Why Should DNA Topoisomerase I Have a Scaffold Activity?



Abstract
Simple Summary
Abstract
1. Introduction
2. Canonical Activities of Topo I
2.1. DNA Structure and the Need to Evolve Enzymes for Solving Its Topological Problems
2.2. DNA Topoisomerases
2.3. DNA Topoisomerase IB
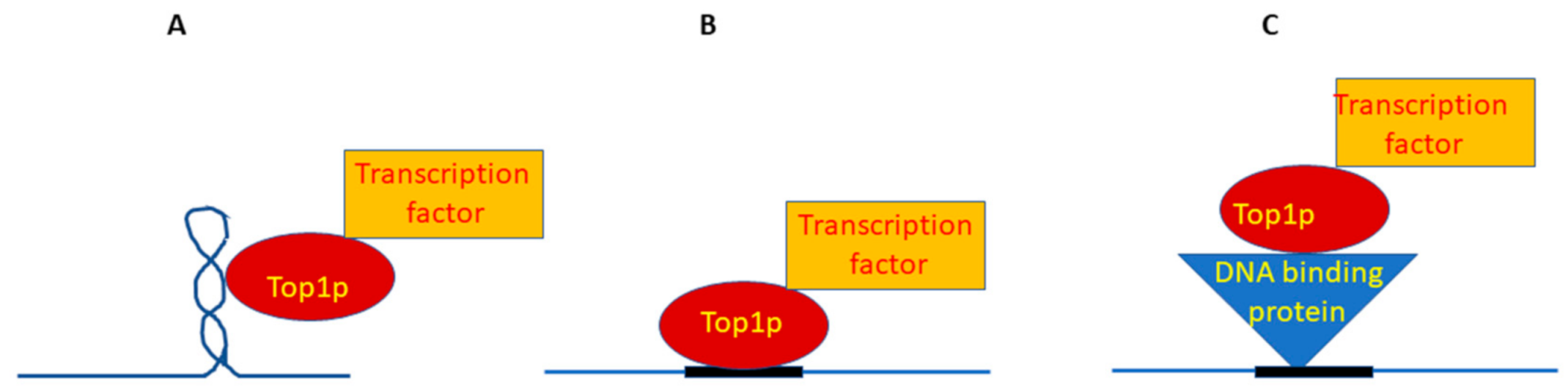
2.4. How Is a Region Recognized Where Topological Problems Have to Be Solved?
3. Catalysis Independent Functions of Topo I
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fuller, F.B. Decomposition of the linking number of a closed ribbon: A problem from molecular biology. Proc. Natl. Acad. Sci. USA 1978, 75, 3557–3561. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C. DNA Topoisomerases. Annu. Rev. Biochem. 1996, 65, 635–692. [Google Scholar] [CrossRef]
- Lee, M.P.; Brown, S.D.; Chen, A.; Hsieh, T.S. DNA topoisomerase I is essential in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1993, 90, 6656–6660. [Google Scholar] [CrossRef]
- Morham, S.G.; Kluckman, K.D.; Voulomanos, N.; Smithies, O. Targeted disruption of the mouse topoisomerase I gene by camptothecin selection. Mol. Cell. Biol. 1996, 16, 6804–6809. [Google Scholar] [CrossRef] [PubMed]
- Capranico, G.; Marinello, J.; Baranello, L. Dissecting the transcriptional functions of human DNA topoisomerase I by selective inhibitors: Implications for physiological and therapeutic modulation of enzyme activity. Biochim. Biophys. Acta (BBA) Bioenerg. 2010, 1806, 240–250. [Google Scholar] [CrossRef]
- Chen, S.H.; Chan, N.-L.; Hsieh, T.-S. New Mechanistic and Functional Insights into DNA Topoisomerases. Annu. Rev. Biochem. 2013, 82, 139–170. [Google Scholar] [CrossRef] [PubMed]
- Pommier, Y.; Sun, Y.; Huang, S.-Y.N.; Nitiss, J.L. Roles of eukaryotic topoisomerases in transcription, replication and genomic stability. Nat. Rev. Mol. Cell Biol. 2016, 17, 703–721. [Google Scholar] [CrossRef]
- Bansal, S.; Bajaj, P.; Pandey, S.; Tandon, V. Topoisomerases: Resistance versus Sensitivity, How Far We Can Go? Med. Res. Rev. 2016, 37, 404–438. [Google Scholar] [CrossRef]
- Champoux, J.J.; Dulbecco, R. An Activity from Mammalian Cells That Untwists Superhelical DNA—A Possible Swivel for DNA Replication. Proc. Natl. Acad. Sci. USA 1972, 69, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.F.; Wang, J.C. Supercoiling of the DNA template during transcription. Proc. Natl. Acad. Sci. USA 1987, 84, 7024–7027. [Google Scholar] [CrossRef]
- Tsao, Y.-P.; Wu, H.-Y.; Liu, L.F. Transcription-driven supercoiling of DNA: Direct biochemical evidence from in vitro studies. Cell 1989, 56, 111–118. [Google Scholar] [CrossRef]
- Wu, H.-Y.; Shyy, S.; Wang, J.C.; Liu, L.F. Transcription generates positively and negatively supercoiled domains in the template. Cell 1988, 53, 433–440. [Google Scholar] [CrossRef]
- Hsiang, Y.H.; Hertzberg, R.; Hecht, S.; Liu, L.F. Camptothecin induces protein-linked DNA breaks via mammalian DNA topoisomerase I. J. Biol. Chem. 1985, 260, 14873–14878. [Google Scholar] [CrossRef]
- Camilloni, G.; Di Martino, E.; Caserta, M.; Di Mauro, E. Eukaryotic DNA topoisomerase I reaction is topology dependent. Nucleic Acids Res. 1988, 16, 7071–7085. [Google Scholar] [CrossRef]
- Caserta, M.; Amadei, A.; Camilloni, G.; Di Mauro, E. Regulation of the function of eukaryotic DNA topoisomerase I: Analysis of the binding step and of the catalytic constants of topoisomerization as a function of DNA topology. Biochemistry 1990, 29, 8152–8157. [Google Scholar] [CrossRef]
- Liu, L.F.; Desai, S.D.; Li, T.-K.; Mao, Y.; Sun, M.; Sim, S.-P. Mechanism of Action of Camptothecin. Ann. N. Y. Acad. Sci. 2006, 922, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Labourier, E.; Forné, T.; Divita, G.; Derancourt, J.; François Riou, J.; Antoine, E.; Cathala, G.; Brunel, C.; Tazi, J. Specific phosphorylation of SR proteins by mammalian DNA topoisomerase I. Nat. Cell Biol. 1996, 381, 80–82. [Google Scholar] [CrossRef]
- Rossi, F.; Labourier, E.; Gallouzi, I.-E.; Derancourt, J.; Allemand, E.; Divita, G.; Tazi, J. The C-terminal domain but not the tyrosine 723 of human DNA topoisomerase I active site contributes to kinase activity. Nucleic Acids Res. 1998, 26, 2963–2970. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jeong, S. SR Proteins: Binders, Regulators, and Connectors of RNA. Mol. Cells 2017, 40, 1–9. [Google Scholar] [CrossRef]
- Long, J.C.; Caceres, J.F. The SR protein family of splicing factors: Master regulators of gene expression. Biochem. J. 2008, 417, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Soret, J.; Mathieu Gabut, C.D.; Glenda Kohlhagen, J.S.; Yves Pommier, J.T. Altered Ser-ine/Arginine-Rich Protein Phosphorylation and Exonic Enhancer-Dependent Splicing in Mammalian Cells Lacking Topoiso-merase I. Cancer Res. 2003, 63, 8203–8211. [Google Scholar]
- Tuduri, S.; Crabbé, L.; Conti, C.; Tourrière, H.; Holtgreve-Grez, H.; Jauch, A.; Pantesco, V.; De Vos, J.; Thomas, A.; Theillet, C.; et al. Topoisomerase I suppresses genomic instability by preventing interference between replication and transcription. Nat. Cell Biol. 2009, 11, 1315–1324. [Google Scholar] [CrossRef]
- Tazi, J.; Rossi, F.; Labourier, E.; Gallouzi, I.-E.; Brunel, C.; Antoine, E. DNA topoisomerase I: Customs officer at the border between DNA and RNA worlds? J. Mol. Med. 1997, 75, 786–800. [Google Scholar] [CrossRef] [PubMed]
- D’Alfonso, A.; Di Felice, F.; Carlini, V.; Wright, C.M.; Hertz, M.I.; Bjornsti, M.-A.; Camilloni, G. Molecular Mechanism of DNA Topoisomerase I-Dependent rDNA Silencing: Sir2p Recruitment at Ribosomal Genes. J. Mol. Biol. 2016, 428, 4905–4916. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Sternglanz, R. Identification and Characterization of the Genes for Two Topoisomerase I-Interacting Proteins from Saccharomyces Cerevisiae. Yeast 1999, 15, 35–41. [Google Scholar] [CrossRef]
- Katou, Y.; Kanoh, Y.; Bando, M.; Noguchi, H.; Tanaka, H.; Ashikari, T.; Sugimoto, K.; Shirahige, K. S-phase checkpoint proteins Tof1 and Mrc1 form a stable replication-pausing complex. Nat. Cell Biol. 2003, 424, 1078–1083. [Google Scholar] [CrossRef] [PubMed]
- Nedelcheva, M.N.; Roguev, A.; Dolapchiev, L.B.; Shevchenko, A.; Taskov, H.B.; Shevchenko, A.; Stewart, A.F.; Stoynov, S.S. Uncoupling of Unwinding from DNA Synthesis Implies Regulation of MCM Helicase by Tof1/Mrc1/Csm3 Checkpoint Complex. J. Mol. Biol. 2005, 347, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Brito, I.L.; Villén, J.; Gygi, S.P.; Amon, A.; Moazed, D. Inhibition of homologous recombination by a cohesin-associated clamp complex recruited to the rDNA recombination enhancer. Genes Dev. 2006, 20, 2887–2901. [Google Scholar] [CrossRef]
- Kretzschmar, M.; Meisterernst, M.; Roeder, R.G. Identification of human DNA topoisomerase I as a cofactor for activa-tor-dependent transcription by RNA polymerase II. Proc. Natl. Acad. Sci. USA 1993, 90, 11508–11512. [Google Scholar] [CrossRef]
- Merino, A.; Madden, K.R.; Lane, W.S.; Champoux, J.J.; Reinberg, D. DNA topoisomerase I is involved in both repression and activation of transcription. Nat. Cell Biol. 1993, 365, 227–232. [Google Scholar] [CrossRef]
- Shykind, B.M.; Kim, J.; Stewart, L.; Champoux, J.J.; Sharp, P.A. Topoisomerase I Enhances TFIID-TFIIA Complex Assem-bly during Activation of Transcription. Genes Dev. 1997, 11, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Bhalla, N. Moonlighting Proteins. Annu. Rev. Genet. 2020, 54, 265–285. [Google Scholar] [CrossRef] [PubMed]
- Bonven, B.J.; Gocke, E.; Westergaard, O. A high affinity topoisomerase I binding sequence is clustered at DNAase I hypersensitive sites in tetrahymena R-chromatin. Cell 1985, 41, 541–551. [Google Scholar] [CrossRef]
- Vogelauer, M.; Camilloni, G. Site-specific in vivo cleavages by DNA topoisomerase I in the regulatory regions of the 35 S rRNA in Saccharomyces cerevisiae are transcription independent. J. Mol. Biol. 1999, 293, 19–28. [Google Scholar] [CrossRef]
- Di Felice, F.; Egidi, A.; D’Alfonso, A.; Camilloni, G. Fob1p recruits DNA topoisomerase I to ribosomal genes locus and contributes to its transcriptional silencing maintenance. Int. J. Biochem. Cell Biol. 2019, 110, 143–148. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Topoisomerase | Cut/s on DNA Strands | Cofactor(s) Requirement | Covalent Intermediate | Relaxation | Examples |
|---|---|---|---|---|---|
| IA | Single | Mg++ | 5′-OH | Negative supercoil | Top3α human Top3βhuman Top3 yeast |
| IB | Single | No | 3′-OH | Negative and positive supercoil | Top1 human Top1 yeast Top1 Mitochondrial |
| IIA | Double | ATP, Mg++ | 5′-OH | Negative and positive supercoil | Top2 yeast Top2α human Top2β human |
| IIB | Double | ATP, Mg++ | 5′-OH | Negative and positive supercoil | Top VI Arabidopsis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Felice, F.; Camilloni, G. Why Should DNA Topoisomerase I Have a Scaffold Activity? Biology 2021, 10, 190. https://doi.org/10.3390/biology10030190
Di Felice F, Camilloni G. Why Should DNA Topoisomerase I Have a Scaffold Activity? Biology. 2021; 10(3):190. https://doi.org/10.3390/biology10030190
Chicago/Turabian StyleDi Felice, Francesca, and Giorgio Camilloni. 2021. "Why Should DNA Topoisomerase I Have a Scaffold Activity?" Biology 10, no. 3: 190. https://doi.org/10.3390/biology10030190
APA StyleDi Felice, F., & Camilloni, G. (2021). Why Should DNA Topoisomerase I Have a Scaffold Activity? Biology, 10(3), 190. https://doi.org/10.3390/biology10030190