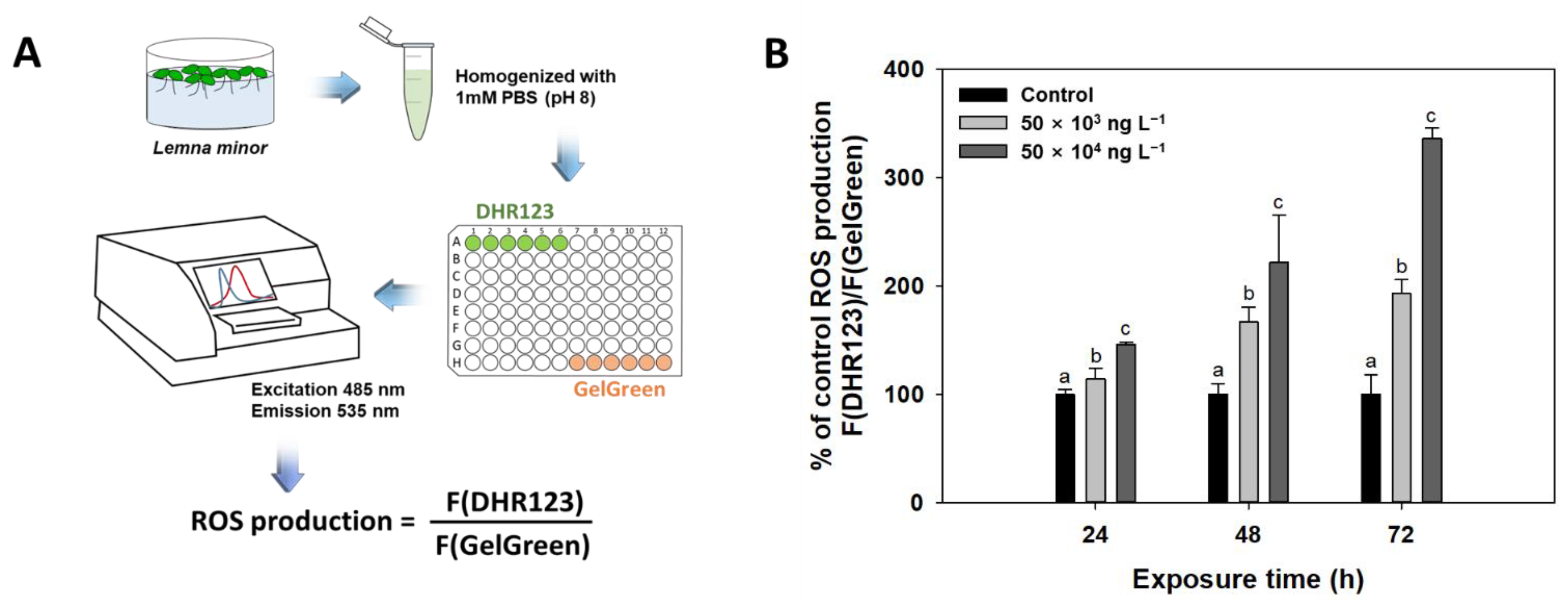
2.2.4. Reactive Oxygen Species (ROS) Measurements
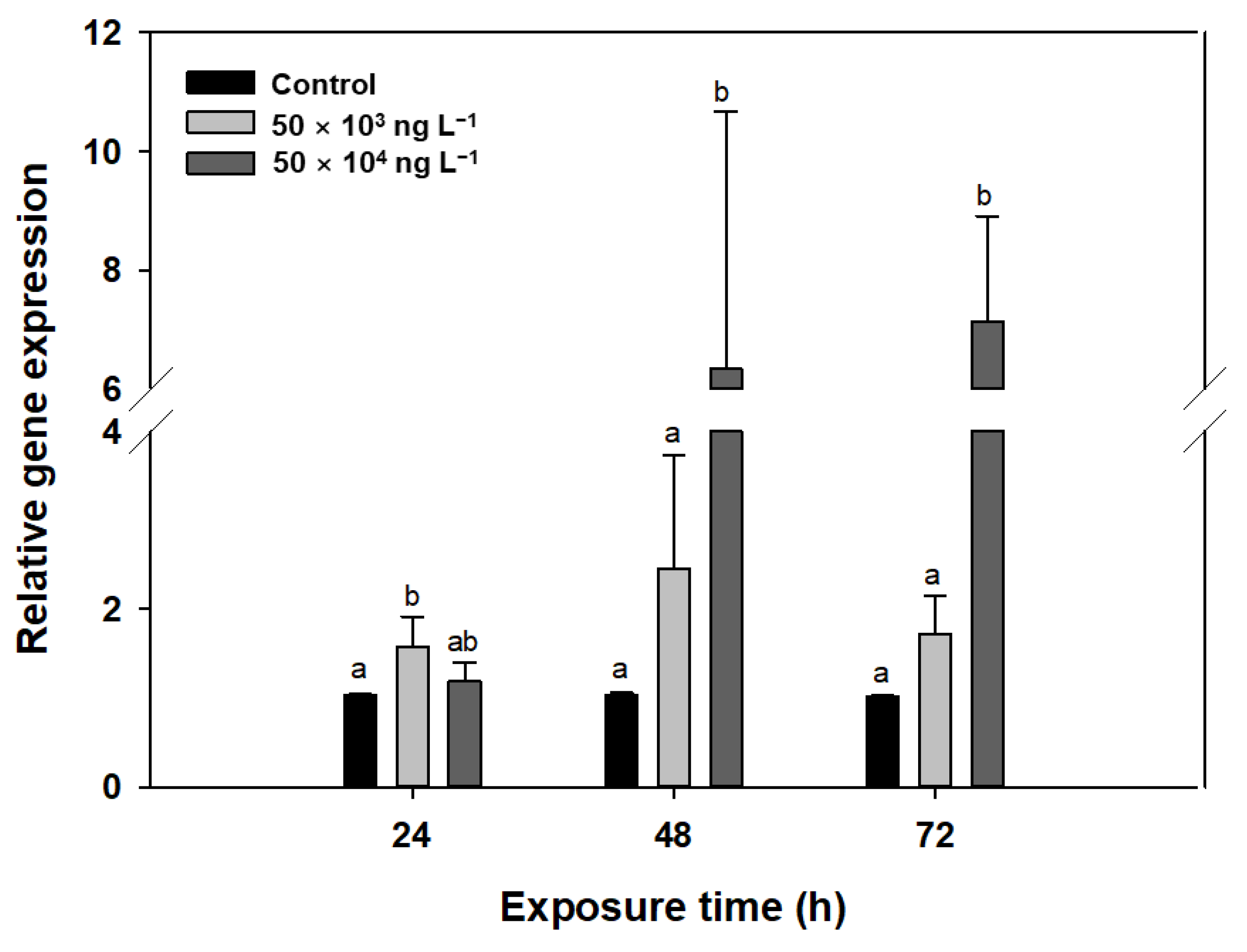
2.2.5. RNA Extraction, cDNA Synthesis, and RT-PCR Analysis
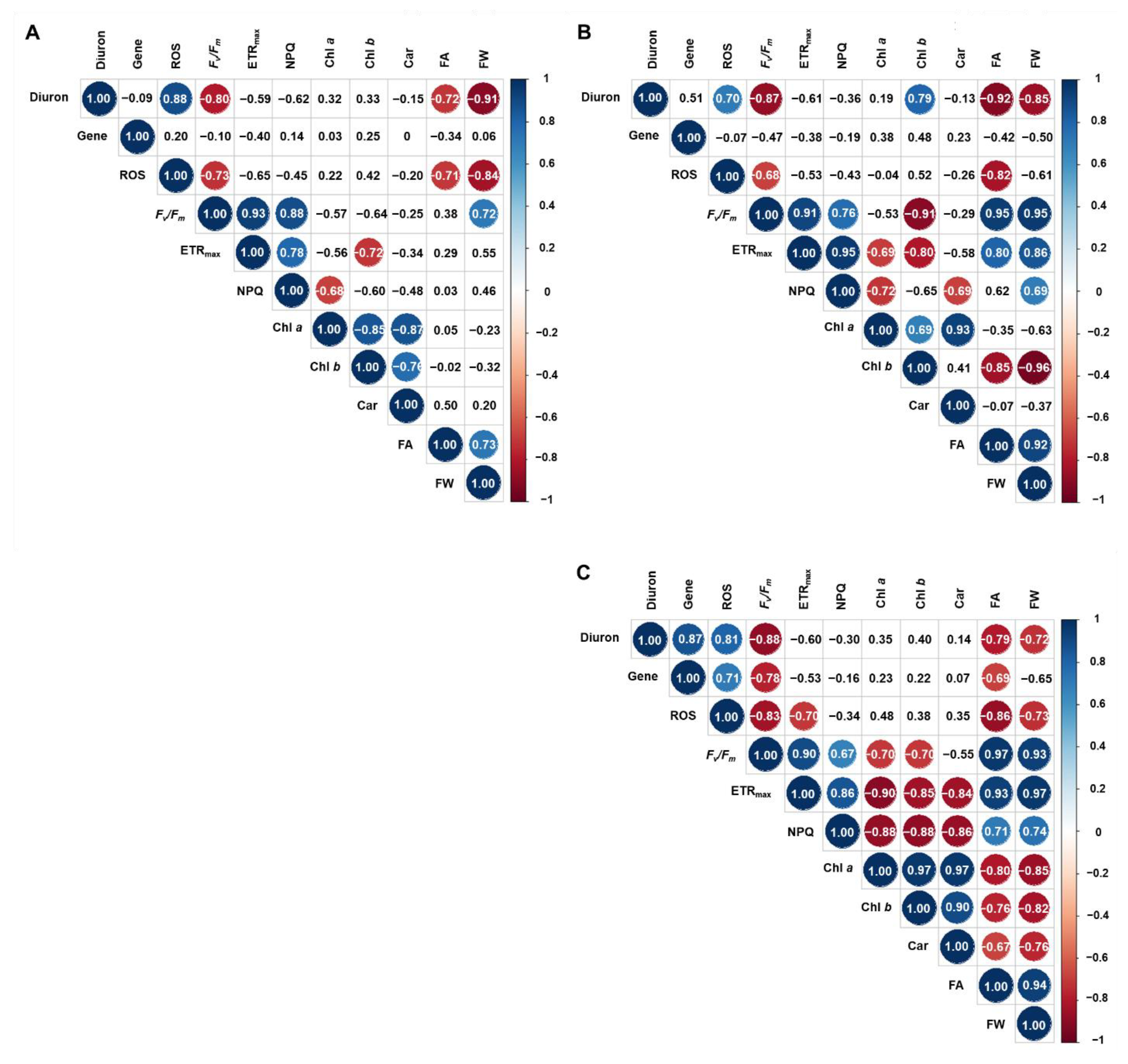
2.3. Statistical Analysis
3. Results and Discussion
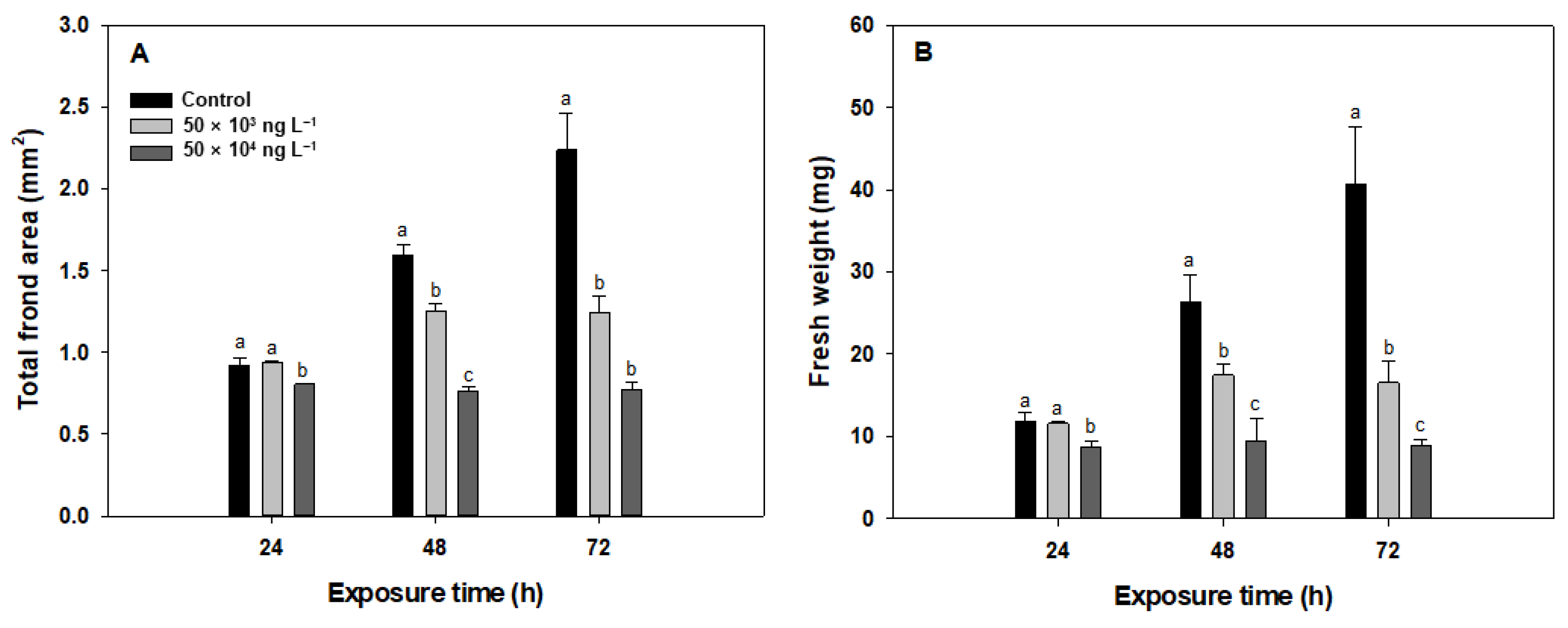
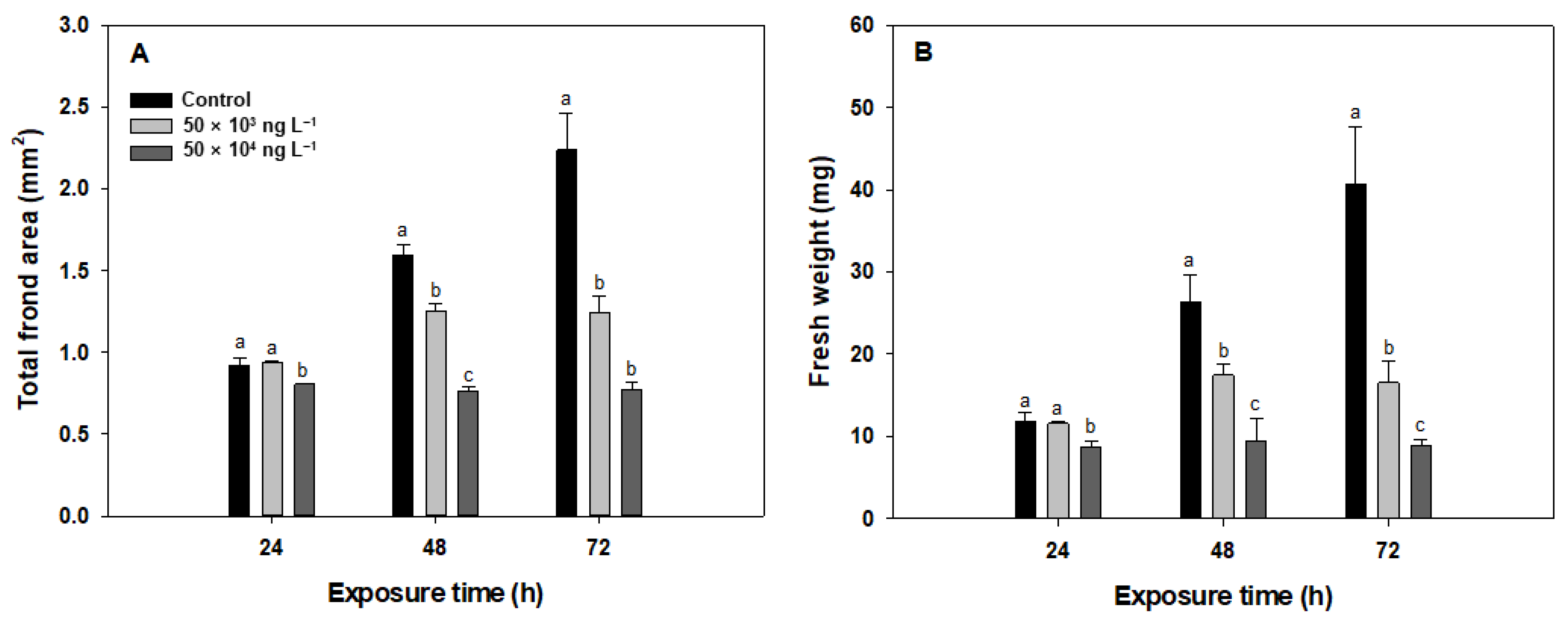
3.1. Total Frond Area and Fresh Weight
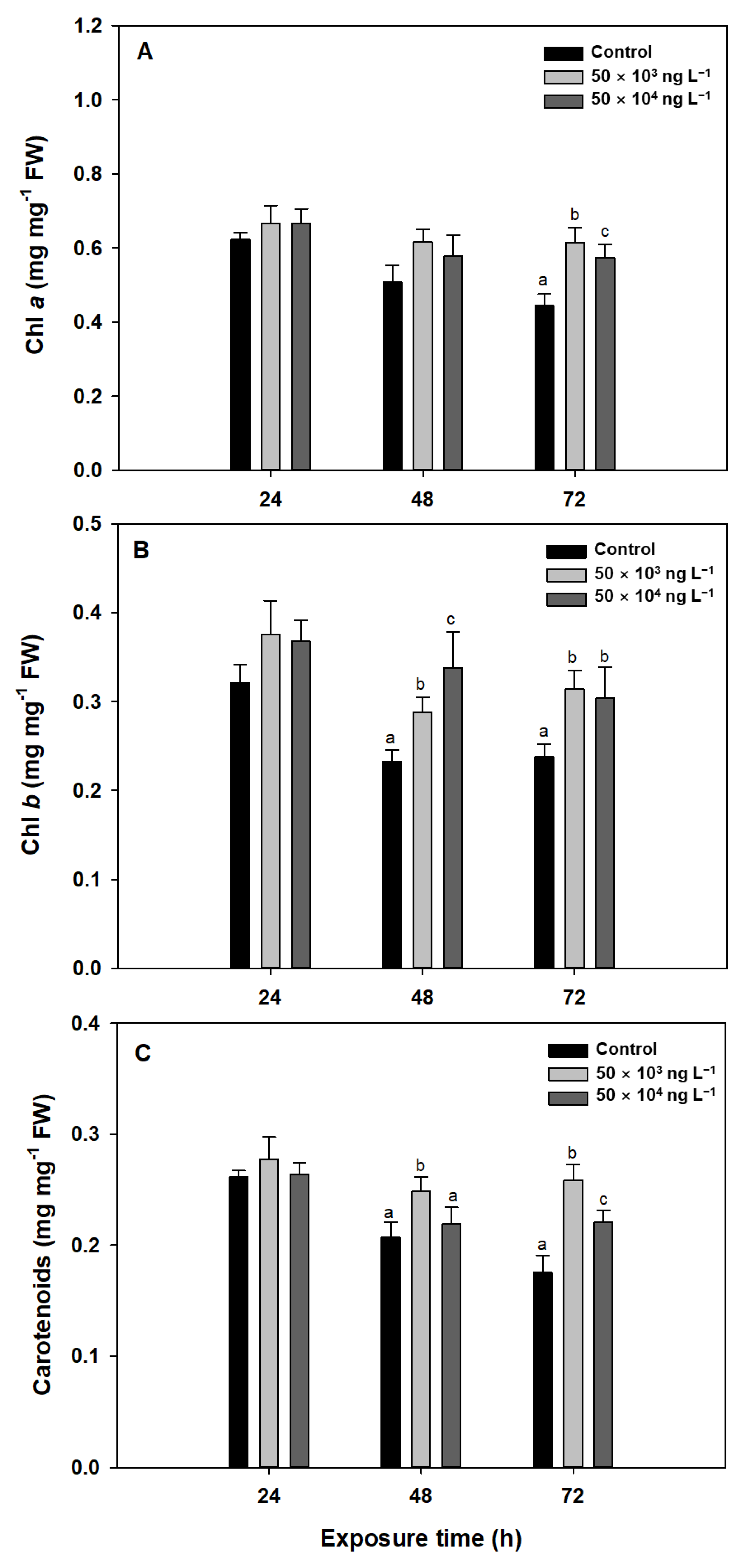
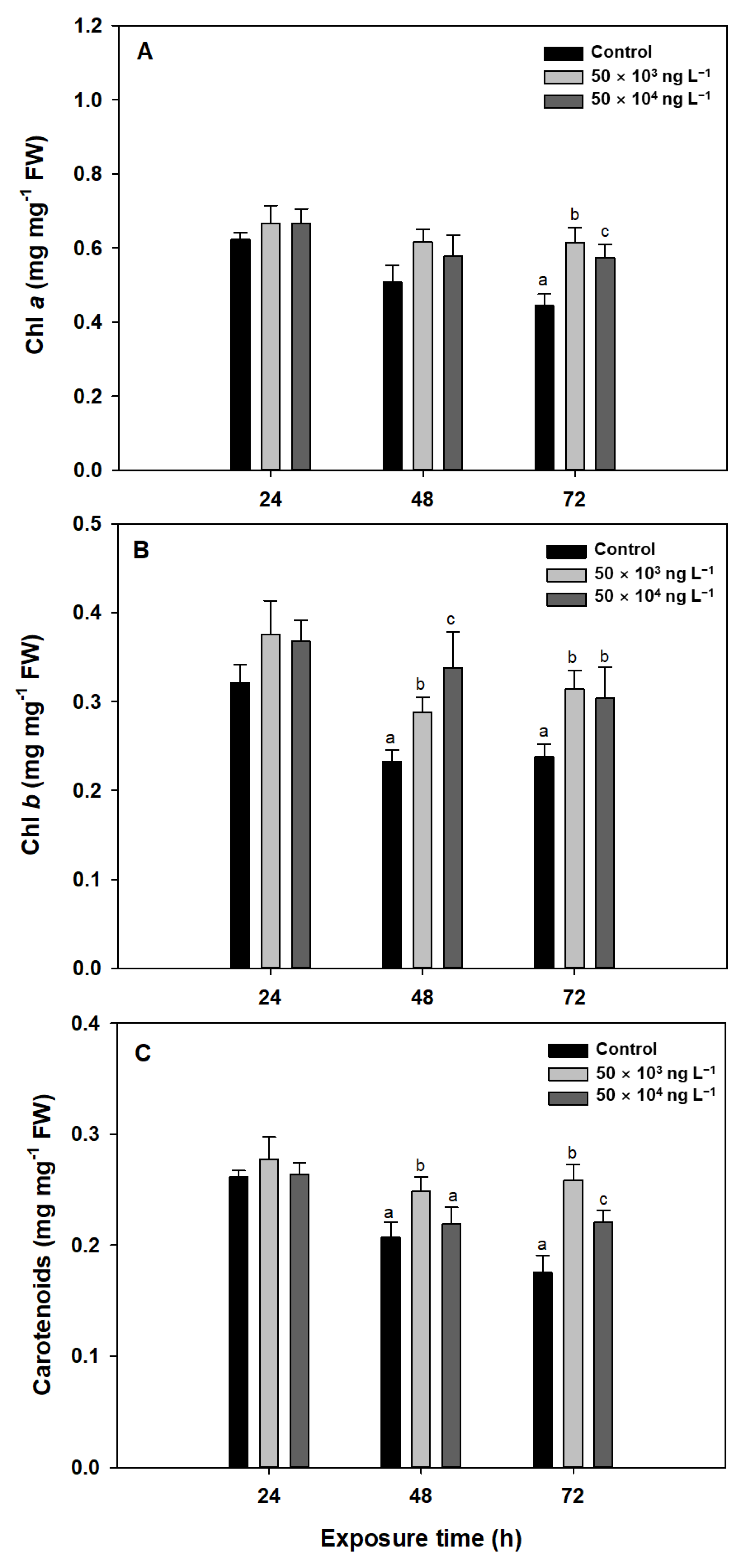
3.2. Photosynthetic Pigments
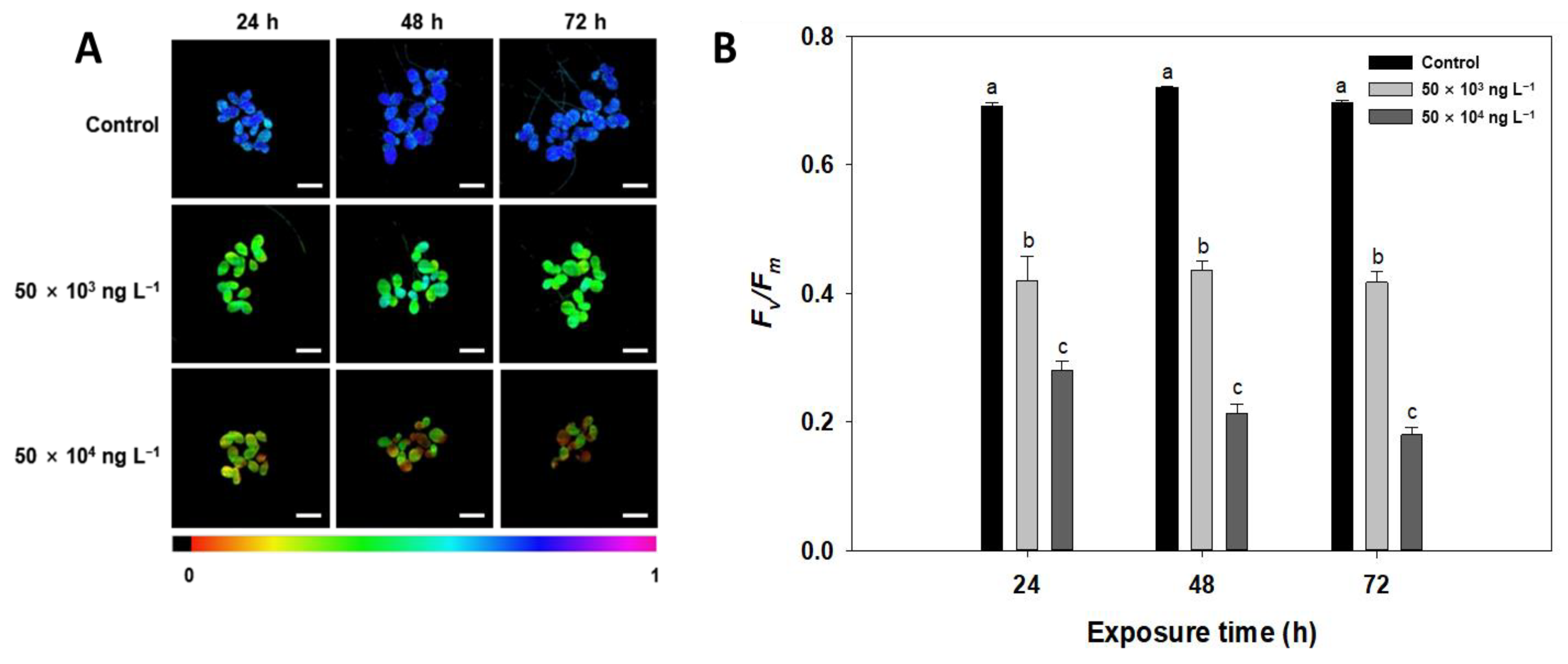
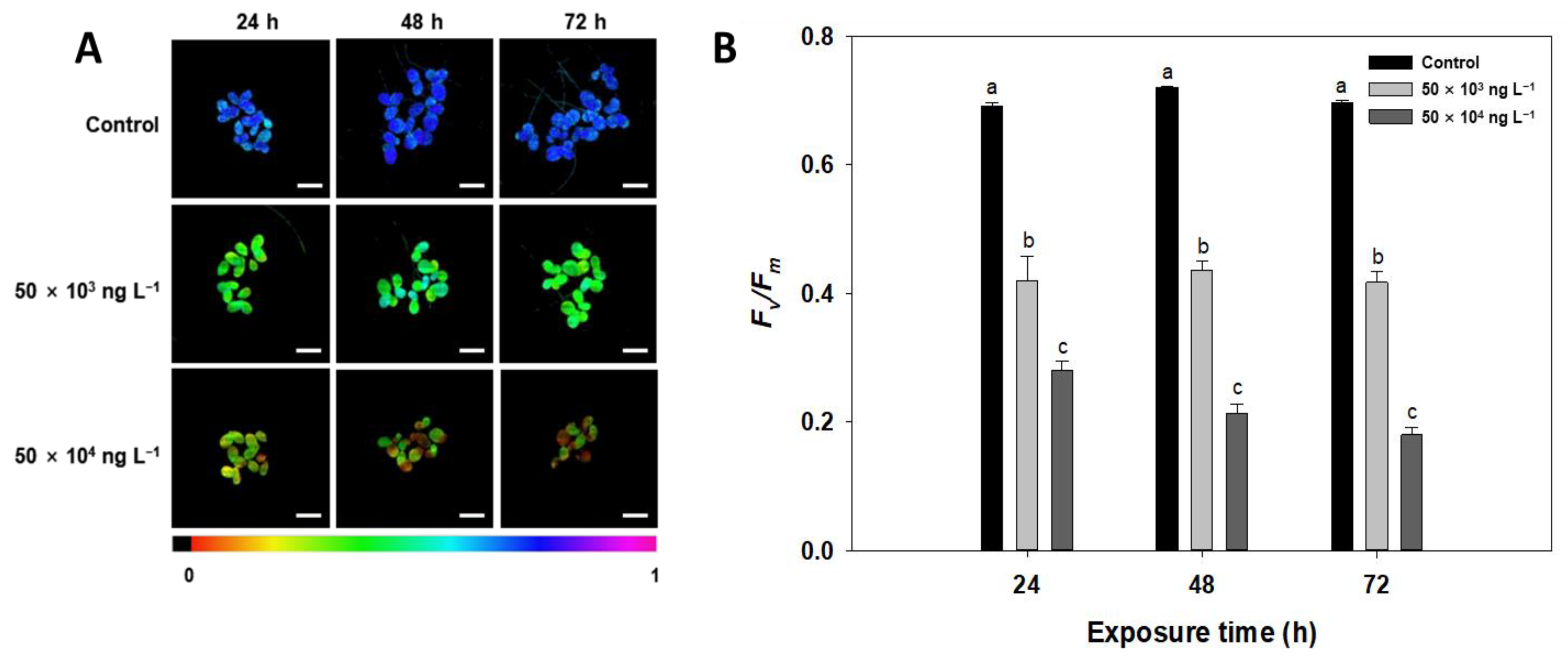
3.3. Chlorophyll a Fluorescence
3.4. Reactive Oxygen Species (ROS)
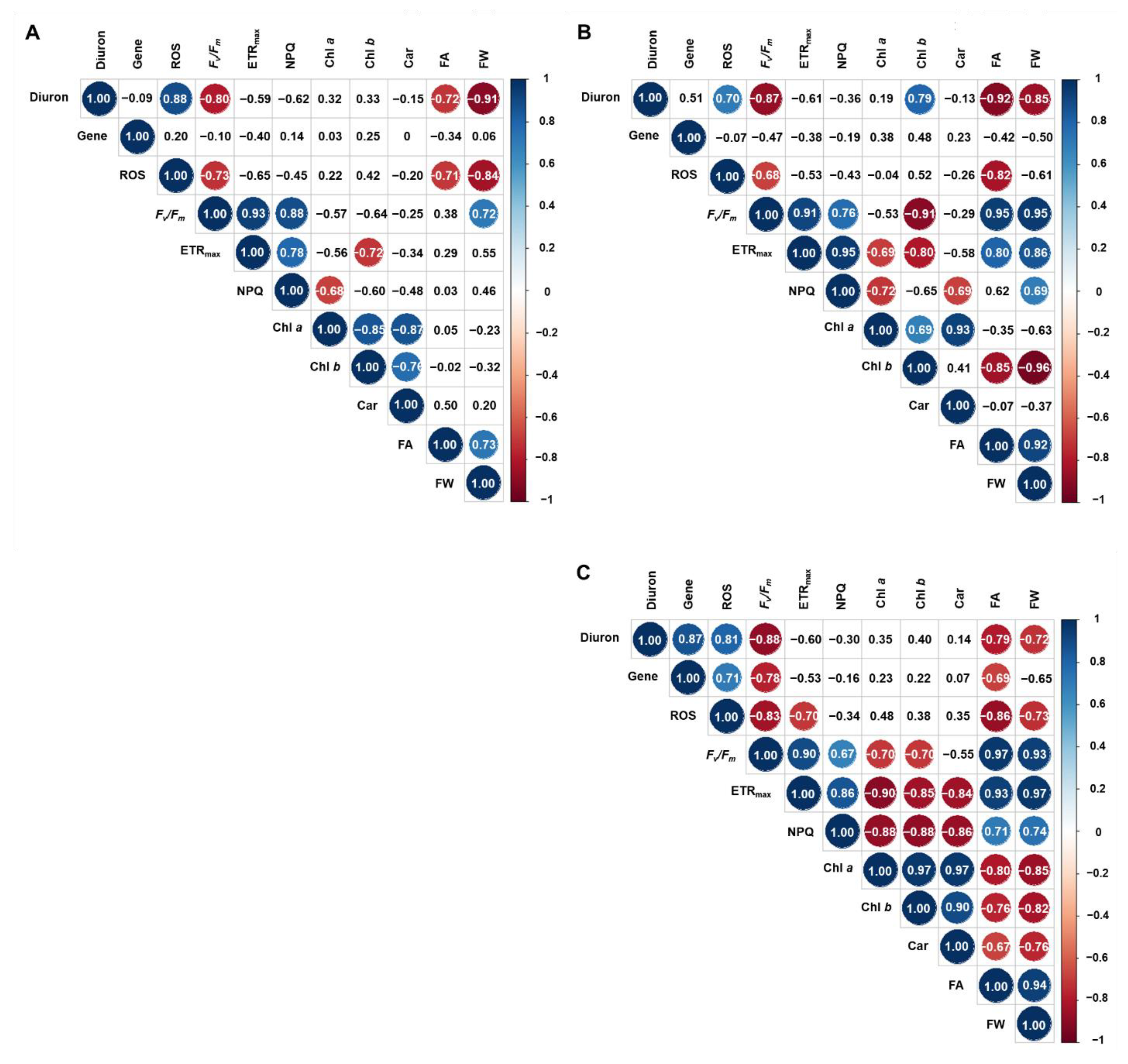
3.5. Gene Transcription
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fatima, M.; Mandiki, S.; Douxfils, J.; Silvestre, F.; Coppe, P.; Kestemont, P. Combined effects of herbicides on biomarkers reflecting immune–endocrine interactions in goldfish: Immune and antioxidant effects. Aquat. Toxicol. 2007, 81, 159–167. [Google Scholar] [CrossRef]
- Klöppel, H.; Kördel, W.; Stein, B. Herbicide transport by surface runoff and herbicide retention in a filter strip—rainfall and runoff simulation studies. Chemosphere 1997, 35, 129–141. [Google Scholar] [CrossRef]
- Prado, R.; Rioboo, C.; Herrero, C.; Cid, A. The herbicide paraquat induces alterations in the elemental and biochemical composition of non-target microalgal species. Chemosphere 2009, 76, 1440–1444. [Google Scholar] [CrossRef] [Green Version]
- Hernández, A.F.; Parrón, T.; Tsatsakis, A.M.; Requena, M.; Alarcón, R.; López-Guarnido, O. Toxic effects of pesticide mixtures at a molecular level: Their relevance to human health. Toxicology 2013, 307, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.S.; Choo, K.-s.; Yea, S.S.; Seo, Y.; Han, T. Effects of the phenylurea herbicide diuron on the physiology of Saccharina japonica aresch. Toxicol. Environ. Health Sci. 2010, 2, 188–199. [Google Scholar] [CrossRef]
- Duan, Q.; Duan, L.; Liu, Y.; Naidu, R.; Zhang, H.; Lei, Y. A novel in-situ passive sampling technique in the application of monitoring diuron in the aquatic environment. Environ. Technol. Innov. 2020, 20, 101073. [Google Scholar] [CrossRef]
- Giacomazzi, S.; Cochet, N. Environmental impact of diuron transformation: A review. Chemosphere 2004, 56, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Ralph, P.J. Herbicide toxicity of Halophila ovalis assessed by chlorophyll a fluorescence. Aquat. Bot. 2000, 66, 141–152. [Google Scholar] [CrossRef]
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of photosystem II under environmental stress. Biochim. Biophys. Acta (BBA) Bioenerg. 2007, 1767, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Lu, Y.; Goltsev, V.; Strasser, R.J.; Kalaji, H.M.; Wang, H.; Wang, X.; Chen, S.; Qiang, S. Comparative effect of tenuazonic acid, diuron, bentazone, dibromothymoquinone and methyl viologen on the kinetics of Chl a fluorescence rise OJIP and the MR820 signal. Plant Physiol. Biochem. 2020, 156, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Tiwari, S. Responses of plants to herbicides: Recent advances and future prospectives. In Plant Life Under Changing Environment: Responses and Management; Tripathi, D.K., Singh, V.P., Chauhan, D.K., Sharma, S., Prasad, S.M., Dubey, N.K., Ramawat, N., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 237–250. [Google Scholar]
- Kusumayuni, E.; Sriyani, N.; Hapsoro, D.; Utomo, S. Long-term application of diuron herbicides caused Eleusine indica weeds to become resistant to diuron. IOP Conf. Ser. Earth Environ. Sci. 2021, 739, 12034. [Google Scholar] [CrossRef]
- Jones, R. The ecotoxicological effects of photosystem II herbicides on corals. Mar. Pollut. Bull. 2005, 51, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, A.; Bados, P.; Estaun, A.R.; de Alencastro, L.F.; Taibi, S.; Einhorn, J.; Mougin, C. Concentrations and specific loads of glyphosate, diuron, atrazine, nonylphenol and metabolites thereof in French urban sewage sludge. Chemosphere 2007, 69, 1368–1373. [Google Scholar] [CrossRef] [PubMed]
- Camenzuli, L.; Scheringer, M.; Gaus, C.; Ng, C.A.; Hungerbühler, K. Describing the environmental fate of diuron in a tropical river catchment. Sci. Total Environ. 2012, 440, 178–185. [Google Scholar] [PubMed]
- Kaonga, C.C.; Takeda, K.; Sakugawa, H. Diuron, Irgarol 1051 and fenitrothion contamination for a river passing through an agricultural and urban area in Higashi Hiroshima City, Japan. Sci. Total Environ. 2015, 518, 450–458. [Google Scholar] [CrossRef]
- Bamba, D.; Atheba, P.; Robert, D.; Trokourey, A.; Dongui, B. Photocatalytic degradation of the diuron pesticide. Environ. Chem. Lett. 2008, 6, 163–167. [Google Scholar]
- Tan, C.; Gao, N.; Deng, Y.; An, N.; Deng, J. Heat-activated persulfate oxidation of diuron in water. Chem. Eng. J. 2012, 203, 294–300. [Google Scholar] [CrossRef]
- Macounová, K.; Krýsová, H.; Ludvík, J.; Jirkovský, J. Kinetics of photocatalytic degradation of diuron in aqueous colloidal solutions of Q-TiO2 particles. J. Photochem. Photobiol. A Chem. 2003, 156, 273–282. [Google Scholar] [CrossRef]
- Oturan, M.A.; Edelahi, M.C.; Oturan, N.; Aaron, J.-J. Kinetics of oxidative degradation/mineralization pathways of the phenylurea herbicides diuron, monuron and fenuron in water during application of the electro-Fenton process. Appl. Catal. B Environ. 2010, 97, 82–89. [Google Scholar] [CrossRef]
- Tomlin, C. The Pesticide Manual; British Crop Protection Council: Surrey, UK, 1997. [Google Scholar]
- Richardson, S.D.; Ternes, T.A. Water analysis: Emerging contaminants and current issues. Anal. Chem. 2018, 90, 398–428. [Google Scholar] [CrossRef]
- Park, J.-S.; Brown, M.T.; Han, T. Phenol toxicity to the aquatic macrophyte Lemna paucicostata. Aquat. Toxicol. 2012, 106, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.S.; Han, T. Physiological response of Lemna species to herbicides and its probable use in toxicity testing. Toxicol. Environ. Health Sci. 2010, 2, 39–49. [Google Scholar] [CrossRef]
- Lahive, E.; O’Halloran, J.; Jansen, M. Differential sensitivity of four Lemnaceae species to zinc sulphate. Environ. Exp. Bot. 2011, 71, 25–33. [Google Scholar] [CrossRef]
- Park, J.; Brown, M.T.; Depuydt, S.; Kim, J.K.; Won, D.-S.; Han, T. Comparing the acute sensitivity of growth and photosynthetic endpoints in three Lemna species exposed to four herbicides. Environ. Pollut. 2017, 220, 818–827. [Google Scholar] [CrossRef] [Green Version]
- Landolt, E.; Kandeler, R. Biosystematic investigations in the family of duckweeds (Lemnaceae), Vol. 4: The family of Lemnaceae-a monographic study, Vol. 2 (phytochemistry, physiology, application, bibliography). Veroeffentlichungen Geobot. Inst. ETH Stiftung Ruebel 1987, 95, 638. [Google Scholar]
- Landolt, E. Lemnaceae duckweed family. J. Ariz. Nev. Acad. Sci. 1992, 26, 10–14. [Google Scholar]
- Scherr, C.; Simon, M.; Spranger, J.; Baumgartner, S. Test system stability and natural variability of a Lemna gibba L. bioassay. PLoS ONE 2008, 3, e3133. [Google Scholar] [CrossRef] [PubMed]
- Wang, W. Literature review on duckweed toxicity testing. Environ. Res. 1990, 52, 7–22. [Google Scholar] [CrossRef]
- Giddings, J.M.; Arts, G.; Hommen, U. The relative sensitivity of macrophyte and algal species to herbicides and fungicides: An analysis using species sensitivity distributions. Integr. Environ. Assess. Manag. 2013, 9, 308–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliferis, K.A.; Materzok, S.; Paziotou, G.N.; Chrysayi-Tokousbalides, M. Lemna minor L. as a model organism for ecotoxicological studies performing 1H NMR fingerprinting. Chemosphere 2009, 76, 967–973. [Google Scholar] [CrossRef]
- Mkandawire, M.; Teixeira da Silva, J.A.; Dudel, E.G. The Lemna bioassay: Contemporary issues as the most standardized plant bioassay for aquatic ecotoxicology. Crit. Rev. Environ. Sci. Technol. 2014, 44, 154–197. [Google Scholar] [CrossRef]
- Kummerová, M.; Zezulka, Š.; Babula, P.; Tříska, J. Possible ecological risk of two pharmaceuticals diclofenac and paracetamol demonstrated on a model plant Lemna minor. J. Hazard. Mater. 2016, 302, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Kostopoulou, S.; Ntatsi, G.; Arapis, G.; Aliferis, K.A. Assessment of the effects of metribuzin, glyphosate, and their mixtures on the metabolism of the model plant Lemna minor L. applying metabolomics. Chemosphere 2020, 239, 124582. [Google Scholar] [CrossRef] [PubMed]
- Nestler, H.; Groh, K.J.; Schönenberger, R.; Behra, R.; Schirmer, K.; Eggen, R.I.; Suter, M.J.-F. Multiple-endpoint assay provides a detailed mechanistic view of responses to herbicide exposure in Chlamydomonas reinhardtii. Aquat. Toxicol. 2012, 110, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, R.A. Mineral requirements of Lemna minor. Plant Physiol. 1946, 21, 42–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K. Chlorophyll fluorescence signatures of leaves during the autumnal chlorophyll breakdown. J. Plant Physiol. 1987, 131, 101–110. [Google Scholar] [CrossRef]
- Schreiber, U.; Quayle, P.; Schmidt, S.; Escher, B.I.; Mueller, J.F. Methodology and evaluation of a highly sensitive algae toxicity test based on multiwell chlorophyll fluorescence imaging. Biosens. Bioelectron. 2007, 22, 2554–2563. [Google Scholar] [CrossRef]
- Platt, T.; Gallegos, C.; Harrison, W.G. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. J. Mar. Res. 1981, 38, 687–701. [Google Scholar]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.T.; Park, J.T.; Choi, K.; Choi, H.J.C.; Jung, C.W.; Kim, G.R.; Lee, Y.S.; Park, S.C. Chemical screening identifies ROCK as a target for recovering mitochondrial function in Hutchinson-Gilford progeria syndrome. Aging Cell 2017, 16, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.-i.; Ihara, T.; Tamura, H.; Tanaka, S.; Ikeda, T.; Kajihara, H.; Dissanayake, C.; Abdel-Motaal, F.F.; El-Sayed, M.A. α-Tomatine, the major saponin in tomato, induces programmed cell death mediated by reactive oxygen species in the fungal pathogen Fusarium oxysporum. FEBS Lett. 2007, 581, 3217–3222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, R.; Zhu, Q.; Tang, X.; Zhao, Q. Transcriptomic and physiological analysis of common duckweed Lemna minor responses to NH4+ toxicity. BMC Plant Biol. 2016, 16, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleczkowski, L.A. Inhibitors of photosynthetic enzymes/carriers and metabolism. Annu. Rev. Plant Biol. 1994, 45, 339–367. [Google Scholar] [CrossRef]
- Williams, S.L.; Carranza, A.; Kunzelman, J.; Datta, S.; Kuivila, K.M. Effects of the herbicide diuron on cordgrass (Spartina foliosa) reflectance and photosynthetic parameters. Estuaries Coasts 2009, 32, 146–157. [Google Scholar] [CrossRef]
- Lichtenthaler, H.; Meier, D.; Retzlaff, G.; Hamm, R. Distribution and effects of bentazon in crop plants and weeds. Zeitschrift für Naturforschung C 1982, 37, 889–897. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chloroplast biogenesis, its inhibition and modification by new herbicide compounds. Zeitschrift für Naturforschung C 1984, 39, 492–499. [Google Scholar] [CrossRef]
- Macinnis-Ng, C.M.; Ralph, P.J. Short-term response and recovery of Zostera capricorni photosynthesis after herbicide exposure. Aquat. Bot. 2003, 76, 1–15. [Google Scholar] [CrossRef]
- Percival, G.C. The use of chlorophyll fluorescence to identify chemical and environmental stress in leaf tissue of three oak (Quercus) species. J. Arboric. 2005, 31, 215. [Google Scholar]
- Bai, X.; Sun, C.; Xie, J.; Song, H.; Zhu, Q.; Su, Y.; Qian, H.; Fu, Z. Effects of atrazine on photosynthesis and defense response and the underlying mechanisms in Phaeodactylum tricornutum. Environ. Sci. Pollut. Res. 2015, 22, 17499–17507. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.C.; Flores, F.; Kaserzon, S.; Reeks, T.A.; Negri, A.P. Toxicity of the herbicides diuron, propazine, tebuthiuron, and haloxyfop to the diatom Chaetoceros muelleri. Sci. Rep. 2020, 10, 1–14. [Google Scholar]
- DeEll, J.R.; van Kooten, O.; Prange, R.K.; Murr, D.P. Applications of chlorophyll fluorescence techniques in postharvest physiology. Hortic. Rev. 1999, 23, 69–107. [Google Scholar]
- Björkman, O.; Demmig, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar] [CrossRef]
- Snel, J.F.; Vos, J.H.; Gylstra, R.; Brock, T.C. Inhibition of photosystem II (PSII) electron transport as a convenient endpoint to assess stress of the herbicide linuron on freshwater plants. Aquat. Ecol. 1998, 32, 113–123. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Complementary PS II quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the saturation pulse method. PAM Appl. Notes 2008, 1, 201–247. [Google Scholar]
- Frankart, C.; Eullaffroy, P.; Vernet, G. Comparative effects of four herbicides on non-photochemical fluorescence quenching in Lemna minor. Environ. Exp. Bot. 2003, 49, 159–168. [Google Scholar] [CrossRef]
- Han, T.; Han, Y.-S.; Park, C.Y.; Jun, Y.S.; Kwon, M.J.; Kang, S.-H.; Brown, M.T. Spore release by the green alga Ulva: A quantitative assay to evaluate aquatic toxicants. Environ. Pollut. 2008, 153, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Brack, W.; Frank, H. Chlorophyll a fluorescence: A tool for the investigation of toxic effects in the photosynthetic apparatus. Ecotoxicol. Environ. Saf. 1998, 40, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; Juneau, P. Oxidative stress in duckweed (Lemna minor L.) induced by glyphosate: Is the mitochondrial electron transport chain a target of this herbicide? Environ. Pollut. 2016, 218, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Philosoph-Hadas, S.; Meir, S.; Akiri, B.; Kanner, J. Oxidative defense systems in leaves of three edible herb species in relation to their senescence rates. J. Agric. Food Chem. 1994, 42, 2376–2381. [Google Scholar] [CrossRef]
- Teisseire, H.; Vernet, G. Ascorbate and glutathione contents in duckweed, Lemna minor, as biomarkers of the stress generated by copper, folpet and diuron. Biomarkers 2000, 5, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Eichelmann, H.; Talts, E.; Oja, V.; Padu, E.; Laisk, A. Rubisco in planta kcat is regulated in balance with photosynthetic electron transport. J. Exp. Bot. 2009, 60, 4077–4088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Zhu, N.; Lavoie, M.; Wang, J.; Qian, H.; Fu, Z. Copper toxicity to Phaeodactylum tricornutum: A survey of the sensitivity of various toxicity endpoints at the physiological, biochemical, molecular and structural levels. Biometals 2014, 27, 527–537. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequence |
|---|---|
| 18S Rrna * (housekeeping gene) | Forward: 5′AGAGGAACAGTCGGGGGCATT-3′ Reverse: 5′-CGGCATCGTTTACGGTTGAGA-3′ |
| rbcL | Forward: 5′-GTCCATGTACCAGTAGAAGATTCGGC-3′ Reverse: 5′-ATGTCACCACAAACAGAGACTAAAGC-3′ |
| Diuron (ng L−1) | 24 h Exposure | 48 h Exposure | 72 h Exposure | |||
|---|---|---|---|---|---|---|
| ETRmax | NPQ | ETRmax | NPQ | ETRmax | NPQ | |
| 0 | 34 a ± 0.71 | 0.55 a ± 0.001 | 34 a ± 0.99 | 0.59 a ± 0.006 | 35 a ± 1.9 | 0.56 a ± 0.02 |
| 50 × 103 | 0.75 b ± 0.09 | 0.47 b ± 0.02 | 1.5 b ± 0.12 | 0.28 b ± 0.004 | 1.5 b ± 0.24 | 0.31 b ± 0.01 |
| 50 × 104 | 0.12 b ± 0.09 | 0.45 b ± 0.02 | 0.01 b ± 0.00 | 0.36 c ± 0.01 | 0.01 b ± 0.00 | 0.38 b ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Depuydt, S.; Shin, K.; Choi, S.; Kim, G.; Lee, Y.H.; Park, J.T.; Han, T.; Park, J. Assessment of Various Toxicity Endpoints in Duckweed (Lemna minor) at the Physiological, Biochemical, and Molecular Levels as a Measure of Diuron Stress. Biology 2021, 10, 684. https://doi.org/10.3390/biology10070684
Lee H, Depuydt S, Shin K, Choi S, Kim G, Lee YH, Park JT, Han T, Park J. Assessment of Various Toxicity Endpoints in Duckweed (Lemna minor) at the Physiological, Biochemical, and Molecular Levels as a Measure of Diuron Stress. Biology. 2021; 10(7):684. https://doi.org/10.3390/biology10070684
Chicago/Turabian StyleLee, Hojun, Stephen Depuydt, Kisik Shin, Soyeon Choi, Geonhee Kim, Yun Haeng Lee, Joon Tae Park, Taejun Han, and Jihae Park. 2021. "Assessment of Various Toxicity Endpoints in Duckweed (Lemna minor) at the Physiological, Biochemical, and Molecular Levels as a Measure of Diuron Stress" Biology 10, no. 7: 684. https://doi.org/10.3390/biology10070684
APA StyleLee, H., Depuydt, S., Shin, K., Choi, S., Kim, G., Lee, Y. H., Park, J. T., Han, T., & Park, J. (2021). Assessment of Various Toxicity Endpoints in Duckweed (Lemna minor) at the Physiological, Biochemical, and Molecular Levels as a Measure of Diuron Stress. Biology, 10(7), 684. https://doi.org/10.3390/biology10070684