
Immuno-Electron and Confocal Laser Scanning Microscopy of the Glycocalyx


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Formalin Fixation
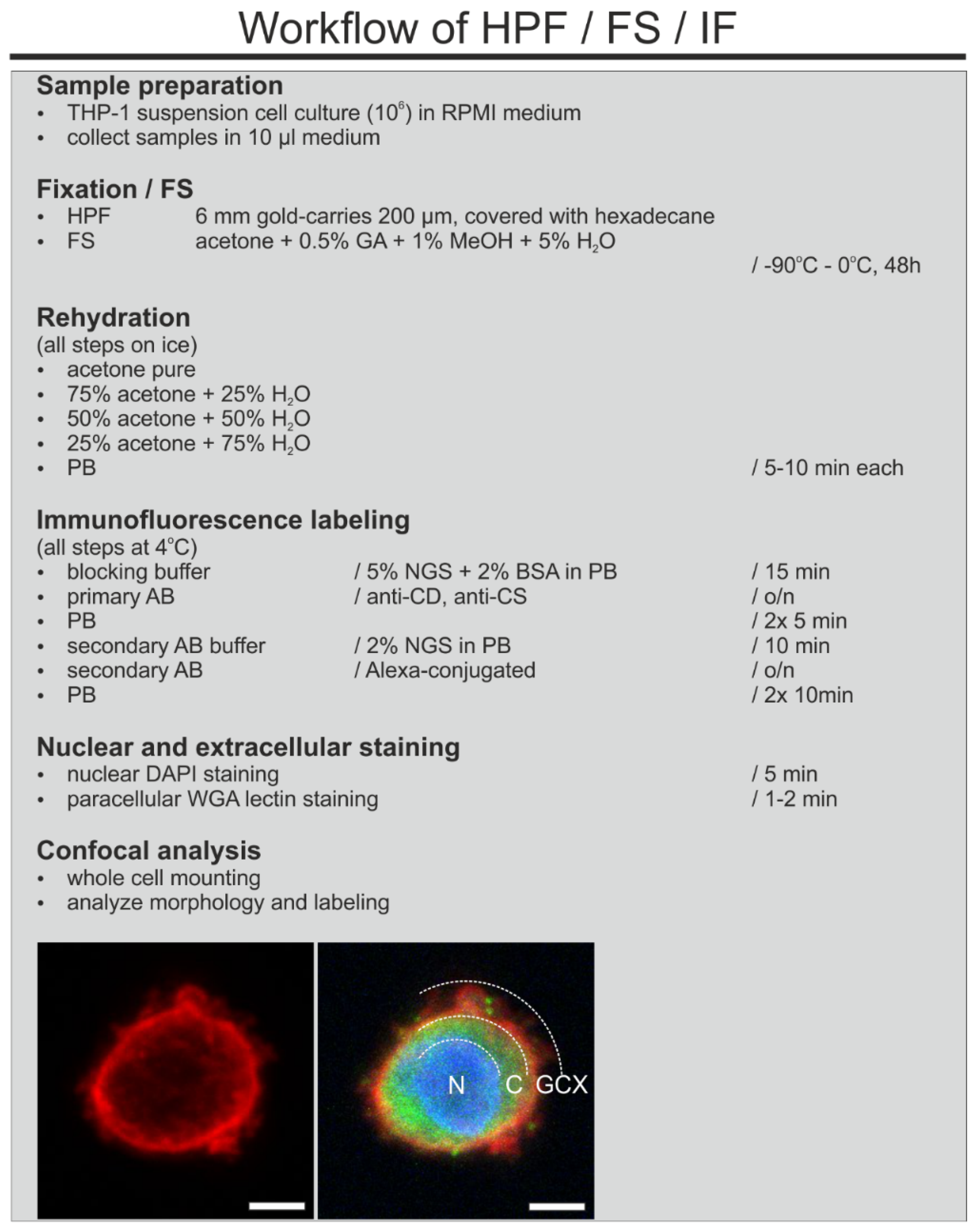
2.3. High Pressure Freezing
2.4. Freeze Substitution and Rehydration
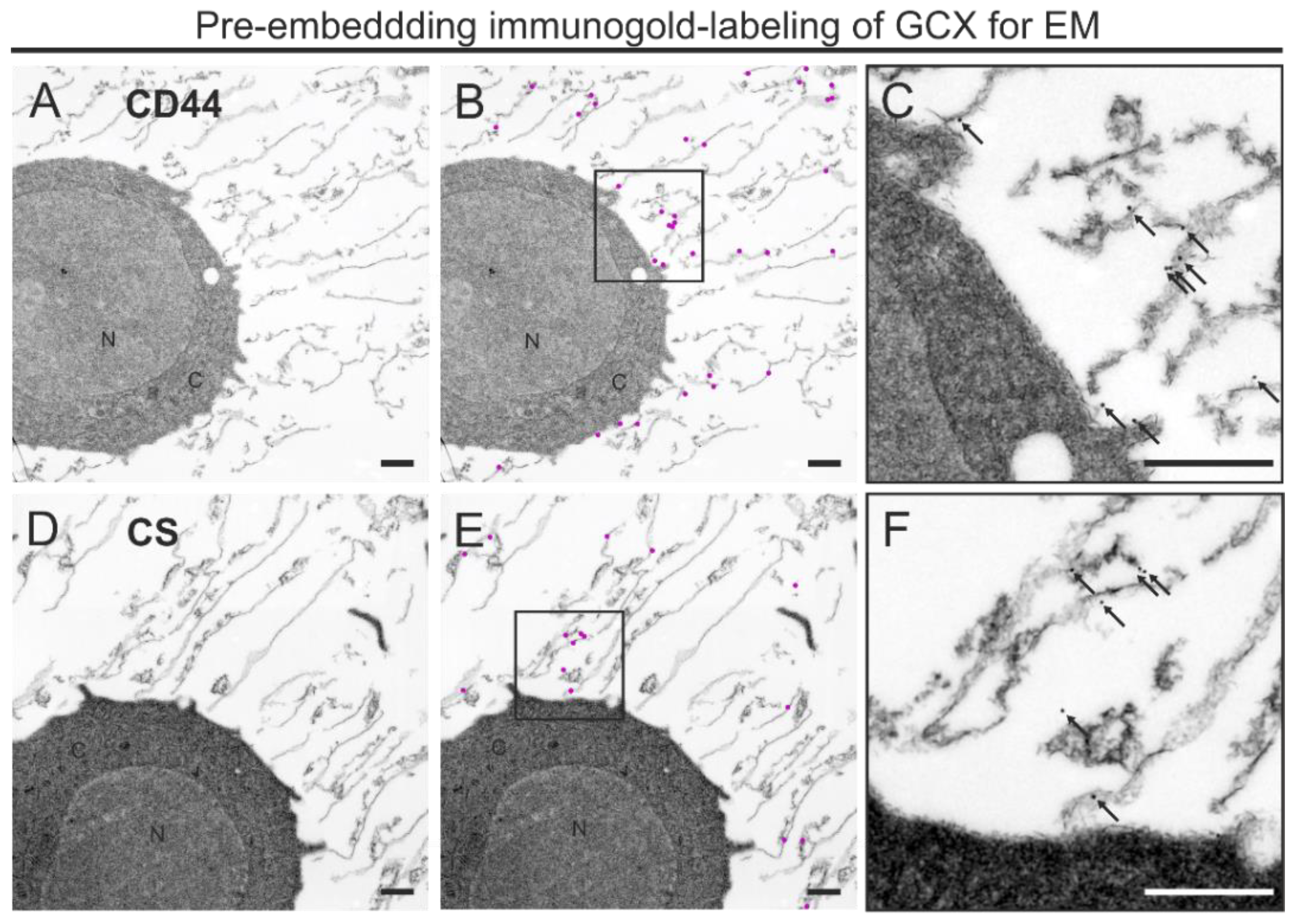
2.5. Pre-Embedding Immunogold Labeling and Processing for TEM
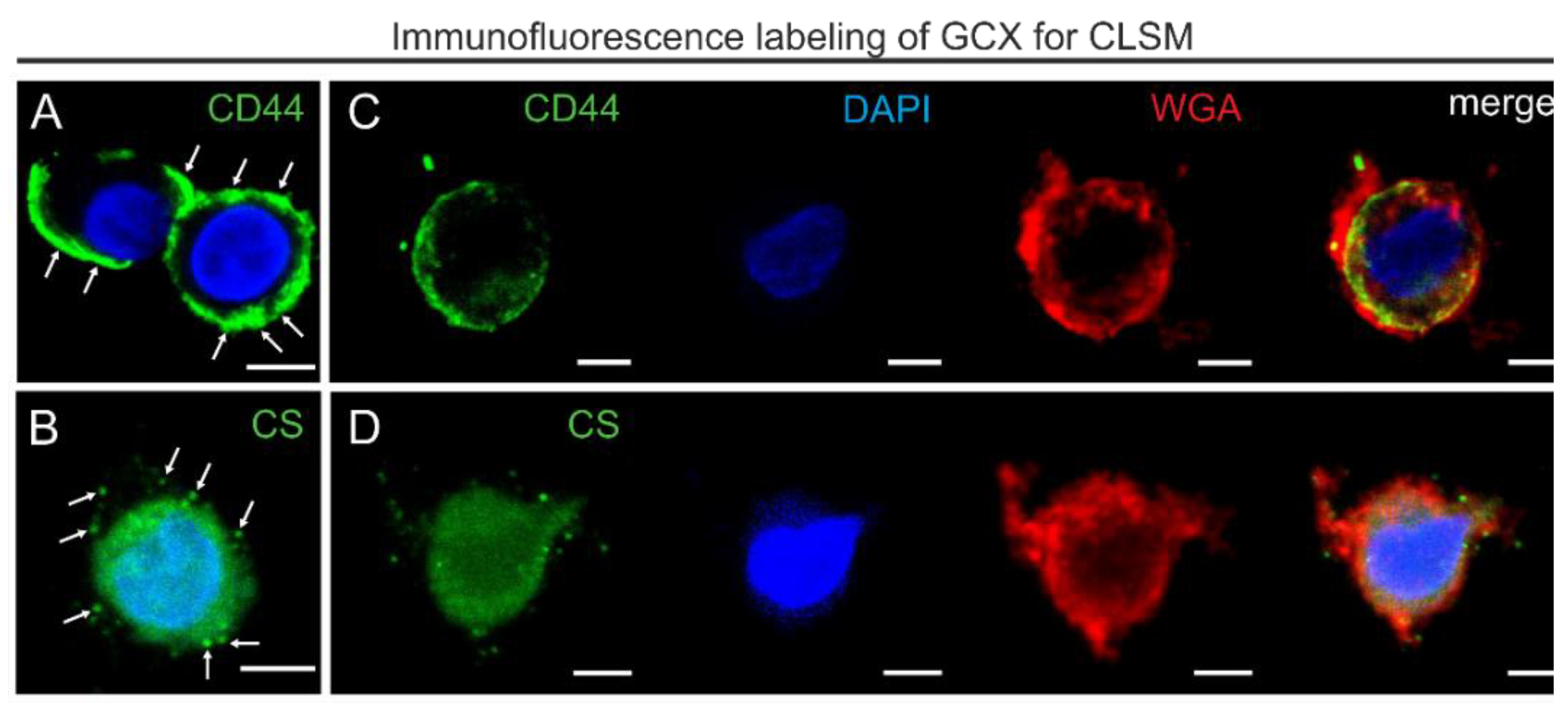
2.6. Immunofluorescence Labeling and Confocal Imaging
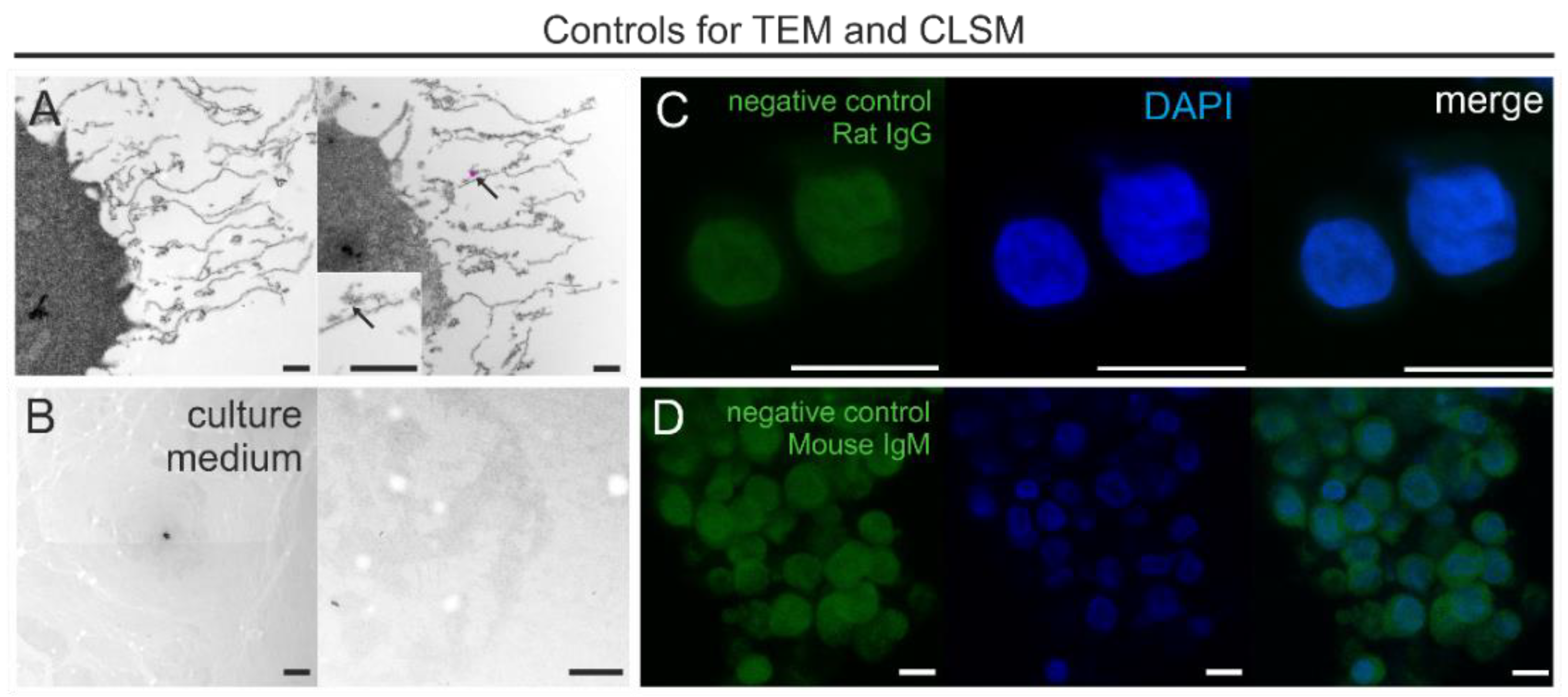
2.7. Control Experiments
3. Results
3.1. Antigen Presentation after Formalin Fixation of THP-1 Cells
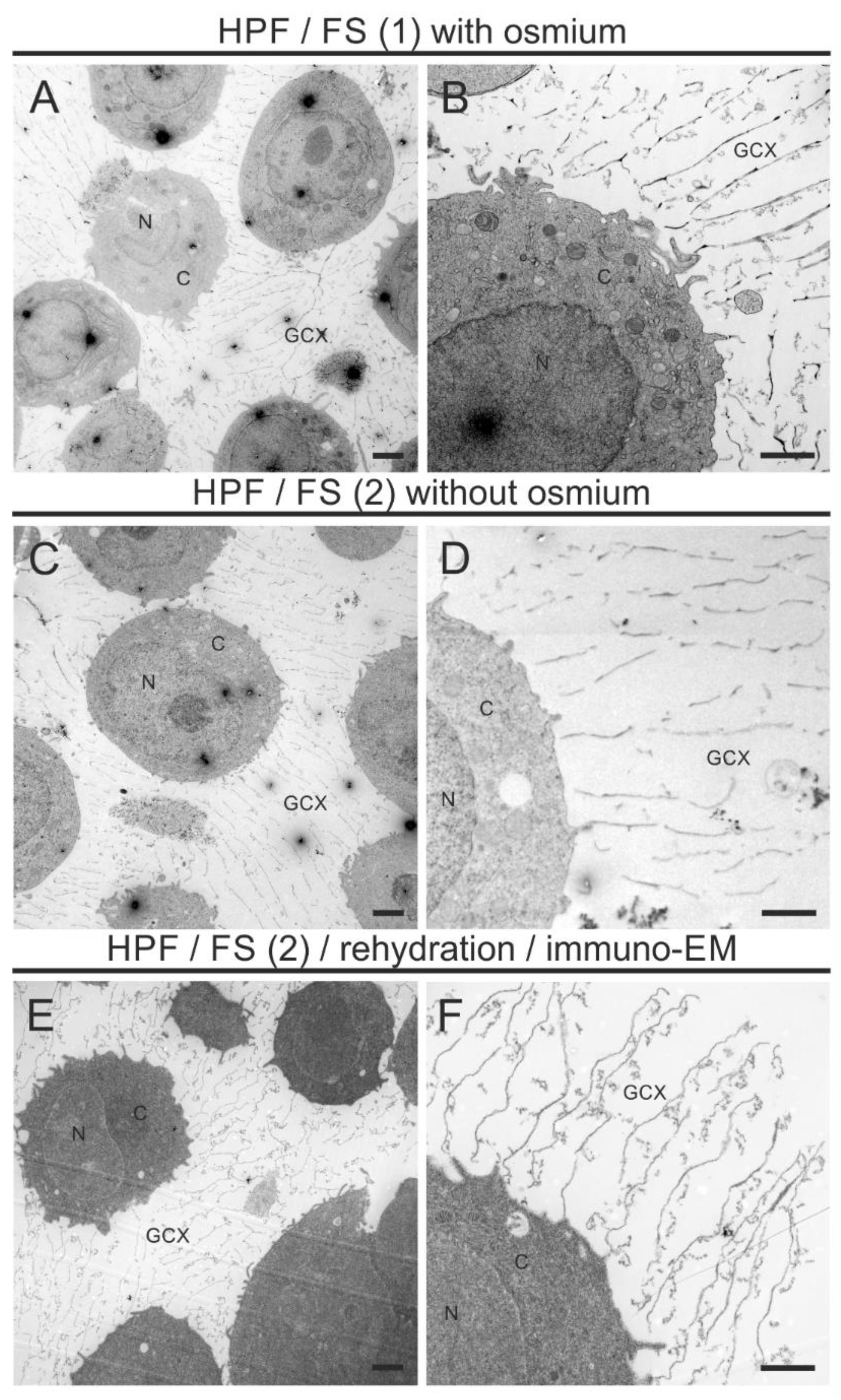
3.2. A Combined HPF, FS, Rehydration, and Pre-Embedding Immunogold Method for Preserved Ultrastructure and Antigen Localization in the GCX of THP-1 Cells by TEM
3.3. Adjustment of the Method for Confocal Laser Scanning Microscopy
3.4. Control Experiments
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Lieleg, O.; Ribbeck, K. Biological hydrogels as selective diffusion barriers. Trends Cell Biol. 2011, 21, 543–551. [Google Scholar] [CrossRef]
- Lipowsky, H.H. The endothelial glycocalyx as a barrier to leukocyte adhesion and its mediation by extracellular proteases. Ann. Biomed. Eng. 2012, 40, 840–848. [Google Scholar] [CrossRef]
- Constantinou, P.E.; Morgado, M.; Carson, D.D. Transmembrane mucin expression and function in embryo implantation and placentation. Adv. Anat. Embryol. Cell Biol. 2015, 216, 51–68. [Google Scholar]
- Tarbell, J.M.; Cancel, L.M. The glycocalyx and its significance in human medicine. J. Intern. Med. 2016, 280, 97–113. [Google Scholar] [CrossRef]
- Reitsma, S.; Slaaf, D.W.; Vink, H.; van Zandvoort, M.A.; oude Egbrink, M.G. The endothelial glycocalyx: Composition, functions, and visualization. Pflugers Arch. 2007, 454, 345–359. [Google Scholar] [CrossRef]
- Ushiyama, A.; Kataoka, H.; Iijima, T. Glycocalyx and its involvement in clinical pathophysiologies. J. Intensive. Care 2016, 4. [Google Scholar] [CrossRef]
- Chevalier, L.; Selim, J.; Genty, D.; Baste, J.M.; Piton, N.; Boukhalfa, I.; Hamzaoui, M.; Pareige, P.; Richard, V. Electron microscopy approach for the visualization of the epithelial and endothelial glycocalyx. Morphologie 2017, 101, 55–63. [Google Scholar] [CrossRef]
- Ebong, E.E.; Macaluso, F.P.; Spray, D.C.; Tarbell, J.M. Imaging the endothelial glycocalyx in vitro by rapid freezing/freeze substitution transmission electron microscopy. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1908–1915. [Google Scholar] [CrossRef]
- Poller, W.C.; Lowa, N.; Schleicher, M.; Munster-Wandowski, A.; Taupitz, M.; Stangl, V.; Ludwig, A.; Wiekhorst, F. Initial interaction of citrate-coated iron oxide nanoparticles with the glycocalyx of thp-1 monocytes assessed by real-time magnetic particle spectroscopy and electron microscopy. Sci. Rep. 2020, 10, 3591. [Google Scholar] [CrossRef] [PubMed]
- Studer, D.; Humbel, B.M.; Chiquet, M. Electron microscopy of high pressure frozen samples: Bridging the gap between cellular ultrastructure and atomic resolution. Histochem. Cell Biol. 2008, 130, 877–889. [Google Scholar] [CrossRef]
- Hess, M.W.; Vogel, G.F.; Yordanov, T.E.; Witting, B.; Gutleben, K.; Ebner, H.L.; de Araujo, M.E.G.; Filipek, P.A.; Huber, L.A. Combining high-pressure freezing with pre-embedding immunogold electron microscopy and tomography. Traffic 2018, 19, 639–649. [Google Scholar] [CrossRef]
- Deetz, J.S.; Behrman, E.J. Reaction of osmium reagents with amino acids and proteins. Reactivity of amino acid residues and peptide bond cleavage. Int. J. Pept. Protein Res. 1981, 17, 495–500. [Google Scholar] [CrossRef]
- Studer, D.; Michel, M.; Muller, M. High-pressure freezing comes of age. Scanning Microsc. 1989, 253–269. [Google Scholar]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and characterization of a human acute monocytic leukemia cell line (thp-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef]
- Makatsori, E.; Karamanos, N.K.; Papadogiannakis, N.; Hjerpe, A.; Anastassiou, E.D.; Tsegenidis, T. Synthesis and distribution of glycosaminoglycans in human leukemic b- and t-cells and monocytes studied using specific enzymic treatments and high-performance liquid chromatography. Biomed Chromatogr. 2001, 15, 413–417. [Google Scholar] [CrossRef]
- Makatsori, E.; Lamari, F.N.; Theocharis, A.D.; Anagnostides, S.; Hjerpe, A.; Tsegenidis, T.; Karamanos, N.K. Large matrix proteoglycans, versican and perlecan, are expressed and secreted by human leukemic monocytes. Anticancer. Res. 2003, 23, 3303–3309. [Google Scholar]
- Ambrosius, M.; Kleesiek, K.; Gotting, C. Quantitative determination and comparison of the glycosaminoglycan delta-disaccharide composition in 22 different human cell lines. Cell Biol. Int. 2009, 33, 848–852. [Google Scholar] [CrossRef]
- Allen, A.K.; Neuberger, A.; Sharon, N. Purification, composition and specificity of wheat-germ agglutinin. Biochem. J. 1973, 131, 155–162. [Google Scholar] [CrossRef]
- Janczyk, P.; Hansen, S.; Bahramsoltani, M.; Plendl, J. The glycocalyx of human, bovine and murine microvascular endothelial cells cultured in vitro. J. Electron. Microsc. 2010, 59, 291–298. [Google Scholar] [CrossRef]
- Devaraj, S.; Yun, J.M.; Adamson, G.; Galvez, J.; Jialal, I. C-reactive protein impairs the endothelial glycocalyx resulting in endothelial dysfunction. Cardiovasc. Res. 2009, 84, 479–484. [Google Scholar] [CrossRef]
- Chappell, D.; Jacob, M.; Paul, O.; Rehm, M.; Welsch, U.; Stoeckelhuber, M.; Conzen, P.; Becker, B.F. The glycocalyx of the human umbilical vein endothelial cell: An impressive structure ex vivo but not in culture. Circ. Res. 2009, 104, 1313–1317. [Google Scholar] [CrossRef] [PubMed]
- Sosinsky, G.E.; Crum, J.; Jones, Y.Z.; Lanman, J.; Smarr, B.; Terada, M.; Martone, M.E.; Deerinck, T.J.; Johnson, J.E.; Ellisman, M.H. The combination of chemical fixation procedures with high pressure freezing and freeze substitution preserves highly labile tissue ultrastructure for electron tomography applications. J. Struct. Biol. 2008, 161, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.L.; Zheng, Y.; Pierce, A.; Ackermann, M.; Horstmann, H.; Kuner, T.; Ronchi, P.; Schwab, Y.; Konietzke, P.; Wunnemann, F.; et al. Mesopolysaccharides: The extracellular surface layer of visceral organs. PLoS ONE 2020, 15, e0238798. [Google Scholar] [CrossRef]
- Alphonsus, C.S.; Rodseth, R.N. The endothelial glycocalyx: A review of the vascular barrier. Anaesthesia 2014, 69, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Jacob, M.; Bruegger, D.; Rehm, M.; Welsch, U.; Conzen, P.; Becker, B.F. Contrasting effects of colloid and crystalloid resuscitation fluids on cardiac vascular permeability. Anesthesiology 2006, 104, 1223–1231. [Google Scholar] [CrossRef] [PubMed]
- Stevens, A.P.; Hlady, V.; Dull, R.O. Fluorescence correlation spectroscopy can probe albumin dynamics inside lung endothelial glycocalyx. Am. J. Physiol. Lung C 2007, 293, L328–L335. [Google Scholar] [CrossRef] [PubMed]
- Jacob, M.; Paul, O.; Mehringer, L.; Chappell, D.; Rehm, M.; Welsch, U.; Kaczmarek, I.; Conzen, P.; Becker, B.F. Albumin augmentation improves condition of guinea pig hearts after 4 hr of cold ischemia. Transplantation 2009, 87, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.; Orenstein, J.M. Processing tissue and cells for transmission electron microscopy in diagnostic pathology and research. Nat. Protoc. 2007, 2, 2439–2450. [Google Scholar] [CrossRef]
- Testillano, P.S.; Sanchezpina, M.A.; Olmedilla, A.; Ollacarizqueta, M.A.; Tandler, C.J.; Risueno, M.C. A specific ultrastructural method to reveal DNA—The nama-ur. J. Histochem. Cytochem. 1991, 39, 1427–1438. [Google Scholar] [CrossRef]
- Svitkina, T.M.; Verkhovsky, A.B.; Borisy, G.G. Improved procedures for electron microscopic visualization of the cytoskeleton of cultured cells. J. Struct. Biol. 1995, 115, 290–303. [Google Scholar] [CrossRef]
- Walther, P.; Ziegler, A. Freeze substitution of high-pressure frozen samples: The visibility of biological membranes is improved when the substitution medium contains water. J. Microsc. 2002, 208, 3–10. [Google Scholar] [CrossRef]
- Oliver, C.; Jamur, M.C. Fixation and embedding. Methods Mol. Biol. 2010, 588, 353–362. [Google Scholar]
- van Donselaar, E.; Posthuma, G.; Zeuschner, D.; Humbel, B.M.; Slot, J.W. Immunogold labeling of cryosections from high-pressure frozen cells. Traffic 2007, 8, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Avnur, Z.; Geiger, B. Immunocytochemical localization of native chondroitin-sulfate in tissues and cultured-cells using specific monoclonal-antibody. Cell 1984, 38, 811–822. [Google Scholar] [CrossRef]
- Cichy, J.; Pure, E. The liberation of cd44. J. Cell Biol. 2003, 161, 839–843. [Google Scholar] [CrossRef]
- Braumueller, H.; Ramadani, M.; Beger, H.G.; Gansauge, F. A possible mechanism for metastasation and invasion of cancer cells by mimicking macrophage invasiveness. Gastroenterology 2000, 118, A1506. [Google Scholar] [CrossRef]
- Schumacher, U.; Thielke, E.; Adam, E. A dot blot technique for the analysis of interactions of lectins with glycosaminoglycans. Histochem. J. 1992, 24, 453–455. [Google Scholar] [CrossRef]
- Kataoka, H.; Ushiyama, A.; Kawakami, H.; Akimoto, Y.; Matsubara, S.; Iijima, T. Fluorescent imaging of endothelial glycocalyx layer with wheat germ agglutinin using intravital microscopy. Microsc Res. Techniq. 2016, 79, 31–37. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Supplier | Host | Dilution |
|---|---|---|---|
| CD44 (IM7) FITC | Invitrogen 14-0441-85 | rat/IgG2b monoclonal | CLSM, 1:500 |
| CD44 (IM7) | Invitrogen 14-0441-82 | rat/IgG2b monoclonal | TEM, 1:100 CLSM, 1:500 |
| CS-56 | Abcam ab11570 | mouse/IgM monoclonal | TEM, 1:100 CLSM, 1:500 |
| THP-1 Cells no. | Standard FS (+ OsO4) | Optimized FS (ø OsO4) | ||
|---|---|---|---|---|
| GCX (µm) | CB (µm) | GCX (µm) | CB (µm) | |
| 37 | 6.60 ± 0.30 * | 13.71 ± 0.32 ** | 6.45 ± 0.26 * | 13.11 ± 0.25 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Twamley, S.G.; Stach, A.; Heilmann, H.; Söhl-Kielczynski, B.; Stangl, V.; Ludwig, A.; Münster-Wandowski, A. Immuno-Electron and Confocal Laser Scanning Microscopy of the Glycocalyx. Biology 2021, 10, 402. https://doi.org/10.3390/biology10050402
Twamley SG, Stach A, Heilmann H, Söhl-Kielczynski B, Stangl V, Ludwig A, Münster-Wandowski A. Immuno-Electron and Confocal Laser Scanning Microscopy of the Glycocalyx. Biology. 2021; 10(5):402. https://doi.org/10.3390/biology10050402
Chicago/Turabian StyleTwamley, Shailey Gale, Anke Stach, Heike Heilmann, Berit Söhl-Kielczynski, Verena Stangl, Antje Ludwig, and Agnieszka Münster-Wandowski. 2021. "Immuno-Electron and Confocal Laser Scanning Microscopy of the Glycocalyx" Biology 10, no. 5: 402. https://doi.org/10.3390/biology10050402
APA StyleTwamley, S. G., Stach, A., Heilmann, H., Söhl-Kielczynski, B., Stangl, V., Ludwig, A., & Münster-Wandowski, A. (2021). Immuno-Electron and Confocal Laser Scanning Microscopy of the Glycocalyx. Biology, 10(5), 402. https://doi.org/10.3390/biology10050402