
Rehydration Process in Rustyback Fern (Asplenium ceterach L.): Profiling of Volatile Organic Compounds


 , , and
, , and
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Plant Material
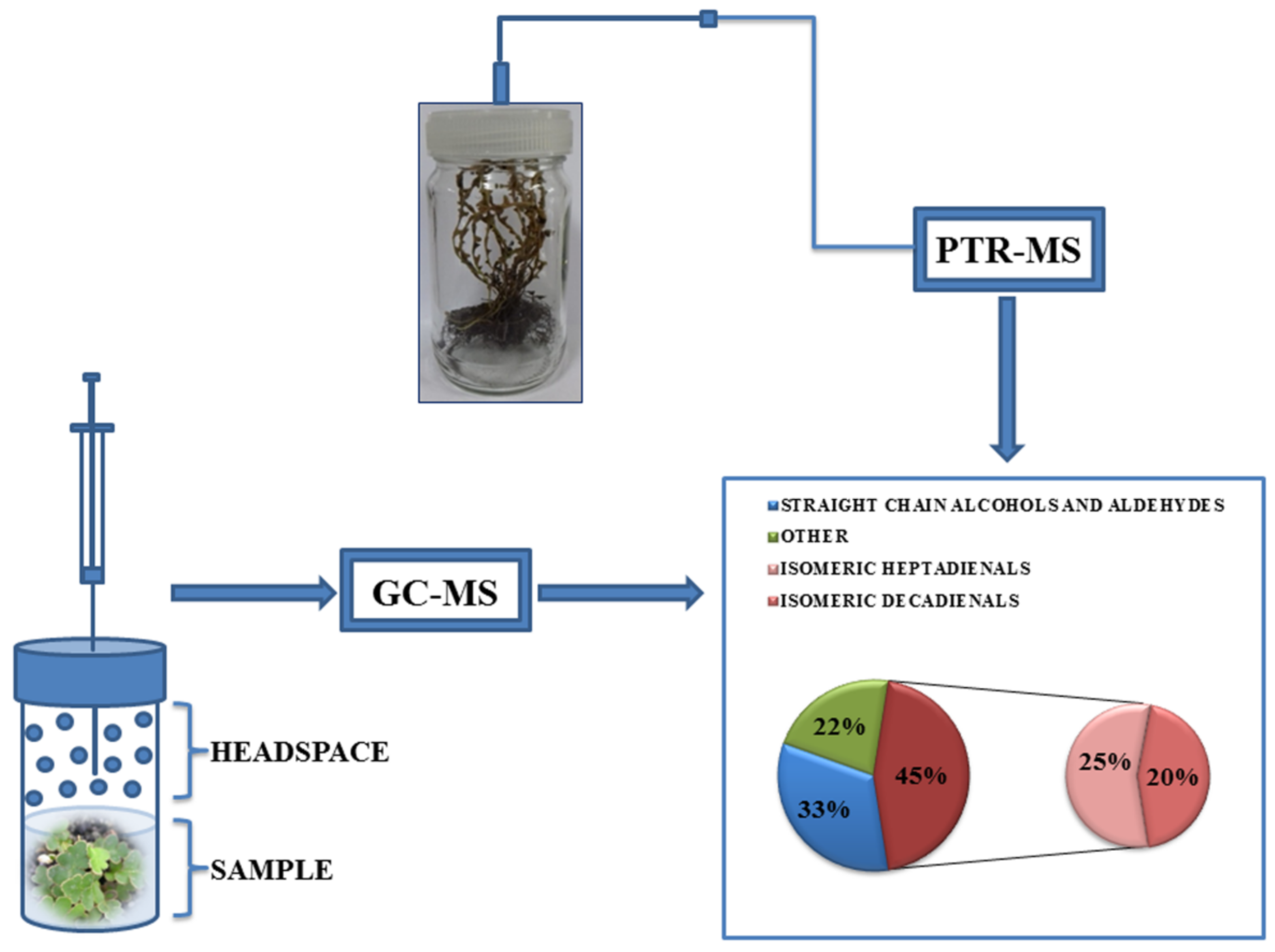
2.2. Headspace GC–MS Analysis of Volatiles
2.3. Online PTR–MS Measurements of Volatiles
2.4. Determination of Relative Water Content
2.5. Measurement of Lipid Peroxidation
2.6. Statistical Analysis
3. Results and Discussion
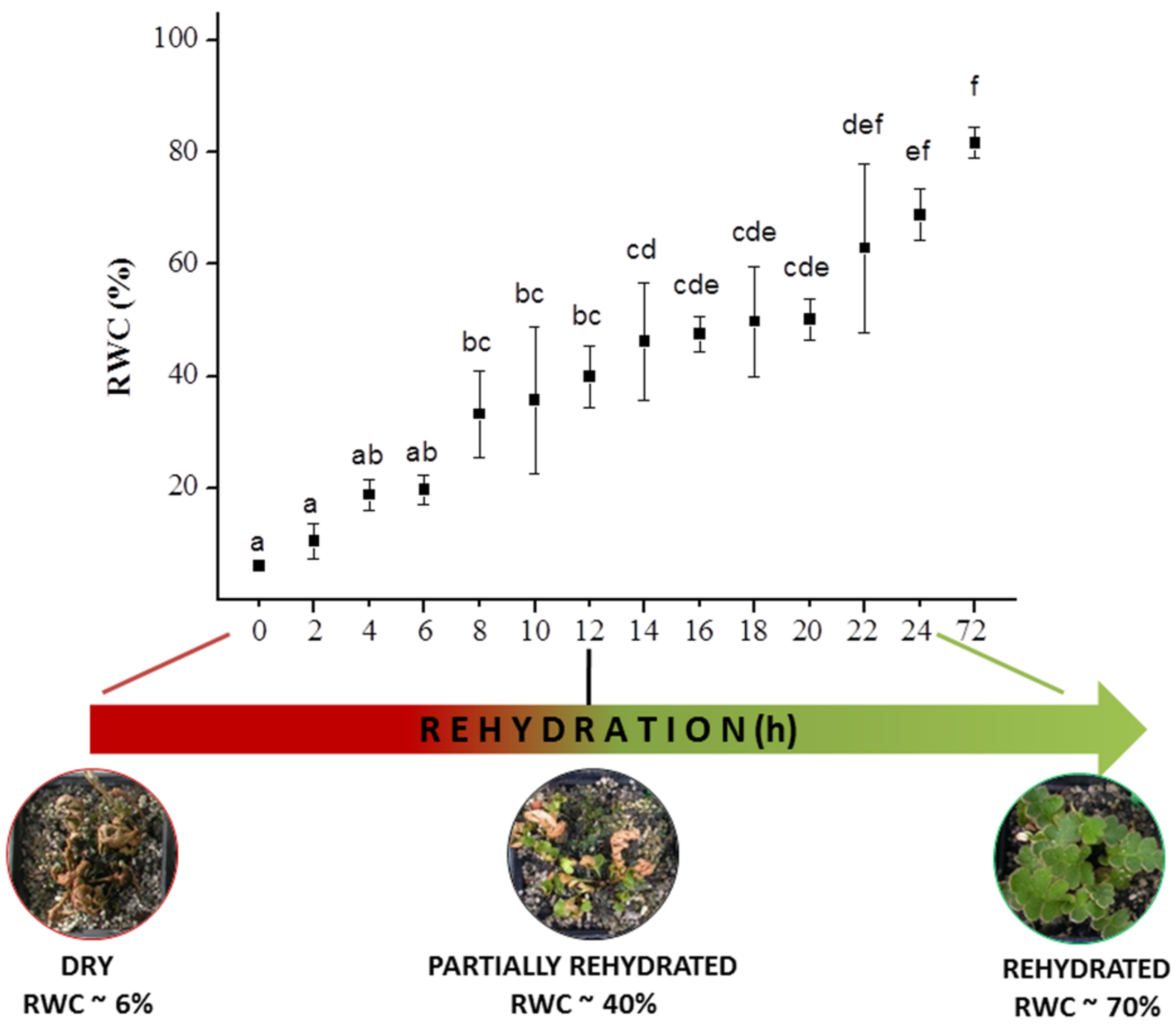
3.1. Relative Water Content Analysis
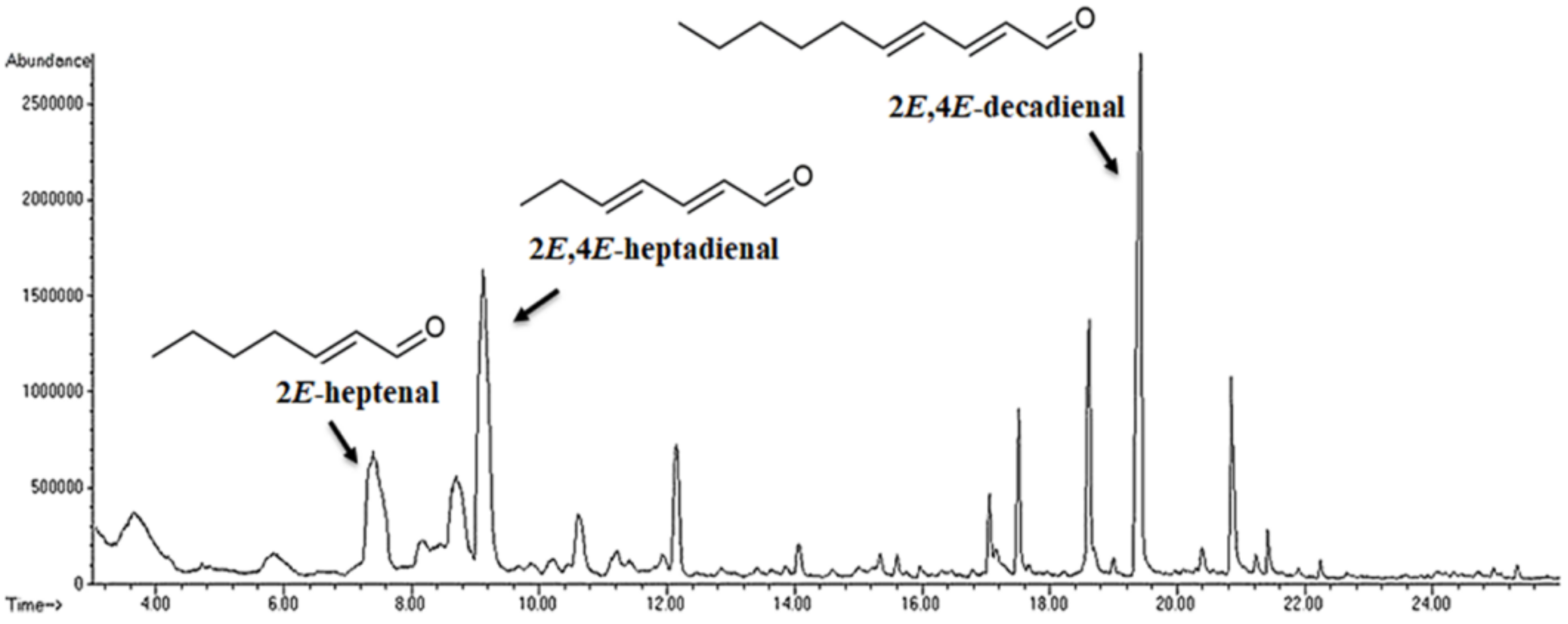
3.2. Emission of Volatiles during Rehydration
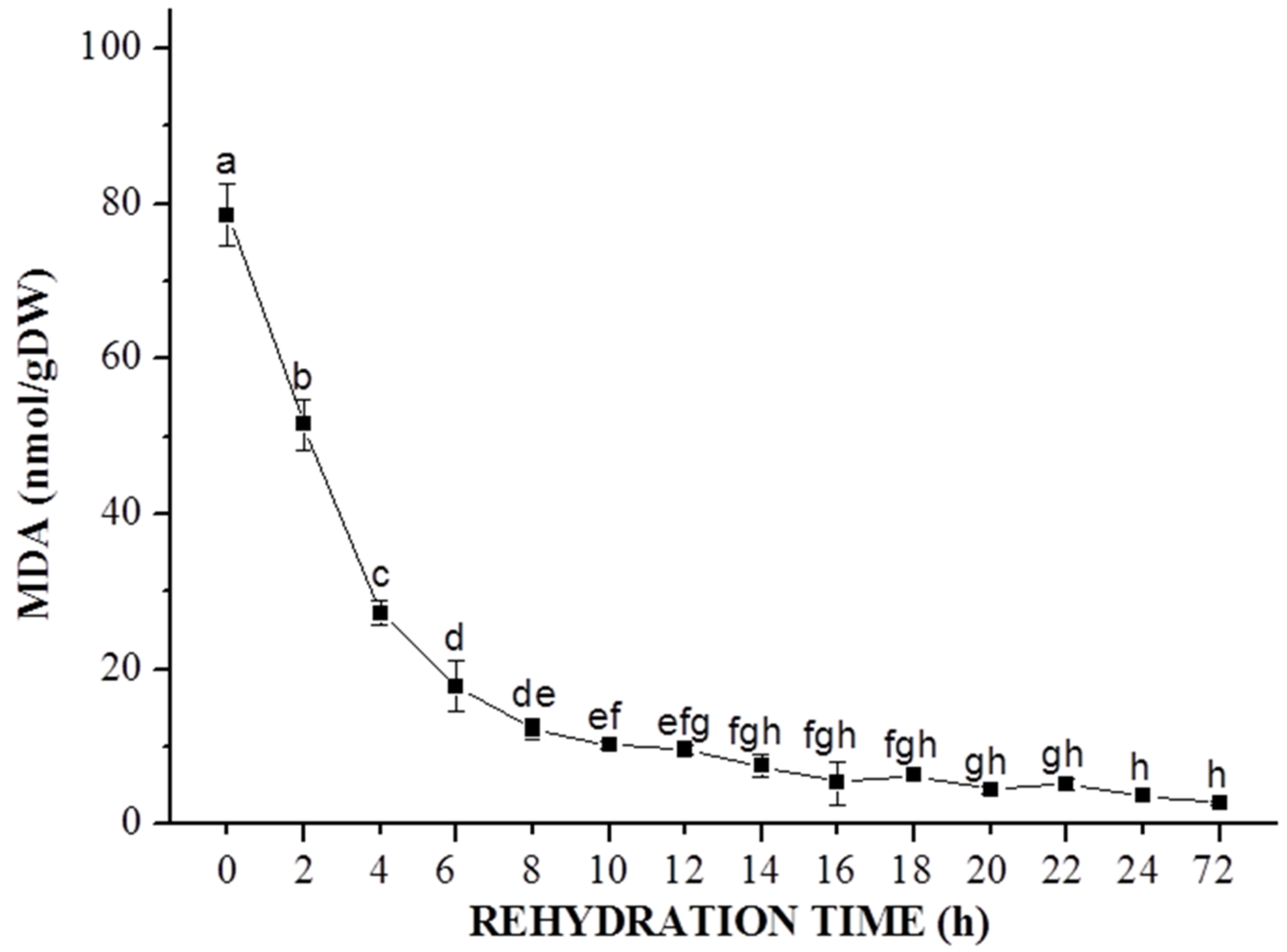
3.3. Estimation of Lipid Peroxidase Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bernacchia, G.; Salamini, F.; Bartels, D. Molecular characterization of the rehydration process in the resurrection plant Craterostigma plantagineum. Plant Physiol. 1996, 111, 1043–1050. [Google Scholar] [CrossRef] [Green Version]
- Georgieva, K.; Mihailova, G.; Velitchkova, M.; Popova, A. Recovery of photosynthetic activity of resurrection plant Haberlea rhodopensis from drought- and freezing-induced desiccation. Photosynthetica 2020, 58, 911–921. [Google Scholar] [CrossRef]
- Beckett, M.; Loreto, F.; Velikova, V.; Brunetti, C.; Di Fernando, M.; Tattini, M.; Calfapietra, C.; Farrant, J.M. Photosynthetic limitations and volatile and non-volatile isoprenoids in the poikilochlorophyllous resurrection plant Xerophyta humilis during dehydration and rehydration. Plant Cell Environ. 2012, 35, 2061–2074. [Google Scholar] [CrossRef] [PubMed]
- Farrant, J.M.; Brandt, W.; Lindsey, G.G. An overview of mechanisms of desiccation tolerance in selected angiosperm resurrection plants. Plant Stress 2007, 1, 72–84. [Google Scholar]
- Oliver, M.J.; Farrant, J.M.; Hilhorst, H.W.; Mundree, S.; Williams, B.; Bewley, J.D. Desiccation tolerance: Avoiding cellular damage during drying and rehydration. Annu. Rev. Plant Biol. 2020, 71, 435–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenkranz, M.; Schnitzler, J.-P. Plant Volatiles. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2016. [Google Scholar] [CrossRef]
- Jardine, K.; Abrell, L.; Kurc, S.A.; Huxman, T.; Ortega, J.; Guenther, A. Volatile organic compound emissions from Larrea tridentata (creosotebush). Atmos. Chem. Phys. 2010, 10, 12191–12206. [Google Scholar] [CrossRef] [Green Version]
- Spiteller, G. The relationship between changes in the cell wall, lipid peroxidation, proliferation, senescence and cell death. Physiol. Plant. 2003, 119, 5–18. [Google Scholar] [CrossRef]
- Foyer, C. Oxygen metabolism and electron transport in photosynthesis. In The Molecular Biology of Free Radical Scavenging System; Scandalios, J., Ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1997; pp. 587–621. [Google Scholar]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant. Biosyst. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Cooper, K.; Farrant, J.M. Recovery of the resurrection plant Craterostigmawilmsii fromdesiccation: Protection versus repair. J. Exp. Bot. 2002, 53, 1805–1813. [Google Scholar] [CrossRef] [Green Version]
- Schwab, K.B.; Schreiber, U.; Heber, U. Response of photosynthesis and respiration of resurrection plants to desiccation and rehydration. Planta 1989, 177, 217–227. [Google Scholar] [CrossRef]
- Froissard, D.; Rapior, S.; Bessière, J.-M.; Buatois, B.; Fruchier, A. Asplenioideae Species as a Reservoir of Volatile Organic Compounds with Potential Therapeutic Properties. Nat. Prod. Commun. 2015, 10, 1079–1083. [Google Scholar] [CrossRef] [Green Version]
- Hammami, S.; Snène, A.; El Mokni, R.; Faidi, K.; Falconieri, D.; Dhaouadi, H.; Piras, A.; Mighri, Z.; Porcedda, S. Essential Oil Constituents and Antioxidant Activity of Asplenium Ferns. J. Chromatogr. Sci. 2016, 54, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Živković, S.; Skorić, M.; Šiler, B.; Dmitrović, S.; Filipović, B.; Nikolić, T.; Mišić, D. Phytochemical characterization and antioxidant potential of rustyback fern (Asplenium ceterach L.). Lek. Sirovine 2017, 37, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Tomou, E.-M.; Skaltsa, H. Phytochemical Investigation of the Fern Asplenium ceterach (Aspleniaceae). Nat. Prod. Commun. 2018, 13, 849–850. [Google Scholar]
- Živković, S.; Milutinović, M.; Maksimović, V.; Ćirić, A.; Ivanov, M.; Božunović, J.; Banjanac, T.; Mišić, D. Antioxidant and antimicrobial activity of two Asplenium species. S. Afr. J. Bot. 2020, 132, 180–187. [Google Scholar] [CrossRef]
- Adams, R. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Lindinger, W.; Hansel, A.; Jordan, A. On-line monitoring of volatile organic compounds at pptV levels by means of proton-transfer-reaction mass spectrometry (PTR-MS): Medical applications, food control, and environmental research. Int. J. Mass Spectrom. Ion Process. 1998, 173, 191–241. [Google Scholar] [CrossRef]
- Ellis, A.M.; Mayhew, C.A. Proton Transfer Reaction Mass Spectrometry: Principles and Applications; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Hodges, M.D.; De Long, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Bewley, J.D.; Krochko, J.E. Desiccation-tolerance. In Encyclopaedia of Plant Physiology, Physiological Ecology II; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1982; Volume 12/B, pp. 325–378. [Google Scholar]
- Farrant, J.M.; Cooper, K.; Kruger, L.A.; Sherwin, H.W. The effect of drying rate on the survival of three desiccation-tolerant angiosperm species. Ann. Bot. 1999, 84, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Farrant, J.M. Mechanisms of Desiccation Tolerance in Angiosperm Resurrection Plants. In Plant Desiccation Tolerance; Jenks, M., Wood, A., Eds.; Blackwell Publishing: Wallingford, IA, USA, 2007; pp. 51–90. [Google Scholar]
- Lazarević, M.; Rakić, T.; Šinžar-Sekulić, J. Morphological differences between the flowers of Ramonda serbica, R. nathaliae and their hybrid. Bot. Serb. 2014, 38, 91–98. [Google Scholar]
- Rakić, T.; Lazarević, M.; Jovanović, Ž.S.; Radović, S.; Šiljak-Yakovlev, S.; Stevanović, B.; Stevanović, V. Resurrection plants of the genus Ramonda: Prospective survival strategies–unlock further capacity of adaptation, or embark on the path of evolution? Front. Plant Sci. 2014, 4, 550. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.; Sun, Y.; Luo, S.; Zhang, C.; Zhou, H.; Lin, H. Efficient modulation of photosynthetic apparatus confers desiccation tolerance in the resurrection plant Boea hygrometrica. Plant Cell Physiol. 2017, 58, 1976–1990. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Noel, J.P.; Dudareva, N. Biosynthesis of Plant Volatiles: Nature’s Diversity and Ingenuity. Science 2006, 311, 808–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Loreto, F.; Schnitzler, J. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Tholl, D.; Boland, W.; Hansel, A.; Loreto, F.; Röse, U.S.R.; Schnitzler, J.P. Practical approaches to plant volatile analysis. Plant J. 2006, 45, 540–560. [Google Scholar] [CrossRef]
- Majchrzak, T.; Wojnowski, W.; Lubinska-Szczygeł, M.; Różańska, A.; Namieśnik, J.; Dymerski, T. PTR-MS and GC-MS as complementary techniques for analysis of volatiles: A tutorial review. Anal. Chim. Acta 2018, 1035, 1–13. [Google Scholar] [CrossRef]
- Rapior, S.; Breheret, S.; Talou, T.; Pélissier, Y.; Bessière, J.M. The anise-like odor of Clitocybeodora, Lentinelluscochleatus and Agaricusessettei. Mycologia 2002, 94, 373–376. [Google Scholar] [CrossRef]
- Rapior, S.; Breheret, S.; Talou, T.; Pélissier, Y.; Milhau, M.; Bessière, J.M. Volatile components of fresh Agrocybe aegerita and Tricholoma sulfureum. Cryptogam. Mycol. 1998, 19, 15–23. [Google Scholar]
- Fons, F.; Froissard, D.; Bessière, J.M.; Buatois, B.; Rapior, S. Biodiversity of volatile organic compounds from five French ferns. Nat. Prod. Commun. 2010, 5, 1655–1658. [Google Scholar] [CrossRef] [Green Version]
- Froissard, D.; Fons, F.; Bessière, J.M.; Buatois, B.; Rapior, S. Volatiles of French ferns and “fougère” scent in perfumery. Nat. Prod. Commun. 2011, 6, 1723–1726. [Google Scholar] [CrossRef] [Green Version]
- Fons, F.; Rapior, S.; Gargadennec, A.; Andary, C.; Bessière, J.M. Volatile components of Plantago lanceolata (Plantaginaceae). Acta Bot. Gall. 1998, 145, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Ho, H.Y.; Kou, R.; Tseng, H.K. Semiochemicals from the predatory stink bug Eocanthecona furcellata (Wolff): Components of metathoracic gland, dorsal abdominal gland, and sternal gland secretions. J. Chem. Ecol. 2003, 29, 2101–2114. [Google Scholar] [CrossRef]
- Fons, F.; Froissard, D.; Bessière, J.-M.; Fruchier, A.; Buatois, B.; Rapior, S. Volatile Composition of Six Horsetails: Prospects and Perspectives. Nat. Prod. Commun. 2013, 8, 509–512. [Google Scholar] [CrossRef]
- Watson, S.B. Aquatic taste and odor: A primary signal of drinking-water integrity. J. Toxicol. Environ. Health Part A 2004, 67, 1779–1795. [Google Scholar] [CrossRef]
- Zhao, Y.; Jianwei, Y.; Ming, S.; Wei, A.; Min, Y. A fishy odor episode in a north China reservoir: Occurrence, origin, and possible odor causing compounds. J. Environ. Sci. 2013, 25, 2361–2366. [Google Scholar] [CrossRef]
- Grosch, W. Lipid Degradation Products and Flavour. In Food Flavours; Morton, I.D., Mac Leod, A.J., Eds.; Elsevier: Amsterdam, The Netherlands, 1982; pp. 325–398. [Google Scholar]
- Brunke, E.J.; Hammerschmidt, F.J.; Schmaus, G.; Akgul, A. The essential oil of Rhuscoriaria L. fruits. Flavour Fragr. J. 1993, 8, 209–214. [Google Scholar] [CrossRef]
- Andreou, A.; Brodhun, F.; Feussner, I. Biosynthesis of oxylipins in nonmammals. Prog. Lipid Res. 2009, 48, 148–170. [Google Scholar] [CrossRef]
- Ruocco, N.; Albarano, L.; Esposito, R.; Zupo, V.; Costantini, M.; Ianora, A. Multiple Roles of Diatom-Derived Oxylipins within Marine Environments and Their Potential Biotechnological Applications. Mar. Drugs 2020, 18, 342. [Google Scholar] [CrossRef]
- Bhattacharjee, S. The language of reactive oxygen species signaling in plants. J. Bot. 2012, 2012, 985298. [Google Scholar] [CrossRef] [Green Version]
- Hansel, A.; Jordan, A.; Holzinger, R.; Prazeller, P.; Vogel, W.; Lindinger, W. Proton transfer reaction mass spectrometry: On-line trace gas analysis at the ppb level. Int. J. Mass Spectrom. Ion Process 1995, 149–150, 609–619. [Google Scholar] [CrossRef]
- Lindinger, W.; Jordan, A. Proton-transfer-reaction mass spectrometry (PTR–MS): On-line monitoring of volatile organic compounds at pptV levels. Chem. Soc. Rev. 1998, 27, 347. [Google Scholar] [CrossRef]
- Lacko, M.; Piel, F.; Mauracher, A.; Spanel, P. Chemical ionization of glyoxal and formaldehyde with H3O+ ions using SIFT-MS under variable system humidity. Phys. Chem. Chem. Phys. 2020, 22, 10170–10178. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Min, D.B. Analysis of Volatile Compounds from Chlorophyll Photosensitized Linoleic Acid by Headspace Solid-phase Microextraction (HS-SPME). Food Sci. Biotechnol. 2010, 19, 611–616. [Google Scholar] [CrossRef]
- Kanavouras, A.; Hernandez-Münoz, P.; Coutelieris, F.; Selke, S. Oxidation-Derived Flavor Compounds as Quality Indicators for Packaged Olive Oil. J. Am. Oil Chem. Soc. 2004, 81, 251–257. [Google Scholar] [CrossRef]
- Kiralan, M.; Çalik, G.; Kiralan, S.; Ramadan, M.F. Monitoring stability and volatile oxidation compounds of cold-pressed flax seed, grape seed and black cumin seed oils upon photo-oxidation. Grasas Aceites 2019, 70, e295. [Google Scholar] [CrossRef]
- Potter, T.L.; Fagerson, I.S. Composition of coriander leaf volatiles. J. Agric. Food Chem. 1990, 38, 2054–2056. [Google Scholar] [CrossRef]
- Gogus, F.; Ozel, M.Z.; Keskin, H.; KocakYanık, D.; Lewis, A.C. Volatiles of fresh and commercial sweet red pepper pastes: Processing methods and microwave assisted extraction. Int. J. Food Prop. 2015, 18, 1625–1634. [Google Scholar] [CrossRef]
- Zhang, J.-H.; Sun, H.; Chen, S.; Zeng, L.; Wang, T.-T. Antifungal activity, mechanism studies on α-Phellandrene and Nonanal against Penicilliumcyclopium. Bot. Stud. 2017, 58, 13. [Google Scholar] [CrossRef] [Green Version]
- Eberl, F.; Hammerbacher, A.; Gershenzon, J.; Unsicker, S.B. Leaf rust infection reduces herbivore-induced volatile emission in black poplar and attracts a generalist herbivore. New Phytol. 2018, 220, 760–772. [Google Scholar] [CrossRef] [Green Version]
- Kigathi, R.N.; Weisser, W.; Reichelt, M.; Gershenzon, J.; Unsicker, S.B. Plant volatile emission depends on the species composition of the neighboring plant community. BMC Plant Biol. 2019, 19, 58. [Google Scholar] [CrossRef]
- Jelen, H.H.; Krawczyk, J.; Larsen, T.O.; Jarosz, A.; Gołebniak, B. Main compounds responsible for off-odour of strawberries infected by Phytophthoracactorum. Lett. Appl. Microbiol. 2005, 40, 255–259. [Google Scholar] [CrossRef]
- Alché, J.d.D. A concise appraisal of lipid oxidation and lipoxidation in higher plants. Redox Biol. 2019, 23, 101136. [Google Scholar] [CrossRef]
- Kong, W.; Liu, F.; Zhang, C.; Zhang, J.; Feng, H. Non-destructive determination of Malondialdehyde (MDA) distribution in oilseed rape leaves by laboratory scale NIR hyperspectral imaging. Sci. Rep. 2016, 6, 35393. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Bravo, L.A.; Copolovici, L. Changes in photosynthetic rate and stress volatile emissions through desiccation-rehydration cycles in desiccation-tolerant epiphytic filmy ferns (Hymenophyllaceae). Plant Cell Environ. 2018, 41, 1605–1617. [Google Scholar] [CrossRef]
- John, S.P.; Hasenstein, K.H. Biochemical responses of the desiccation-tolerant resurrection fern Pleopeltis polypodioides to dehydration and rehydration. J. Plant Physiol. 2018, 228, 12–18. [Google Scholar] [CrossRef]
- NeeragundaShivaraj, Y.; Plancot, B.; Ramdani, Y.; Gügi, B.; Kambalagere, Y.; Jogaiah, S.; Driouich, A.; RamasandraGovind, S. Physiological and biochemical responses involved in vegetative desiccation tolerance of resurrection plant Selaginella brachystachya. 3 Biotech 2021, 11, 135. [Google Scholar] [CrossRef]
- Wang, Y.L.; Gao, S.S.; He, X.Y.; Li, Y.; Li, P.Y.; Zhang, Y.; Chen, W. Growth, secondary metabolites and enzyme activity responses of two edible fern species to drought stress and rehydration in Northeast China. Agronomy 2019, 9, 137. [Google Scholar] [CrossRef] [Green Version]
- Pandey, V.; Ranjan, S.; Deeba, F.; Pandey, A.K.; Singh, R.; Shirke, P.A.; Pathre, U.V. Desiccation-induced physiological and biochemical changes in resurrection plant, Selaginellabryopteris. J. Plant Physiol. 2010, 167, 1351–1359. [Google Scholar] [CrossRef]
- Djilianov, D.; Ivanov, S.; Moyankova, D.; Miteva, L.; Kirova, E.; Alexieva, V.; Joudi, M.; Peshev, D.; Van den Ende, W. Sugar ratios, glutathione redox status and phenols in the resurrection species Haberlea rhodopensis and the closely related non-resurrection species Chirita eberhardtii. Plant Biol. 2011, 13, 767–776. [Google Scholar] [CrossRef]
- Li, A.; Wang, D.; Yu, B.; Yu, X.; Li, W. Maintenance or collapse: Responses of extraplastidic membrane lipid composition to desiccation in the resurrection plant Paraisometrum mileense. PLoS ONE 2014, 9, e103430. [Google Scholar] [CrossRef] [Green Version]
- Banupriya, T.G.; Ramyashree, C.; Akash, D.; Yathish, N.S.; Sharthchandra, R.G. Studies on the mechanism of desiccation tolerance in the resurrection fern Adiantumraddianum. J. Appl. Biol. Biotechnol. 2020, 8, 6–14. [Google Scholar]
- Lubaina, A.S.; Brijithlal, N.D.; Murugan, K. Unraveling desiccation and rehydration tolerance mechanism in the fern, Adiantumlatifolium. Biosci. Biotechnol. Res. Commun. 2016, 9, 672–679. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Constituent | KIE | KIL | tR | % (m/m) 1 |
|---|---|---|---|---|---|
| 1 | Hex-1-ene-3-ol | 829.5 | 832 | 3.66 | 4.48 |
| 2 | n-Heptanal | 903.8 | 901 | 5.88 | 2.32 |
| 3 | (2E)-Heptenal | 954.6 | 947 | 7.40 | 11.54 |
| 4 | 1-Octen-3-ol | 980.0 | 974 | 8.16 | 2.37 |
| 5 | 3-Octanone | 985.1 | 979 | 8.31 | 0.90 |
| 6 | 2-Pentyl furan | 988.8 | 984 | 8.42 | 1.13 |
| 7 | (2E,4Z)-Heptadienal | 1000.1 | 988 | 8.76 | 8.43 |
| 8 | (2E,4E)-Heptadienal | 1010.7 | 1005 | 9.13 | 17.66 |
| 9 | (2E)-Octen-1-al | 1043.1 | 1034 | 10.23 | 0.96 |
| 10 | Oct-3-ene-1-ol | 1054.2 | 1044 | 10.61 | 3.05 |
| 11 | n-Octanol | 1071.2 | 1063 | 11.19 | 1.32 |
| 12 | (2E)-Octen-1-ol | 1078.5 | 1067 | 11.44 | 0.71 |
| 13 | n-Undecane | 1092.4 | 1100 | 11.93 | 1.40 |
| 14 | n-Nonanal | 1099.1 | 1100 | 12.15 | 4.24 |
| 15 | (2E)-Nonen-1-al | 1154.2 | 1157 | 14.07 | 0.89 |
| 16 | n-Dodecane | 1190.9 | 1200 | 15.35 | 0.58 |
| 17 | n-Decanal | 1198.3 | 1201 | 15.61 | 0.41 |
| 18 | g-Octalactone | 1241.2 | 1250 | 17.05 | 1.55 |
| 19 | n.i. | 1244.2 | / | 17.15 | 1.06 |
| 20 | (2E)-Decenal | 1254.8 | 1260 | 17.51 | 2.81 |
| 21 | (2E,4Z)-Decadienal | 1287.4 | 1292 | 18.62 | 5.68 |
| 22 | n-Undecanal | 1299.0 | 1305 | 19.00 | 0.42 |
| 23 | (2E,4E)-Decadienal | 1311.6 | 1315 | 19.42 | 15.03 |
| 24 | n.i. | 1342.1 | / | 20.40 | 0.70 |
| 25 | (2E)-Undecenal | 1355.8 | 1357 | 20.85 | 4.51 |
| 26 | n.i. | 1367.8 | / | 21.23 | 0.39 |
| 27 | n.i. | 1373.3 | / | 21.41 | 0.79 |
| 28 | n-Dodecanal | 1398.8 | 1408 | 22.24 | 0.30 |
| 29 | n-Tridecanal | 1499.5 | 1509 | 25.33 | 0.21 |
| 30 | (6Z)-Pentadecen-2-one | 1655.5 | 1667 | 29.79 | 0.16 |
| 31 | (2E)-Tridecenol acetate | 1701.1 | 1703 | 31.07 | 0.18 |
| 32 | Hexadecanoic acid | 1965.8 | 1959 | 37.75 | 0.30 |
| 33 | n.i. | 2062.5 | / | 40.02 | 0.23 |
| 34 | Linoleic acid | 2146.9 | 2132 | 41.89 | 0.77 |
| 35 | n.i. | 2153.2 | / | 42.02 | 0.32 |
| 36 | n.i. | 2282.9 | / | 44.79 | 0.28 |
| 37 | n.i. | 2455.6 | / | 48.25 | 0.43 |
| 38 | n.i. | 2460.5 | / | 48.35 | 0.66 |
| 39 | Squalene | 2808.6 | 2814 | 54.74 | 0.31 |
| 40 | n.i. | 3062.7 | / | 59.96 | 0.52 |
| Number of detected constituents | 40 | 100% | |||
| Number of identified constituents | 30 | 94.61% | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Živković, S.; Skorić, M.; Ristić, M.; Filipović, B.; Milutinović, M.; Perišić, M.; Puač, N. Rehydration Process in Rustyback Fern (Asplenium ceterach L.): Profiling of Volatile Organic Compounds. Biology 2021, 10, 574. https://doi.org/10.3390/biology10070574
Živković S, Skorić M, Ristić M, Filipović B, Milutinović M, Perišić M, Puač N. Rehydration Process in Rustyback Fern (Asplenium ceterach L.): Profiling of Volatile Organic Compounds. Biology. 2021; 10(7):574. https://doi.org/10.3390/biology10070574
Chicago/Turabian StyleŽivković, Suzana, Marijana Skorić, Mihailo Ristić, Biljana Filipović, Milica Milutinović, Mirjana Perišić, and Nevena Puač. 2021. "Rehydration Process in Rustyback Fern (Asplenium ceterach L.): Profiling of Volatile Organic Compounds" Biology 10, no. 7: 574. https://doi.org/10.3390/biology10070574
APA StyleŽivković, S., Skorić, M., Ristić, M., Filipović, B., Milutinović, M., Perišić, M., & Puač, N. (2021). Rehydration Process in Rustyback Fern (Asplenium ceterach L.): Profiling of Volatile Organic Compounds. Biology, 10(7), 574. https://doi.org/10.3390/biology10070574