
Comparison of Culturing and Metabarcoding Methods to Describe the Fungal Endophytic Assemblage of Brachypodium rupestre Growing in a Range of Anthropized Disturbance Regimes


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
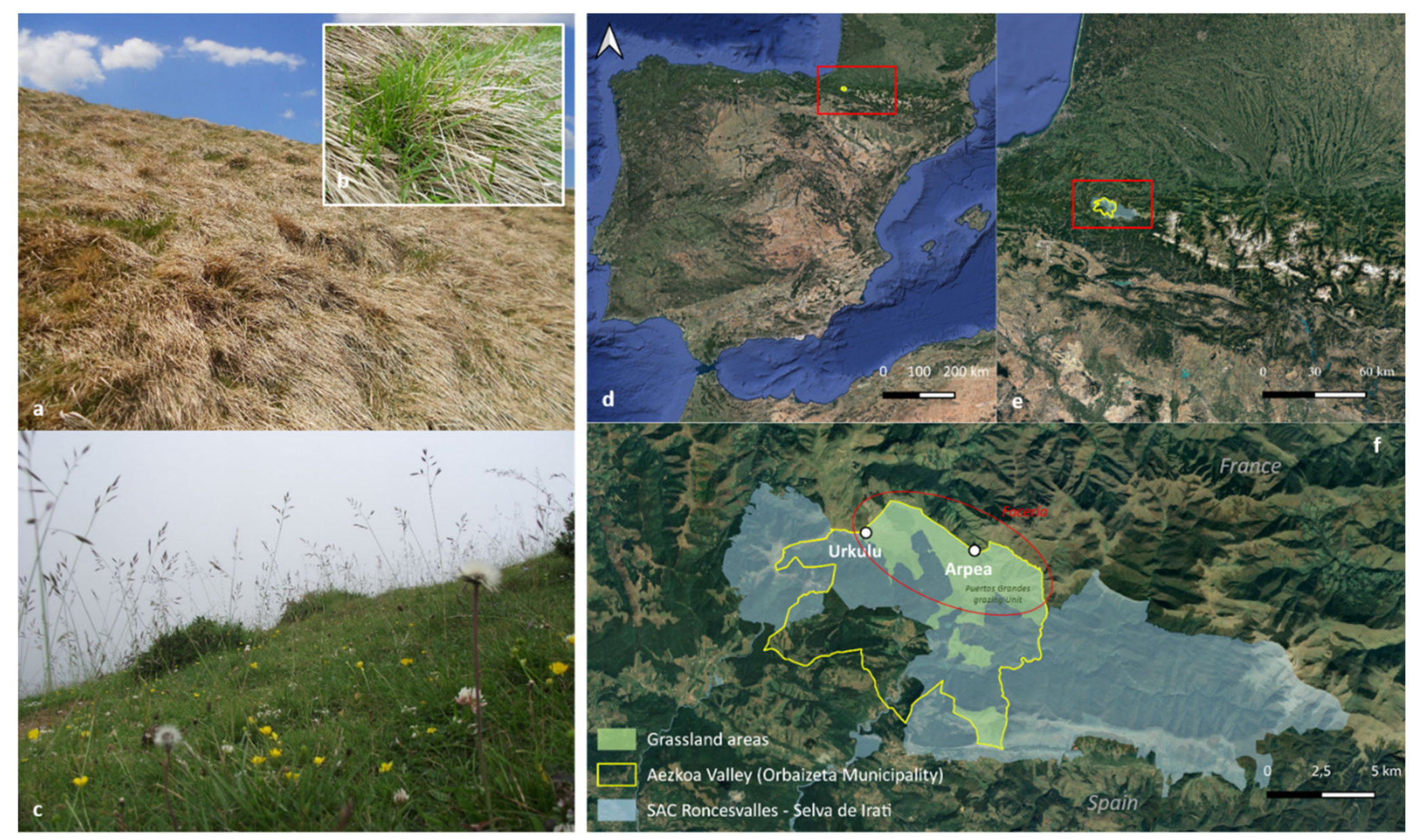
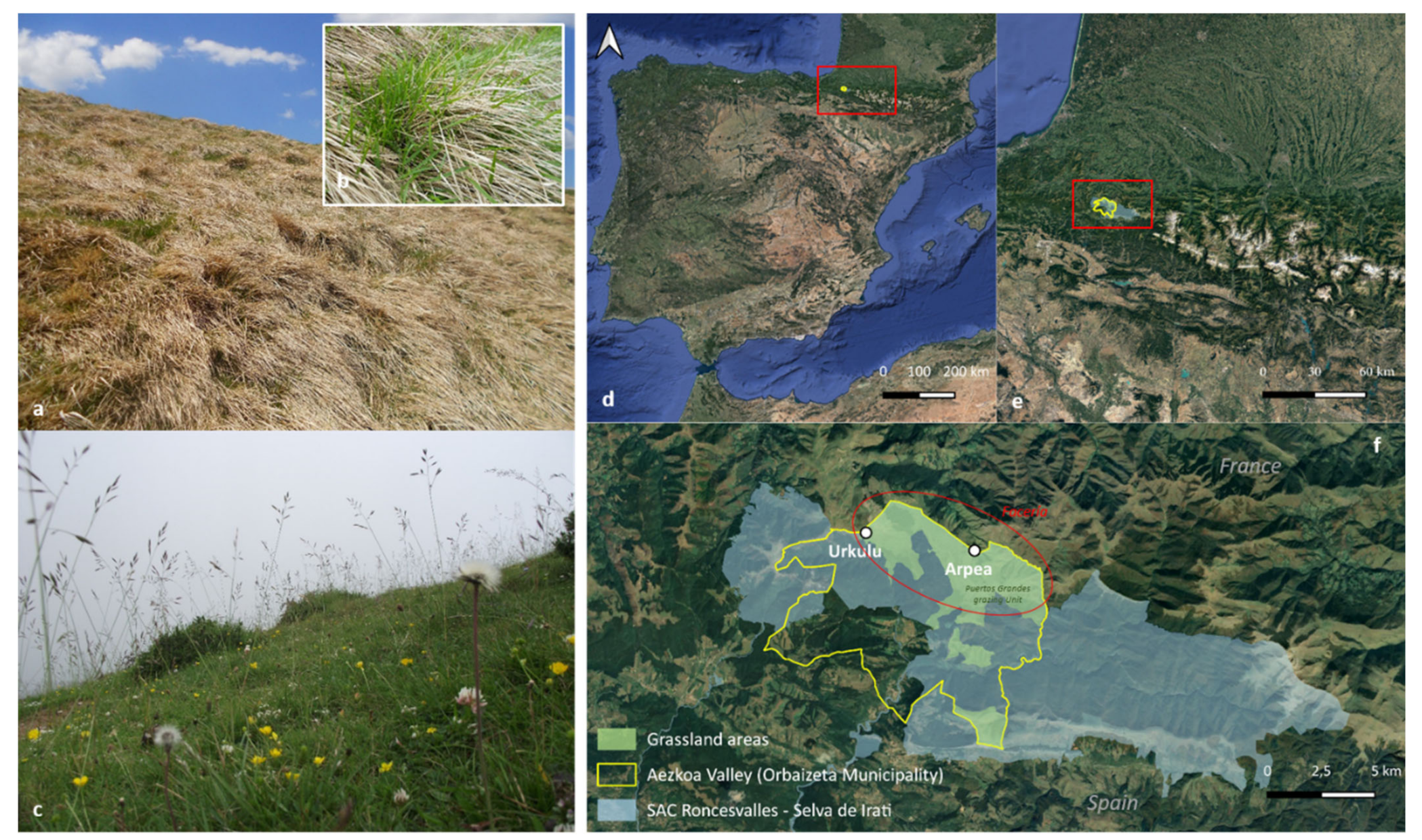
2.1. The Study Area
2.2. Plant Sampling
2.3. Isolation and Identification of Fungi Using the Culturing Method
2.4. Metabarcoding Analysis and Taxonomic Assignment
2.5. Data Analysis
3. Results
3.1. Comparison of B. rupestre Mycobiome Obtained by Culturing and Metabarcoding Methods
3.2. The Mycobiome of B. rupestre According to the Metabarcoding Method
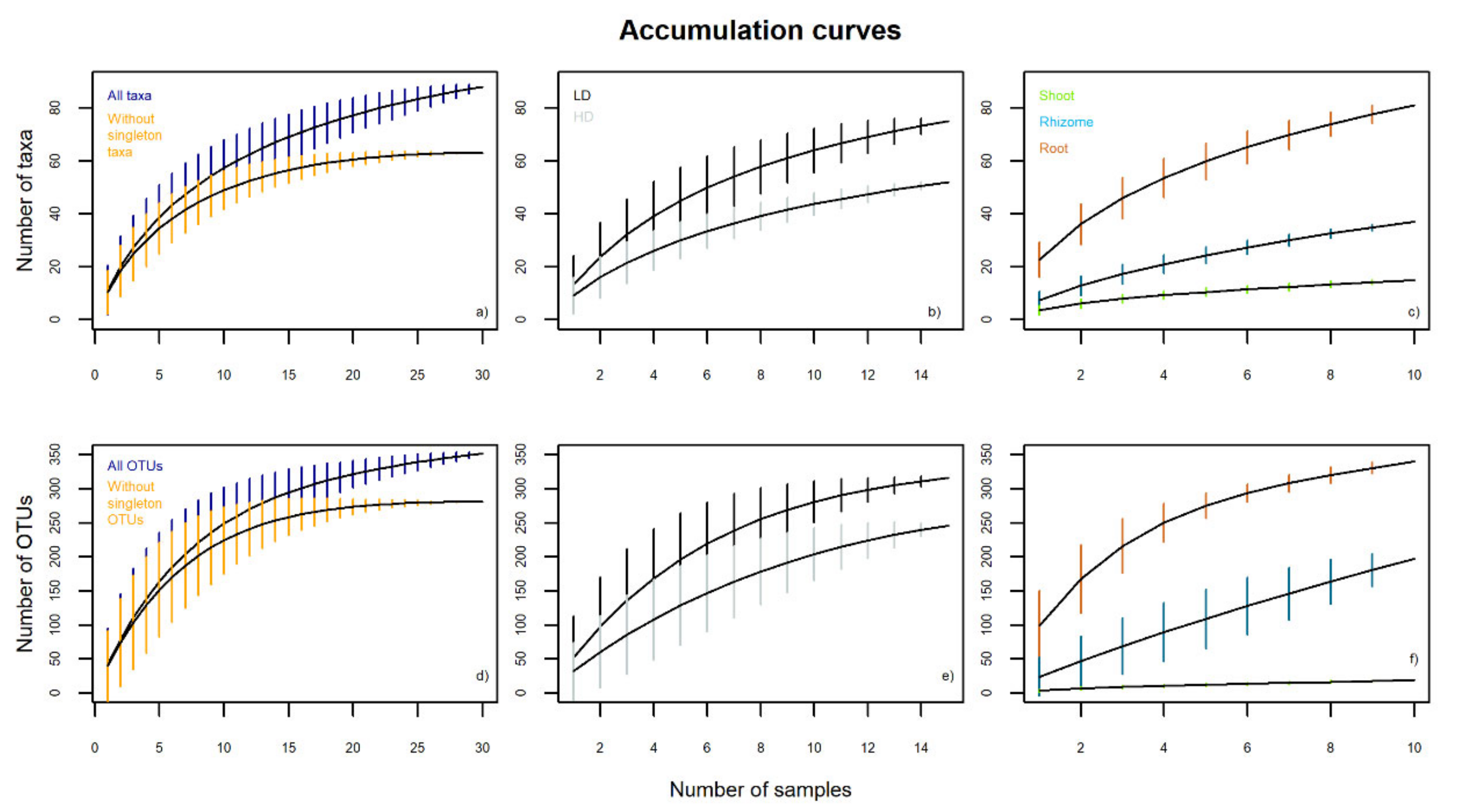
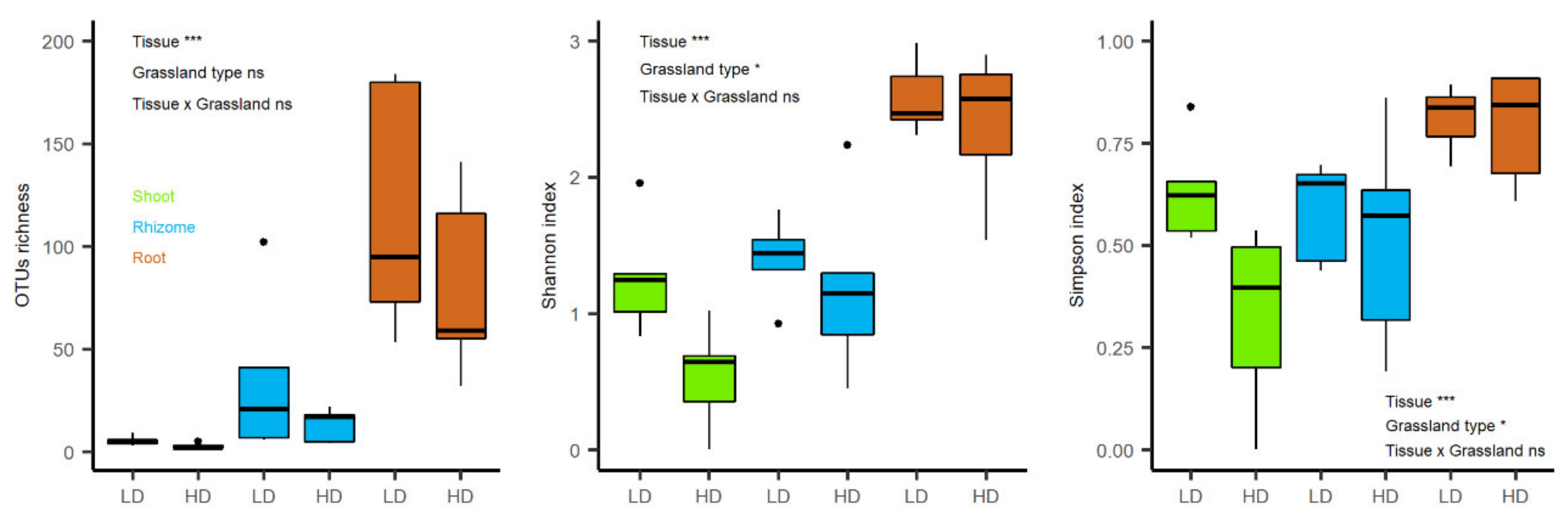
3.2.1. Fungal Endophytic Richness and Diversity
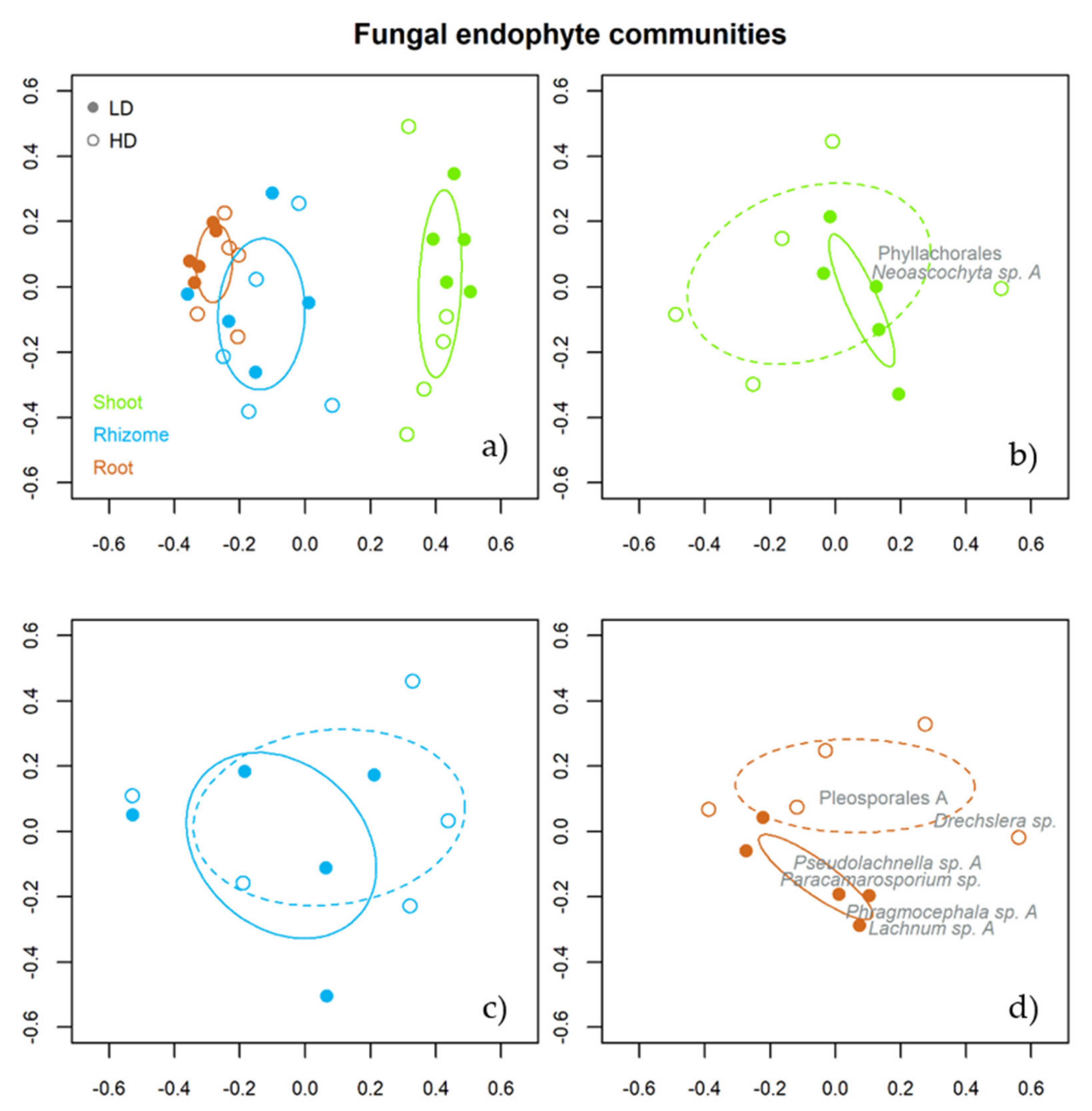
3.2.2. Taxonomic Assemblages for Grassland Types and Tissues
3.2.3. Indicator Species of the Fungal Assemblages
4. Discussion
4.1. The Mycobiome of B. rupestre According to the Metabarcoding Data
4.2. Culturing vs. Metabarcoding Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Match Taxon (NCBI) | Match Taxon (UNITE) | |||||||
|---|---|---|---|---|---|---|---|---|
| Accession Number | Greatest Percentage Identity (%) | Accession Number | Greatest Percentage Identity (%) | Taxon Proposed | Accession Number | |||
| 1 | Gerronema sp. | NR_166278 | 82.37 | Delicatula integrella | UDB034203 | 99.77 | Agaricales A | OK430888 |
| 2 | Gerronema sp. | NR_166278 | 78.9 | Mycena sp. | KT224934 | 89 | Agaricales B | OK430889 |
| 3 | Ramariopsis flavescens | NR_119913 | 85.06 | Agaricales | JX456916 | 95.2 | Agaricales C | OK430890 |
| 4 | Gerronema indigoticum | NR_166278 | 78.06 | Mycenaceae | KT224934 | 94.29 | Agaricales D | OK430891 |
| 5 | Laccaria aurantia | NR_154113 | 78.57 | Mycena floridula | MH856660 | 99.35 | Agaricales E | OK430892 |
| 6 | Radulotubus resupinatus | NR_153458 | 84.66 | Agaricomycetes | LR864837 | 99.32 | Agaricomycetes | OK430893 |
| 7 | Lachnellula hyalina | NR_165202 | 93.02 | Albotricha sp. | JN995639 | 100 | Albotricha sp. A | OK430894 |
| 8 | Lachnellula hyalina | NR_165202 | 91.59 | Albotricha sp. | JN995639 | 98.71 | Albotricha sp. B | OK430895 |
| 9 | Lachnellula hyalina | NR_165202 | 91.59 | Albotricha sp. | HM136666 | 100 | Albotricha sp. C | OK430896 |
| 10 | Funiliomyces biseptatus | NR_159862 | 96.39 | Acremonium sp. | MT911439 | 100 | Ascomycota A | OK430897 |
| 11 | Tricladium terrestre | NR_160144 | 93.67 | Ascomycota sp. | KR266584 | 93.67 | Ascomycota B | OK430898 |
| 12 | Auricularia scissa | NR_125807 | 80.48 | Oliveonia sp. | MT235652 | 97.16 | Auriculariales | OK430899 |
| 13 | Hydnum albidum | NR_164025 | 78.7 | Sistotrema sp. | KC965692 | 93.87 | Cantharellales | OK430900 |
| 14 | Codinaeae sp. | NR_168799 | 97.52 | Codinaea sp. | MT626587 | 98.35 | Chaetosphaeriaceae | OK430901 |
| 15 | Chalara hyalocuspica | NR_137568 | 91.25 | Chalara sp. | MK965778 | 98.33 | Chalara sp. | OK430902 |
| 16 | Cladophialophora tengchongensis | NR_172399 | 90.07 | Cladophialophora sp. | KP889848 | 100 | Cladophialophora sp. | OK430903 |
| 17 | Coccomyces pinicola | NR_158295 | 83.54 | Coccomyces dentatus | KU986782 | 93.82 | Coccomyces sp. | OK430904 |
| 18 | Conlarium duplumascospora | NR_138382 | 94.9 | Conlarium sp. | MK164654 | 96.85 | Conlarium sp. | OK430905 |
| 19 | Laburnicola centaurear | NR_154131 | 93.6 | Laburnicola sp. | MK018553 | 97.95 | Didymosphaeriaceae | OK430906 |
| 20 | Pseudoseptoria collariana | NR_156560 | 97.63 | Pseudoseptoria donacis | MH859141 | 99.6 | Dothideales | OK430907 |
| 21 | Roussoella thailandica | NR_155717 | 80.56 | Dothideomycetes | KJ827952 | 95 | Dothideomycetes A | OK430908 |
| 22 | Pirozynskiella laurisilvica | NR_153488 | 91 | Capnodiales | KX403688 | 91 | Dothideomycetes B | OK430909 |
| 23 | Drechslera sp. | NR_164466 | 92.89 | Drechslera sp. | MT816433 | 99.6 | Drechslera sp. | OK430910 |
| 24 | Entoloma luteofuscum | NR_152900 | 95.24 | Entoloma conferendum | MT741744 | 100 | Entoloma sp. | OK430911 |
| 25 | Epicoccum phragmospora | NR_165920 | 99.19 | Epicoccum sp. | MW054426 | 100 | Epicoccum sp. | OK430912 |
| 26 | Falciphora oryzae | NR_153972 | 98.86 | Falciphora oryzae | MH201898 | 99.23 | Falciphora sp. | OK430913 |
| 27 | Glarea lozoyensis | NR_137138 | 98.48 | Glarea sp. | KT268823 | 100 | Glarea sp. | OK430914 |
| 28 | Glarea lozoyensis | NR_137138 | 95.96 | Glarea sp. | KF617491 | 100 | Helotiaceae | OK430915 |
| 29 | Loramyces macrosporus | NR_138379 | 89.8 | Loramyces sp. | KF618060 | 99.58 | Helotiales A | OK430916 |
| 30 | Loramyces macrosporus | NR_138379 | 89.07 | Mollisia sp. | UDB0778890 | 99.59 | Helotiales B | OK430917 |
| 31 | Triposporium cycadicola | NR_156587 | 89.71 | Hymenoscyphus sp. | HQ625461 | 99.58 | Helotiales C | OK430918 |
| 32 | Bisporella shangrilana | NR_153628 | 97.02 | Helotiales | LR863043 | 99.58 | Helotiales D | OK430919 |
| 33 | Hyaloscypha finlandica | NR_121279 | 92.27 | Hyaloscypha vraolstadiae | KC876248 | 96.23 | Hyaloscyphaceae | OK430920 |
| 34 | Lachnellula hyalina | NR_165202 | 91.12 | Lachnum sp. | MT913626 | 96.61 | Lachnum sp. A | OK430921 |
| 35 | Lachnum fusiforme | NR_154122 | 89.91 | Lachnum sp. | MK808968 | 97.45 | Lachnum sp. B | OK430922 |
| 36 | Proliferodiscus sp. | NR_164304 | 86.67 | Lachnum sp. | MH628228 | 99.57 | Lachnum sp. C | OK430923 |
| 37 | Leohumicola minima | NR_121307 | 100 | Leohumicola sp. | FM999596 | 100 | Leohumicola sp. | OK430924 |
| 38 | Variabilispora flava | NR_165906 | 86.83 | Helotiales | AY969994 | 95.65 | Leotiomycetes | OK430925 |
| 39 | Menispora ciliata | NR_171740 | 99.5 | Menispora ciliata | MH860017 | 99.12 | Menispora sp. | OK430926 |
| 40 | Microdochium phragmitis | NR_132916 | 100 | Microdochium phragmitis | MN077456 | 100 | Microdochium sp. | OK430927 |
| 41 | Phialocephala sp. | NR_119482 | 90.38 | Phialocephala sp. | MG066460 | 97.88 | Mollisiaceae A | OK430928 |
| 42 | Mollisia scopiformis | NR_119460 | 93.22 | Phialocephala sp. | MK808244 | 98.72 | Mollisiaceae B | OK430929 |
| 43 | Mollisia monilioides | NR_171261 | 96.22 | Phialocephala sp. | MT911435 | 100 | Mollisiaceae C | OK430930 |
| 44 | Mollisia prismatica | NR_171258 | 91.9 | Phialocephala sp. | MK965789 | 99.57 | Mollisiaceae D | OK430931 |
| 45 | Mollisia asteliae | NR_173037 | 95.15 | Mollisia sp. | MH633925 | 100 | Mollisiaceae E | OK430932 |
| 46 | Mollisia diesbachiana | NR_171259 | 96.77 | Mollisia sp. | MT179560 | 100 | Mollisiaceae F | OK430933 |
| 47 | Mortierella gemmifera | NR_111559 | 94.81 | Mortierellaceae | LR863033 | 99.43 | Mortierella sp. | OK430934 |
| 48 | Podila horticola | NR_111572 | 99.09 | Mortierella sp. | DQ388818 | 99.7 | Mortierellaceae | OK430935 |
| 49 | Mycena fulgoris | NR_163300 | 93.29 | Mycena sp. | JF519186 | 98.4 | Mycena sp. A | OK430936 |
| 50 | Mycena fulgoris | NR_163300 | 93.29 | Mycena sp. | MK961197 | 99.67 | Mycena sp. B | OK430937 |
| 51 | Mycena fulgoris | NR_163300 | 93.31 | Mycena arcangeliana | JF908402 | 99.35 | Mycena sp. C | OK430938 |
| 52 | Mycena fulgoris | NR_163300 | 87.99 | Mycena sp. | UDB020406 | 100 | Mycena sp. D | OK430939 |
| 53 | Mycena fulgoris | NR_163300 | 89.64 | Mycena sp. | HQ625481 | 99.32 | Mycena sp. E | OK430940 |
| 54 | Cercospora coniogrammes | NR_147260 | 97.89 | Cercospora sp. | MN970528 | 97.89 | Mycosphaerellaceae | OK430941 |
| 55 | Myrmecridium spartii | NR_155376 | 96.25 | Myrmecridium sp. | MW133876 | 98.32 | Myrmecridium sp. | OK430942 |
| 56 | Pseudomassariella vexata | NR_164217 | 87.78 | Fusidium sp. | HG936132 | 100 | Nectriaceae | OK430943 |
| 57 | Neoascochyta europaea | NR_136131 | 97.03 | Neoascochyta europaea | MK190674 | 97.17 | Neoascochyta sp. A | OK430944 |
| 58 | Neoascochyta soli | NR_158269 | 100 | Neoascochyta paspali | MT373264 | 100 | Neoascochyta sp. B | OK430945 |
| 59 | Ophiosphaerella aquatica | NR_154352 | 89.96 | Ophiosphaerella sp. | MH063799 | 98.38 | Ophiosphaerella sp. | OK430946 |
| 60 | Paracamarosporium fagi | NR_154318 | 99.18 | Paracamarosporium fagi | MN244221 | 99.18 | Paracamarosporium sp. | OK430947 |
| 61 | Parasola parvula | NR_160509 | 94.43 | Parasola schroeteri | UDB024639 | 99.67 | Parasola sp. | OK430948 |
| 62 | Periconia epilithographicola | NR_157477 | 94.55 | Periconia sp. | MG543950 | 100 | Periconia sp. | OK430949 |
| 63 | Pezicula rhizophila | NR_155659 | 100 | Pezicula sp. | MN385513 | 100 | Pezicula sp. | OK430950 |
| 64 | Parastagonospora poagena | NR_168147 | 97.94 | Parastagonospora nodorum | MN313349 | 99.17 | Phaeosphaeriaceae | OK430951 |
| 65 | Phragmocephala garethjonessi | NR_147636 | 92.21 | Phragmocephala garethjonessi | MN660752 | 92.21 | Phragmocephala sp. A | OK430952 |
| 66 | Phragmocephala garethjonessi | NR_147636 | 90.2 | Phragmocephala atra | MN660752 | 90.61 | Phragmocephala sp. B | OK430953 |
| 67 | Phyllachora sp. | NR_156611 | 85 | Phyllachora graminis | AF257111 | 96.68 | Phyllachorales | OK430954 |
| 68 | Pleotrichocladium opacum | NR_155696 | 94.21 | Pleosporales | KY228531 | 99.58 | Pleosporales A | OK430955 |
| 69 | Camposporium multiseptatum | NR_171863 | 100 | Camposporium sp. | MN758889 | 100 | Pleosporales B | OK430956 |
| 70 | Anteaglonium rubescens | NR_164489 | 89.92 | Lophiostoma sp. | EU977287 | 93.17 | Pleosporales C | OK430957 |
| 71 | Pseudolachnella fusiformis | NR_154280 | 94.24 | Pseudolachnella fusiformis | AB934080 | 94.24 | Pseudolachnella sp. A | OK430958 |
| 72 | Pseudolachnella fusiformis | NR_154280 | 93.78 | Pseudolachnella fusiformis | AB934080 | 93.77 | Pseudolachnella sp. B | OK430959 |
| 73 | Puccinia aizazii | NR_158929 | 99.2 | Puccinia brachypodii | GQ457303 | 100 | Puccinia sp. | OK430960 |
| 74 | Plectosphaerella niemeijerarum | NR_156677 | 88.24 | Plectosphaerellaceae | MK762215 | 88.23 | Sordariomycetes A | OK430961 |
| 75 | Phaeoacrenonium cinereum | NR_ 132066 | 80.62 | Sordaryomycetes | KP050604 | 80.62 | Sordariomycetes B | OK430962 |
| 76 | Cordana pauciseptata | NR_154771 | 88.98 | Sordariales | UDB067041 | 96.69 | Sordariomycetes C | OK430963 |
| 77 | Neomyrmecridium guizhouense | NR_170024 | 82.45 | Sordariomycetes | LR865231 | 100 | Sordariomycetes D | OK430964 |
| 78 | Atractospora verruculosa | NR_153542 | 89.53 | Sordariales | EU754966 | 100 | Sordariomycetes E | OK430965 |
| 79 | Subulicistidium oberwinkleri | NR_159060 | 86.42 | Trechisporales | JF519283 | 100 | Trechisporales A | OK430966 |
| 80 | Subulicystidium oberwinkleri | NR_159060 | 80.53 | Trechisporales | UDB020436 | 83.77 | Trechisporales B | OK430967 |
| 81 | Trichoderma hispanicum | NR_138451 | 99.25 | Trichoderma koningii | MT781958 | 99.24 | Trichoderma sp. | OK430968 |
| 82 | Corinarius hadrocroceus | NR_131854 | 79.62 | Tricholomataceae | KX115676 | 100 | Tricholomataceae A | OK430969 |
| 83 | Mycena seminau | NR_154170 | 88.82 | Tricholomataceae | MH016642 | 99.67 | Tricholomataceae B | OK430970 |
| 84 | Phialocephala humicola | NR_103570 | 87.7 | Chaetosphaeriales | HM136627 | 100 | Unidentified A | OK430971 |
| 85 | Rhodosporidiobolus fluvialis | NR_077089 | 93.65 | Agaricomycetes | UDB0327559 | 100 | Unidentified B | OK430972 |
| 86 | Mycosymbioces mycenaphila | NR_137807 | 85.06 | Helotiales | UDB0779249 | 100 | Unidentified C | OK430973 |
| 87 | Mollisia monilioides | NR_171261 | 90.34 | Helotiales | KT203037 | 96.61 | Unidentified D | OK430974 |
| 88 | Linteromyces quintiniae | NR_171989 | 86.25 | Xylariales | MN218782 | 99.62 | Xylariales | OK430975 |
Appendix B
| ROOT | RHIZOME | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Endophyte Taxon | Relative Abundance (%) | Reads | OTUs | Infected Plants | Relative Abundance (%) | Reads | OTUs | Infected Plants | ||||||||
| LD | HD | LD | HD | LD | HD | LD | HD | LD | HD | LD | HD | LD | HD | LD | HD | |
| Helotiaceae | 24.51 | 31.09 | 64,132 | 51,968 | 114 | 116 | 2 | 4 | 25.95 | 16.49 | 12,267 | 4897 | 94 | 18 | 2 | 4 |
| Lachnum sp. A | 35.08 | 12.5 | 91,790 | 20,889 | 36 | 20 | 5 | 5 | 29.13 | 9.01 | 13,771 | 2676 | 5 | 7 | 4 | 4 |
| Albotricha sp. A | 1.71 | 18.83 | 4465 | 31,473 | 7 | 6 | 3 | 3 | 5.23 | 50.58 | 2473 | 15,018 | 4 | 6 | 3 | 3 |
| Helotiales A | 12.43 | 6.03 | 32,534 | 10,072 | 40 | 28 | 5 | 5 | 10.98 | 2.77 | 5188 | 823 | 25 | 1 | 2 | 3 |
| Agaricales A | 3.55 | 9.65 | 9281 | 16,124 | 3 | 3 | 2 | 4 | 0.05 | 0 | 24 | 0 | 2 | 0 | 1 | 0 |
| Mycena sp. A | 2.03 | 0.17 | 5323 | 289 | 10 | 2 | 4 | 1 | 20.88 | 0 | 9870 | 0 | 1 | 0 | 1 | 0 |
| Mollisiaceae C | 4.17 | 0.45 | 10,913 | 745 | 2 | 1 | 4 | 1 | 0.42 | 0.04 | 198 | 13 | 2 | 1 | 1 | 1 |
| Pleosporales A | 0.56 | 4.04 | 1476 | 6751 | 2 | 4 | 3 | 5 | 0 | 0.04 | 0 | 12 | 0 | 1 | 0 | 1 |
| Glarea sp. | 0.41 | 3.94 | 1060 | 6589 | 2 | 1 | 2 | 1 | 0 | 0.29 | 0 | 86 | 0 | 1 | 0 | 1 |
| Mollisiaceae B | 0.43 | 1.89 | 1118 | 3161 | 1 | 3 | 2 | 3 | 2.35 | 1.44 | 1111 | 429 | 1 | 3 | 2 | 2 |
| Mollisiaceae D | 0.89 | 1.07 | 2330 | 1782 | 1 | 2 | 2 | 1 | 0.78 | 3.56 | 369 | 1056 | 1 | 2 | 2 | 2 |
| Chaetosphaeriaceae | 1.76 | 0 | 4608 | 0 | 4 | 0 | 1 | 0 | 0.07 | 0 | 33 | 0 | 1 | 0 | 1 | 0 |
| Mycena sp. B | 0 | 2.08 | 0 | 3479 | 0 | 3 | 0 | 1 | 0 | 3.34 | 0 | 993 | 0 | 1 | 0 | 1 |
| Tricholomataceae B | 0 | 1.48 | 0 | 2474 | 0 | 1 | 0 | 2 | 0 | 4.21 | 0 | 1251 | 0 | 1 | 0 | 1 |
| Lachnum sp. B | 0.38 | 1.27 | 1007 | 2119 | 11 | 7 | 4 | 4 | 0.87 | 0.14 | 411 | 43 | 1 | 1 | 1 | 1 |
| Cantharellales | 1.3 | 0 | 3397 | 0 | 2 | 0 | 1 | 0 | ||||||||
| Parasola sp. | 0 | 0.9 | 0 | 1503 | 0 | 1 | 0 | 1 | 0 | 5.81 | 0 | 1725 | 0 | 3 | 0 | 1 |
| Unidentified A | 1.21 | 0.01 | 3174 | 19 | 2 | 1 | 1 | 1 | ||||||||
| Ophiosphaerella sp. | 0.96 | 0.32 | 2513 | 535 | 2 | 1 | 2 | 1 | 0.11 | 0.06 | 50 | 17 | 1 | 1 | 1 | 1 |
| Mollisiaceae A | 0.88 | 0.4 | 2309 | 666 | 4 | 3 | 4 | 5 | 0.03 | 0.28 | 13 | 83 | 1 | 1 | 1 | 1 |
| Drechslera sp. | 0.03 | 1.43 | 87 | 2388 | 2 | 2 | 2 | 5 | ||||||||
| Paracamarosporium sp. | 0.92 | 0 | 2419 | 0 | 1 | 0 | 4 | 0 | 0.06 | 0 | 28 | 0 | 1 | 0 | 2 | 0 |
| Agaricales C | 0.58 | 0.07 | 1514 | 114 | 1 | 1 | 2 | 1 | ||||||||
| Auriculariales | 0.5 | 0 | 1308 | 0 | 2 | 0 | 1 | 0 | ||||||||
| Tricholomataceae A | 0.48 | 0 | 1266 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Unidentified B | 0.13 | 0.43 | 340 | 718 | 1 | 1 | 3 | 2 | 0.19 | 0.29 | 90 | 86 | 1 | 1 | 1 | 1 |
| Pseudolachnella sp. B | 0.42 | 0.04 | 1097 | 72 | 1 | 1 | 3 | 2 | 0.11 | 0 | 51 | 0 | 1 | 0 | 1 | 0 |
| Trichoderma sp. | 0.41 | 0.01 | 1076 | 15 | 1 | 1 | 2 | 1 | ||||||||
| Didymosphaeriaceae | 0 | 0.58 | 0 | 963 | 0 | 2 | 0 | 1 | ||||||||
| Conlarium sp. | 0.35 | 0 | 925 | 0 | 1 | 0 | 3 | 0 | 0.07 | 0 | 35 | 0 | 1 | 0 | 1 | 0 |
| Helotiales C | 0.34 | 0 | 900 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Phragmocephala sp. A | 0.28 | 0 | 734 | 0 | 2 | 0 | 4 | 0 | ||||||||
| Agaricales B | 0.26 | 0.02 | 675 | 32 | 2 | 1 | 1 | 1 | ||||||||
| Menispora sp. | 0.27 | 0 | 702 | 0 | 1 | 0 | 2 | 0 | ||||||||
| Cladophialophora sp. | 0.23 | 0.04 | 604 | 74 | 1 | 1 | 3 | 3 | 0.04 | 0 | 18 | 0 | 1 | 0 | 1 | 0 |
| Pleosporales B | 0.11 | 0.2 | 299 | 341 | 1 | 2 | 1 | 2 | ||||||||
| Pseudolachnella sp. A | 0.23 | 0.01 | 606 | 11 | 2 | 1 | 4 | 1 | ||||||||
| Mortierellaceae | 0.11 | 0 | 283 | 0 | 2 | 0 | 1 | 0 | 0.43 | 0 | 201 | 0 | 3 | 0 | 1 | 0 |
| Mollisiaceae F | 1.01 | 0 | 476 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Chalara sp. | 0.06 | 0.03 | 168 | 57 | 2 | 2 | 3 | 2 | 0.4 | 0.09 | 187 | 26 | 1 | 1 | 1 | 1 |
| Sordariomycetes D | 0.17 | 0 | 435 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Helotiales B | 0.03 | 0 | 87 | 0 | 1 | 0 | 1 | 0 | 0.21 | 0.73 | 99 | 218 | 1 | 1 | 1 | 1 |
| Agaricomycetes | 0.12 | 0 | 325 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Ascomycota B | 0.1 | 0 | 266 | 0 | 1 | 0 | 1 | 0 | 0.09 | 0 | 41 | 0 | 1 | 0 | 1 | 0 |
| Mollisiaceae E | 0 | 0.13 | 0 | 216 | 0 | 1 | 0 | 1 | 0 | 0.31 | 0 | 91 | 0 | 1 | 0 | 1 |
| Microdochium sp. | 0.07 | 0 | 187 | 0 | 1 | 0 | 3 | 0 | 0.25 | 0 | 119 | 0 | 1 | 0 | 1 | 0 |
| Mortierella sp. | 0.12 | 0 | 305 | 0 | 2 | 0 | 1 | 0 | ||||||||
| Sordariomycetes B | 0.11 | 0 | 297 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Pezicula sp. | 0 | 0.16 | 0 | 272 | 0 | 1 | 0 | 2 | ||||||||
| Coccomyces sp. | 0.09 | 0.02 | 227 | 31 | 1 | 1 | 2 | 1 | ||||||||
| Albotricha sp. B | 0 | 0.13 | 0 | 217 | 0 | 2 | 0 | 1 | 0 | 0.06 | 0 | 19 | 0 | 1 | 0 | 1 |
| Leohumicola sp. | 0.06 | 0.03 | 166 | 57 | 1 | 1 | 4 | 2 | 0.03 | 0 | 13 | 0 | 1 | 0 | 1 | 0 |
| Lachnum sp. C | 0.09 | 0 | 228 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Pleosporales C | 0 | 0.13 | 0 | 213 | 0 | 1 | 0 | 3 | ||||||||
| Nectriaceae | 0.08 | 0 | 212 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Phragmocephala sp. B | 0.06 | 0.03 | 147 | 45 | 1 | 1 | 1 | 2 | ||||||||
| Sordariomycetes C | 0 | 0.11 | 0 | 188 | 0 | 1 | 0 | 2 | ||||||||
| Ascomycota A | 0.06 | 0 | 168 | 0 | 1 | 0 | 3 | 0 | ||||||||
| Sordariomycetes E | 0.06 | 0 | 167 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Trechisporales B | 0 | 0.09 | 12 | 151 | 1 | 1 | 1 | 1 | ||||||||
| Trechisporales A | 0.03 | 0 | 76 | 0 | 1 | 0 | 2 | 0 | 0 | 0.28 | 0 | 82 | 0 | 1 | 0 | 1 |
| Agaricales D | 0.06 | 0 | 153 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Myrmecridium sp. | 0.06 | 0 | 149 | 0 | 1 | 0 | 2 | 0 | ||||||||
| Dothideomycetes A | 0.04 | 0 | 117 | 0 | 1 | 0 | 1 | 0 | 0.06 | 0 | 30 | 0 | 1 | 0 | 1 | 0 |
| Entoloma sp. | 0.06 | 0 | 144 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Falciphora sp. | 0.05 | 0 | 141 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Leotiomycetes | 0.05 | 0 | 139 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Mycena sp. D | 0.05 | 0.01 | 120 | 14 | 1 | 1 | 1 | 1 | ||||||||
| Mycena sp. E | 0 | 0.08 | 0 | 133 | 0 | 1 | 0 | 1 | ||||||||
| Unidentified D | 0.05 | 0 | 132 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Agaricales E | 0.05 | 0 | 128 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Dothideomycetes B | 0 | 0.08 | 0 | 127 | 0 | 1 | 0 | 1 | ||||||||
| Unidentified C | 0.05 | 0 | 124 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Albotricha sp. C | 0.05 | 0 | 123 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Helotiales D | 0.05 | 0 | 121 | 0 | 1 | 0 | 1 | 0 | ||||||||
| Neoascochyta sp. B | 0.01 | 0 | 17 | 0 | 1 | 0 | 1 | 0 | 0.22 | 0 | 102 | 0 | 1 | 0 | 3 | 0 |
| Mycena sp. C | 0.03 | 0 | 86 | 0 | 1 | 0 | 1 | 0 | 0 | 0.07 | 0 | 22 | 0 | 1 | 0 | 1 |
| Hyaloscyphaceae | 0.03 | 0.02 | 73 | 27 | 1 | 1 | 2 | 1 | ||||||||
| Periconia sp. | 0.02 | 0 | 51 | 0 | 1 | 0 | 2 | 0 | ||||||||
| Epicoccum sp. | 0 | 0 | 12 | 0 | 1 | 0 | 1 | 0 | 0 | 0.09 | 0 | 26 | 0 | 1 | 0 | 1 |
| Phaeosphaeriaceae | 0 | 0.02 | 0 | 28 | 0 | 1 | 0 | 1 | ||||||||
| Xylariales sp. | 0.04 | 0.01 | 105 | 12 | 1 | 1 | 2 | 1 | ||||||||
| 100 | 100 | 261673 | 167154 | 100 | 100 | 47268 | 29692 | |||||||||
References
- Bud, R. Biotechnology in the Twentieth Century. Soc. Stud. Sci. 1991, 21, 415–457. [Google Scholar] [CrossRef]
- United Nations Convention on Biological Diversity; United Nations: Rio de Janeiro, Brazil, 1992.
- Carthey, A.J.R.; Blumstein, D.T.; Tetu, S.G.; Gillings, M.R.; Gallagher, R.V. Conserving the holobiont. Funct. Ecol. 2019, 34, 764–776. [Google Scholar] [CrossRef]
- Guerrero, R.; Margulis, L.; Berlanga, M. Symbiogenesis: The holobiont as a unit of evolution. Int. Microbiol. 2013, 16, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Tikhonovich, I.A.; Provorov, N.A. Microbiology is the basis of sustainable agriculture: An opinion. Ann. Appl. Biol. 2011, 159, 155–168. [Google Scholar] [CrossRef]
- Saikkonen, K.; Lehtonen, P.; Helander, M.; Koricheva, J.; Faeth, S.H. Model systems in ecology: Dissecting the endophyte-grass literature. Trends Plant Sci. 2006, 11, 428–433. [Google Scholar] [CrossRef]
- Saikkonen, K.; Faeth, S.H.; Helander, M.; Sullivan, T.J. Fungal endophytes: A continuum of interactions with host plants. Annu. Rev. Ecol. Syst. 1998, 29, 319–343. [Google Scholar] [CrossRef]
- Hume, D.E.; Sewell, J.C. Agronomic advantages conferred by endophyte infection of perennial ryegrass (Lolium perenne L.) and tall fescue (Festuca arundinacea Schreb.) in Australia. Crop Pasture Sci. 2014, 65, 747–757. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binet, M.N.; Sage, L.; Malan, C.; Ement, J.C.C.L.; Redecker, D.; Wipf, D.; Geremia, R.A.; Lavorel, S.; Mouhamadou, B. Effects of mowing on fungal endophytes and arbuscular mycorrhizal fungi in subalpine grasslands. Fungal Ecol. 2013, 6, 248–255. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Hyde, K.D.; Soytong, K. The fungal endophyte dilemma. Fungal Divers. 2008, 33, 133–173. [Google Scholar]
- Verma, V.C.; Gange, A.C. Advances in Endophytic Research; Springer: London, UK, 2014; ISBN 9788132215752. [Google Scholar]
- Chi, W.C.; Chen, W.; He, C.C.; Guo, S.Y.; Cha, H.J.; Tsang, L.M.; Ho, T.W.; Pang, K.L. A highly diverse fungal community associated with leaves of the mangrove plant Acanthus ilicifolius var. xiamenensis revealed by isolation and metabarcoding analyses. PeerJ 2019, 7, e7293. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2019, 47, D259–D264. [Google Scholar] [CrossRef] [PubMed]
- Ebach, M.C.; Valdecasas, A.G.; Wheeler, Q.D. Impediments to taxonomy and users of taxonomy: Accessibility and impact evaluation. Cladistics 2011, 27, 550–557. [Google Scholar] [CrossRef]
- Sun, X.; Guo, L.D. Endophytic fungal diversity: Review of traditional and molecular techniques. Mycology 2012, 3, 65–76. [Google Scholar] [CrossRef]
- Li, W. Analysis and comparison of very large metagenomes with fast clustering and functional annotation. BMC Bioinform. 2009, 10, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bricca, A.; Tardella, F.M.; Tolu, F.; Goia, I.; Ferrara, A.; Catorci, A. Disentangling the effects of disturbances from those of dominant tall grass features in driving the functional variation of restored grassland in a sub-mediterranean context. Diversity 2020, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Bobbink, R.; Willems, J.H. Increasing dominance of Brachypodium pinnatum (L.) Beauv. in chalk grasslands: A threat to a species-rich ecosystem. Biol. Conserv. 1987, 40, 301–314. [Google Scholar] [CrossRef]
- Catorci, A.; Cesaretti, S.; Gatti, R.; Ottaviani, G. Abiotic and biotic changes due to spread of Brachypodium genuense (DC.) Roem. & Schult. in sub-Mediterranean meadows. Community Ecol. 2011, 12, 117–125. [Google Scholar] [CrossRef]
- Bąba, W.; Kurowska, M.; Kompała-Baba, A.; Wilczek, A.; Długosz, J.; Szarejko, I. Genetic diversity of the expansive grass Brachypodium pinnatum in a changing landscape: Effect of habitat age. Flora 2012, 207, 346–353. [Google Scholar] [CrossRef]
- Buckland, S.M.; Thompson, K.; Hodgson, J.G.; Grime, J.P. Grassland invasions: Effects of manipulations of climate and management. J. Appl. Ecol. 2001, 38, 301–309. [Google Scholar]
- Durán, M.; Canals, R.M.; Sáez, J.L.; Ferrer, V.; Lera-López, F. Disruption of traditional land use regimes causes an economic loss of provisioning services in high-mountain grasslands. Ecosyst. Serv. 2020, 46, 101200. [Google Scholar] [CrossRef]
- Múgica, L.; Canals, R.M.; San Emeterio, L.; Peralta, J. Decoupling of traditional burnings and grazing regimes alters plant diversity and dominant species competition in high-mountain grasslands. Sci. Total Environ. 2021, 790, 147917. [Google Scholar] [CrossRef] [PubMed]
- San Emeterio, L.; Durán, M.; Múgica, L.; Jiménez, J.J.; Canals, R.M. Relating the spatial distribution of a tall-grass to fertility islands in a temperate mountain grassland. Soil Biol. Biochem. 2021, 163, 108455. [Google Scholar] [CrossRef]
- Catorci, A.; Cesaretti, S.; Tardella, F.M. Effect of tall-grass invasion on the flowering-related functional pattern of submediterranean hay-meadows. Plant Biosyst. 2014, 148, 1127–1137. [Google Scholar] [CrossRef]
- Canals, R.M.; Pedro, J.; Rupérez, E.; San-Emeterio, L. Nutrient pulses after prescribed winter fires and preferential patterns of N uptake may contribute to the expansion of Brachypodium pinnatum (L.) P. Beauv. in highland grasslands. Appl. Veg. Sci. 2014, 17, 419–428. [Google Scholar] [CrossRef]
- Canals, R.M.; San Emeterio, L.; Durán, M.; Múgica, L. Plant-herbivory feedbacks and selective allocation of a toxic metal are behind the stability of degraded covers dominated by Brachypodium pinnatum in acidic soils. Plant Soil 2017, 415, 373–386. [Google Scholar] [CrossRef]
- Bąba, W.; Kalaji, H.M.; Kompała-Bąba, A.; Goltsev, V. Acclimatization of photosynthetic apparatus of tor grass (Brachypodium pinnatum) during expansion. PLoS ONE 2016, 11, e0156201. [Google Scholar] [CrossRef] [Green Version]
- Tardella, F.M.; Malatesta, L.; Goia, I.G.; Catorci, A. Effects of long-term mowing on coenological composition and recovery routes of a Brachypodium rupestre-invaded community: Insight into the restoration of sub-Mediterranean productive grasslands. Rend. Lincei. Sci. Fis. Nat. 2018, 29, 329–341. [Google Scholar] [CrossRef]
- Meijer, G.; Leuchtmann, A. Multistrain infections of the grass Brachypodium sylvaticum by its fungal endophyte Epichloe sylvatica. New Phytol. 1999, 141, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Miwa, E.; Okane, I.; Ishiga, Y.; Sugawara, K.; Yamaoka, Y. Confirmation of taxonomic status of an Epichloë species on Brachypodium sylvaticum in Japan. Mycoscience 2017, 58, 147–153. [Google Scholar] [CrossRef]
- Zabalgogeazcoa, I.; Ciudad, A.G.; Leuchtmann, A.; Vázquez-de-Aldana, B.R.; Criado, B.G. Effects of choke disease in the grass Brachypodium phoenicoides. Plant Pathol. 2008, 57, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-de-Aldana, B.R.; Zabalgogeazcoa, I.; Garcia-Ciudad, A.; Garcia-Criado, B. Ergovaline occurrence in grasses infected by fungal endophytes of semi-arid pastures in Spain. J. Sci. Food Agric. 2003, 83, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Leuchtmann, A.; Schardl, C.L. Mating compatibility and phylogenetic relationships among two new species of Epichloë and other congeneric European species. Mycol. Res. 1998, 102, 1169–1182. [Google Scholar] [CrossRef]
- Durán, M.; San Emeterio, L.; Múgica, L.; Zabalgogeazcoa, I.; Vázquez-de-Aldana, B.R.; Canals, R.M. Disruption of traditional grazing and fire regimes shape the fungal endophyte assemblages of the tall-grass Brachypodrium rupestre. Front. Microbiol. 2021, 12, 679729. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Ed.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Niu, B.; Gao, Y.; Fu, L.; Li, W. CD-HIT Suite: A web server for clustering and comparing biological sequences. Bioinformatics 2010, 26, 680–682. [Google Scholar] [CrossRef]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding needles in haystacks: Linking scientific names, reference specimens and molecular data for Fungi. Database 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Turenne, C.Y.; Sanche, S.E.; Hoban, D.J.; Karlowsky, J.A.; Kabani, A.M. Rapid identification of fungi by using the ITS2 genetic region and automated fluorescent capillary electrophoresis system. J. Clin. Microbiol. 1999, 1846–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Caporaso, J.G. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2011, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, A.J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; Hara, R.B.O.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 17 October 2021).
- Roberts, D.W. Labdsv: Ordination and Multivariate Analysis for Ecology. Available online: https://cran.r-project.org/web/packages/labdsv/index.html (accessed on 17 October 2021).
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Sun, X.; Kosman, E.; Sharon, O.; Ezrati, S.; Sharon, A. Significant host- and environment-dependent differentiation among highly sporadic fungal endophyte communities in cereal crops-related wild grasses. Environ. Microbiol. 2020, 22, 3357–3374. [Google Scholar] [CrossRef]
- Harrison, J.G.; Griffin, E.A. The diversity and distribution of endophytes across biomes, plant phylogeny and host tissues: How far have we come and where do we go from here? Environ. Microbiol. 2020, 22, 2107–2123. [Google Scholar] [CrossRef] [Green Version]
- Wearn, J.A.; Sutton, B.C.; Morley, N.J.; Gange, A.C. Species and organ specificity of fungal endophytes in herbaceous grassland plants. J. Ecol. 2012, 100, 1085–1092. [Google Scholar] [CrossRef]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Herrera, J.; Khidir, H.H.; Eudy, D.M.; Porras-Alfaro, A.; Natvig, D.O.; Sinsabaugh, R.L. Shifting fungal endophyte communities colonize Bouteloua gracilis: Effect of host tissue and geographical distribution. Mycologia 2010, 102, 1012–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudgers, J.A.; Kivlin, S.N.; Whitney, K.D.; Price, M.V.; Waser, N.M.; Harte, J. Responses of high-altitude graminoids and soil fungi to 20 years of experimental warming. Ecology 2014, 95, 1918–1928. [Google Scholar] [CrossRef] [PubMed]
- Frank, A.C.; Paola, J.; Guzm, S.; Shay, J.E. Transmission of bacterial endophytes. Microorganisms 2017, 5, 70. [Google Scholar] [CrossRef] [Green Version]
- Compant, S.; Saikkonen, K.; Mitter, B.; Campisano, A.; Mercado-Blanco, J. Editorial special issue: Soil, plants and endophytes. Plant Soil 2016, 405, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Márquez, S.S.; Bills, G.F.; Zabalgogeazcoa, I. The endophytic mycobiota of the grass Dactylis glomerata. Fungal Divers. 2007, 27 I, 171–195. [Google Scholar]
- Sánchez-Márquez, S.; Bills, G.F.; Acuña, L.D.; Zabalgogeazcoa, I. Endophytic mycobiota of leaves and roots of the grass Holcus lanatus. Fungal Divers. 2010, 41, 115–123. [Google Scholar] [CrossRef]
- Høyer, A.K.; Hodkinson, T.R. Hidden fungi: Combining culture-dependent and -independent DNA barcoding reveals inter-plant variation in species richness of endophytic root fungi in Elymus repens. J. Fungi 2021, 7, 466. [Google Scholar] [CrossRef] [PubMed]
- Tejesvi, M.V.; Ruotsalainen, A.L.; Markkola, A.M.; Pirttilä, A.M. Root endophytes along a primary succession gradient in northern Finland. Fungal Divers. 2010, 41, 125–134. [Google Scholar] [CrossRef]
- Hernández-Restrepo, M.; Groenewald, J.Z.; Elliott, M.L.; Canning, G.; McMillan, V.E.; Crous, P.W. Take-all or nothing. Stud. Mycol. 2016, 83, 19–48. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Restrepo, M.; Bezerra, J.D.P.; Tan, Y.P.; Wiederhold, N.; Crous, P.W.; Guarro, J.; Gené, J. Re-evaluation of Mycoleptodiscus species and morphologically similar fungi. Persoonia Mol. Phylogeny Evol. Fungi 2019, 42, 205–227. [Google Scholar] [CrossRef] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Tian, W.; Gao, Z.; Wang, G.; Zhao, H. Phylogenetic utility of rRNA ITS2 sequence-structure under functional constraint. Int. J. Mol. Sci. 2020, 21, 6395. [Google Scholar] [CrossRef] [PubMed]
- Mbareche, H.; Veillette, M.; Bilodeau, G.; Duchaine, C. Comparison of the performance of ITS1 and ITS2 as barcodes in amplicon-based sequencing of bioaerosols. PeerJ 2020, 2020, e8523. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.E.; Henk, D.A.; Eells, R.L.; Lutzoni, F.; Vilgalys, R. Diversity and phylogenetic affinities of foliar fungal endophytes in loblolly pine inferred by culturing and environmental PCR. Mycologia 2007, 99, 185–206. [Google Scholar] [CrossRef]
- Tanney, J.B.; Seifert, K.A. Mollisiaceae: An overlooked lineage of diverse endophytes. Stud. Mycol. 2020, 95, 293–380. [Google Scholar] [CrossRef]
- Tanney, J.B.; Douglas, B.; Seifert, K.A. Sexual and asexual states of some endophytic Phialocephala species of Picea. Mycologia 2016, 108, 255–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bills, G.F.; Platas, G.; Peláez, F.; Masurekar, P. Reclassification of a pneumocandin-producing anamorph, Glarea lozoyensis gen. et sp. nov., previously identified as Zalerion arboricola. Mycol. Res. 1999, 103, 179–192. [Google Scholar] [CrossRef]




| Study Site | ARPEA | URKULU | |
|---|---|---|---|
| Type of Grassland | LD = Low Diversity | HD = High Diversity | |
| General description | Location | −1°10′57″ W | –1°14′38″ W |
| 43°2′12″ N | 43°2′49″ N | ||
| Soil classification (WRB) | Cambic Umbrisol | Dystric Cambisol | |
| Altitude (m.a.s.l.) | 893 | 1256 | |
| Slope (%) | 40 | 45 | |
| Management | Burning recurrence | High 1–2 years | Low 6–7 years |
| Type of burning | Large grassland areas | Bush-to-bush | |
| Grazing level | Low to nonexistent | Moderate to high | |
| B. rupestre cover (%) | >80% | <25% |
| Match Taxon (NCBI) | Match Taxon (UNITE) | Taxon Proposed | GenBank Accession Number | |||||
|---|---|---|---|---|---|---|---|---|
| Accessio Number | Greatest Percentage Identity (%) | Accession Number | Greatest Percentage Identity (%) | |||||
| 1 | Lachnellula hyalina | NR_165202 | 90.11 | Albotricha sp. | HM136666 | 98.22 | Albotricha sp. | MW789554 |
| 2 | Codinaea paniculata | NR_166297 | 99.74 | Codinaea sp. | MT118230 | 99.74 | Codinaea sp. | MW789567 |
| 3 | Paracamarosporium sp. | NR_154318 | 94.28 | Paracamarosporium sp. | MT882131 | 97.6 | Didymosphaeriaceae | MW789559 |
| 4 | Drechslera sp. | NR_153992 | 94.43 | Drechslera sp. | UDB0174425 | 100 | Drechslera sp | MW789560 |
| 5 | Falciphora oryzae | NR_153972 | 96.69 | Falciphora sp. | UDB0162916 | 99.76 | Falciphora sp. | MW789558 |
| 6 | Glarea lozoyensis | NR_137138 | 96.18 | Glarea sp. | KF617491 | 99.58 | Helotiaceae | MW789565 |
| 7 | Ilyonectria leucospermi | NR_152889 | 99.36 | Ilyonectria crassa | MT294410 | 100 | Ilyonectria sp. | MW789566 |
| 8 | Lachnellula hualina | NR_165202 | 88.89 | Lachnum virgineum | MT133783 | 98.15 | Lachnum sp. | MW789564 |
| 9 | Microdochium phragmitis | NR_132916 | 100 | Microdochium phragmitis | MH861162 | 100 | Microdochium phragmitis | MW789562 |
| 10 | Mollisia asteliae | NR_173037 | 96.44 | Mollisia sp. | KJ188683 | 98.69 | Mollisia sp. | MW789555 |
| 11 | Phialocephala spaheroides | NR_121302 | 95.71 | Loramyces sp. | KF618060 | 99.36 | Mollisiaceae | MW789556 |
| 12 | Neoascochyta dactylidis | NR_170041 | 100 | Neoascochyta sp. | MT185527 | 100 | Neoascochyta sp. | MW789561 |
| 13 | Omnidemptus graminis | NR_164058 | 100 | Omnidemptus graminis | MK487758 | 100 | Omnidemptus graminis | MW789553 |
| 14 | Phialocephala sphaeroides | NR_121302 | 89 | Phialocephala sp. | JN995646 | 98.87 | Phialocephala sp. | MW789563 |
| 15 | Paraphaeosphaeria michotii | NR_155640 | 91.41 | Pleosporales | MN450621 | 100 | Pleosporales | MW789557 |
| Type of Grassland | Tissue | ||||||
|---|---|---|---|---|---|---|---|
| Shoot | Rhizome | Root | |||||
| Metabarcoding method | Reads | LD | 313,621 | 4680 | 47,268 | 261,673 | |
| HD | 200,050 | 3204 | 29,692 | 167,154 | |||
| Total | 513,671 | 7884 | 76,960 | 428,827 | |||
| OTUs | LD | 316 | 12 | 165 | 305 | ||
| HD | 246 | 11 | 58 | 236 | |||
| Total | 352 | 19 | 197 | 340 | |||
| Taxa | LD | 75 | 10 | 27 | 69 | ||
| HD | 52 | 10 | 23 | 45 | |||
| Total | 88 | 15 | 37 | 82 | |||
| Culture method | Taxa | LD | 10 | 2 | 3 | 6 | |
| HD | 8 | 1 | 3 | 5 | |||
| Total | 15 | 2 | 5 | 11 | |||
| Sample | Culture Method | Match Methods | Metabarcoding | |||
|---|---|---|---|---|---|---|
| Isolated Taxa | Taxa | OTUs | Reads | |||
| Shoot LD | 1 | Neoascochyta sp. | ✓ | 5 | 5 | 233 |
| 2 | Omnidemptus graminis | × | 5 | 6 | 1639 | |
| 3 | Omnidemptus graminis | × | 7 | 9 | 1619 | |
| 4 | Omnidemptus graminis | × | 3 | 3 | 207 | |
| 5 | × | 4 | 4 | 982 | ||
| Shoot HD | 6 | × | 2 | 2 | 229 | |
| 7 | Omnidemptus graminis | × | 2 | 2 | 37 | |
| 8 | Omnidemptus graminis | × | 5 | 5 | 644 | |
| 9 | Omnidemptus graminis | × | 1 | 1 | 13 | |
| 10 | × | 3 | 3 | 2281 | ||
| Rhizome LD | 1 | × | 8 | 102 | 12,546 | |
| 2 | × | 11 | 21 | 24,377 | ||
| 3 | Didymosphaeriaceae | × | 4 | 6 | 1312 | |
| Helotiaceae | ✓ | |||||
| 4 | × | 5 | 7 | 831 | ||
| 5 | Mollisiaceae | ✓ | 12 | 41 | 8202 | |
| Rhizome HD | 6 | Helotiaceae | ✓ | 6 | 22 | 5621 |
| 7 | Helotiaceae | ✓ | 3 | 4 | 267 | |
| 8 | Phialocephala sp. | × | 11 | 17 | 15,380 | |
| 9 | × | 4 | 5 | 2035 | ||
| 10 | Microdochium phragmitis | × | 11 | 18 | 6389 | |
| Root LD | 1 | Didymosphaeriaceae | × | 15 | 180 | 49,606 |
| 2 | Falciphora sp. | ✓ | 29 | 53 | 40,482 | |
| Codinaea sp. | × | |||||
| 3 | Didymosphaeriaceae | × | 27 | 184 | 70,132 | |
| 4 | Mollisia sp. | × | 34 | 73 | 52,335 | |
| 5 | Pleosporales | × | 31 | 95 | 49,118 | |
| Didymosphaeriaceae | × | |||||
| Lachnum sp. | ✓ | |||||
| Root HD | 6 | Helotiaceae | ✓ | 18 | 141 | 62,044 |
| 7 | Mollisiaceae | ✓ | 20 | 116 | 23,703 | |
| 8 | Albotricha sp. | ✓ | 16 | 32 | 12,379 | |
| 9 | Albotricha sp. | ✓ | 20 | 55 | 47,814 | |
| 10 | Drechslera sp. | ✓ | 17 | 59 | 21,214 | |
| Ilyonectria sp. | × | |||||
| ROOT | RHIZOME | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Endophyte Taxon | Relative Abundance (%) | Reads | OTUs | Infected Plants | Relative Abundance (%) | Reads | OTUs | Infected Plants | ||||||||
| LD | HD | LD | HD | LD | HD | LD | HD | LD | HD | LD | HD | LD | HD | LD | HD | |
| Helotiaceae | 24.51 | 31.09 | 64,132 | 51,968 | 114 | 116 | 2 | 4 | 25.95 | 16.49 | 12,267 | 4897 | 94 | 18 | 2 | 4 |
| Lachnum sp. A | 35.08 | 12.5 | 91,790 | 20,889 | 36 | 20 | 5 | 5 | 29.13 | 9.01 | 13,771 | 2676 | 5 | 7 | 4 | 4 |
| Albotricha sp. A | 1.71 | 18.83 | 4465 | 31,473 | 7 | 6 | 3 | 3 | 5.23 | 50.58 | 2473 | 15,018 | 4 | 6 | 3 | 3 |
| Helotiales A | 12.43 | 6.03 | 32,534 | 10,072 | 40 | 28 | 5 | 5 | 10.98 | 2.77 | 5188 | 823 | 25 | 1 | 2 | 3 |
| Agaricales A | 3.55 | 9.65 | 9281 | 16,124 | 3 | 3 | 2 | 4 | 0.05 | 0 | 24 | 0 | 2 | 0 | 1 | 0 |
| Mycena sp. A | 2.03 | 0.17 | 5323 | 289 | 10 | 2 | 4 | 1 | 20.88 | 0 | 9870 | 0 | 1 | 0 | 1 | 0 |
| Mollisiaceae C | 4.17 | 0.45 | 10,913 | 745 | 2 | 1 | 4 | 1 | 0.42 | 0.04 | 198 | 13 | 2 | 1 | 1 | 1 |
| Pleosporales A | 0.56 | 4.04 | 1476 | 6751 | 2 | 4 | 3 | 5 | 0 | 0.04 | 0 | 12 | 0 | 1 | 0 | 1 |
| Glarea sp. | 0.41 | 3.94 | 1060 | 6589 | 2 | 1 | 2 | 1 | 0 | 0.29 | 0 | 86 | 0 | 1 | 0 | 1 |
| Mollisiaceae B | 0.43 | 1.89 | 1118 | 3161 | 1 | 3 | 2 | 3 | 2.35 | 1.44 | 1111 | 429 | 1 | 3 | 2 | 2 |
| Mollisiaceae D | 0.89 | 1.07 | 2330 | 1782 | 1 | 2 | 2 | 1 | 0.78 | 3.56 | 369 | 1056 | 1 | 2 | 2 | 2 |
| Chaetosphaeriaceae | 1.76 | 0 | 4608 | 0 | 4 | 0 | 1 | 0 | 0.07 | 0 | 33 | 0 | 1 | 0 | 1 | 0 |
| Mycena sp. B | 0 | 2.08 | 0 | 3479 | 0 | 3 | 0 | 1 | 0 | 3.34 | 0 | 993 | 0 | 1 | 0 | 1 |
| Tricholomataceae B | 0 | 1.48 | 0 | 2474 | 0 | 1 | 0 | 2 | 0 | 4.21 | 0 | 1251 | 0 | 1 | 0 | 1 |
| Lachnum sp. B | 0.38 | 1.27 | 1007 | 2119 | 11 | 7 | 4 | 4 | 0.87 | 0.14 | 411 | 43 | 1 | 1 | 1 | 1 |
| Cantharellales | 1.3 | 0 | 3397 | 0 | 2 | 0 | 1 | 0 | ||||||||
| Parasola sp. | 0 | 0.9 | 0 | 1503 | 0 | 1 | 0 | 1 | 0 | 5.81 | 0 | 1725 | 0 | 3 | 0 | 1 |
| Unidentified A | 1.21 | 0.01 | 3174 | 19 | 2 | 1 | 1 | 1 | ||||||||
| Ophiosphaerella sp. | 0.96 | 0.32 | 2513 | 535 | 2 | 1 | 2 | 1 | 0.11 | 0.06 | 50 | 17 | 1 | 1 | 1 | 1 |
| Mollisiaceae A | 0.88 | 0.4 | 2309 | 666 | 4 | 3 | 4 | 5 | 0.03 | 0.28 | 13 | 83 | 1 | 1 | 1 | 1 |
| Drechslera sp. | 0.03 | 1.43 | 87 | 2388 | 2 | 2 | 2 | 5 | ||||||||
| SHOOT | ||||||||
|---|---|---|---|---|---|---|---|---|
| Endophyte Taxon | Relative Abundance (%) | Reads | OTUs | Infected Plants | ||||
| LD | HD | LD | HD | LD | HD | LD | HD | |
| Phaeosphaeriaceae | 34.08 | 58.80 | 1595 | 1884 | 2 | 1 | 4 | 2 |
| Phyllachorales | 20.49 | 0 | 959 | 0 | 1 | 0 | 4 | 0 |
| Puccinia sp. | 16.05 | 0 | 751 | 0 | 3 | 0 | 1 | 0 |
| Neoascochyta sp. A | 14.94 | 0.53 | 699 | 17 | 1 | 1 | 4 | 1 |
| Sordariomycetes A | 0 | 17.45 | 0 | 559 | 0 | 1 | 0 | 1 |
| Mycosphaerellaceae | 0 | 12.73 | 0 | 408 | 1 | 0 | 0 | 1 |
| Microdochium sp. | 5.58 | 1.59 | 261 | 51 | 1 | 1 | 4 | 1 |
| Dothideales | 3.65 | 0 | 171 | 0 | 1 | 0 | 2 | 0 |
| Ophiosphaerella sp. | 0 | 4.68 | 0 | 150 | 0 | 1 | 0 | 1 |
| Epicoccum sp. | 2.01 | 0.75 | 94 | 24 | 1 | 1 | 1 | 1 |
| Helotiaceae | 1.41 | 1.56 | 66 | 50 | 1 | 2 | 2 | 3 |
| Periconia sp. | 1.56 | 0 | 73 | 0 | 1 | 0 | 1 | 0 |
| Lachnum sp. A | 0 | 1.28 | 0 | 41 | 0 | 1 | 0 | 1 |
| Phragmocephala sp. B | 0 | 0.63 | 0 | 20 | 1 | 0 | 0 | 1 |
| Unidentified C | 0.23 | 0 | 11 | 0 | 1 | 0 | 1 | 0 |
| 100 | 100 | 4680 | 3204 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durán, M.; San Emeterio, L.; Canals, R.M. Comparison of Culturing and Metabarcoding Methods to Describe the Fungal Endophytic Assemblage of Brachypodium rupestre Growing in a Range of Anthropized Disturbance Regimes. Biology 2021, 10, 1246. https://doi.org/10.3390/biology10121246
Durán M, San Emeterio L, Canals RM. Comparison of Culturing and Metabarcoding Methods to Describe the Fungal Endophytic Assemblage of Brachypodium rupestre Growing in a Range of Anthropized Disturbance Regimes. Biology. 2021; 10(12):1246. https://doi.org/10.3390/biology10121246
Chicago/Turabian StyleDurán, María, Leticia San Emeterio, and Rosa Maria Canals. 2021. "Comparison of Culturing and Metabarcoding Methods to Describe the Fungal Endophytic Assemblage of Brachypodium rupestre Growing in a Range of Anthropized Disturbance Regimes" Biology 10, no. 12: 1246. https://doi.org/10.3390/biology10121246
APA StyleDurán, M., San Emeterio, L., & Canals, R. M. (2021). Comparison of Culturing and Metabarcoding Methods to Describe the Fungal Endophytic Assemblage of Brachypodium rupestre Growing in a Range of Anthropized Disturbance Regimes. Biology, 10(12), 1246. https://doi.org/10.3390/biology10121246