
The Functions of β-Defensin in Flounder (Paralichthys olivaceus): Antibiosis, Chemotaxis and Modulation of Phagocytosis


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. RNA Extraction and cDNA Synthesis
2.3. Cloning and Sequence Analysis
2.4. Preparation of Recombinant fBD
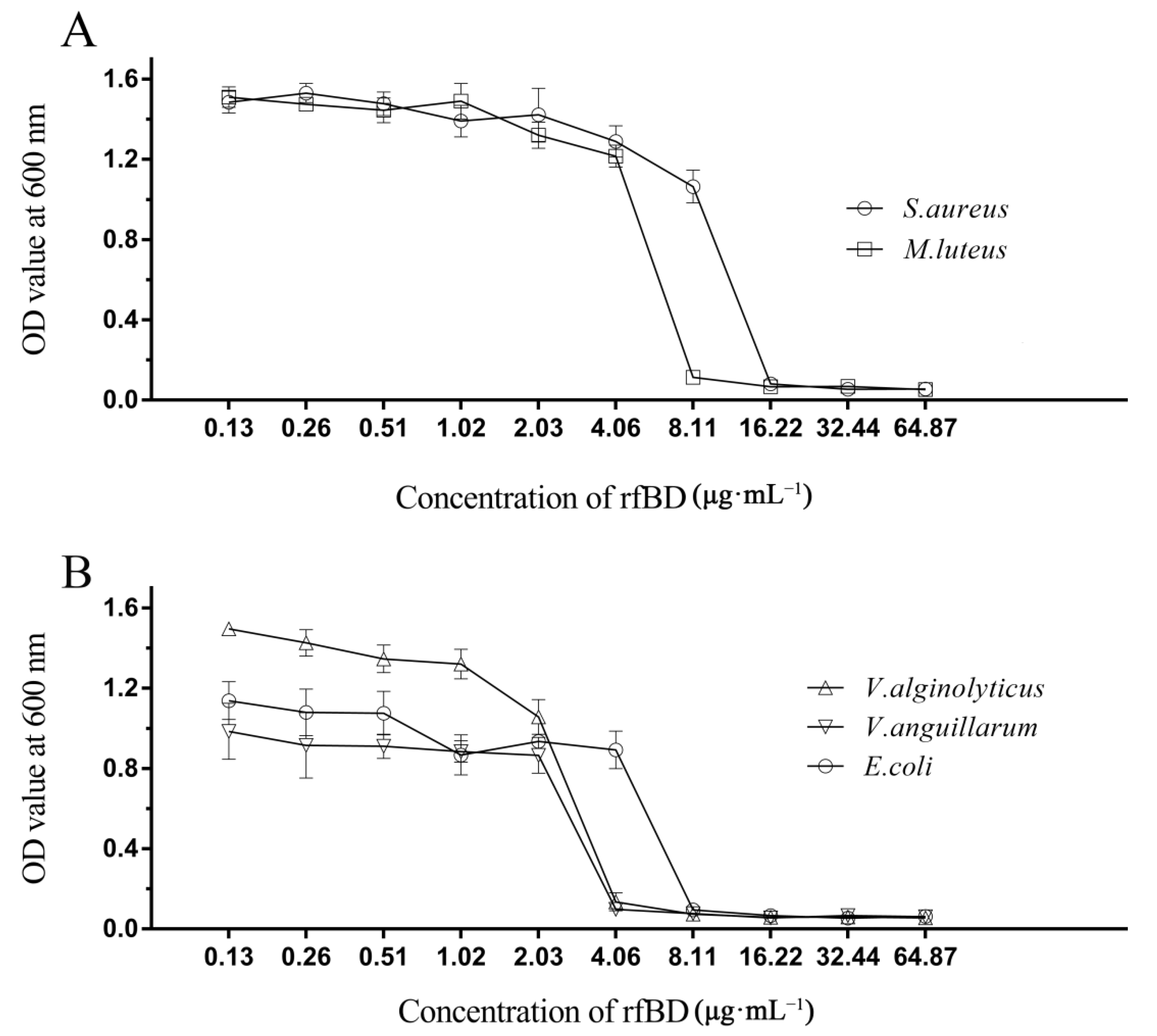
2.5. Antimicrobial Activity Assay
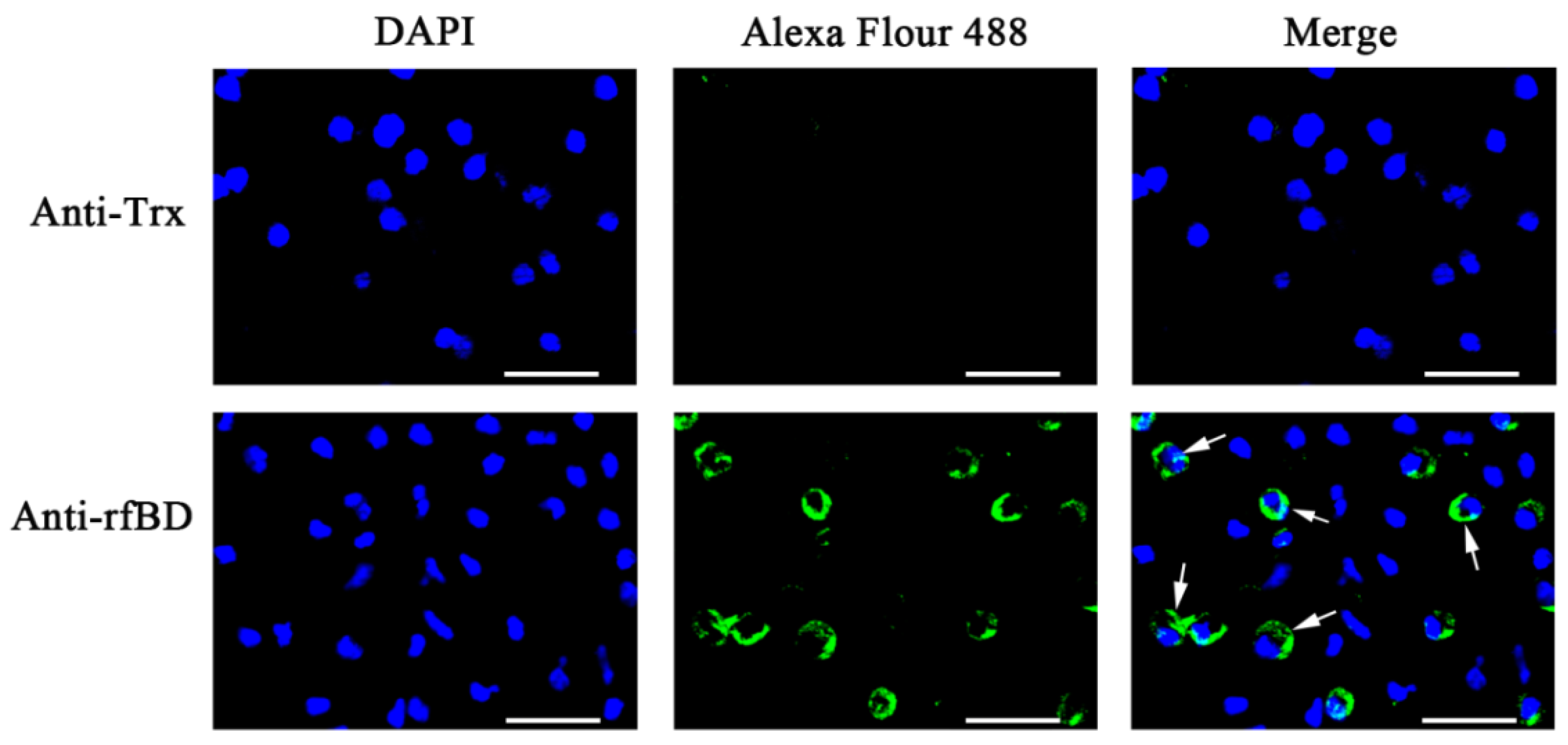
2.6. Production of Antibody and Indirect Immunofluorescence Assay
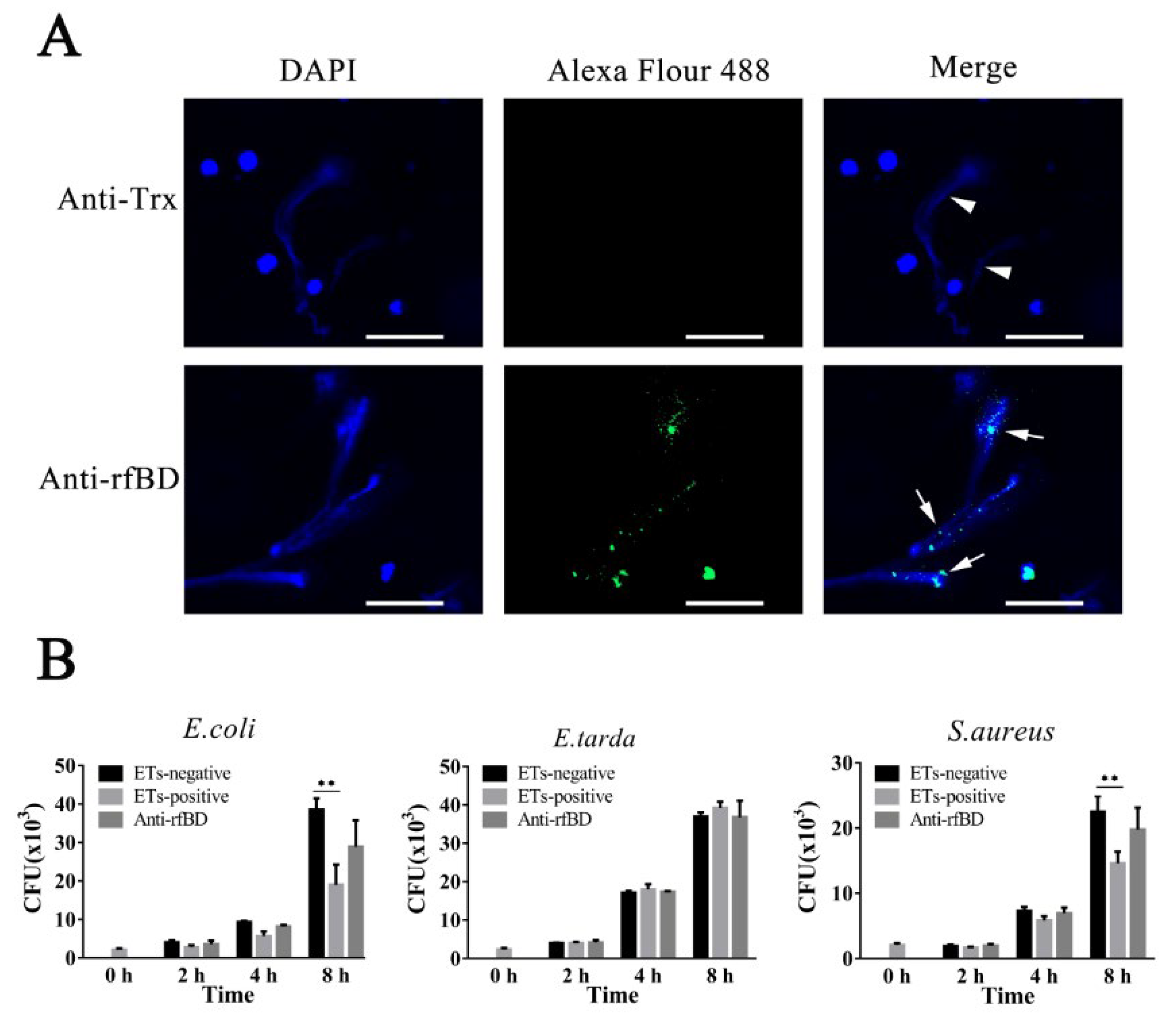
2.7. Co-localization rfBD and Extracellular Traps
2.8. Survival of ETs-Trapped Bacteria
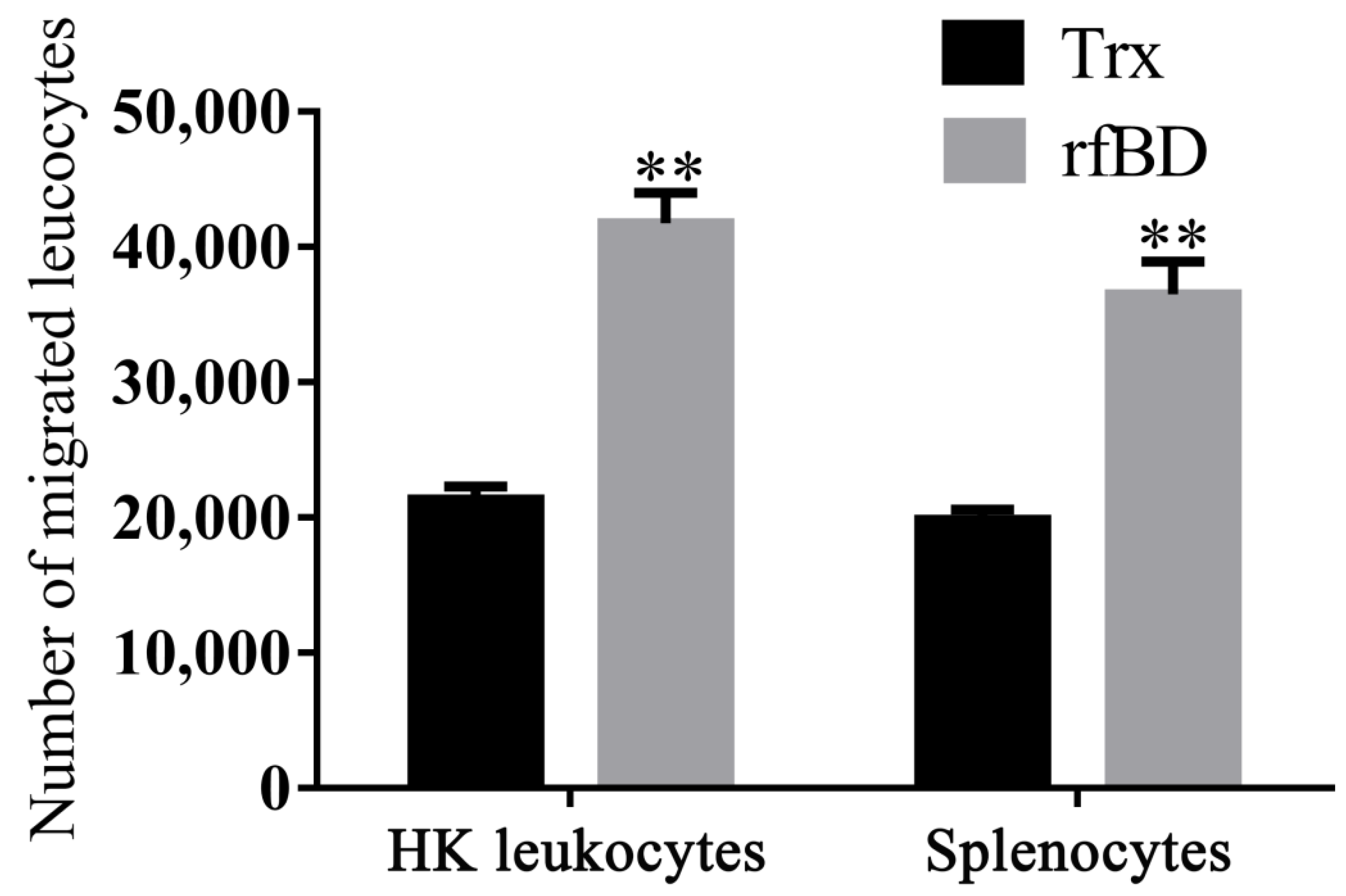
2.9. In Vitro Chemotaxis Assay
2.10. Phagocytic Assay
2.11. Statistical Analysis
3. Result
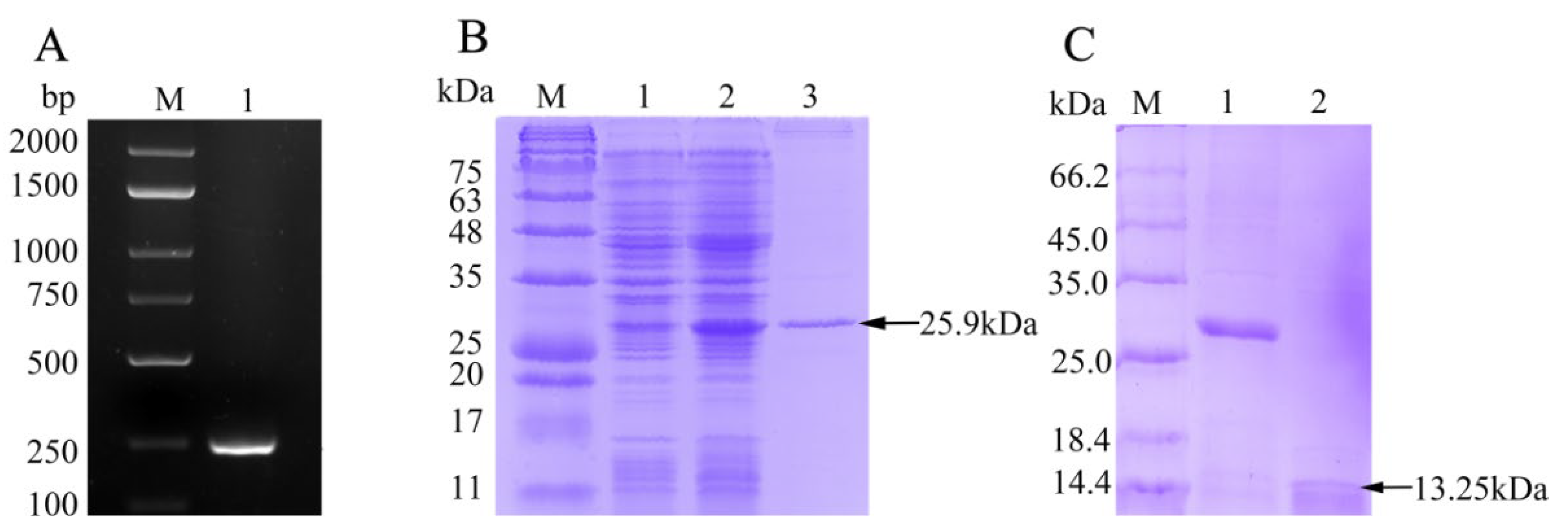
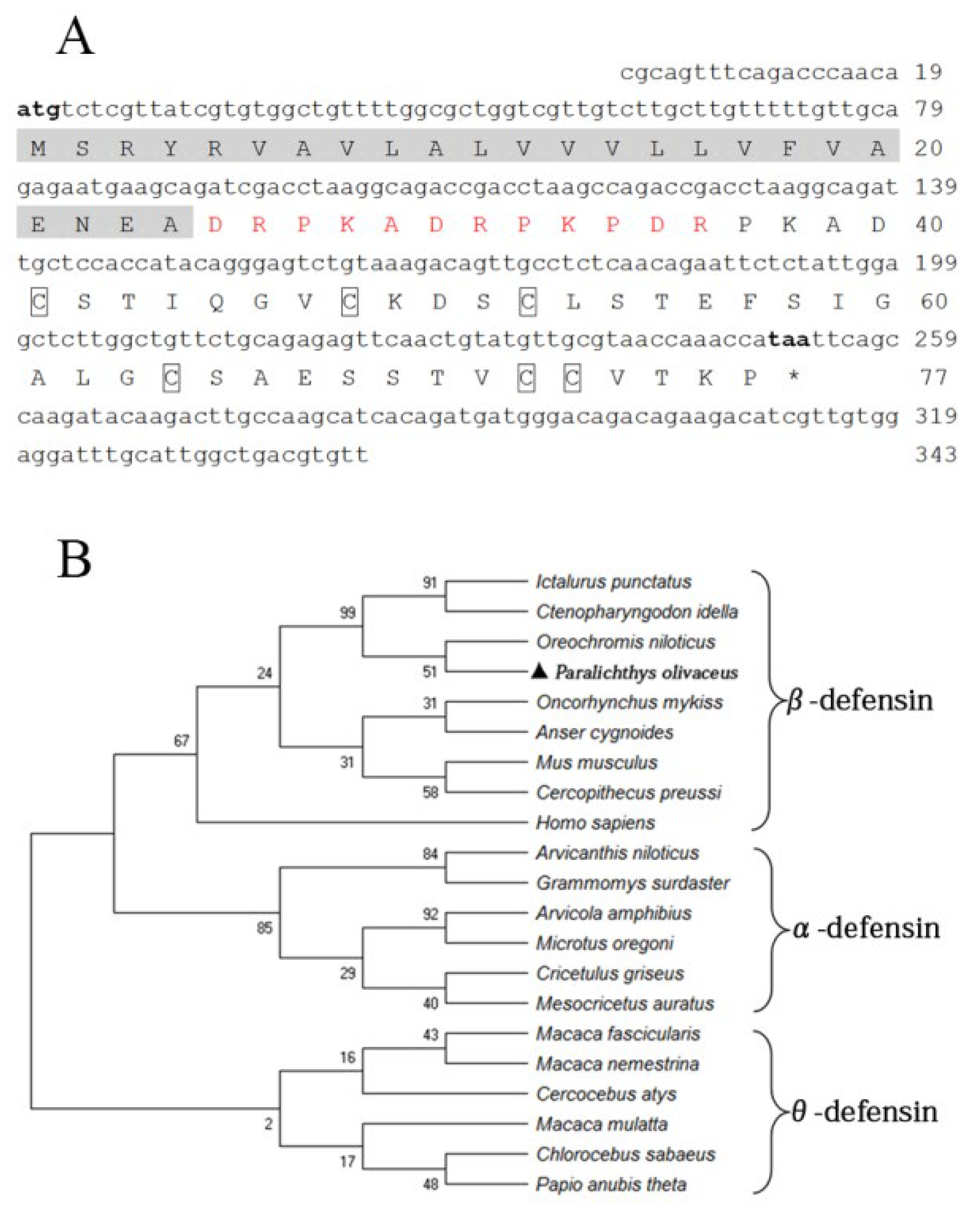
3.1. Cloning and Sequence Analysis of fBD
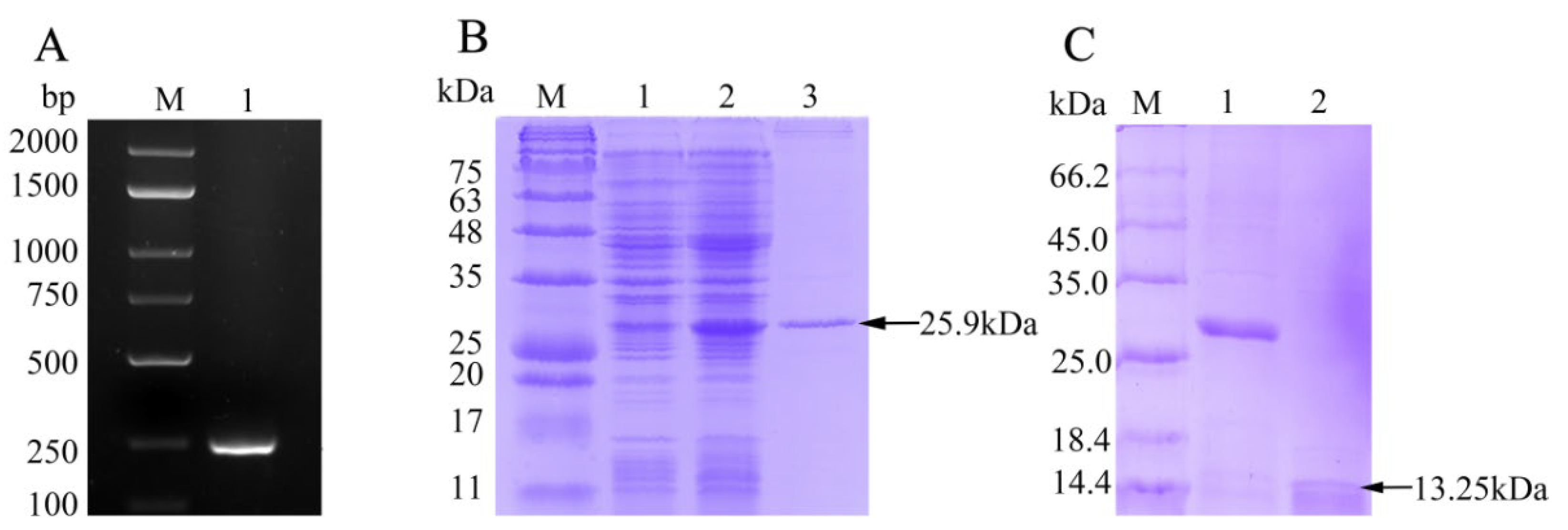
3.2. rfBD Production and Antibody Verification
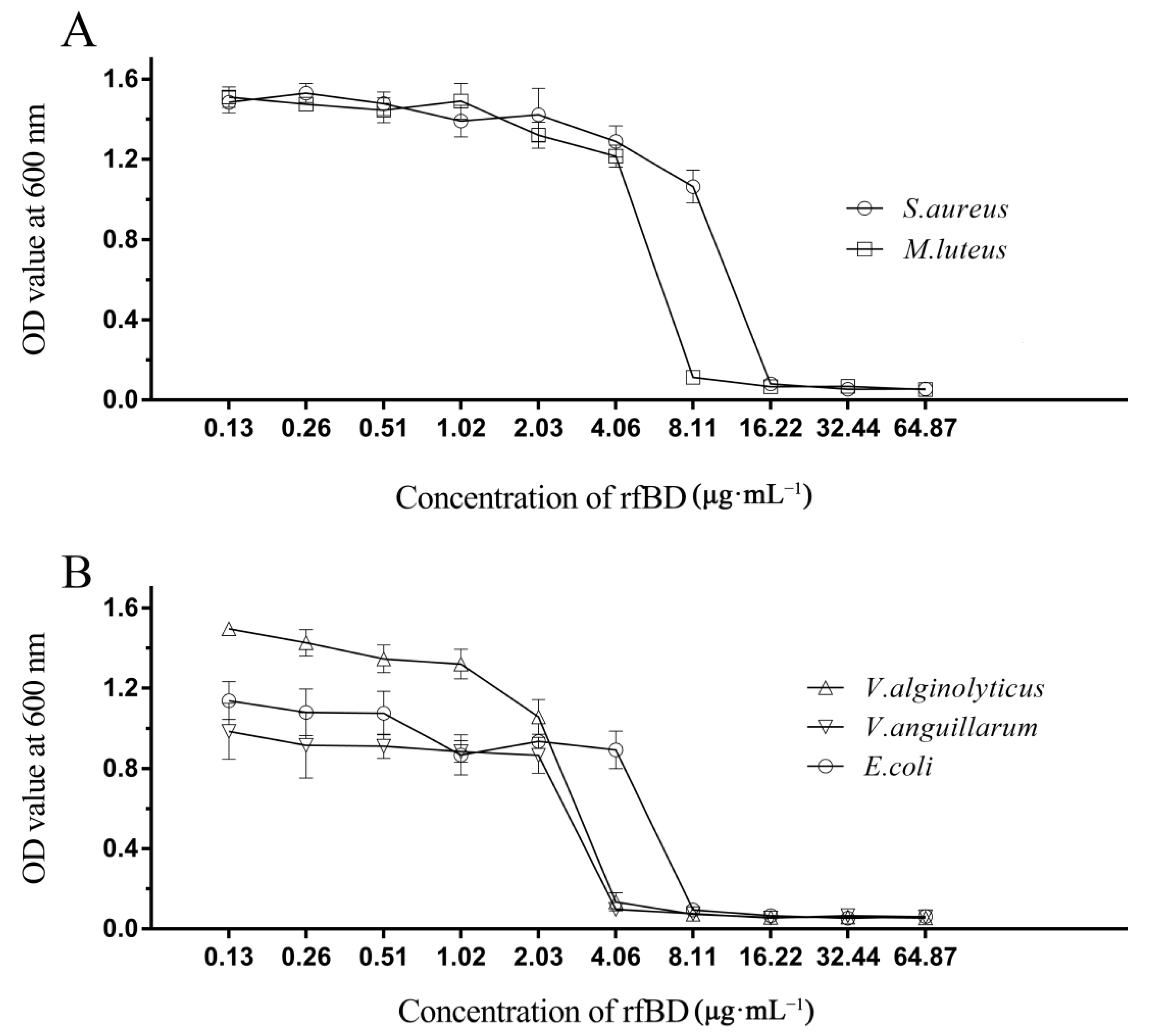
3.3. Antimicrobial Activity of rfBD
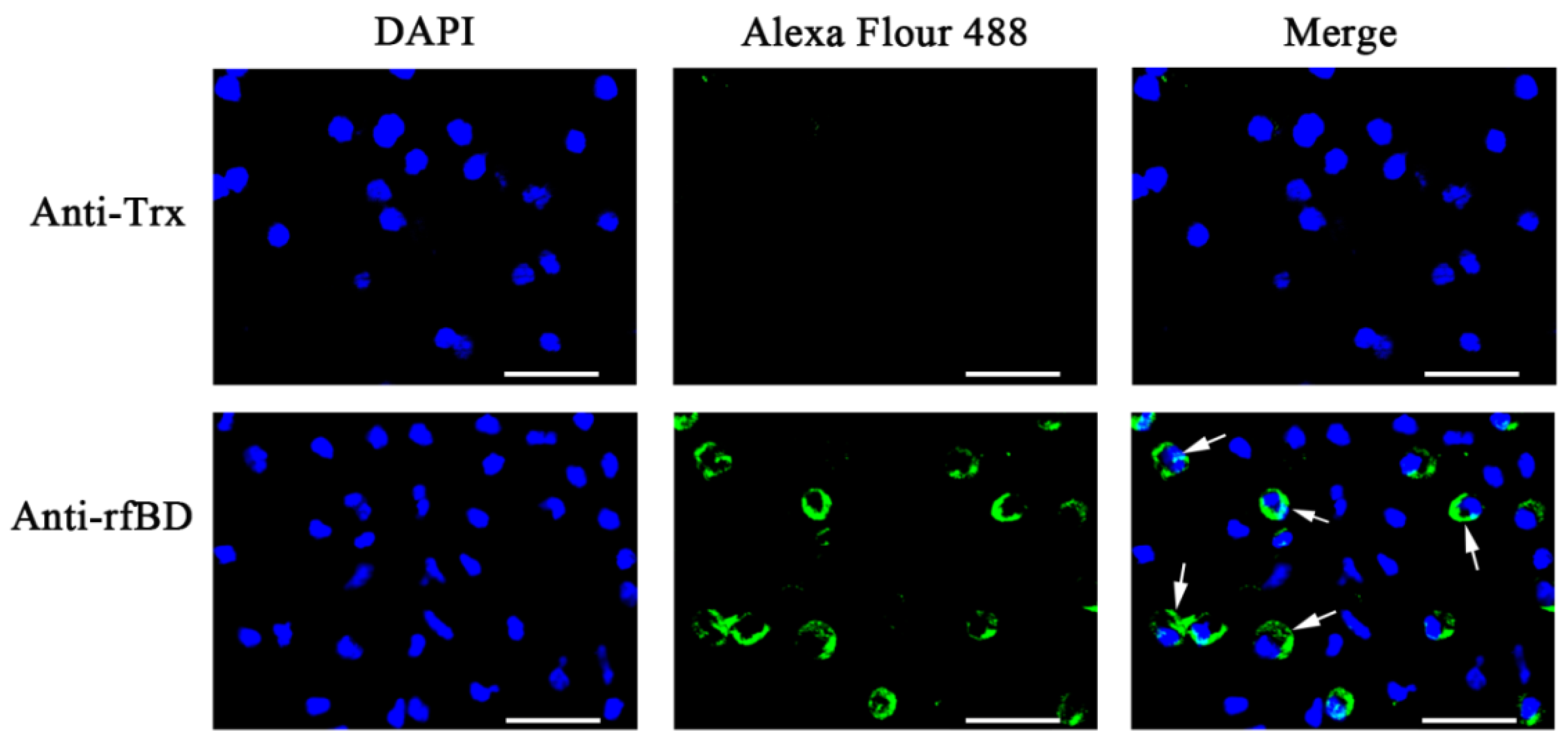
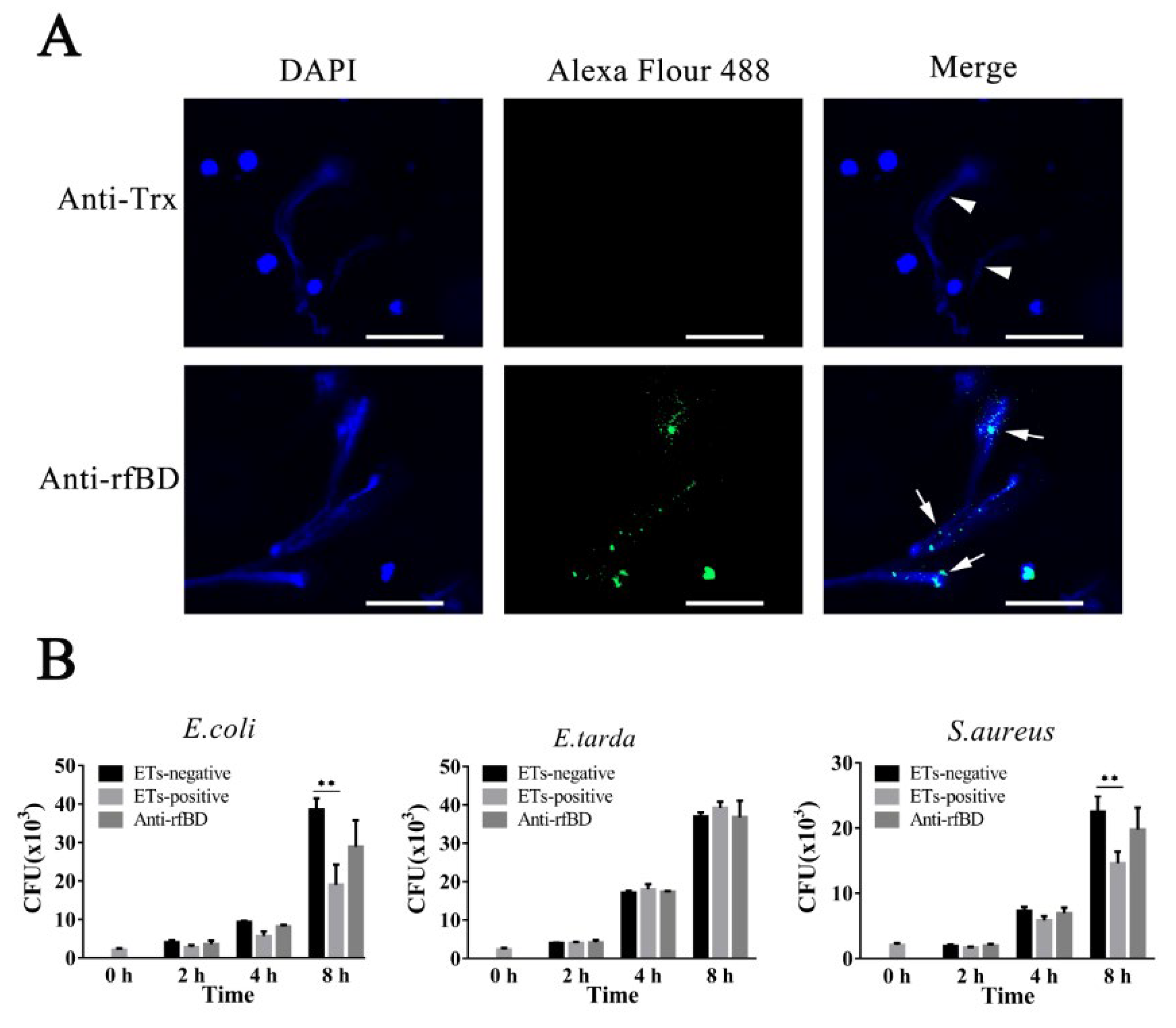
3.4. Localization and Effect of fBD on the ETs
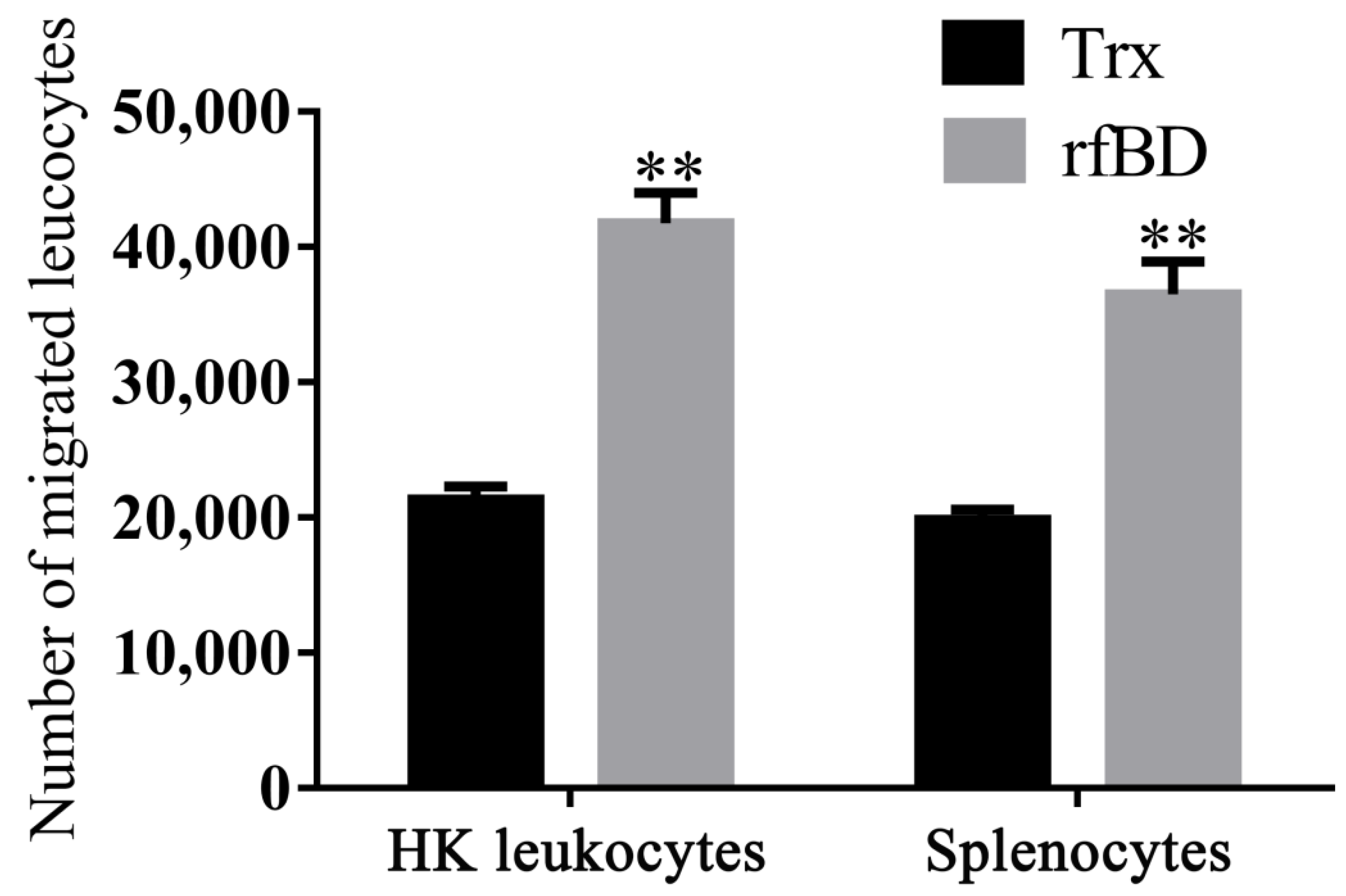
3.5. Effects of rfBD on Chemotaxis of Leukocytes
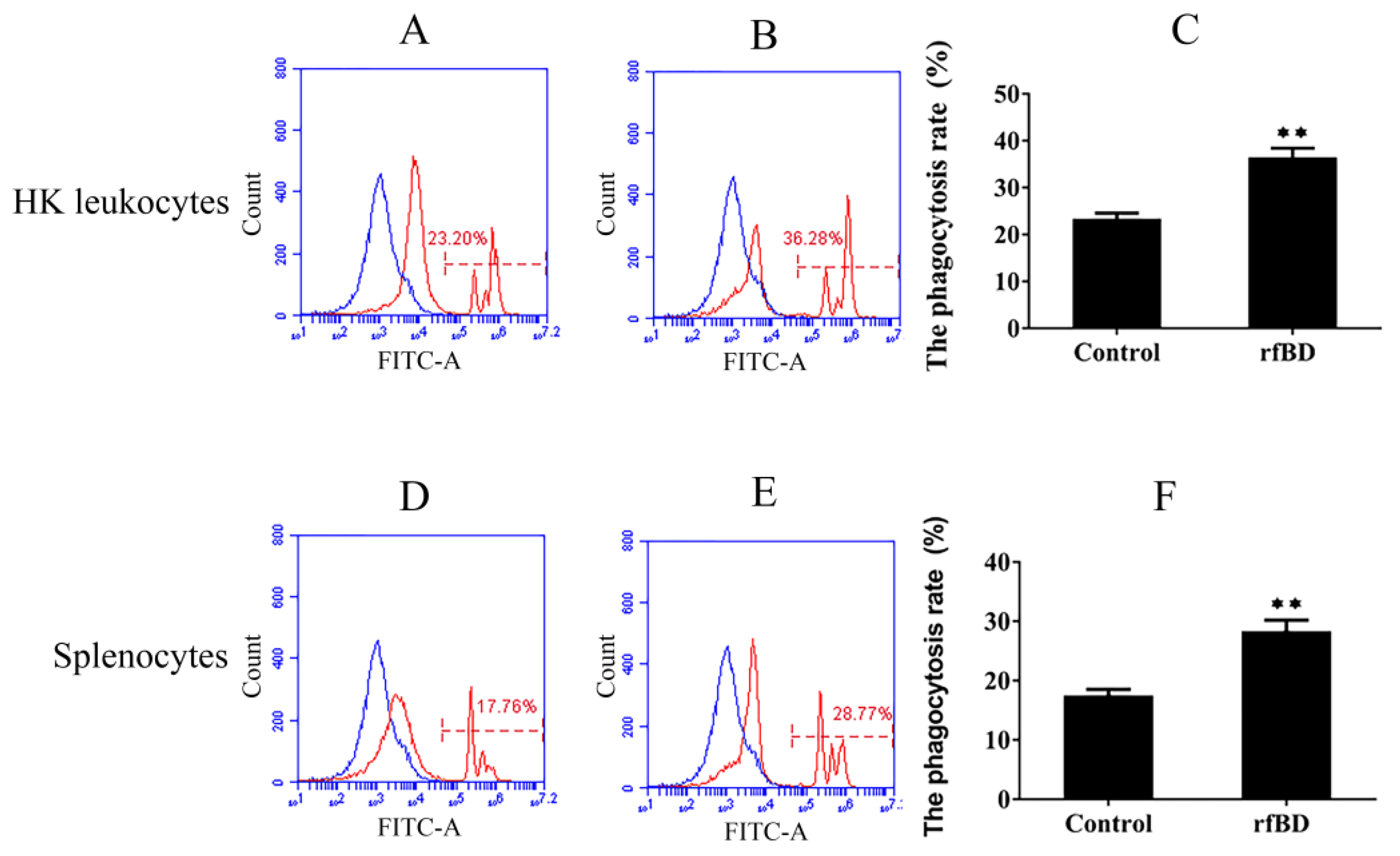
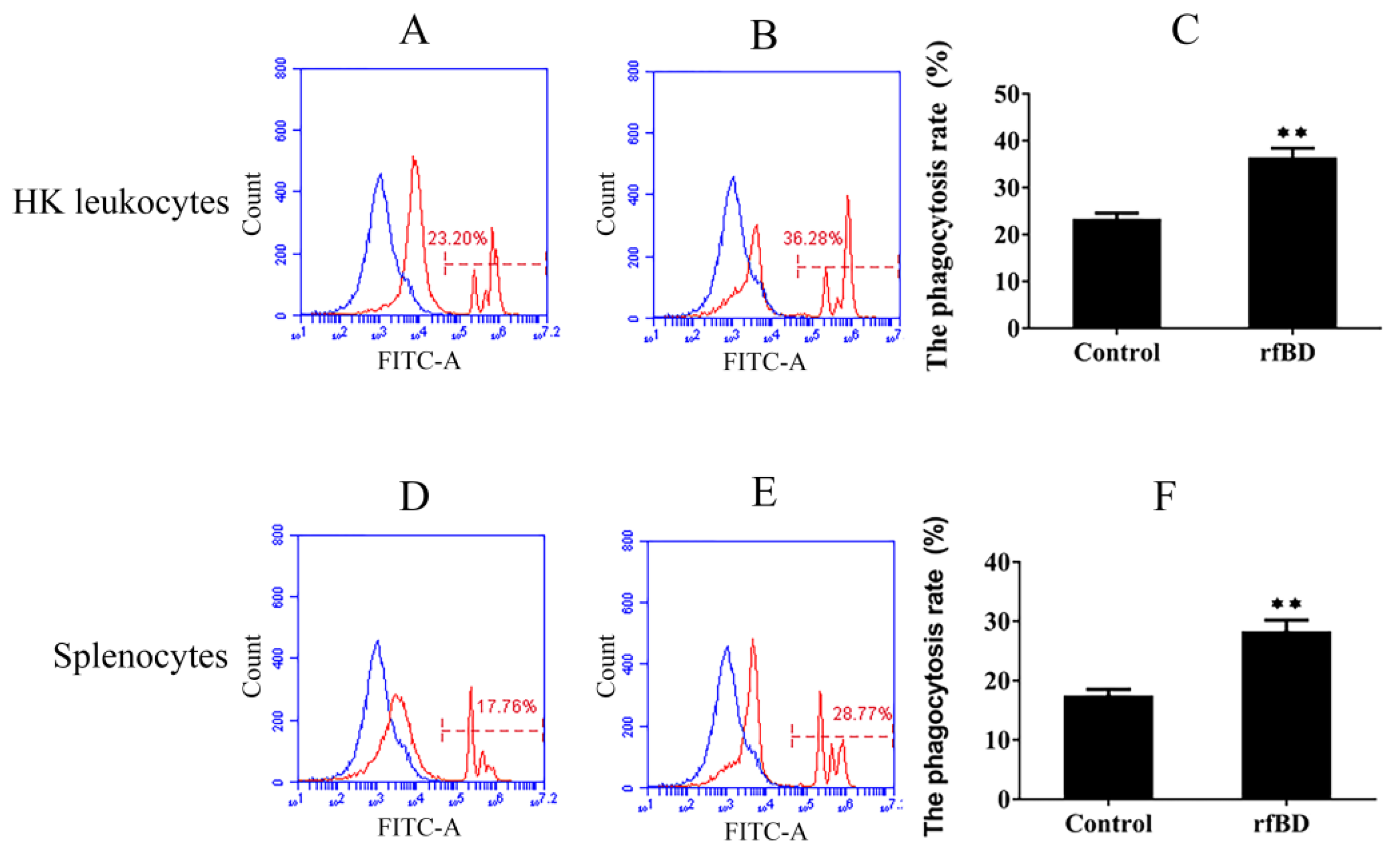
3.6. Effects of rfBD on the Phagocytosis of Lymphocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ellis, A.E. Innate host defense mechanisms of fish against viruses and bacteria. Dev. Comp. Immunol. 2001, 25, 827–839. [Google Scholar] [CrossRef]
- Oliva, R.; Chino, M.; Pane, K.; Pistorio, V.; De Santis, A.; Pizzo, E.; D’Errico, G.; Pavone, V.; Lombardi, A.; Del Vecchio, P.; et al. Exploring the role of unnatural amino acids in antimicrobial peptides. Sci. Rep. 2018, 8, 8888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valero, Y.; Chaves-Pozo, E.; Cuesta, A. NK-lysin is highly conserved in European sea bass and gilthead seabream but differentially modulated during the immune response. Fish. Shellfish Immunol. 2020, 99, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cole, A.M.; Hong, T.; Waring, A.J.; Lehrer, R.I. Retrocyclin, an antiretroviral theta-defensin, is a lectin. J. Immunol. 2003, 170, 4708–4716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klotman, M.E.; Chang, T.L. Defensins in innate antiviral immunity. Nat. Rev. Immunol. 2006, 6, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.Q.; Yuan, J.; Osapay, G.; Osapay, K.; Tran, D.; Miller, C.J.; Ouellette, A.J.; Selsted, M.E. A cyclic antimicrobial peptide produced in primate leukocytes by the ligation of two truncated alpha-defensins. Science 1999, 286, 498–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, R.; Mokhtar, S.S.; Phipps, M.E.; Hoh, B.P.; Xu, S. A genome-wide characterization of copy number variations in native populations of Peninsular Malaysia. Eur. J. Hum. Genet. EJHG 2018, 26, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.I.; Pleguezuelos, O.; Zhang, Y.A.; Zou, J.; Secombes, C.J. Identification of a novel cathelicidin gene in the rainbow trout, Oncorhynchus mykiss. Infect. Immun. 2005, 73, 5053–5064. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.L.; Xu, M.Y.; Ji, X.S.; Yu, G.C.; Liu, Y. Cloning, characterization, and expression analysis of hepcidin gene from red sea bream (Chrysophrys major). Antimicrob. Agents Chemother. 2005, 49, 1608–1612. [Google Scholar] [CrossRef] [Green Version]
- Cole, A.M.; Weis, P.; Diamond, G. Isolation and characterization of pleurocidin, an antimicrobial peptide in the skin secretions of winter flounder. J. Biol. Chem. 1997, 272, 12008–12013. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.Y.; Ohira, T.; Hirono, I.; Aoki, T. A pore-forming protein, perforin, from a non-mammalian organism, Japanese flounder, Paralichthys olivaceus. Immunogenetics 2004, 56, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Lauth, X.; Shike, H.; Burns, J.C.; Westerman, M.E.; Ostland, V.E.; Carlberg, J.M.; Van Olst, J.C.; Nizet, V.; Taylor, S.W.; Shimizu, C.; et al. Discovery and Characterization of Two Isoforms of Moronecidin, a Novel Antimicrobial Peptide from Hybrid Striped Bass. J. Biol. Chem. 2002, 277, 5030–5039. [Google Scholar] [CrossRef] [Green Version]
- Richards, R.C.; O’Neil, D.B.; Thibault, P.; Ewart, K.V. Histone H1: An antimicrobial protein of Atlantic salmon (Salmo salar). Biochem. Biophys. Res. Commun. 2001, 284, 549–555. [Google Scholar] [CrossRef]
- Zou, J.; Mercier, C.; Koussounadis, A.; Secombes, C. Discovery of multiple beta-defensin like homologues in teleost fish. Mol. Immunol. 2007, 44, 638–647. [Google Scholar] [CrossRef]
- Ganz, T.; Selsted, M.E.; Szklarek, D.; Harwig, S.S.; Daher, K.; Bainton, D.F.; Lehrer, R.I. Defensins. Natural peptide antibiotics of human neutrophils. J. Clin. Investig. 1985, 76, 1427–1435. [Google Scholar] [CrossRef]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.; Barran, P.E.; Dorin, J.R. Structure-activity relationships in beta-defensin peptides. Biopolymers 2008, 90, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Hou, B.; Ren, F.; Zhou, H.; Zhao, T. Characterization of grass carp (Ctenopharyngodon idella) beta-defensin 1: Implications for its role in inflammation control. Biosci. Biotechnol. Biochem. 2019, 83, 87–94. [Google Scholar] [CrossRef]
- Jiang, H.; Hu, Y.; Wei, X.; Xiao, X.; Jakovlic, I.; Liu, X.; Su, J.; Yuan, G. Chemotactic effect of beta-defensin 1 on macrophages in Megalobrama amblycephala. Fish. Shellfish Immunol. 2018, 74, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, A.; Meseguer, J.; Esteban, M.A. Molecular and functional characterization of the gilthead seabream beta-defensin demonstrate its chemotactic and antimicrobial activity. Mol. Immunol. 2011, 48, 1432–1438. [Google Scholar] [CrossRef]
- Garcia-Valtanen, P.; Martinez-Lopez, A.; Ortega-Villaizan, M.; Perez, L.; Coll, J.M.; Estepa, A. In addition to its antiviral and immunomodulatory properties, the zebrafish beta-defensin 2 (zfBD2) is a potent viral DNA vaccine molecular adjuvant. Antiviral. Res. 2014, 101, 136–147. [Google Scholar] [CrossRef]
- Daniel, C.; Leppkes, M.; Munoz, L.E.; Schley, G.; Schett, G.; Herrmann, M. Extracellular DNA traps in inflammation, injury and healing. Nat. Rev. Nephrol. 2019, 15, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Borregaard, N.; Cowland, J.B. Granules of the Human Neutrophilic Polymorphonuclear Leukocyte. Blood 1997, 89, 3503–3521. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhan, W.; Xing, J.; Sheng, X. Production, characterisation and applicability of monoclonal antibodies to immunoglobulin of Japanese flounder (Paralichthys olivaceus). Fish. Shellfish Immunol. 2007, 23, 982–990. [Google Scholar] [CrossRef]
- Nam, B.H.; Moon, J.Y.; Kim, Y.O.; Kong, H.J.; Kim, W.J.; Lee, S.J.; Kim, K.K. Multiple beta-defensin isoforms identified in early developmental stages of the teleost Paralichthys olivaceus. Fish. Shellfish Immunol. 2010, 28, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Tang, X.; Zhan, W.; Xing, J.; Sheng, X. Immunoglobulin Tau Heavy Chain (IgT) in Flounder, Paralichthys olivaceus: Molecular Cloning, Characterization, and Expression Analyses. Int. J. Mol. Sci. 2016, 17, 1571. [Google Scholar] [CrossRef] [Green Version]
- Lv, C.; Han, Y.; Yang, D.; Zhao, J.; Wang, C.; Mu, C. Antibacterial activities and mechanisms of action of a defensin from manila clam Ruditapes philippinarum. Fish. Shellfish Immunol. 2020, 103, 266–276. [Google Scholar] [CrossRef]
- Tang, X.; Zhan, W.; Sheng, X.; Chi, H. Immune response of Japanese flounder Paralichthys olivaceus to outer membrane protein of Edwardsiella tarda. Fish. Shellfish Immunol. 2010, 28, 333–343. [Google Scholar] [CrossRef]
- Tang, X.; Zhou, L.; Zhan, W. Isolation and characterization of pathogenic Listonella anguillarum of diseased half-smooth tongue sole (Cynoglossus semilaevis Günther). J. Ocean. Univ. China 2008, 7, 343–351. [Google Scholar] [CrossRef]
- Xing, J.; Tian, H.; Wang, L.; Tang, X.; Sheng, X.; Zhan, W. Characterization of CD40(+) leukocytes in flounder (Paralichthys olivaceus) and its response after Hirame novirhabdovirus infection and immunization. Mol. Immunol. 2018, 104, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.L.; Zhao, M.L.; Chi, H.; Sun, L. Histones and chymotrypsin-like elastases play significant roles in the antimicrobial activity of tongue sole neutrophil extracellular traps. Fish. Shellfish Immunol. 2018, 72, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Ruangsri, J.; Kitani, Y.; Kiron, V.; Lokesh, J.; Brinchmann, M.F.; Karlsen, B.O.; Fernandes, J.M. A novel beta-defensin antimicrobial peptide in Atlantic cod with stimulatory effect on phagocytic activity. PLoS ONE 2013, 8, e62302. [Google Scholar]
- Wen, H.; Lan, X.; Cheng, T.; He, N.; Shiomi, K.; Kajiura, Z.; Zhou, Z.; Xia, Q.; Xiang, Z.; Nakagaki, M. Sequence structure and expression pattern of a novel anionic defensin-like gene from silkworm (Bombyx mori). Mol. Biol. Rep. 2009, 36, 711–716. [Google Scholar] [CrossRef]
- Lai, R.; Lomas, L.O.; Jonczy, J.; Turner, P.C.; Rees, H.H. Two novel non-cationic defensin-like antimicrobial peptides from haemolymph of the female tick, Amblyomma hebraeum. Biochem. J. 2004, 379 Pt 3, 681–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.Y.; Zhou, L.; Wang, Y.; Li, Z.; Zhao, J.G.; Zhang, Q.Y.; Gui, J.F. Antibacterial and antiviral roles of a fish beta-defensin expressed both in pituitary and testis. PLoS ONE 2010, 5, e12883. [Google Scholar] [CrossRef]
- Zhao, J.G.; Zhou, L.; Jin, J.Y.; Zhao, Z.; Lan, J.; Zhang, Y.B.; Zhang, Q.Y.; Gui, J.F. Antimicrobial activity-specific to Gram-negative bacteria and immune modulation-mediated NF-kappaB and Sp1 of a medaka beta-defensin. Dev. Comp. Immunol. 2009, 33, 624–637. [Google Scholar] [CrossRef]
- Hermosilla, C.; Caro, T.M.; Silva, L.M.; Ruiz, A.; Taubert, A. The intriguing host innate immune response: Novel anti-parasitic defence by neutrophil extracellular traps. Parasitology 2014, 141, 1489–1498. [Google Scholar] [CrossRef]
- Lominadze, G.; Powell, D.W.; Luerman, G.C.; Link, A.J.; Ward, R.A.; McLeish, K.R. Proteomic analysis of human neutrophil granules. Mol. Cell. Proteom. 2005, 4, 1503–1521. [Google Scholar] [CrossRef] [Green Version]
- Durr, M.; Peschel, A. Chemokines meet defensins: The merging concepts of chemoattractants and antimicrobial peptides in host defense. Infect. Immun. 2002, 70, 6515–6517. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Biragyn, A.; Kwak, L.W.; Oppenheim, J.J. Mammalian defensins in immunity: More than just microbicidal. Trends Immunol. 2002, 23, 291–296. [Google Scholar] [CrossRef]
- Yang, D.; Chen, Q.; Chertov, O.; Oppenheim, J.J. Human neutrophil defensins selectively chemoattract naive T and immature dendritic cells. J. Leukoc. Biol. 2000, 68, 9–14. [Google Scholar] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′→3′) |
|---|---|
| fBD-OF | CGCAGTTTCAGACCCAACAATG |
| fBD-OR | AAGACGTCAGCCAATGCAAATC |
| fBD-IF | CGGGATCCATGTCTCGTTATCGTGTGGCT |
| fBD-IR | CCCTCGAGTTATGGTTTGGTTACGCAACATA |
| T7F | TAATACGACTCACTATAGGG |
| T7R | TGCTAGTTATTGCTCAGCGG |
| Microorganisms | MIC Value * (μg·mL−1) |
|---|---|
| Gram-positive bacteria | |
| S. aureus | 8.11–16.22 |
| M. luteus | 4.06–8.11 |
| Gram-negative bacteria | |
| E. coli | 4.06–8.11 |
| V. alginolyticus | 2.03–4.06 |
| V. anguillarum | 2.03–4.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, X.; Chi, H.; Tang, X.; Xing, J.; Sheng, X.; Zhan, W. The Functions of β-Defensin in Flounder (Paralichthys olivaceus): Antibiosis, Chemotaxis and Modulation of Phagocytosis. Biology 2021, 10, 1247. https://doi.org/10.3390/biology10121247
Hao X, Chi H, Tang X, Xing J, Sheng X, Zhan W. The Functions of β-Defensin in Flounder (Paralichthys olivaceus): Antibiosis, Chemotaxis and Modulation of Phagocytosis. Biology. 2021; 10(12):1247. https://doi.org/10.3390/biology10121247
Chicago/Turabian StyleHao, Xiaokai, Heng Chi, Xiaoqian Tang, Jing Xing, Xiuzhen Sheng, and Wenbin Zhan. 2021. "The Functions of β-Defensin in Flounder (Paralichthys olivaceus): Antibiosis, Chemotaxis and Modulation of Phagocytosis" Biology 10, no. 12: 1247. https://doi.org/10.3390/biology10121247
APA StyleHao, X., Chi, H., Tang, X., Xing, J., Sheng, X., & Zhan, W. (2021). The Functions of β-Defensin in Flounder (Paralichthys olivaceus): Antibiosis, Chemotaxis and Modulation of Phagocytosis. Biology, 10(12), 1247. https://doi.org/10.3390/biology10121247