
Quantitative Analysis of Factors Regulating Angiogenesis for Stem Cell Therapy

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
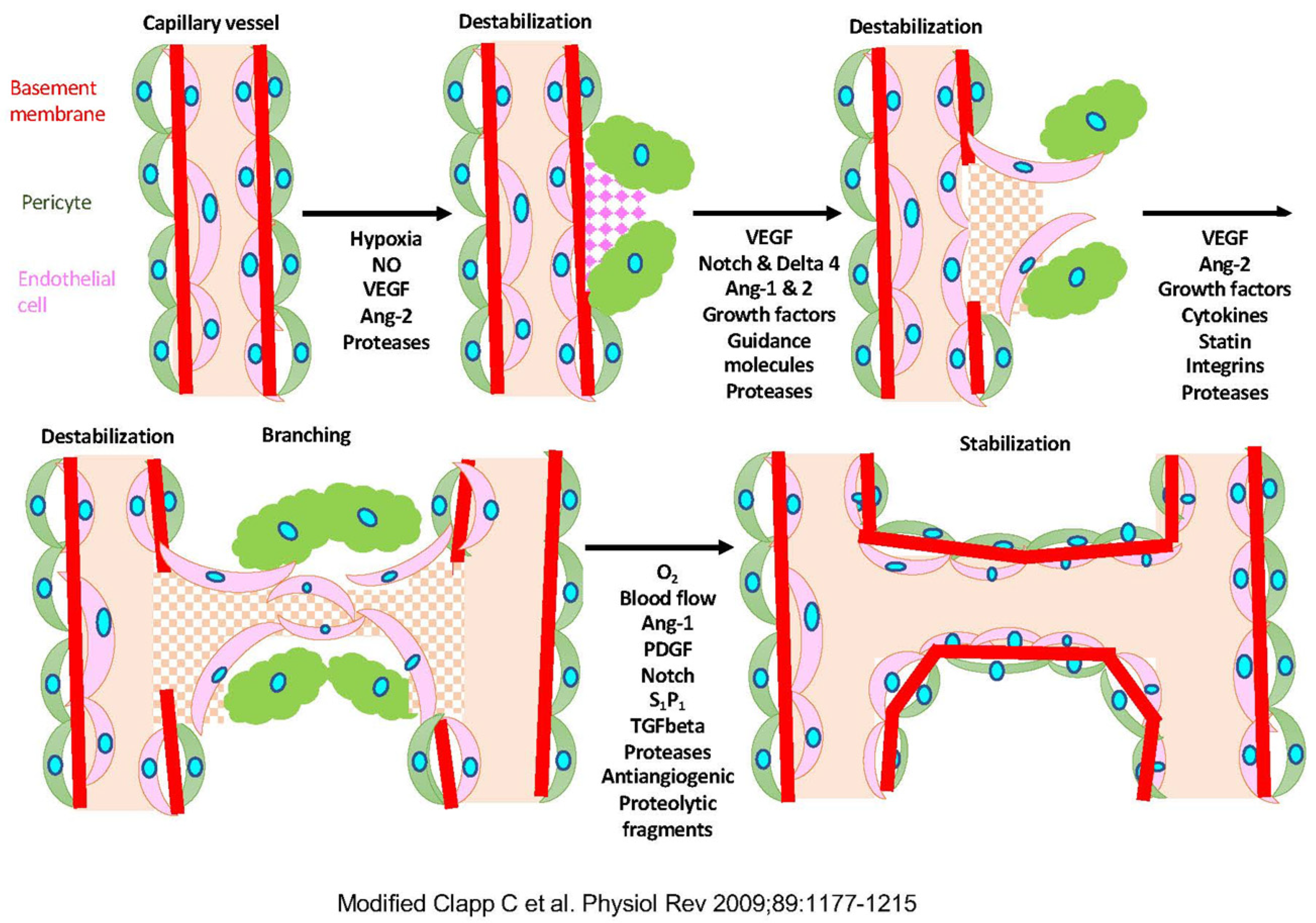
1.1. Angiogenesis
1.2. Disease and Angiogenesis
1.3. Regenerative Medicine and Angiogenesis
1.4. Factors Involved in Angioplasty
1.5. Polyphenols
1.6. Molecular Pathway of Angiogenesis
1.7. The Aim of This Research
2. Materials and Methods
2.1. Materials
2.2. Angiogenesis Assay
2.3. Quantitative Analysis
2.4. Gene Expression Analysis
3. Results
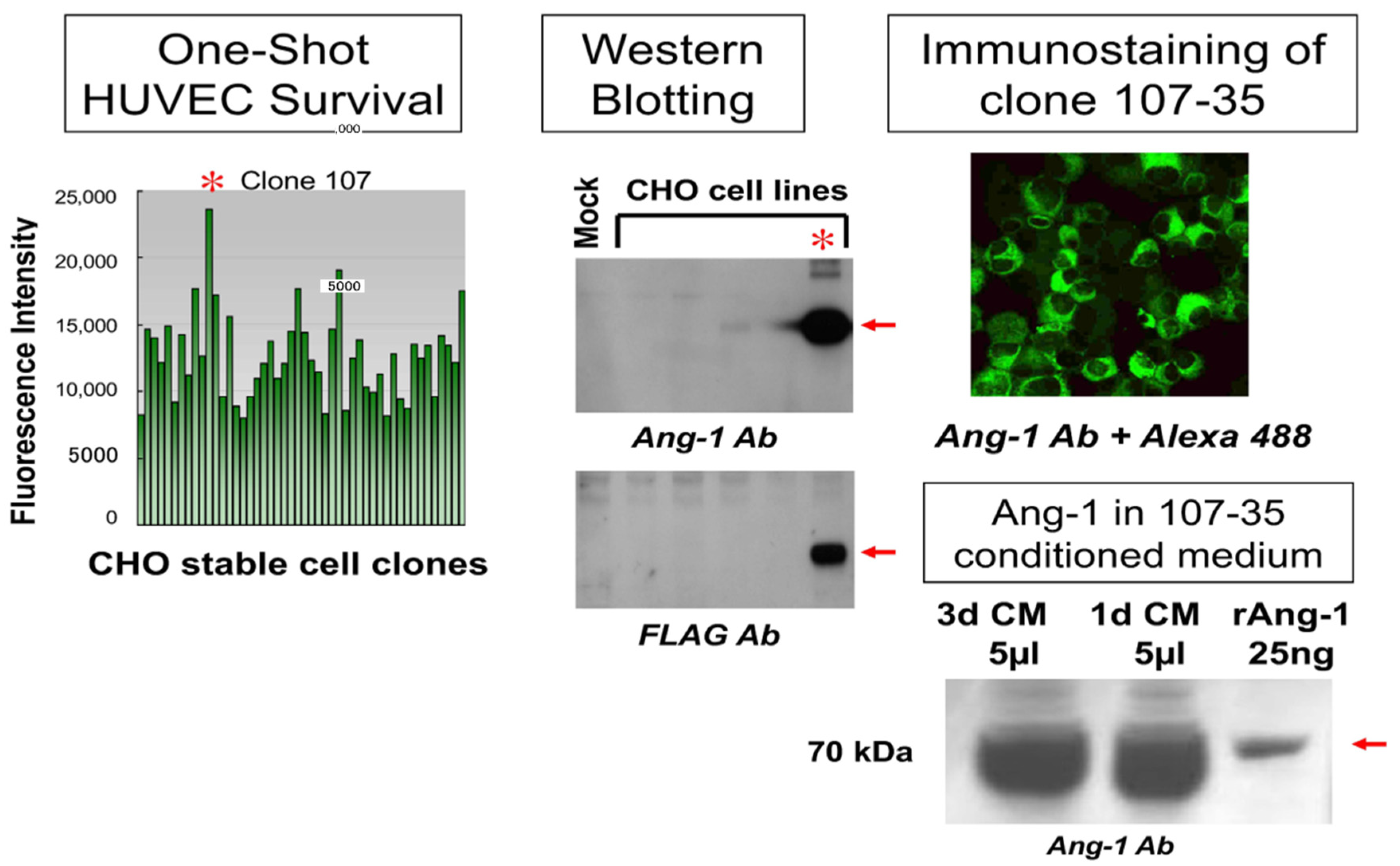
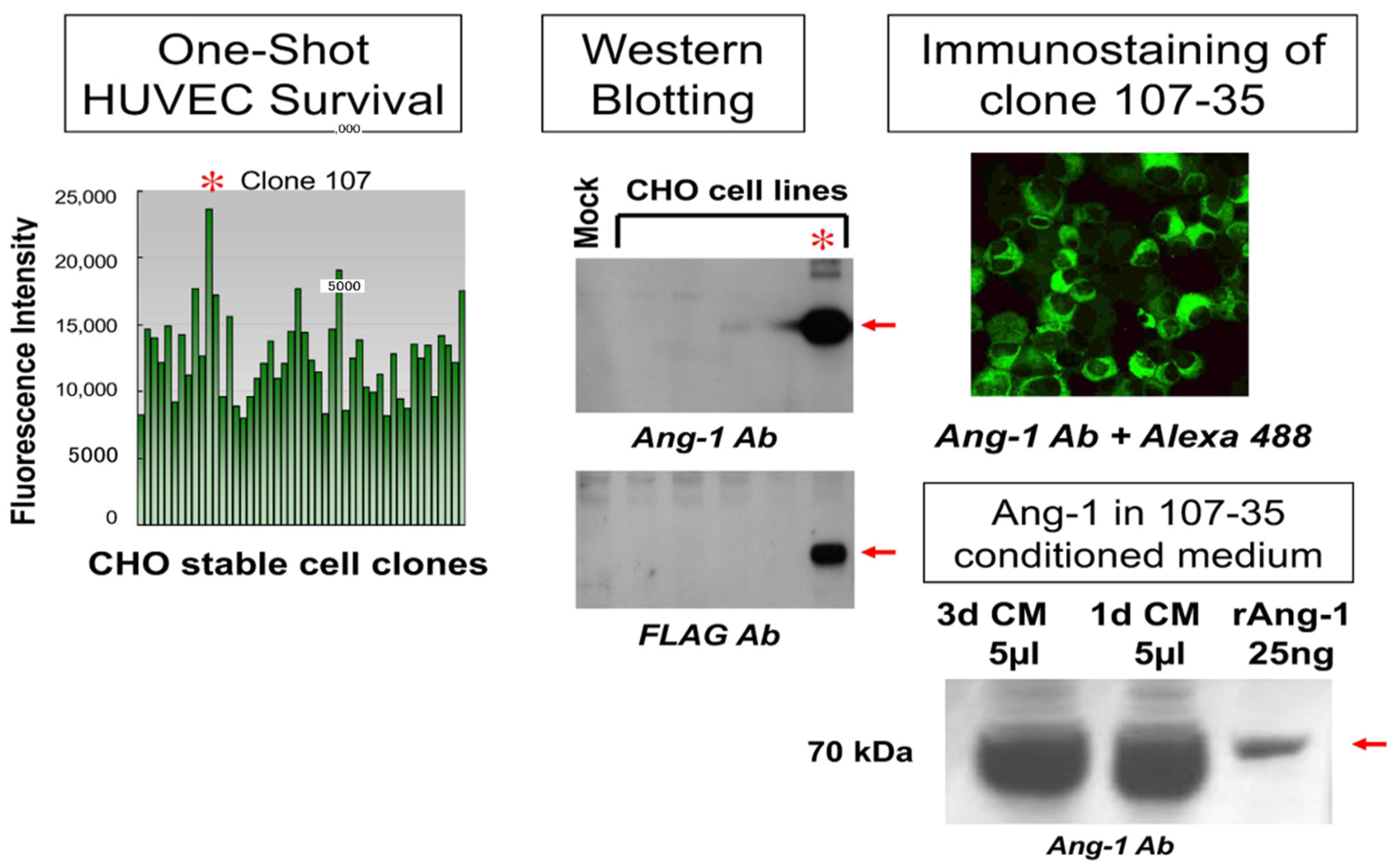
3.1. Establishment and Analysis of rAng-1-Producing CHO Cells
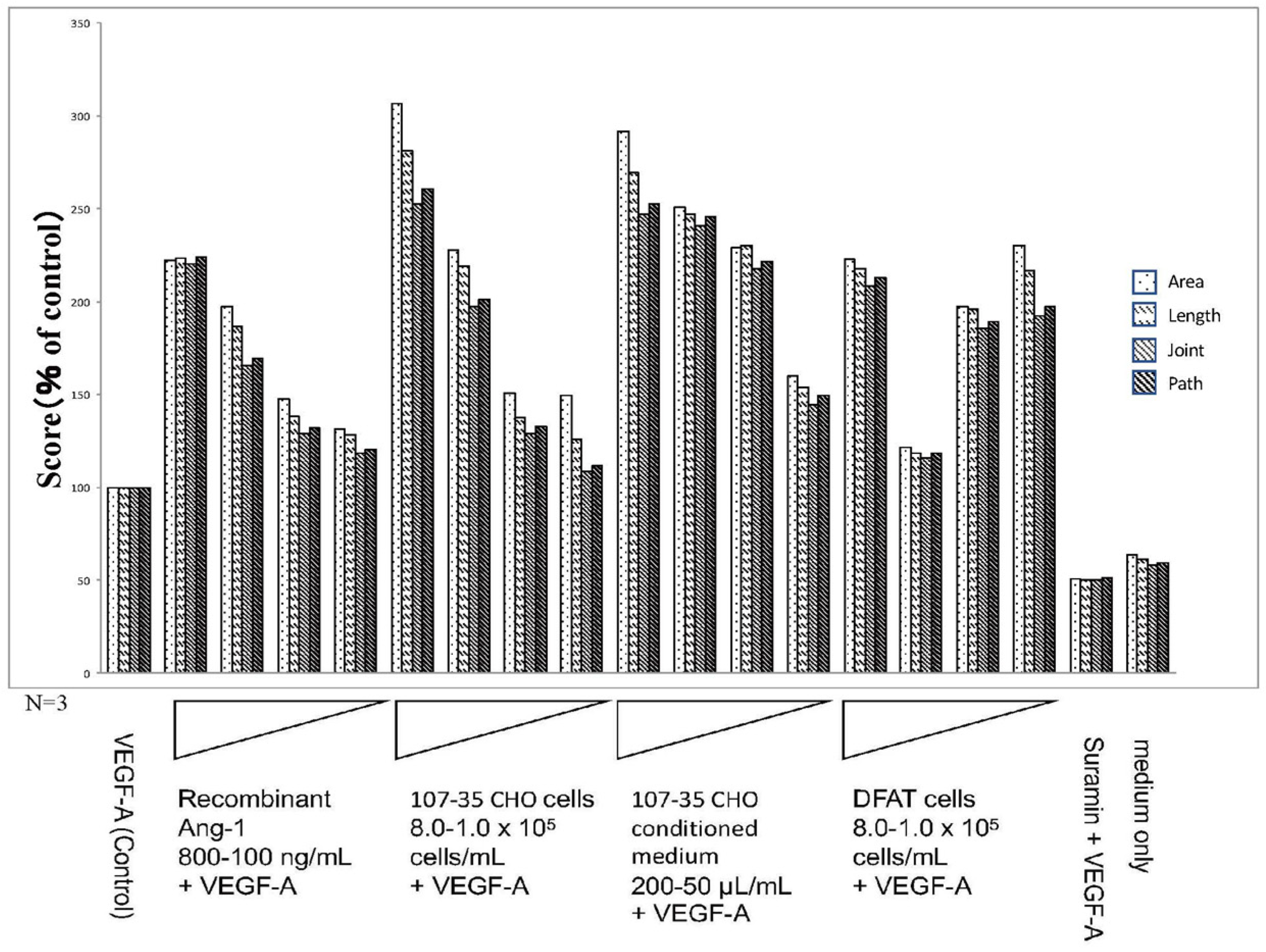
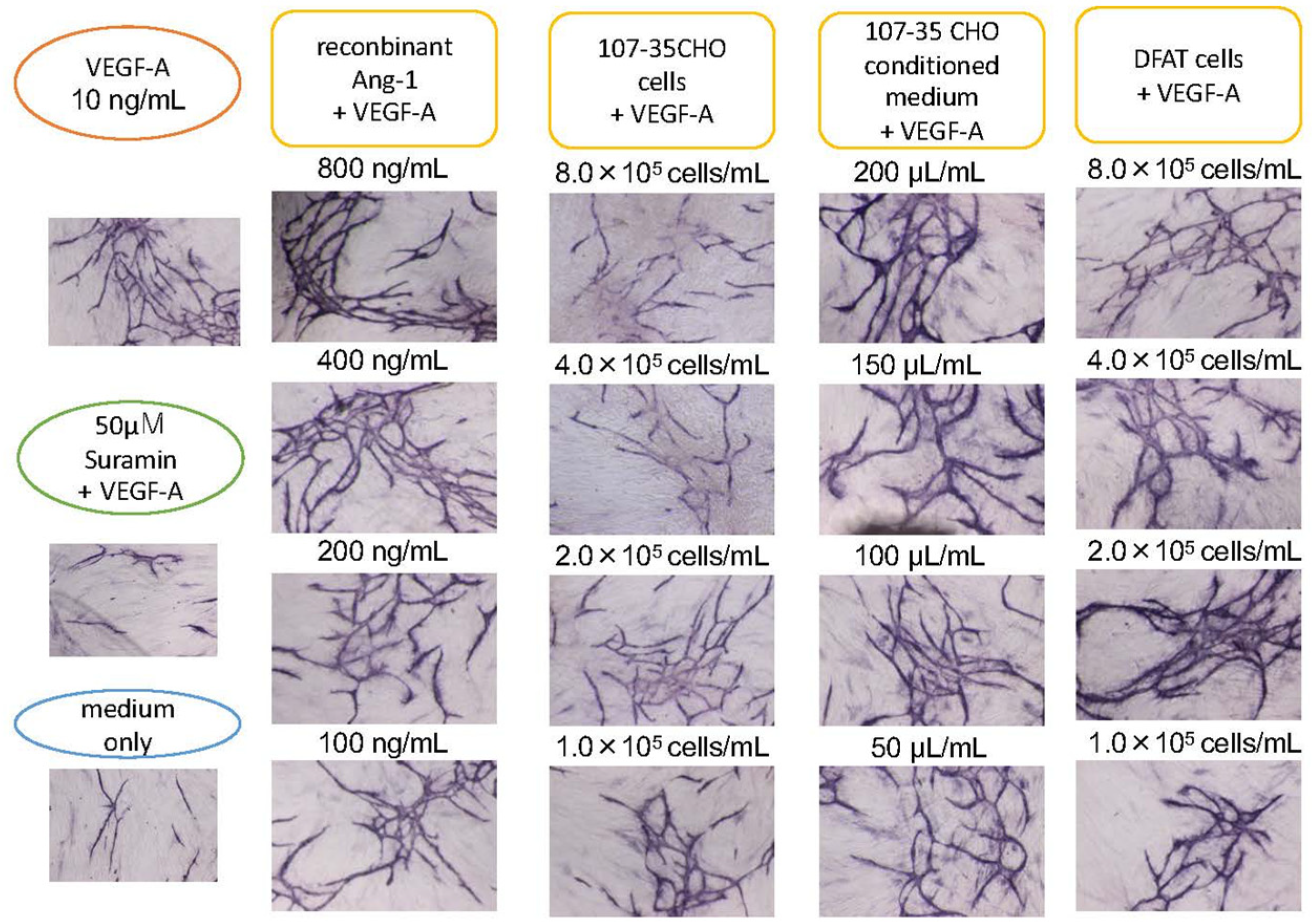
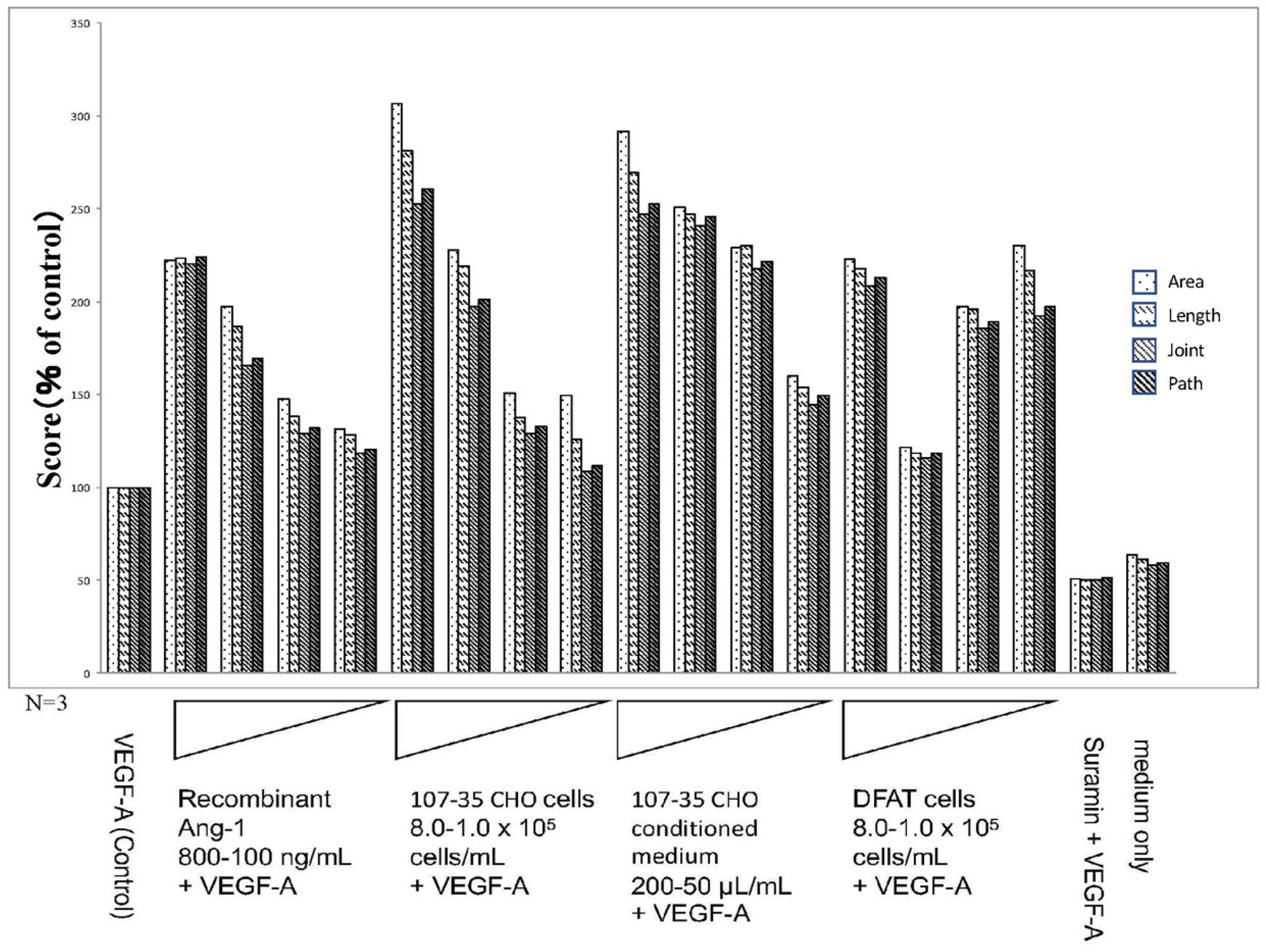
3.2. Promotion of Angiogenesis by rAng-1
3.3. Promotion of Angiogenesis by Co-Culture with DFAT-D1 Cells
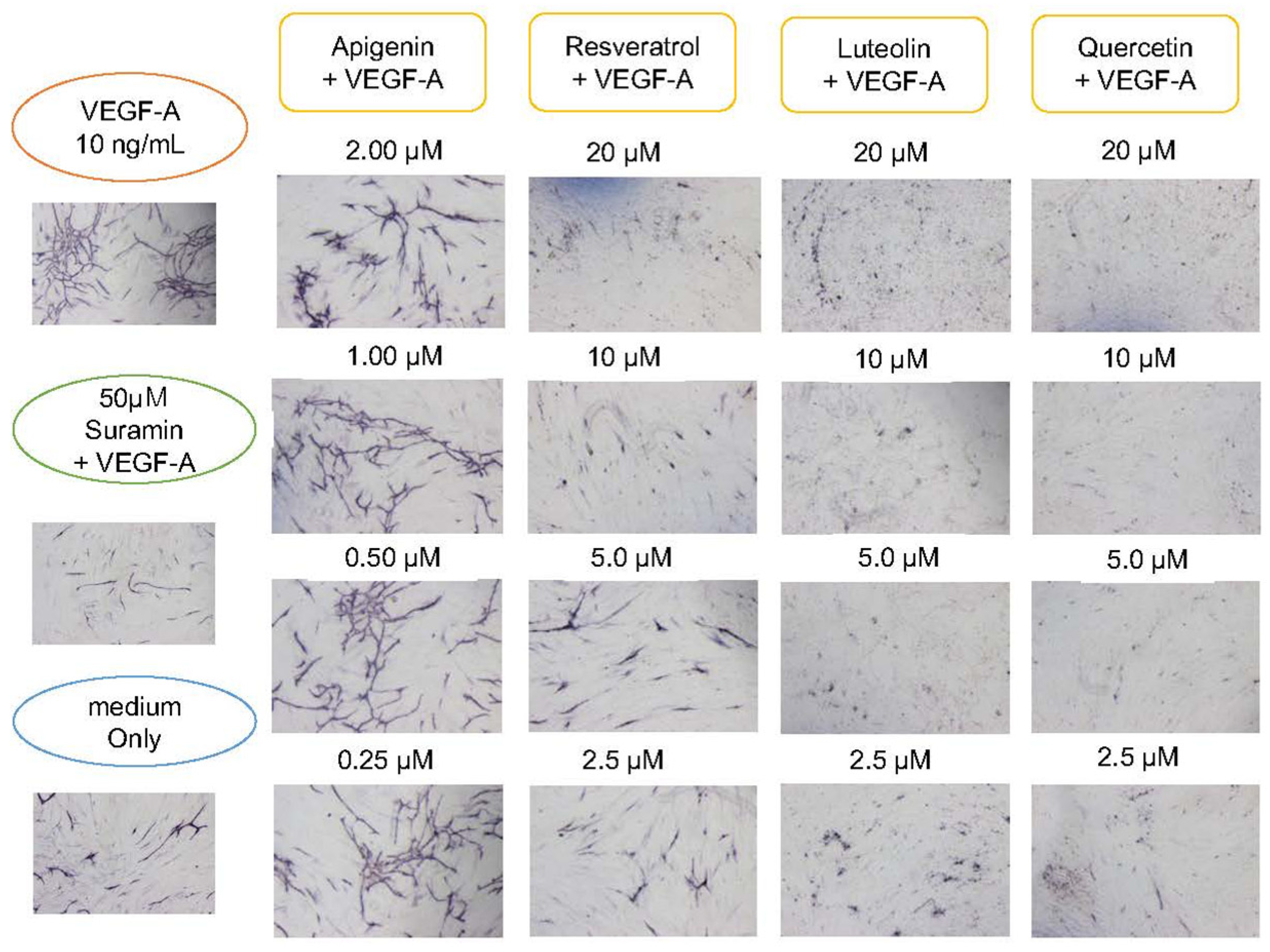
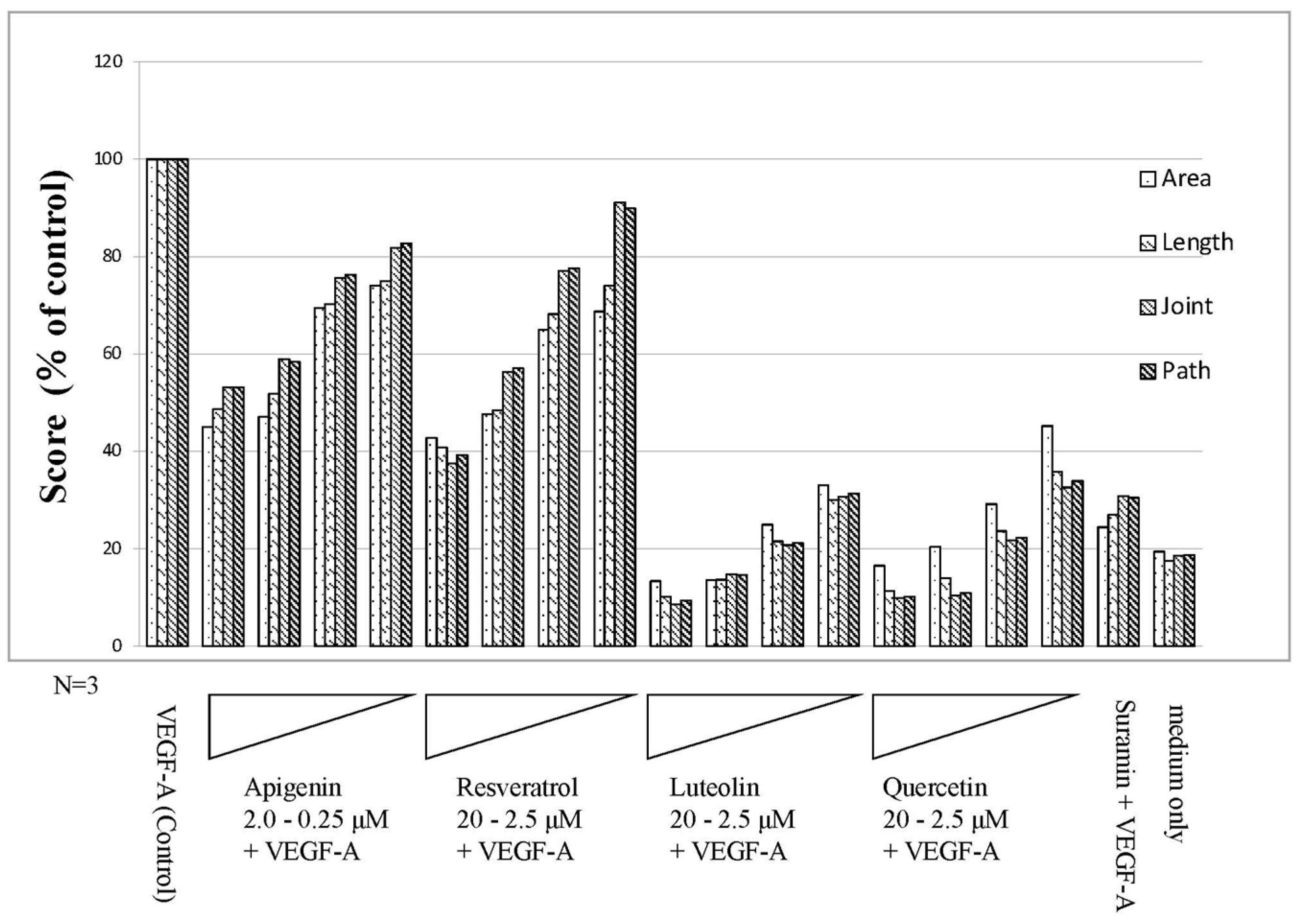
3.4. Angiogenesis Inhibitory Effect of Polyphenols
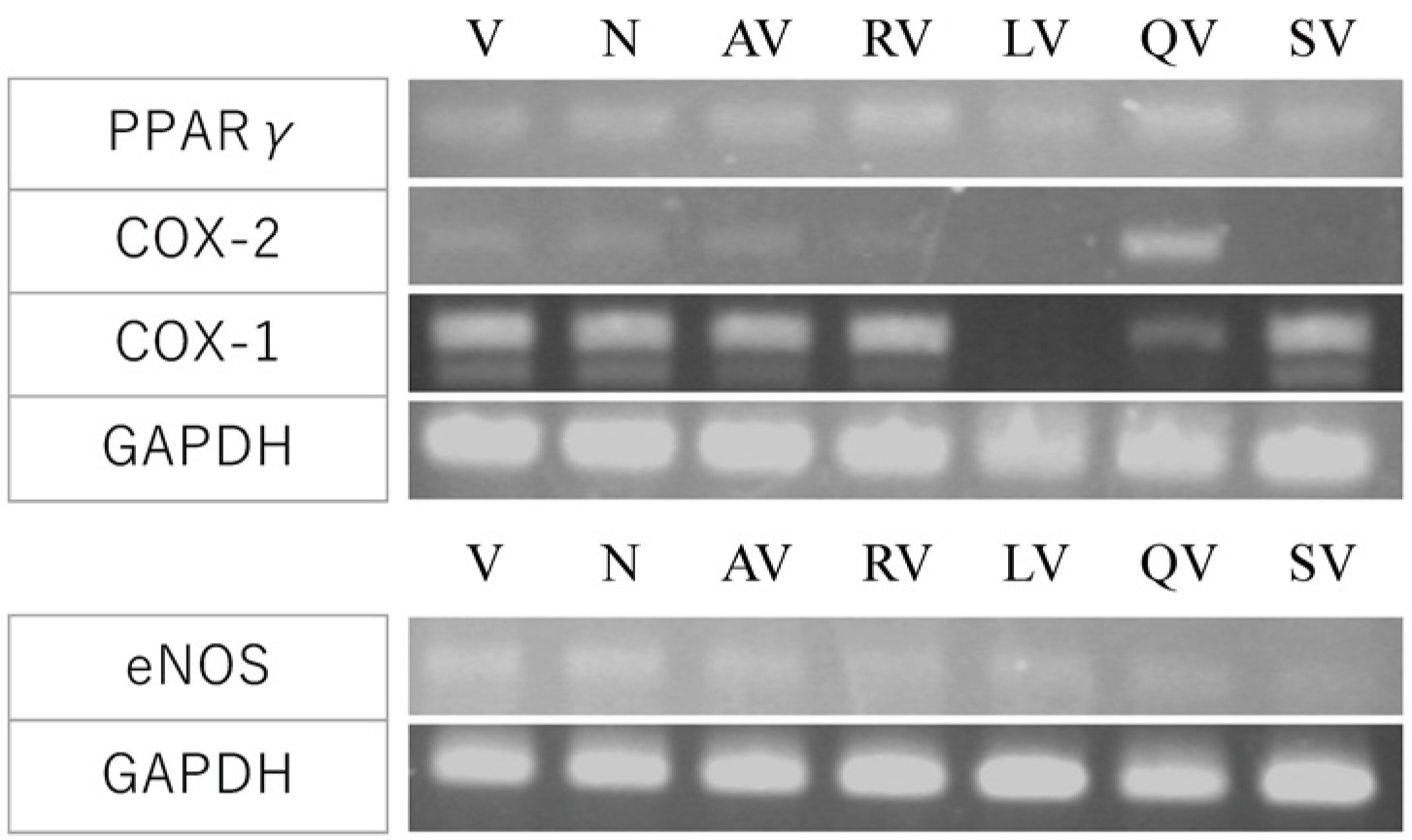
3.5. Investigation of the Molecular Mechanism by Gene Expression Analysis
4. Discussion
4.1. Angiogenesis-Promoting Effect of Ang-1
4.2. Angiogenesis Promoting Effect of DFAT
4.3. Angiogenesis Suppressing Effect of Polyphenol
4.4. Molecular Mechanisms
4.5. Future Perspective
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tepper, O.M.; Capla, J.M.; Galiano, R.D.; Ceradini, D.J.; Callaghan, M.J.; Kleinman, M.E.; Gurtner, G.C. Adult vasculogenesis occurs through in situ recruitment, proliferation, and tubulization of circulating bone marrow-derived cells. Blood 2005, 105, 1068–1077. [Google Scholar] [CrossRef] [Green Version]
- Hristov, M.; Erl, W.; Weber, P.C. Endothelial progenitor cells: Mobilization, differentiation, and homing. Arter. Thromb. Vasc. Biol. 2003, 23, 1185–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risau, W. Mechanisms of angiogenesis. Nature 1997, 386, 671–674. [Google Scholar] [CrossRef]
- Clapp, C.; Thebault, S.; Jeziorski, M.C.; Martinez De La Escalera, G. Peptide hormone regulation of angiogenesis. Physiol. Rev. 2009, 89, 1177–1215. [Google Scholar] [CrossRef]
- Folkman, J. Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Folkman, J.; Klagsburm, M. Angiogenic Factors. Science 1987, 235, 442–447. [Google Scholar] [CrossRef]
- Folkman, J. Angiogenesis: An organizing principle for drug discovery? Nat. Rev. Drug Discov. 2007, 6, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Masumoto, S.; Ono, A.; Ito, A.; Kawabe, Y.; Kamihira, M. Hypoxia-responsive expression of vascular endothelial growth factor for induction of angiogenesis in artificial three-dimensional tissues. J. Biosci. Bioeng. 2021, 132, 399–407. [Google Scholar] [CrossRef]
- Peters, E.B. Endothelial Progenitor Cells for the Vascularization of Engineered Tissues. Tissue Eng. Part B Rev. 2018, 24, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Kano, K.; Kondo, D.; Fukuda, N.; Iribe, Y.; Tanaka, N.; Matsubara, Y.; Sakuma, T.; Satomi, A.; Otaki, M.; et al. Mature adipocyte-derived dedifferentiated fat cells exhibit multilineage potential. J. Cell Physiol. 2007, 215, 210–222. [Google Scholar] [CrossRef]
- Watanabe, H.; Goto, S.; Kato, R.; Komiyama, S.; Nagaoka, Y.; Kazama, T.; Yamamoto, C.; Li, Y.; Konuma, N.; Hagikura, K.; et al. The neovascularization effect of dedifferentiated fat cells. Sci. Rep. 2020, 10, 9211. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Henzel, W.J. Pituitary follicular cells secrete a novel heparin-binding growth factor specific for vascular endothelial cells. Biochem. Biophys. Res. Commun. 1989, 161, 851–858. [Google Scholar] [CrossRef]
- Keck, P.J.; Hauser, S.D.; Krivi, G.; Sanzo, K.; Warren, T.; Feder, J.; Connolly, D.T. Vascular permeability factor, an endothelial cell mitogen related to PDGF. Science 1989, 246, 1306–1312. [Google Scholar] [CrossRef]
- Ferrara, N.; Davis-Smyth, T. The biology of vascular endothelial growth factor. Endocri. Rev. 1997, 18, 4–25. [Google Scholar] [CrossRef]
- Williams, B. Vascular permeability/vascular endothelial growth factors: A potential role in the pathogenesis and treatment of vascular diseases. Vasc. Med. 1996, 1, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D. Signaling vascular morphogenesis and maintenance. Science 1997, 277, 48–50. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid Med. Cell Longev. 2010, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.H. Flavonoids inhibit VEGF/bFGF-induced angiogenesis in vitro by inhibiting the matrix-degrading proteases. J. Cell. Biochem. 2003, 89, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Bagli, E.; Stefaniotou, M.; Morbidelli, L.; Ziche, M.; Psillas, K.; Murphy, C.; Fotsis, T. Luteolin Inhibits Vascular Endothelial Growth Factor-Induced Angiogenesis; Inhibition of Endothelial Cell Survival and Proliferation by Targeting Phosphatidylinositol 3′-Kinase Activity. Cancer Res. 2004, 64, 7936–7946. [Google Scholar] [CrossRef] [Green Version]
- Lam, K.H.; Alex, D.; Lam, I.K.; Tsui, S.K.W.; Yang, Z.F.; Lee, S.M.Y. Nobiletin, a Polymethoxylated Flavonoid from Citrus, Shows Anti-Angiogenic Activity in a Zebrafish In Vivo Model and HUVEC In Vitro Model. J. Cell. Biochem. 2011, 112, 3313–3321. [Google Scholar] [CrossRef] [PubMed]
- Bouloumie, A.; Schini-Kerth, V.B.; Busse, R. Vascular endothelial growth factor up-regulates nitric oxide synthase expression in endothelial cells. Cardiovasc. Res. 1999, 41, 773–780. [Google Scholar] [CrossRef] [Green Version]
- Goncharov, N.V.; Popova, P.I.; Avdonin, P.P.; Kudryavtsev, I.V.; Serebryakova, M.K.; Korf, E.A.; Avdonin, P.V. Markers of Endothelial Cells in Normal and Pathological Conditions. Biochem. (Mosc.) Suppl. Ser. A Membr. Cell Biol. 2020, 14, 167–183. [Google Scholar] [CrossRef]
- Wu, A.W.; Gu, J.; Li, Z.F.; Ji, J.F.; Xu, G.W. COX-2 expression and tumor angiogenesis in colorectal cancer. World J. Gastroenterol. 2004, 10, 2323–2326. [Google Scholar] [CrossRef] [Green Version]
- Meade, E.A.; McIntyre, T.M.; Zimmerman, G.A.; Prescott, S.M. Peroxisome proliferators enhance cyclooxygenase-2 expression in epithelial cells. J. Biol. Chem. 1999, 274, 8328–8334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.L.; Frucht, H. Activation of the PPAR pathway induces apoptosis and COX-2 inhibition in HT-29 human colon cancer cells. Carcinogenesis 2001, 22, 1379–1383. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Luo, J.; Rana, J.S.; Laham, R.; Sellke, F.W.; Li, J. Involvement of COX-2 in VEGF-induced angiogenesis via P38 and JNK pathways in vascular endothelial cells. Cardiovasc. Res. 2006, 69, 512–519. [Google Scholar] [CrossRef] [Green Version]
- Ho, T.C.; Chen, S.L.; Yang, Y.C.; Liao, C.L.; Cheng, H.C.; Tsao, Y.P. PEDF induces p53-mediated apoptosis through PPAR gamma signaling in human umbilical vein endothelial cells. Cardiovasc. Res. 2007, 76, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Yagi, K.; Kondo, D.; Okazaki, Y.; Kano, K. A novel preadipocyte cell line established from mouse adult mature adipocytes. Biochem. Biophys Res. Commun. 2004, 321, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Jeansson, M.; Gawlik, A.; Anderson, G.; Li, C.; Kerjaschki, D.; Henkelman, M.; Quaggin, S.E. Angiopoietin-1 is essential in mouse vasculature during development and in response to injury. J. Clin. Investig. 2011, 121, 2278–2289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindblom, P.; Gerhardt, H.; Liebner, S.; Abramsson, A.; Enge, M.; Hellstrom, M.; Backstrom, G.; Fredriksson, S.; Landegren, U.; Nystrom, H.C.; et al. Endothelial PDGF-B retention is required for proper investment of pericytes in the microvessel wall. Genes Dev. 2003, 17, 1835–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Kazama, T.; Nagaoka, Y.; Inamo, Y.; Mugishima, H.; Takahashi, S.; Matsumoto, T. Influence of Donor Age and Passage Number on Angiogenic Activity in Human Adipose-Derived Stem Cell-Conditioned Media. J. Stem. Cell Res. Ther. 2015, 5, 307. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Agarwal, R.; Lu, J. Anti-angiogenic potential of a cancer chemopreventive flavonoid antioxidant, silymarin: Inhibition of key attributes of vascular endothelial cells and angiogenic cytokine secretion by cancer epithelial cells. Biochem. Biophys. Res. Commun. 2000, 276, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, J.; Itoh, H.; Hirashima, M.; Ogawa, M.; Nishikawa, S.; Yurugi, T.; Naito, M.; Nakao, K.; Nishikawa, S. Flk1-positive cells derived from embryonic stem cells serve as vascular progenitors. Nature 2000, 408, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Potente, M.; Ghaeni, L.; Baldessari, D.; Mostoslavsky, R.; Rossig, L.; Dequiedt, F.; Haendeler, J.; Mione, M.; Dejana, E.; Alt, F.W.; et al. SIRT1 controls endothelial angiogenic functions during vascular growth. Gene Dev. 2007, 21, 2644–2658. [Google Scholar] [CrossRef] [Green Version]
- Nakata, R.; Takahashi, S.; Inoue, H. Recent advances in the study on resveratrol. Biol. Pharm. Bull. 2012, 35, 273–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, G.H.; Jang, E.S.; Kim, J.W.; Jeong, S.H. Prognostic role of plasma level of angiopoietin-1, angiopoietin-2, and vascular endothelial growth factor in hepatocellular carcinoma. World J. Gastroenterol. 2021, 27, 4453–4467. [Google Scholar] [CrossRef] [PubMed]
- Okumo, T.; Furuta, A.; Kimura, T.; Yusa, K.; Asano, K.; Sunagawa, M. Inhibition of Angiogenic Factor Productions by Quercetin In Vitro and In Vivo. Medicines 2021, 8, 22. [Google Scholar] [CrossRef]
- Gobbi, S.; Belluti, F.; Rampa, A.; Bisi, A. Flavonoid-Inspired Vascular Disrupting Agents: Exploring Flavone-8-Acetic Acid and Derivatives in the New Century. Molecules 2021, 26, 4228. [Google Scholar] [CrossRef] [PubMed]
- Forni, C.; Rossi, M.; Borromeo, I.; Feriotto, G.; Platamone, G.; Tabolacci, C.; Mischiati, C.; Beninati, S. Flavonoids: A Myth or a Reality for Cancer Therapy? Molecules 2021, 26, 3583. [Google Scholar] [CrossRef]
- Hu, Z.; Zhao, G.; Gou, W.; Cheng, H. Myricitrin inhibits vascular endothelial growth factor-induced angiogenesis of human umbilical vein endothelial cells and mice. Biomed. Pharm. 2020, 130, 110726. [Google Scholar] [CrossRef]
- Jumabay, M.; Abdmaulen, R.; Urs, S.; Heydarkhan-Hagvall, S.; Chazenbalk, G.D.; Jordan, M.C.; Roos, K.P.; Yao, Y.; Bostrom, K.I. Endothelial differentiation in multipotent cells derived from mouse and human white mature adipocytes. J. Mol. Cell Cardiol. 2012, 53, 790–800. [Google Scholar] [CrossRef] [Green Version]
- Rizzi, A.; Benagian, V.; Ribatti, D. Angiogenesis versus arteriogenesis. Rom. J. Morphol. Embryol. 2017, 58, 15–19. [Google Scholar] [PubMed]
- Zaki, M.M.; Lesha, E.; Said, K.; Kiaee, K.; Robinson-McCarthy, L.; George, H.; Hanna, A.; Appleton, E.; Liu, S.; Ng, A.H.M.; et al. Cell therapy strategies for COVID-19: Current approaches and potential applications. Sci. Adv. 2021, 7, eabg5995. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimazaki, T.; Noro, N.; Hagikura, K.; Matsumoto, T.; Yoshida-Noro, C. Quantitative Analysis of Factors Regulating Angiogenesis for Stem Cell Therapy. Biology 2021, 10, 1212. https://doi.org/10.3390/biology10111212
Shimazaki T, Noro N, Hagikura K, Matsumoto T, Yoshida-Noro C. Quantitative Analysis of Factors Regulating Angiogenesis for Stem Cell Therapy. Biology. 2021; 10(11):1212. https://doi.org/10.3390/biology10111212
Chicago/Turabian StyleShimazaki, Takahiro, Nobuhiro Noro, Kazuhiro Hagikura, Taro Matsumoto, and Chikako Yoshida-Noro. 2021. "Quantitative Analysis of Factors Regulating Angiogenesis for Stem Cell Therapy" Biology 10, no. 11: 1212. https://doi.org/10.3390/biology10111212
APA StyleShimazaki, T., Noro, N., Hagikura, K., Matsumoto, T., & Yoshida-Noro, C. (2021). Quantitative Analysis of Factors Regulating Angiogenesis for Stem Cell Therapy. Biology, 10(11), 1212. https://doi.org/10.3390/biology10111212