
Ancestry Studies in Forensic Anthropology: Back on the Frontier of Racism



Abstract
:Simple Summary
Abstract
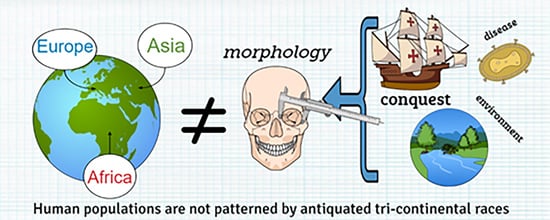
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Landmark Precision and Reliability
2.3. Geometric Morphometrics
2.4. Hierarchical Clustering
2.5. Spatial Analysis
3. Results
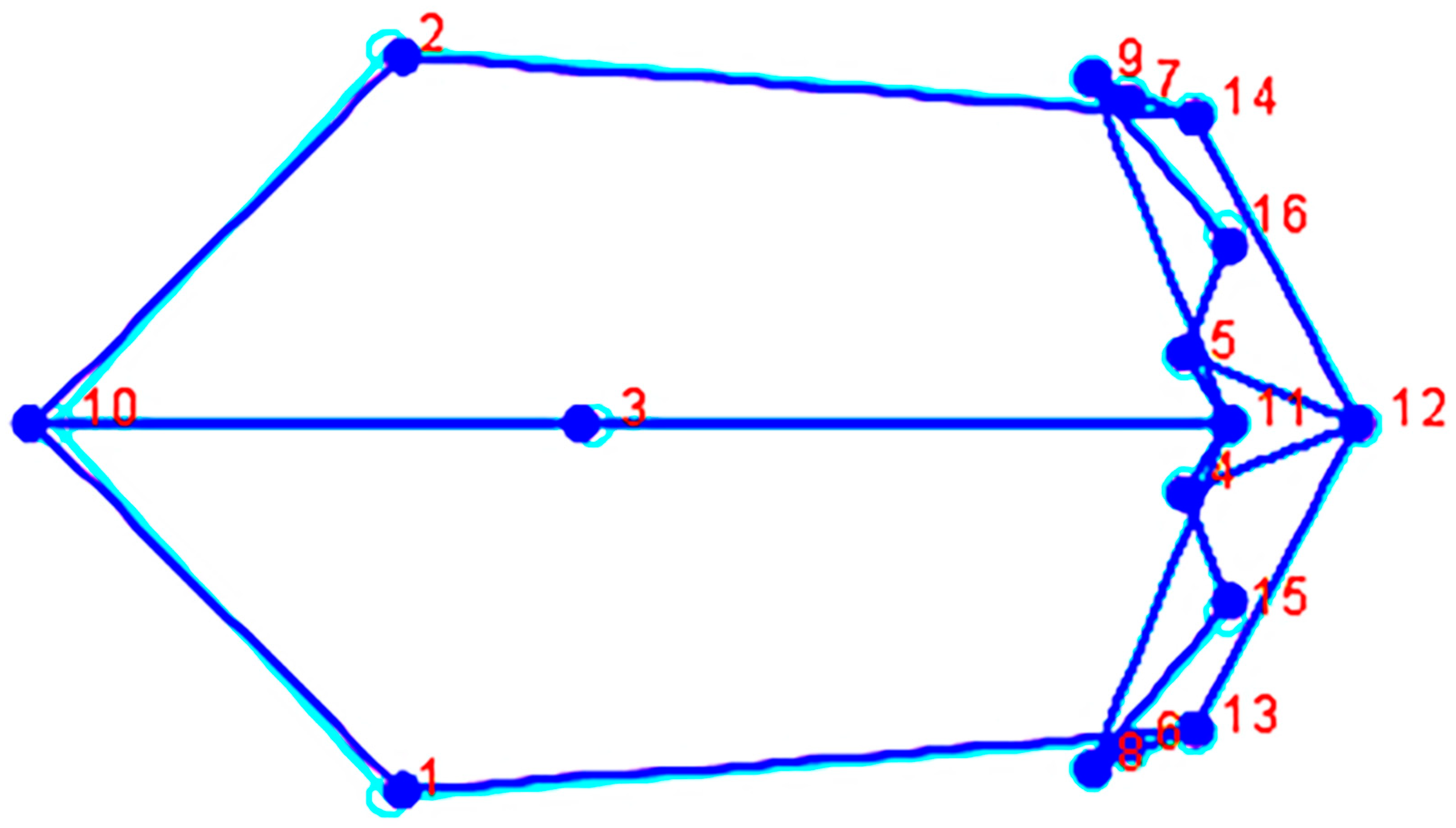
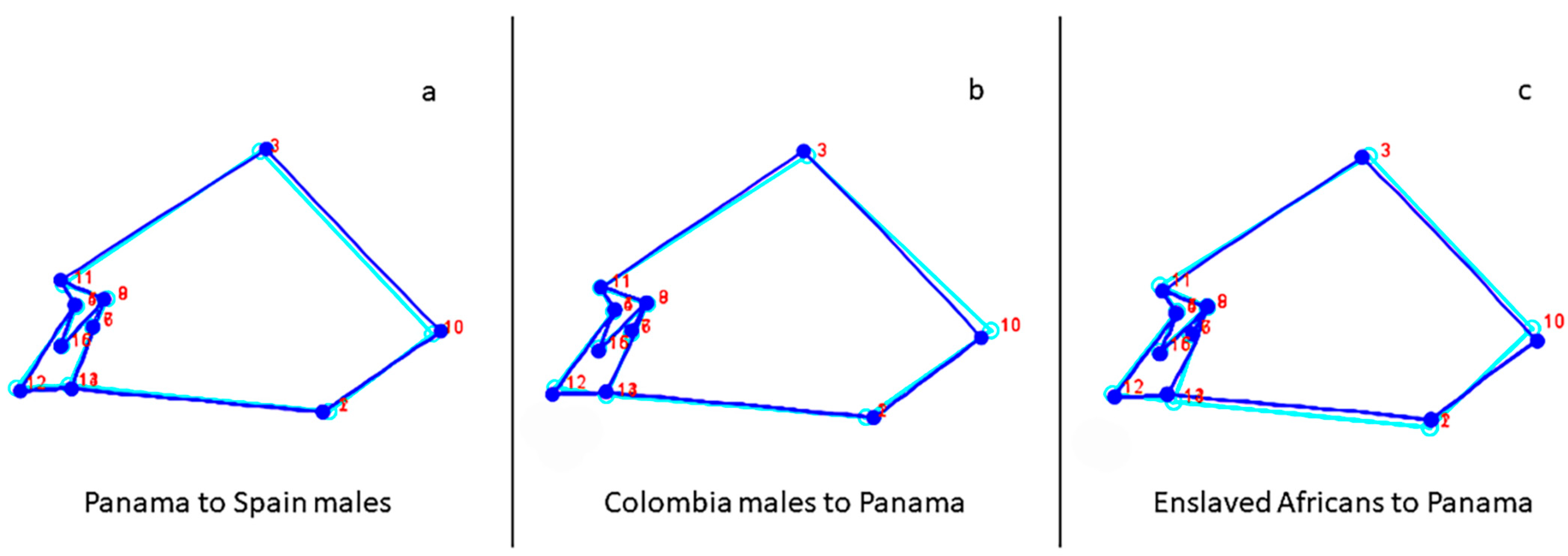
3.1. Geometric Morphometrics
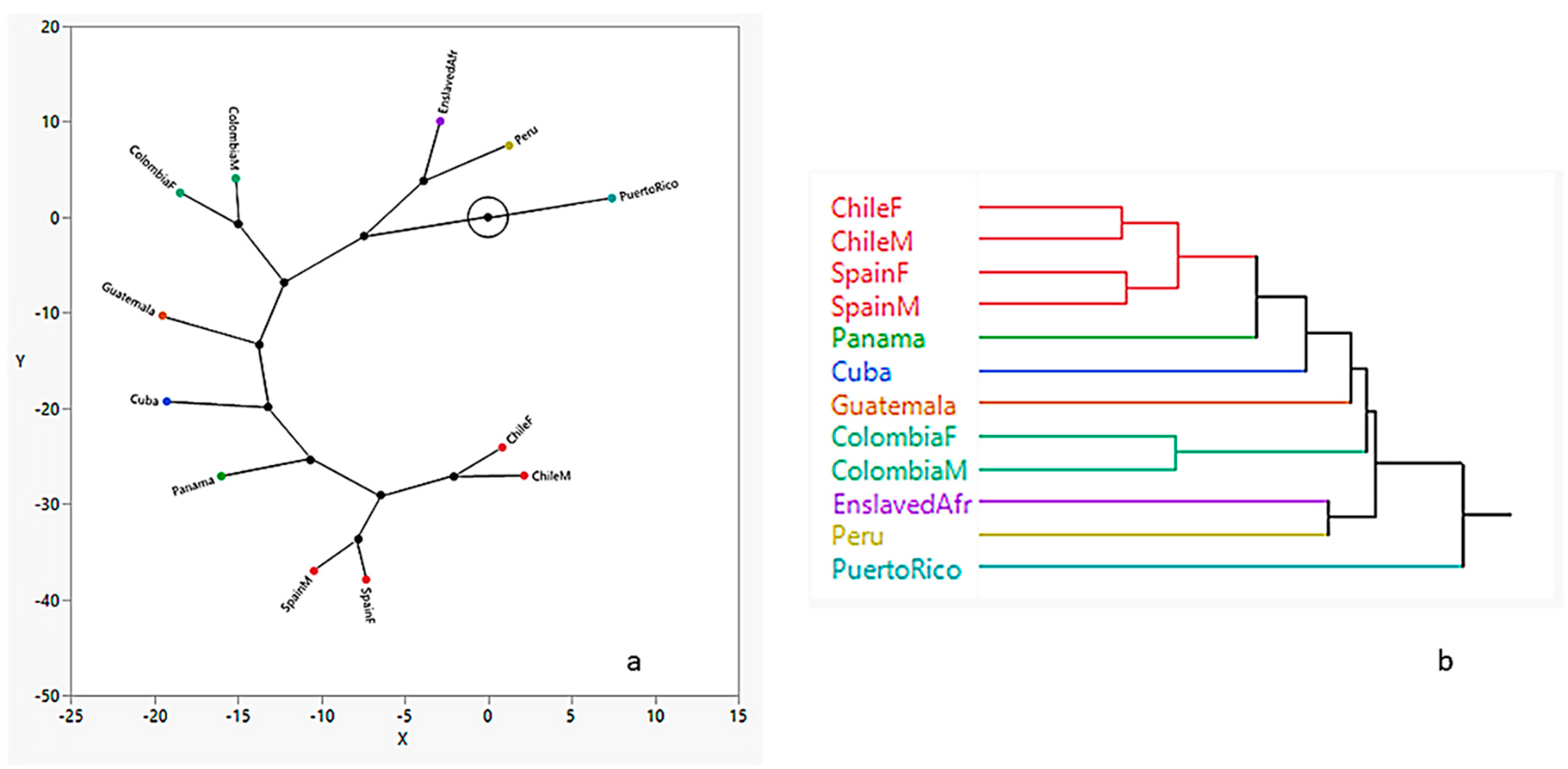
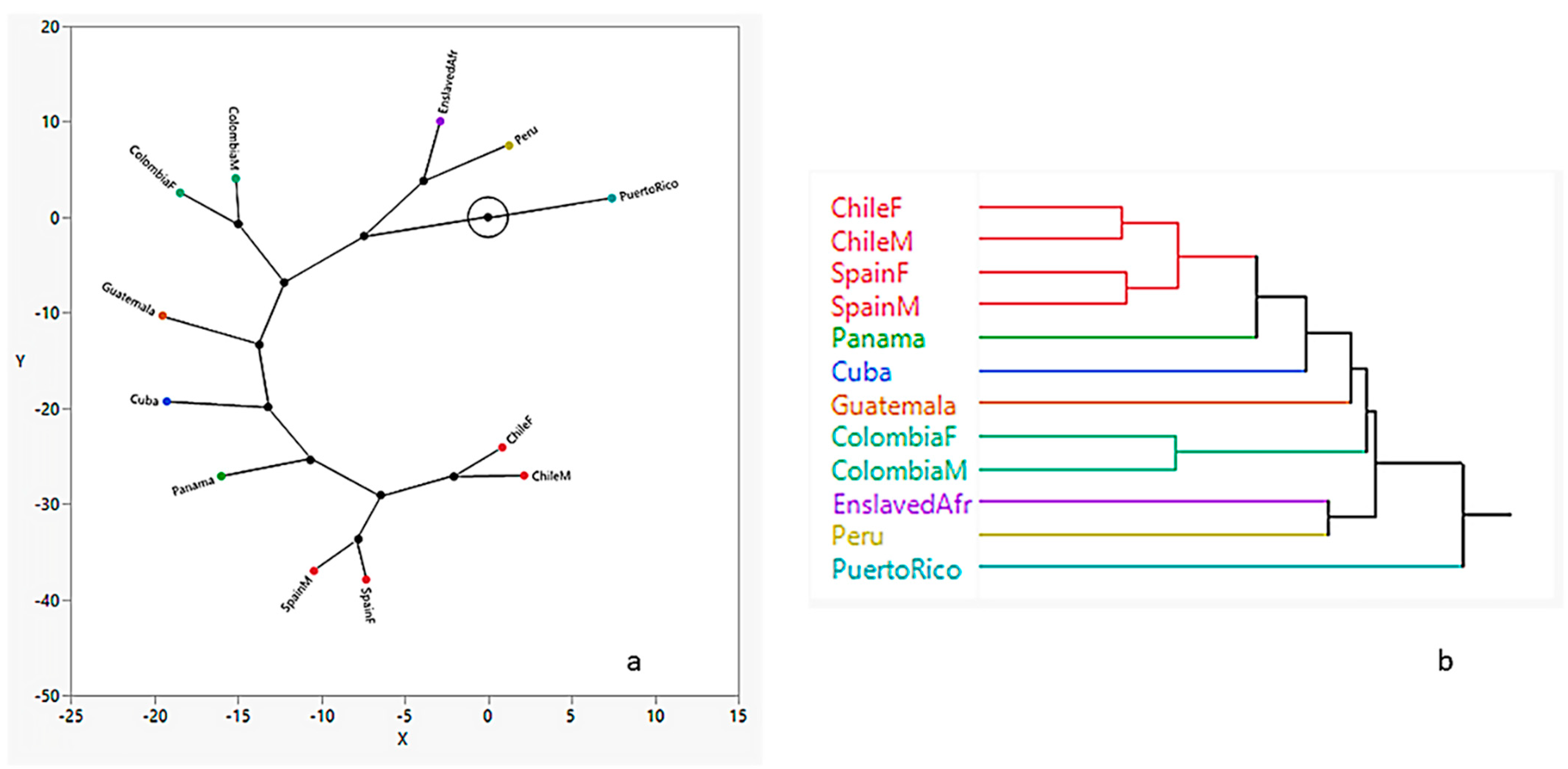
3.2. Hierarchical Clustering
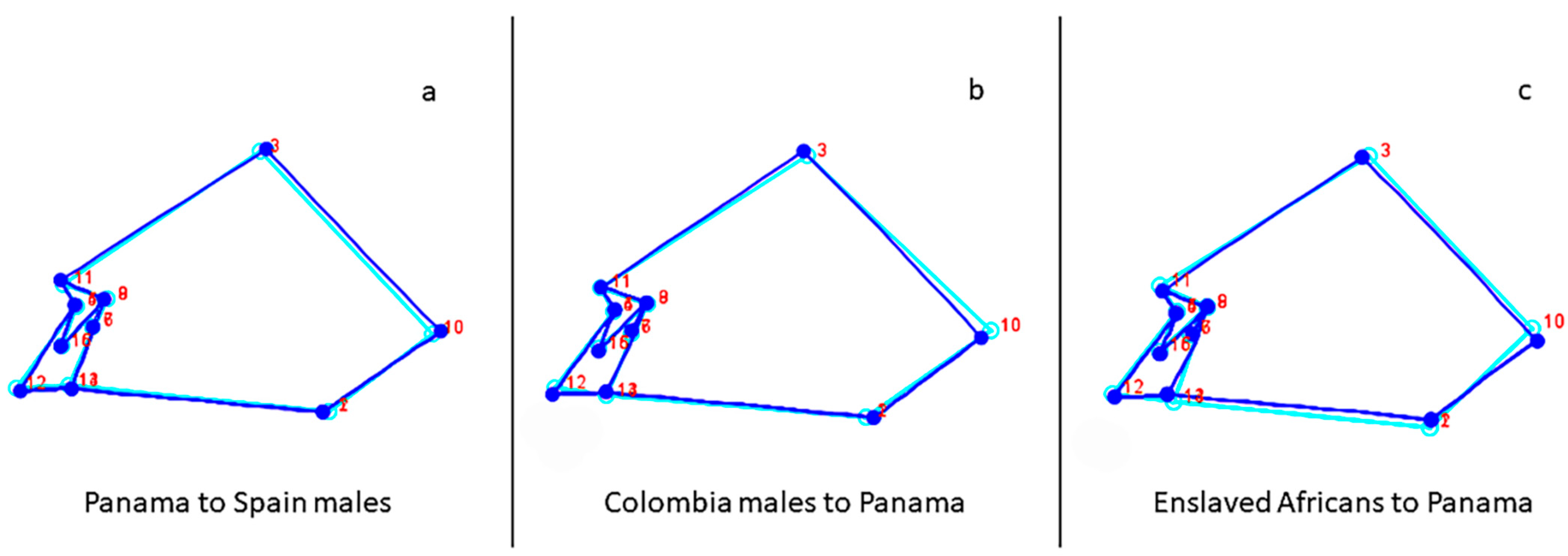
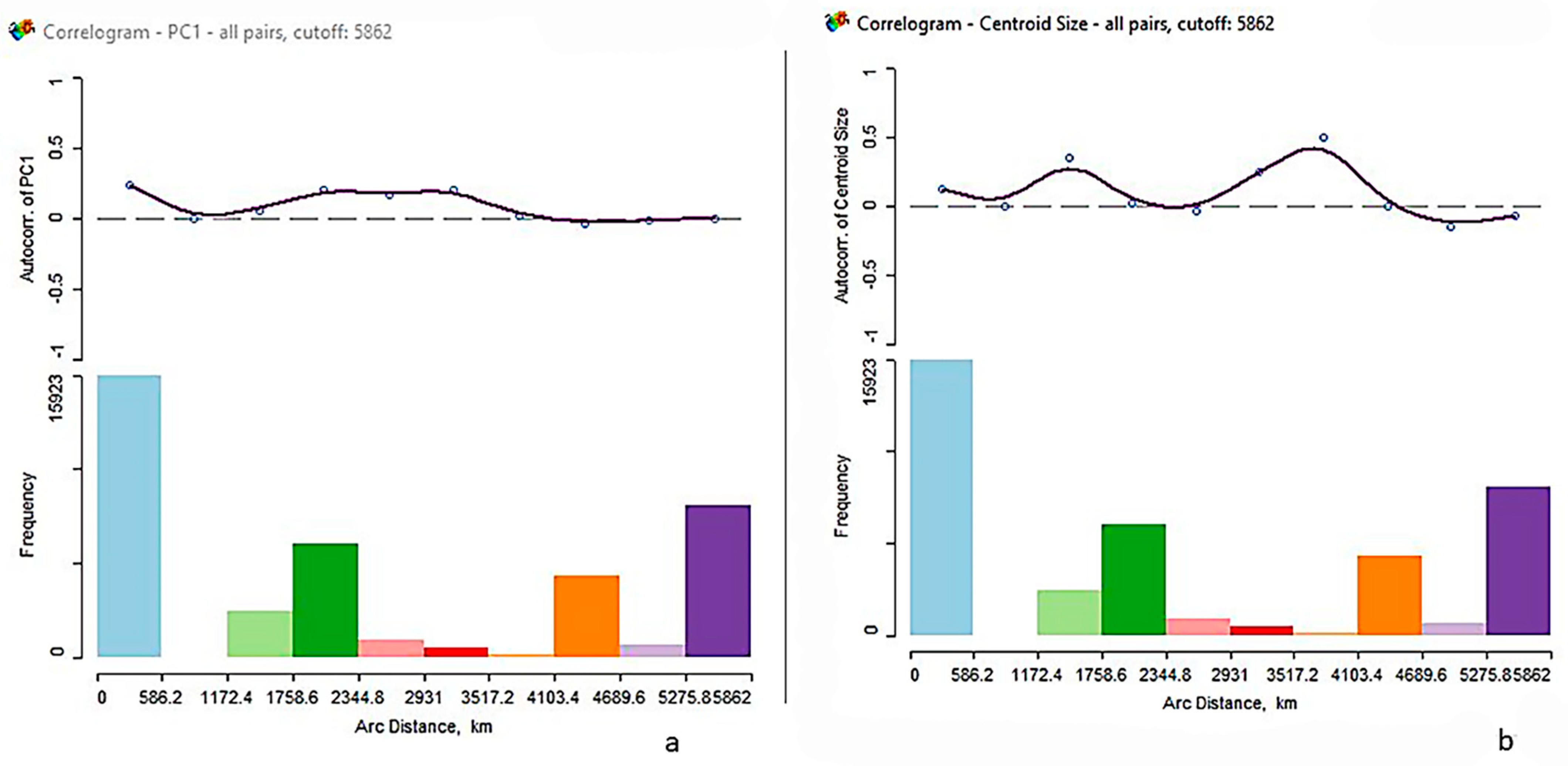
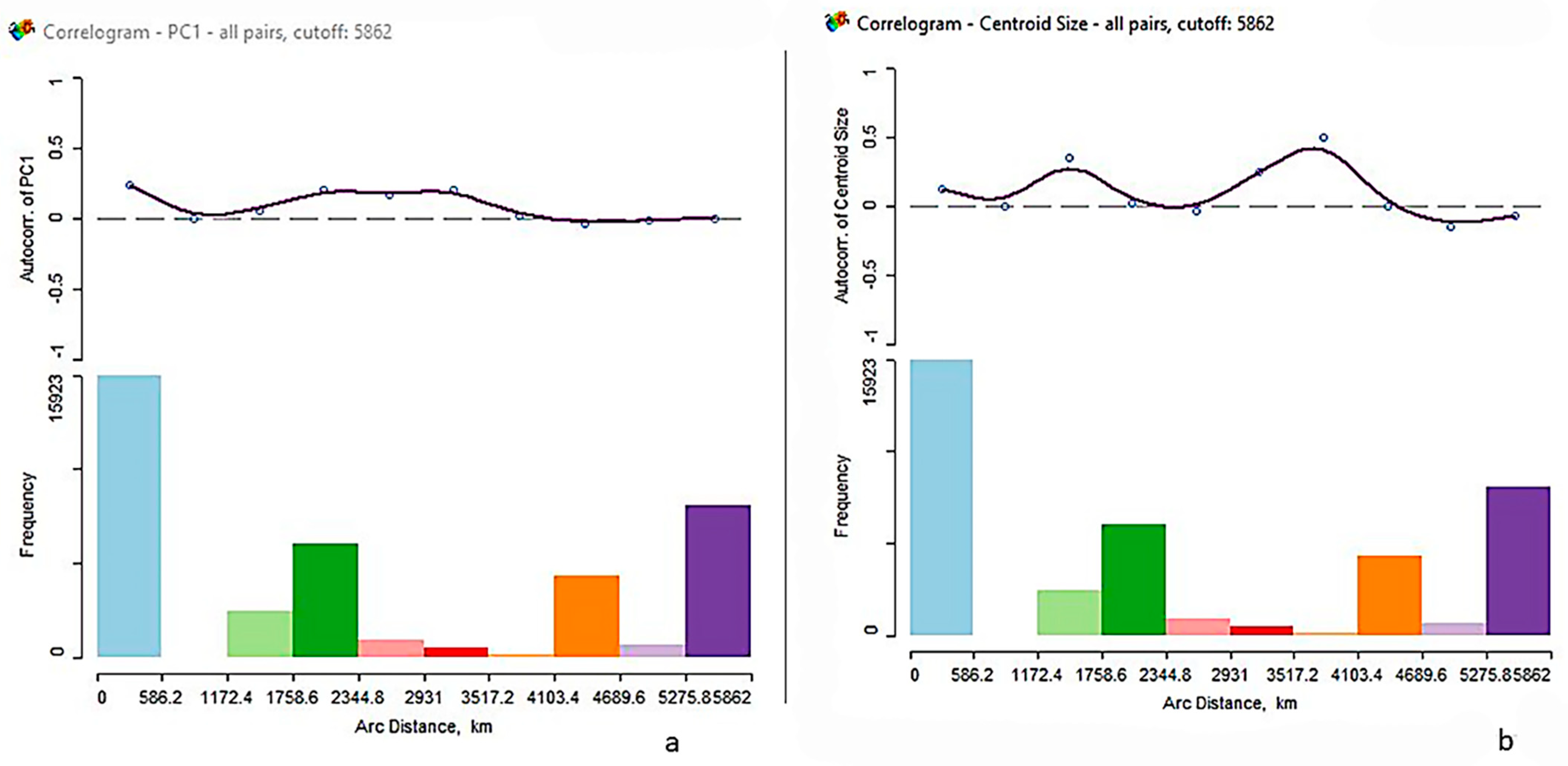
3.3. Spatial Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Langley, N.R.; Tersigni-Tarrant, M.A. Forensic Anthropology: A Comprehensive Introduction; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Smay, D.; Armelagos, G. Galileo Wept: A Critical Assessment of the Use of Race in Forensic Anthropology. Transform. Anthropol. 2000, 9, 19–29. [Google Scholar] [CrossRef]
- Bethard, J.D.; DiGangi, E.A. Letter to the Editor—Moving Beyond a Lost Cause: Forensic Anthropology and Ancestry Estimates in the United States. J. Forensic Sci. 2020, 65, 1791–1792. [Google Scholar] [CrossRef] [PubMed]
- DiGangi, E.A.; Bethard, J.D. Uncloaking a Lost Cause: Decolonizing ancestry estimation in the United States. Am. J. Phys. Anthropol. 2021, 175, 422–436. [Google Scholar] [CrossRef]
- Edgar, H.J.; Hunley, K.L. Race reconciled?: How biological anthropologists view human variation. Am. J. Phys. Anthropol. 2009, 139, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, L.; Stevenson, B.W.; Reynolds, L.T. Race and Anthropology: A Core Concept without Consensus. Anthropol. Educ. Q. 1989, 20, 67–73. [Google Scholar] [CrossRef]
- Sauer, N.J. Forensic Anthropology and the Concept of Race—If Races Don’t Exist, Why Are Forensic Anthropologists So Good at Identifying Them. Soc. Sci. Med. 1992, 34, 107–111. [Google Scholar] [CrossRef]
- Ousley, S.; Jantz, R.; Freid, D. Understanding race and human variation: Why forensic anthropologists are good at identfying race. Am. J. Phys. Anthropol. 2009, 139, 68–76. [Google Scholar] [CrossRef]
- Stull, K.E.; Bartelink, E.J.; Klales, A.R.; Berg, G.E.; Kenyhercz, M.W.; L’Abbé, E.N.; Mariscal, C. Commentary on: Bethard JD, DiGangi EA. Letter to the Editor-Moving beyond a lost cause: Forensic anthropology and ancestry estimates in theUnited States. J. Forensic Sci. 2021, 66, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Goodman, A.H. Bred in the Bone? Sciences 1997, 37, 20–25. [Google Scholar] [CrossRef]
- Blakey, M.L. Scientific racism and the biological concept of race. Lit. Psychol. 1999, 45, 29. [Google Scholar]
- Albanese, J.; Saunders, S.R. Is it possible to escape racial typology in forensic identification? In Forensic Anthropology and Medicine: Complementary Sciences From Recovery to Cause of Death; Schmitt, A., Cunha, E., Pinheiro, J., Eds.; Humana: Ttowa, NJ, USA, 2006; pp. 281–316. [Google Scholar]
- Maier, C.; Craig, A.; Adams, D.M. Language use in ancestry research and estimation. J. Forensic Sci. 2021, 66, 11–24. [Google Scholar] [CrossRef]
- Ross, A.H.; Slice, D.E.; Ubelaker, D.H.; Falsetti, A.B. Population affinities of 19th Century Cuban crania: Implications for identification criteria in South Florida Cuban Americans. J. Forensic Sci. 2002, 49, 1–6. [Google Scholar] [CrossRef]
- Brues, A.M. Forensic diagnosis of race—General race vs specific populations. Soc. Sci. Med. 1992, 34, 125–128. [Google Scholar] [CrossRef]
- Caspari, R. Race, then and now: 1918 revisited. Am. J. Phys. Anthropol. 2018, 165, 924–938. [Google Scholar] [CrossRef] [Green Version]
- Von Cramon-Taubadel, N. Evolutionary insights into global patterns of human cranial diversity: Population history, climatic and dietary effects. J. Anthropol. Sci. 2014, 92, 43–77. [Google Scholar] [PubMed]
- Sneath, P.H.; Sokal, R.R. Numerical Taxonomy. The Principles and Practice of Numerical Classification; Stony Brook University: Stony Brook, NY, USA, 1975. [Google Scholar]
- Ross, A.H. Regional isolation in the Balkan region: An analysis of craniofacial variation. Am. J. Phys. Anthropol. 2004, 124, 73–80. [Google Scholar] [CrossRef]
- Coon, C.S. The Origin of Races; Alfred A. Knopf.: New York, NY, USA, 1962. [Google Scholar]
- Gill, G.W.; Rhine, S. Skeletal Attribution of Race: Methods for Forensic Anthropology; Maxwell Museum of Anthropology: Albuquerque, NM, USA, 1990. [Google Scholar]
- Sardi, M.L.; Rozzi, F.R.; Dahinten, S.L.; Pucciarelli, H.M. Amerindians: Testing the hypothesis about their homogeneity. Comptes Rendus Palevol 2004, 3, 403–409. [Google Scholar] [CrossRef]
- Cardini, A.; O’Higgins, P.; Rohlf, F.J. Seeing Distinct Groups Where There are None: Spurious Patterns from Between-Group PCA. Evol. Biol. 2019, 46, 303–316. [Google Scholar] [CrossRef]
- Rolf, F.J. Why Clusters and Other Patterns Can Seem to be Found in Analyses of High-Dimensional Data. Evol. Biol. 2021, 48, 1–16. [Google Scholar] [CrossRef]
- Ross, A.H.; Williams, S.E. Testin Repeatability and Error of Coordinate Landmark Data Acquired from Crania. J. Forensic Sci. 2008, 53, 782–785. [Google Scholar] [CrossRef]
- Stephen, A.J.; Wegscheider, P.K.; Nelson, A.J.; Dickey, J.P. Quantifying the precision and accuracy of the MicroScribe G2X three-dimensional digitizer. Digit. Appl. Archaeol. Cult. Herit. 2015, 2, 28–33. [Google Scholar] [CrossRef]
- Slice, D.E.; Ross, A.H. 3D-ID: Geometric Morphometric Classification of Crania for Forensic Scientists; NCJRS: Washington, DC, USA, 2009. [Google Scholar]
- Slice, D.E. Morpheus et al. Java Edition. Available online: http:/morphlab.sc.fsu.edu/ (accessed on 28 June 2021).
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Kendall, D.G. The diffusion of shape. Adv. Appl. Prob. 1977, 9, 428–430. [Google Scholar] [CrossRef]
- Zelditch, M.L.; Swiderski, D.L.; Sheets, H.D. Geometric Morphometrics for Biologists: A Primer; Academic Press: Salt Lake City, UT, USA, 2012. [Google Scholar]
- JMP, Version 14.1 ed; SAS Institute Inc.: Cary, NC, USA, 1989.
- Sokal, R.R. The continental population structure of Europe. Annu. Rev. Anthropol. 1991, 20, 119–140. [Google Scholar] [CrossRef]
- Pietrusewsky, M. Traditional morphometrics and biological distance methods and an example. In Biological Anthropology of the Human Skeleton, 3rd ed.; Katzenberg, M.A., Grauer, A.L., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2019; pp. 547–591. [Google Scholar]
- Sokal, R.R.; Uytterschaut, H. Cranial variation in European populations: A spatial autocorrelation study at three time periods. Am. J. Phys. Anthropol. 1987, 74, 21–38. [Google Scholar] [CrossRef]
- Anselin, L.; Syabri, I.; Kho, Y. GeoDa: An Introduction to Spatial Data Analysis. In Handbook of Applied SpatialAanalysis; Springer: Berlin/Heidelberg, Germany, 2010; pp. 73–89. [Google Scholar]
- Hefner, J.T.; Ousley, S.D. Statistical Classification Methods for Estimating Ancestry Using Morphoscopic Traits. J. Forensic Sci. 2014, 59, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Jantz, R.L.; Ousley, S.D. Fordisc 3.1: Computerized Forensic Discriminant Functions; The University of Tennessee: Knoxville, TN, USA, 2005. [Google Scholar]
- Navega, D.; Coelho, C.; Vicente, R.; Ferreira, M.T.; Wasterlain, S.; Cunha, E. AncesTrees: Ancestry estimation with randomized decision trees. Int. J. Leg. Med. 2015, 129, 1145–1153. [Google Scholar] [CrossRef]
- Ross, A.H.; Ubelaker, D.H. Complex Nature of Hominin Dispersals: Ecogeographical and Climatic Evidence for Pre-Contact Craniofacial Variation. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez, O.J. La población del Istmo de Panamá: Estudio de geohistoria; Ediciones de Cultura Hispánica: Madrid, Spain, 1998. [Google Scholar]
- Salzano, F.M.; Bortolini, M.C. The Evolution and Genetics of Latin American Populations; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Price, T.D.; Tiesler, V.; Zabala, P.; Coppa, A.; Freiwald, C.; Schroeder, H.; Cucina, A. (Ahead of Print). Home Is the Sailor: Investigating the Origins of the Inhabitants of La Isabela, the First European Settlement in the New World. Curr. Anthropol. 2020, 61, 5. [Google Scholar] [CrossRef]
- Ross, A.H.; Keegan, W.F.; Pateman, M.; Young, C.B. Faces Divulge the Origins of Caribbean Prehistoric Inhabitants. Sci. Rep. 2020, 10, 147–149. [Google Scholar] [CrossRef]
- Panama Canal Commission. The Panama Canal Review; October 1 Issue; 1979. Available online: https://ufdcimages.uflib.ufl.edu/UF/00/09/73/66/00059/UF00097366_00059.pdf (accessed on 28 June 2021).
- Chiappetta, R.F.; Treleaven, T.; Nixon, E.; Smith, J.D. History and expansion of the Panama Canal. Fragblast 1998, 2, 313–340. [Google Scholar] [CrossRef]
- McCullough, D. The Path Between the Seas: The Creation of the Panama Canal; Simon & Schuster: New York, NY, USA, 1977; pp. 1870–1914. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N | Provenance | Latitude | Longitude |
|---|---|---|---|---|
| Chile | ♀ = 34 ♂ = 37 | Juan Munizaga Collection, Universidad de Chile, Santiago, Chile | −33.45 | −70.67 |
| Colombia | ♀ = 11 ♂ = 53 | Antioquia modern skeletal collection, Escuela Nacional de Criminalística, Medellín, Colombia | 6.230833 | −75.5906 |
| Cuba | 19 | Cemetery Collection, Museo de Montane, Havana, Cuba | 23.11359 | −82.3666 |
| Enslaved Africans | 25 | Morton Collection, University of Pennsylvania, US | −8.83833 | 13.23444 |
| Guatemala | ♂ = 71 | Provided by Kate Spradley | 14.62843 | −90.5227 |
| Puerto Rico | ♂ = 5 | University of Rio Piedras, Puerto Rico | 18.46633 | −66.1057 |
| Panama | 10 | Insituto de Medicina Legal, Panama | 8.983333 | −79.5167 |
| Peru | 7 | C.A. Pound Human ID Lab, University of Florida, US | −12.0464 | −77.0428 |
| Spain | ♀ = 58 ♂ = 67 | Oloriz Collection, Madrid, Spain | 40.41678 | −3.70379 |
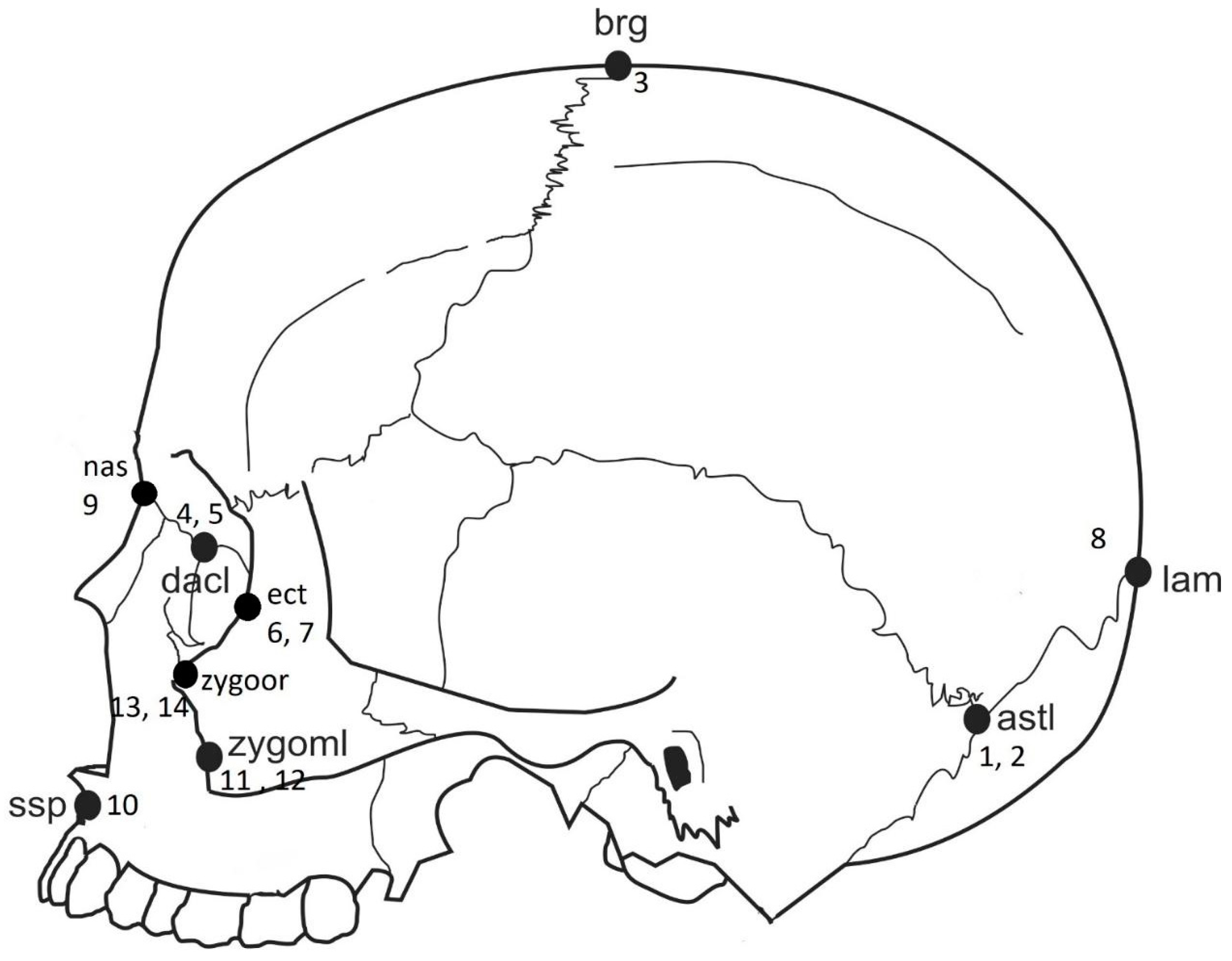
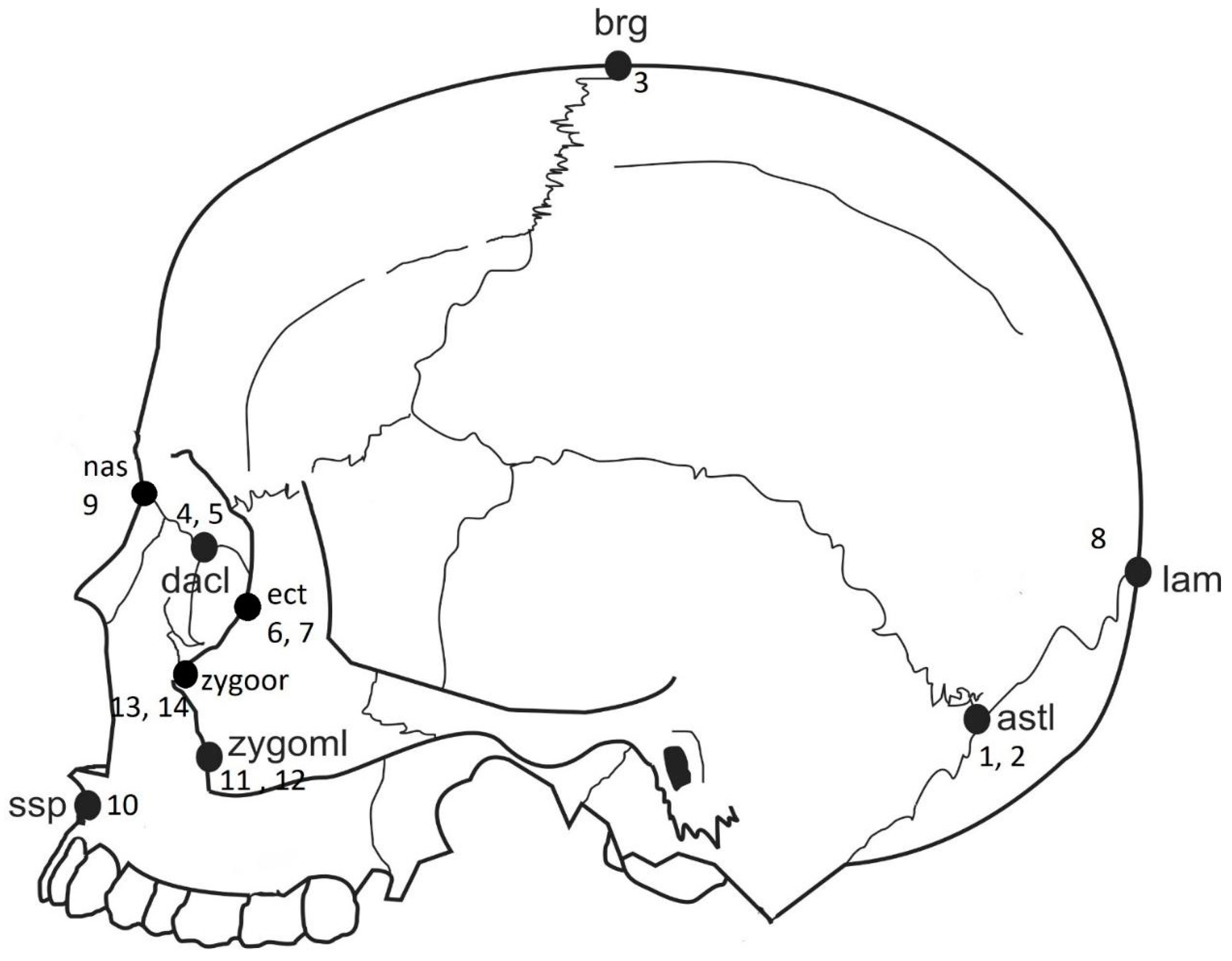
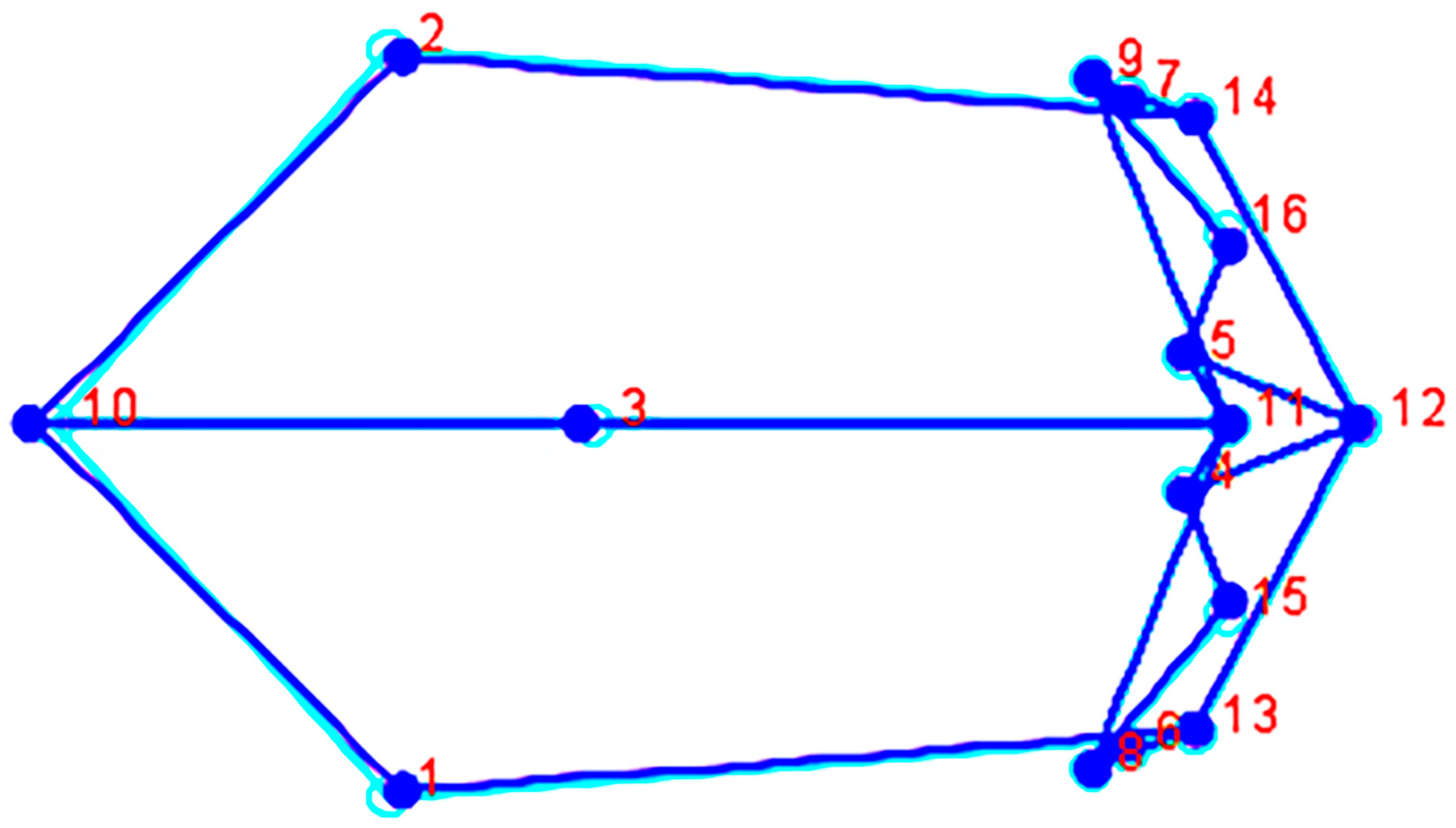
| Landmark Number | Anatomical Landmarks |
|---|---|
| 1, 2 | Asterion, bilateral |
| 3 | Bregma |
| 4, 5 | Dacryon, bilateral |
| 6, 7 | Ectoconchion, bilateral |
| 8,9 | Frontomalare temporale, bilateral |
| 10 | Lambda |
| 11 | Nasion |
| 12 | Subspinale |
| 13, 14 | Zygomaxillare, bilateral |
| 15, 16 | Zygoorbitale, bilateral |
| Moran’s I | Observed | Expected | Std Dev | Z | PR > Z |
|---|---|---|---|---|---|
| PC1 | 0.0695 | −0.0025 | 0.0022 | 32.5 | 0.001 |
| CS | 0.0027 | −0.0025 | 0.0026 | 2.0 | 0.027 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ross, A.H.; Williams, S.E. Ancestry Studies in Forensic Anthropology: Back on the Frontier of Racism. Biology 2021, 10, 602. https://doi.org/10.3390/biology10070602
Ross AH, Williams SE. Ancestry Studies in Forensic Anthropology: Back on the Frontier of Racism. Biology. 2021; 10(7):602. https://doi.org/10.3390/biology10070602
Chicago/Turabian StyleRoss, Ann H., and Shanna E. Williams. 2021. "Ancestry Studies in Forensic Anthropology: Back on the Frontier of Racism" Biology 10, no. 7: 602. https://doi.org/10.3390/biology10070602
APA StyleRoss, A. H., & Williams, S. E. (2021). Ancestry Studies in Forensic Anthropology: Back on the Frontier of Racism. Biology, 10(7), 602. https://doi.org/10.3390/biology10070602