1. Introduction
Animal personalities, or consistent individual differences in behavior that persist through time and across contexts, have been studied intensively in the last decades [
1,
2], and occur in many taxa, including invertebrates [
3]. Personalities have complex effects on fitness and have been suggested to entail ecological and evolutionary implications [
4]. Nevertheless, few studies have empirically tested for the relationships between individual variability in behavior and fitness consequences, and those show mixed results (reviewed in [
5]).
A likely explanation for the variation in the results of past studies is that fitness effects of a behavioral type are often context, condition, and/or sex-dependent [
6,
7,
8,
9]. For example, high activity can help males encounter more females and high aggression can increase their success when competing for access to females, both enhancing reproductive success. However, such attributes may be costly and disadvantageous in an antipredator context, where active and aggressive males may suffer higher survival costs [
10]. A well-known example of such a trade-off across contexts is a model called the aggressive spillover hypothesis, which was proposed to explain sexual cannibalism in the raft spider
Dolomedes fimbriatus [
11]. This model suggests that juvenile female aggression towards prey was selected to increase growth rate and fecundity, but “spills over” into aggression towards courting males in adulthood due to genetic constraints. Consequently, highly voracious females attack and kill approaching males and thereby remain unmated [
11].
The aggressive spillover hypothesis has been discussed frequently, but most studies support it only partly [
12]. Namely, several studies show a correlation between female voracity and precopulatory cannibalism, e.g., in
Dolomedes triton [
13],
Argiope aurantia [
14], and
Lycosa hispanica [
15], but the evidence for maladaptiveness of high female voracity and sexual cannibalism are lacking (reviewed in [
12]) or studies find effects in the opposite direction than predicted [
16]. Furthermore, a follow up study on
Dolomedes fimbriatus found no support for most of the predictions of the aggressive spillover hypothesis. While females exhibited consistent individual differences in voracity towards prey, voracity did not correlate with individual variability in aggression towards males and sexual cannibalism [
16]. However, both female aggression towards males and sexual cannibalism correlated with the relative size difference between mates, where females more likely consumed relatively smaller males [
16]. The latter indicates sexual cannibalism as a mechanism of mate choice based on male size, a pattern found in some other spider species (e.g., [
17,
18]).
While sexual selection studies mostly focus on the effects of body size or other morphological traits (color, ornaments), behavioral traits may also play an important role in intra-sexual competition and mate choice [
19]. The links between behavioral variation and sexual selection have been mainly investigated in vertebrate systems [
8,
19]. For example, in great tits (
Parus major) and zebra finches (
Taeniopygia guttata), behavioral traits are involved in mate choices of both sexes. Similarly, older males of the banded wren (
Thryothorus pleurostictus) sing more consistent songs and are more likely to mate than younger males [
20]. Thus, consistency is another source of potential information in sexual selection contexts, and sexual selection, therefore, likely affects the maintenance of personality traits [
19].
In spiders, the importance of behavioral traits in male–male competition and mate choice is understudied. For instance, in the bridge spider (
Larinioides sclopetarius)
, aggression in males correlates positively to the number of sired offspring, implying that aggressive males have an advantage in sperm competition [
21]. The significance of male aggression in mate choice has been shown in the hermit spider
Nephilingis livida, where more aggressive males are less likely to be cannibalized by females, and also achieve higher copulation rates [
22]. In
D. fimbriatus, however, there is no support for the effects of such among-individual differences in behavior on mating success and sexual cannibalism [
16].
Here, we explore the relationship among individual variation in behaviors, body size measures and mating success in the Mediterranean black widow (
Latrodectus tredecimguttatus). Characteristically for widow spiders (genus
Latrodectus), this species exhibits sexual size dimorphism (SSD) with females several-fold larger than males. Males of
L. tredecimguttatus are often monogamous, though not obligatorily [
23], while females are presumed to be polygamous [
23]. Several mating behaviors associated with SSD are present in this species, including sexual cannibalism, male genital damage, and female genital plugging and mate guarding [
24]. These extreme mating behaviors and SSD indicate widow spiders are under strong sexual selection [
25], which makes them an appropriate study group to explore the role of behavioral traits in the mating context.
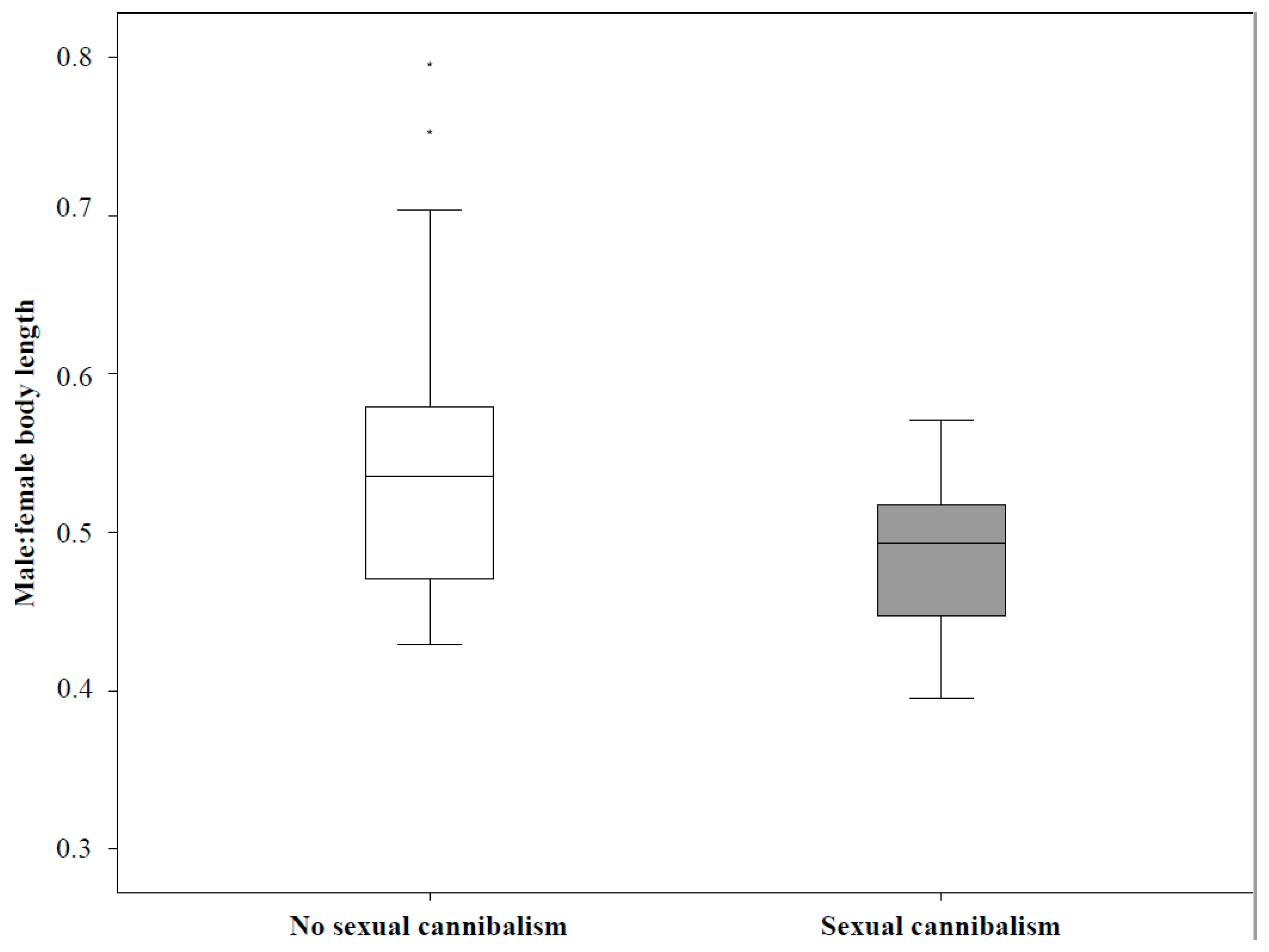
We observed the Mediterranean black widow in the individual and mating context, to test how body size measures and two functionally important behaviors, i.e., male aggression towards rivals and female voracity towards prey, affect mating behaviors, mating success, and sexual cannibalism. Specifically, we asked (i) whether males and females consistently differ in aggression towards a rival and voracity towards prey, respectively; (ii) whether and how behavioral traits correlate to measures of body size in both sexes; (iii) whether and how male aggression and female voracity, together with size measures, affect the behavior of individuals in the mating contexts; and (iv) whether size and behaviors correlate with male mating success (number and duration of copulations) and sexual cannibalism.
To broaden our understanding of extreme sexual behaviors connected to SSD, we additionally quantify mating behaviors in
L. tredecimguttatus in detail. We predicted that both sexes exhibit consistent individual variation in aggressive behavior. If increased aggression correlates to increased foraging success, we expected to find a positive correlation between aggression and body size measures. We predict larger males will win contests more often, and thus be more likely to gain mating opportunities. Furthermore, aggression may be beneficial for mating success, in particular in large males. Aggression in females, however, may limit their mating success if it spills over into aggression towards mates. In females, large body size advertises high fecundity [
26,
27], and we thus predicted that larger females are more attractive to males and achieve higher mating success (number and duration of copulations). Furthermore, we tested two additional hypotheses for sexual cannibalism. The mate-size dimorphism hypothesis predicts that sexual cannibalism depends on relative size difference between the male and the female [
17,
28]. Sexual cannibalism may also allow females to control copulation duration and thereby paternity of males [
29,
30]. Under assumptions that females should benefit from mating with more than one male, the probability of sexual cannibalism should increase with duration of copulation.
2. Material and Methods
2.1. Study Animals
We collected egg sacs of
L. tredecimguttatus in the field around Pula, Croatia (N44.940882°, E13.8718884°) at the beginning of their hatching season in early May 2017. We kept individual egg sacs in plastic containers until hatching. After their second molt, we moved individual spiders into their own plastic containers (5 cm × 10 cm) with a foam cover. From the initial egg sacs, we reared two generations of spiders, which we used for this study. We kept all spiders in standardized laboratory conditions, at 21–24 °C, 40–60% relative humidity, and natural lighting conditions. We sprayed and fed spiders twice per week, using size-appropriate food:
Drosophila melanogaster for small juveniles and males,
Lucilia sericata for middle sized juveniles and larvae of
Tenebrio molitor for adult females. To facilitate web construction for later experiments, we moved penultimate females onto wooden or metal constructions (20 cm × 12 cm × 11 cm), consisting of a horizontal plate and six vertical sticks (
Supplementary Material). The feeding and spraying regimes remained the same for spiders on these constructions. We recorded each experiment using a Panasonic HC-V180 camcorder, and we conducted all behavioral quantifications from recordings.
2.2. Female Voracity towards Prey
We quantified female voracity as her reaction to prey. Each female (N = 54) was tested for voracity twice before being in the mating experiment. After reaching adulthood, all females were fed with one food item, three days later tested for voracity and allowed to eat the prey, and then tested again for voracity three days later, then finally put into a mating trial after another three-day period. Before each experiment, we set the female’s web in front of a black background and between lights to visualize the web and spider for video recording. These spiders are active during the day and night, so the lights likely did not affect results. After 15 min of acclimatization, we placed a mealworm larva on the edge of the enclosure and allowed it to roam between sticky threads of the web (
Supplementary Material). We started quantification when prey first touched the web, and ended once the spider finished manipulating it, i.e., ignored it for at least 45 s or started feeding. We scored the following behaviors.
Latency to first reaction: time from web contact of prey to the first movement of the spider.
Latency to wrap prey: time from web contact of prey to beginning of prey wrapping.
Latency to bite prey: time from web contact of prey to first biting of prey.
Prey manipulation time: time from first reaction to the end of the experiment.
2.3. Aggression in Males
We quantified a males’ aggression as his behavior towards a conspecific male. Each male (N = 59) was tested twice, once on an empty adult female web and once in a mating trial. Female webs were chosen at random. In the first experiments, we randomly selected two males and placed them on an empty female web (
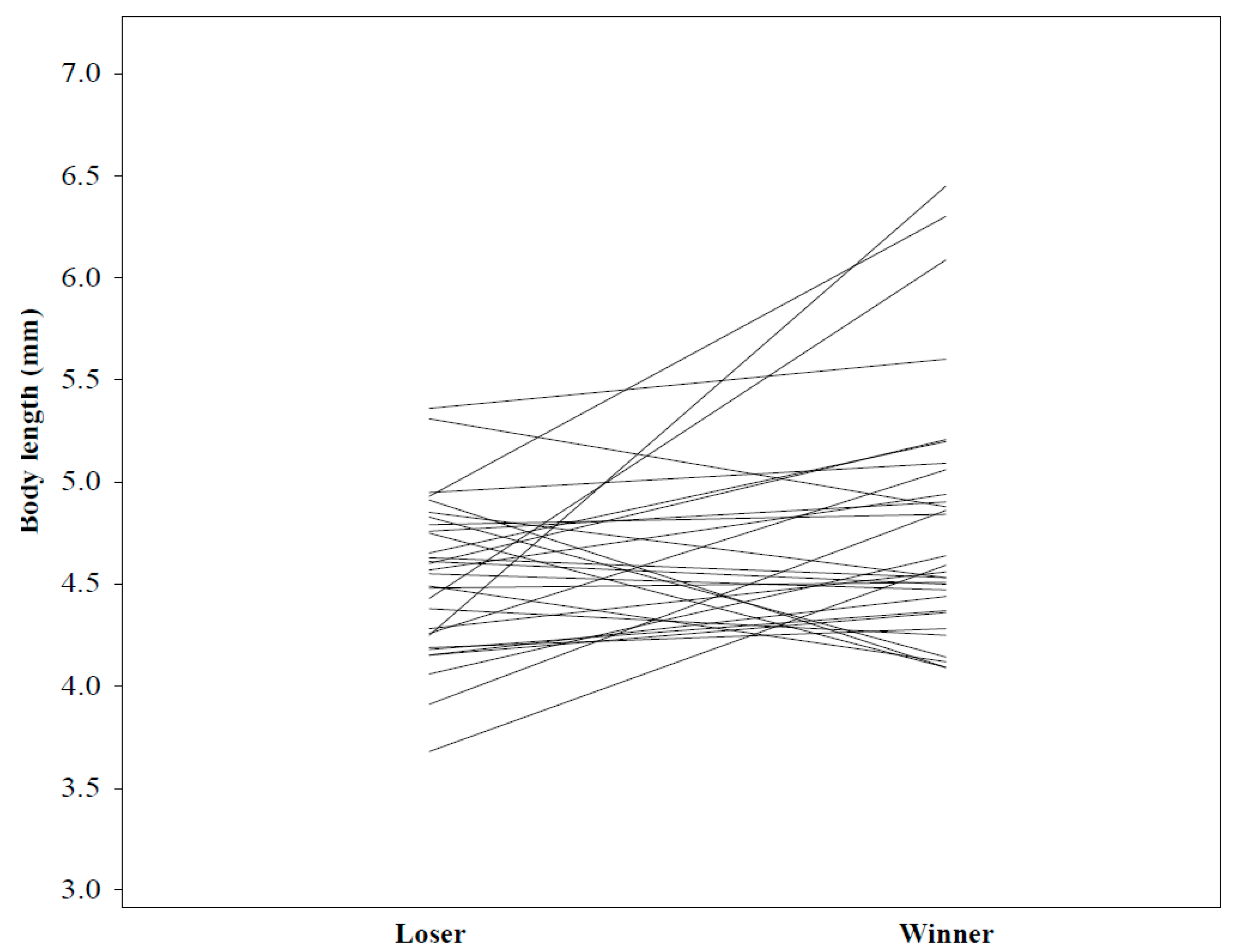
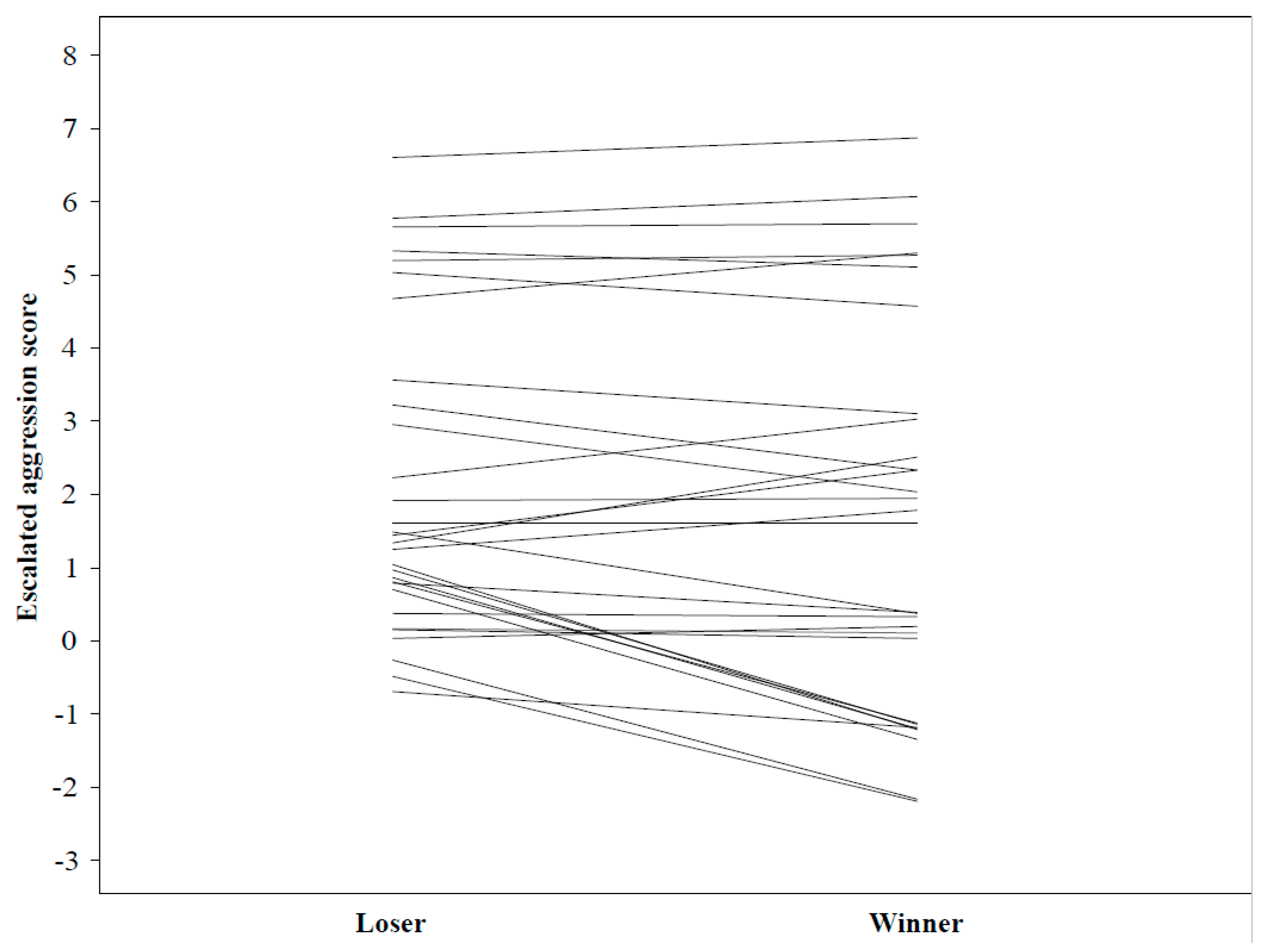
Supplementary Material). Males were paired with a different opponent in each test. Because most males matured within 1 and 2 weeks of each other, male age was ignored when pairing for personality experiments. To distinguish the males, we marked each on one of the legs using nail polish of different colors. We measured several antagonistic behaviors during a period of one hour or until one of the males lost the contest (escaped or fell from the web and stayed off it for more than 30 s). If neither male fled (N = 6), we declared the spider that was closest to the female retreat after one hour, the winner. Males started at opposing ends of the web, so they could meet as naturally as possible. Because males were highly aggressive during the first contest, with several losing a leg, we conducted the second contest as part of the mating trial to prevent larger injuries before mating trials. This contest again concluded if one of the males fell or escaped from the web (and stayed off for more than 30 s), or additionally, if one of the males mated with the female (we considered the mating as a win). If neither male mated or escaped, we again declared the male closest to the female retreat as the winner after 1 h. We immediately removed the losing male, to prevent him from interfering with the mating, in order to know the paternity of offspring for a related study. Males injured in the first experiments were not disqualified from mating experiments, because the loss of a leg did not appear to hinder their behavior. We scored the following behaviors:
Latency to first attack: time from the beginning of the experiment to the first attack. We defined an attack as any rapid movement towards the opponent in close proximity (see
Supplementary Materials).
Number of attacks: total number of attacks that a male conducted towards its opponent. Scored separately for each male.
Number of fights: total number of fights. A fight was defined as an attack that resulted in physical contact of the competing males.
Total fight duration: the summed duration (s) of all fights. We defined fight duration as time from attack until the escape or death of one of the spiders.
Number of movements towards opponent: total number of times that a male responded to the presence of its opponent by stopping his courting behavior and walking towards the opponent, occasionally shaking the web. Chance encounters were not included, and were defined as random encounters between two courting males.
Number of chases: total number of times the male chased his fleeing opponent. We defined a chase as the immediate movement of a male towards a fleeing opponent.
Aggressive web shaking: number of times the male shook the web at his opponent. Since males also use web shaking for signaling to the female, this was only counted when males shook the web near a rival male, not during courtship.
First attacker: whichever male attacked first, measured as binary, yes/no.
Number of escapes: total number of times the male escaped fighting during the experiment. An escape was defined as rapid movement away from the opponent.
Winner: whichever male won the contest (criteria defined above). Measured as binary, yes/no.
Experiment duration: total time of experiment.
2.4. Mating Trials
During mating trials (N = 30), we scored mating and antagonistic behaviors of both males, while competing for the female, as the second contest to quantify male aggression, and continued quantifying the winner’s behavior after removal of the losing male. Mating experiments were done as soon as possible, following the second personality experiment of the female (within 3 weeks of female maturation). If the female killed the male during copulation (N = 9), the experiment was considered over only after the males’ pedipalp was detached from her genitals. If the male survived mating and continued courting, we ended the experiment after a total of three hours, unless the male was currently mating at the three-hour mark. In this case, the experiment was extended until the end of mating (N = 1). We scored the following mating behaviors:
First contact: time from start of experiment to the first physical contact between the male and the female.
Silk deposition: male deposition of silk on the female web. Measured binary (yes/no for occurrence of the behavior) in intervals of one minute, for each male separately.
Web reduction: a male collapsing some of the female’s web using his chelicerae. Measured binary (yes/no for occurrence of the behavior) in intervals of one minute, for each male separately.
Mate binding: male silk deposition on the female’s body. Measured binary (yes/no for occurrence of the behavior) in intervals of one minute, for each male separately.
Oral sexual contact: contact between male chelicerae and the female’s genital opening. Measured binary (yes/no for occurrence of the behavior) in intervals of one minute, for each male separately.
Latency to first insertion: time from beginning of experiment to insertion of the male’s palp into the female’s genital opening.
Inserted palp: which palp was inserted (left/right).
Insertion duration: total time of insertion for each palp separately.
Sexual aggression: measured binary (yes/no) for whether the female attempted to bite the male, grasped at him with her legs, or attempted to cover him in silk.
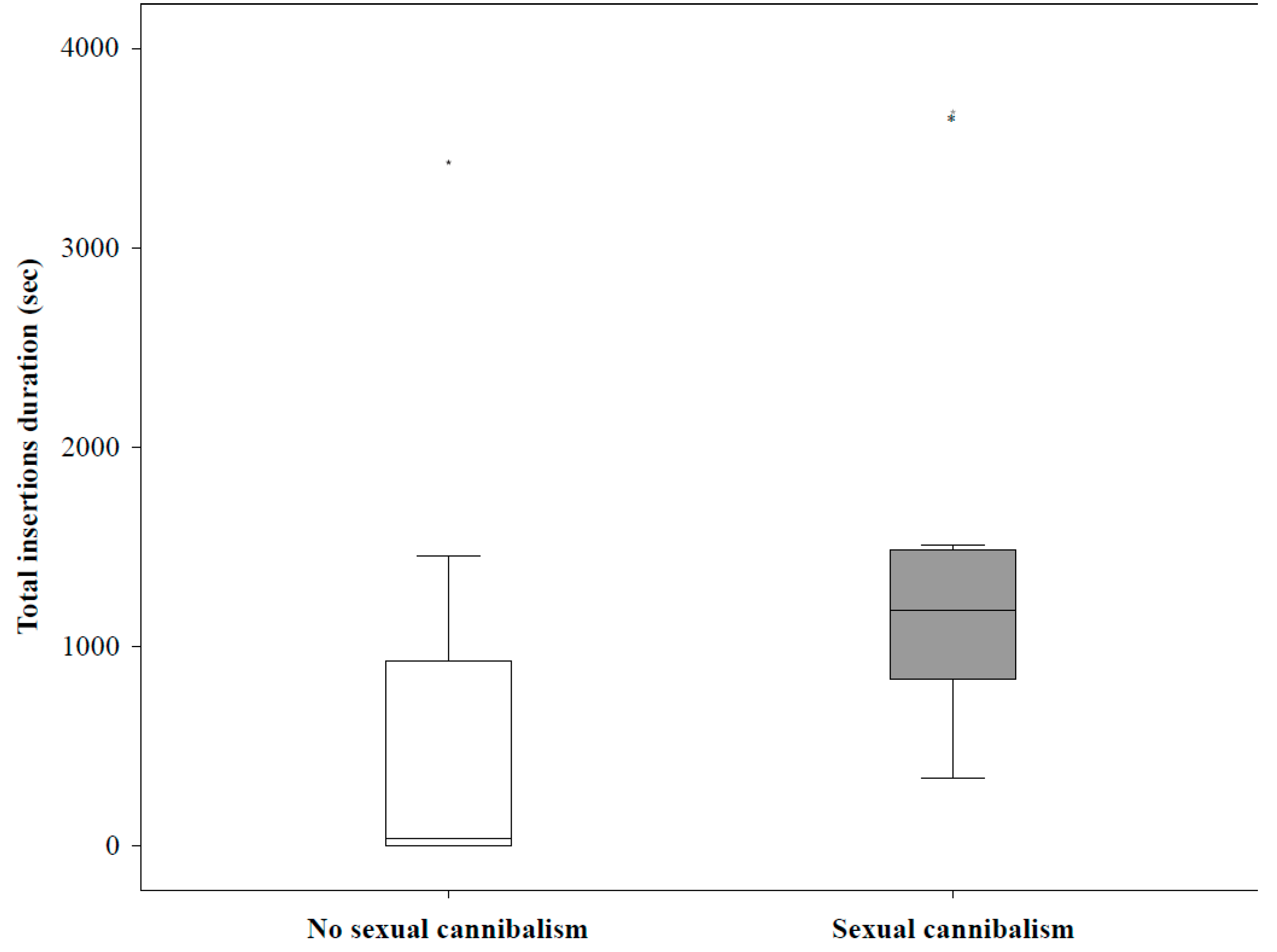
Sexual cannibalism: measured binary (yes/no) for whether the female killed the male during mating.
Total insertions: total number of pedipalp insertions.
Total insertion duration: summed time of all pedipalp insertions.
All behaviors were measured as frequencies, meaning that uninterrupted behavior was not counted more than once. For instance, a male shaking the female’s web for 10 s was only counted as shaking it once, while a male that stopped and moved between shakings had the behavior counted each time he started.
After noticing web reduction and silk deposition behavior by males, we conducted a subsequent experiment, testing for pheromones in female silk. We collected entire female webs on a sterile inoculation loop. Following [
31], we then washed the collected silk in methanol to extract potential pheromones. We soaked sterile filter paper in 50 µL of silk extract, and put each filter paper on a T-shaped climbing structure [
32]. We covered each filter paper with a plastic cage to prevent males from contacting it. We presented 10 males each to their own such silk extract and tested whether they will locate the filter paper and start courtship.
2.5. Body Size Measurements
Before all experiments, we photographed each spider using standardized settings on a Canon EOS 7D DSRL camera equipped with a 50 mm Canon macro lens. We used these photographs to later measure body length (prosoma + opisthosoma), as a linear measure of body size [
33]. Because removing females from their silken retreats caused significant web destruction, we used abdomen volume as a proxy of their body mass [
34,
35]. We measured abdomen volume using the formula
where a = abdomen length, b = abdomen width, and c = abdomen height [
34,
35]. We acquired abdomen measurements from photographs obtained by the above protocol. In addition, we calculated body condition in females. We used standardized residuals of abdomen volume over body length using linear regression.
2.6. Ethical Note
Research on spiders is not restricted by the animal welfare law of the country where the study was conducted. In the field, we collected the minimum number of individuals needed to conduct the research. The spiders were kept in conditions similar to their natural environmental conditions. The spiders were regularly fed with different prey items. The study was based on behavioural observations and all experiments were non-invasive. After the experiments, the spiders remained in the laboratory and were reared until natural death as described above.
2.7. Statistical Analyses
We analyzed whether females and males differ consistently in behaviors related to voracity towards prey and antagonistic behaviors towards rivals, respectively. Due to the large number of observed behaviors, we first reduced them by performing principal component analysis (PCA) with varimax rotation. In females, latency to first reaction, latency to wrap prey, and latency to bite prey were included in PCA, while duration of prey manipulation was analyzed separately, because it was not correlated to the former three behaviors. In males, we included in the PCA number of approaching the rival, number of web shakings, number of chases, attack latency, number of attacks, number of fights and fight duration. Because the two remaining behaviors, the attack first and winner-loser, are binary, we analyzed them separately. To estimate the relevant number of factors to extract, we used Kaiser’s eigenvalue-one rule and the screen test. To estimate an individual’s scoring on the extracted factor, as an underlying measure for a suite of correlated behaviors, we used the Anderson–Rubin method. The PCA determinants of the correlation matrices exceeded 0.0001, confirming there was no multicollinearity or singularity in the data subjected to PCAs. Measures of sampling adequacy indicated that the correlation matrices were appropriate for PCAs (Kaiser–Meyer–Olkin measure of sampling adequacy: >0.5, Bartlett’s test of sphericity: p < 0.05).
We then used generalized mixed models (glmm) to analyze individual repeatability of behaviors. To partition phenotypic variance into between-individual (Vind) and within- individual variance (Ve), we ran Markov Chain Monte Carlo (MCMC) glmm in R [
36], following [
37]. We calculated repeatability as
[
38]. In the models, we used trial (repeat 1, repeat 2) as a fixed factor, and animal ID as a random factor. However, when calculating the repeatability of male aggression, we added the ID of the contest as a random factor to account for the effect of an opponent’s behavior. We chose this approach because an individual’s aggression in the contest may depend in part on his rival’s behavior. We ran models using non informative priors. We checked convergence and mixing properties by visual inspection of the chains and checked the autocorrelation values. To verify that the number of iterations was adequate for MCMC chains to achieve convergence, we ran Heidelberger and Welch’s convergence diagnostics.
Next, we tested for the relationships between body size measures (body length, abdomen volume, and body condition) and behaviors that were significantly repeatable, i.e., voracity towards prey (primary factor from PCA analysis, see Results) and duration of prey manipulation in females, and with escalated aggression (aggression that continues beyond just a confrontation, e.g., fighting; also see
Section 3.2) and winner–loser in males. In females, we used Pearson’s correlation coefficient to test for correlations. We used the means of behaviors across the two repeats, and mean voracity was log transformed. Then, we tested whether an individual’s behavior within the mating context correlates to female aggressive behavior and body size measures (body length, abdomen volume). Using linear regression (continuous variables) and logistic regression (binary variables), we tested whether and how behaviors (voracity towards prey and duration of prey manipulation) and body measures affect mating success (number of copulations, total duration of copulations), aggression towards mates (yes/no), and sexual cannibalism (yes/no). Regressions were run with bootstrapping. In addition, we tested whether female aggression and sexual cannibalism depended on total copulation duration and the relative size difference between a male and female in a pair (males’ length: female length) using regression.
In males, we used MCMCglmm to test for the relationship between male aggressive behaviors (escalated aggression), winning the contest and body size measures. We asked whether and how male aggressive behaviors (escalated aggression), body length and their interaction (aggression × body length) affect winning the contest. We ran this model because we needed to account for an opponent’s behavior, thus we added experiment ID as a random variable.
Finally, we ran linear (continuous variables) or logistic regression (binary variables) to test how male body length, aggression, and courting activity affect mating success (yes, no), number of copulations (i.e., pedipalp insertions), total duration of copulations, received female aggression (yes/no), and occurrences of being cannibalized (yes/no). Regressions were run with bootstrapping.
We reduced the number of the recorded courtship behaviors (occurrences of silk deposition, web reduction, mate binding and oral contact) using PCA, as described above.
For the descriptions of mating behaviors, we analyzed only winning males from the mating contests, as losing males did not have a chance to fully court females. We report descriptive statistics as medians ± interquartile ranges.








{kind=link}
{kind=link}
{kind=link}
{kind=link}