
Two-Dimensional Gel Electrophoresis to Study the Activity of Type IIA Topoisomerases on Plasmid Replication Intermediates
 ,
,  and
and 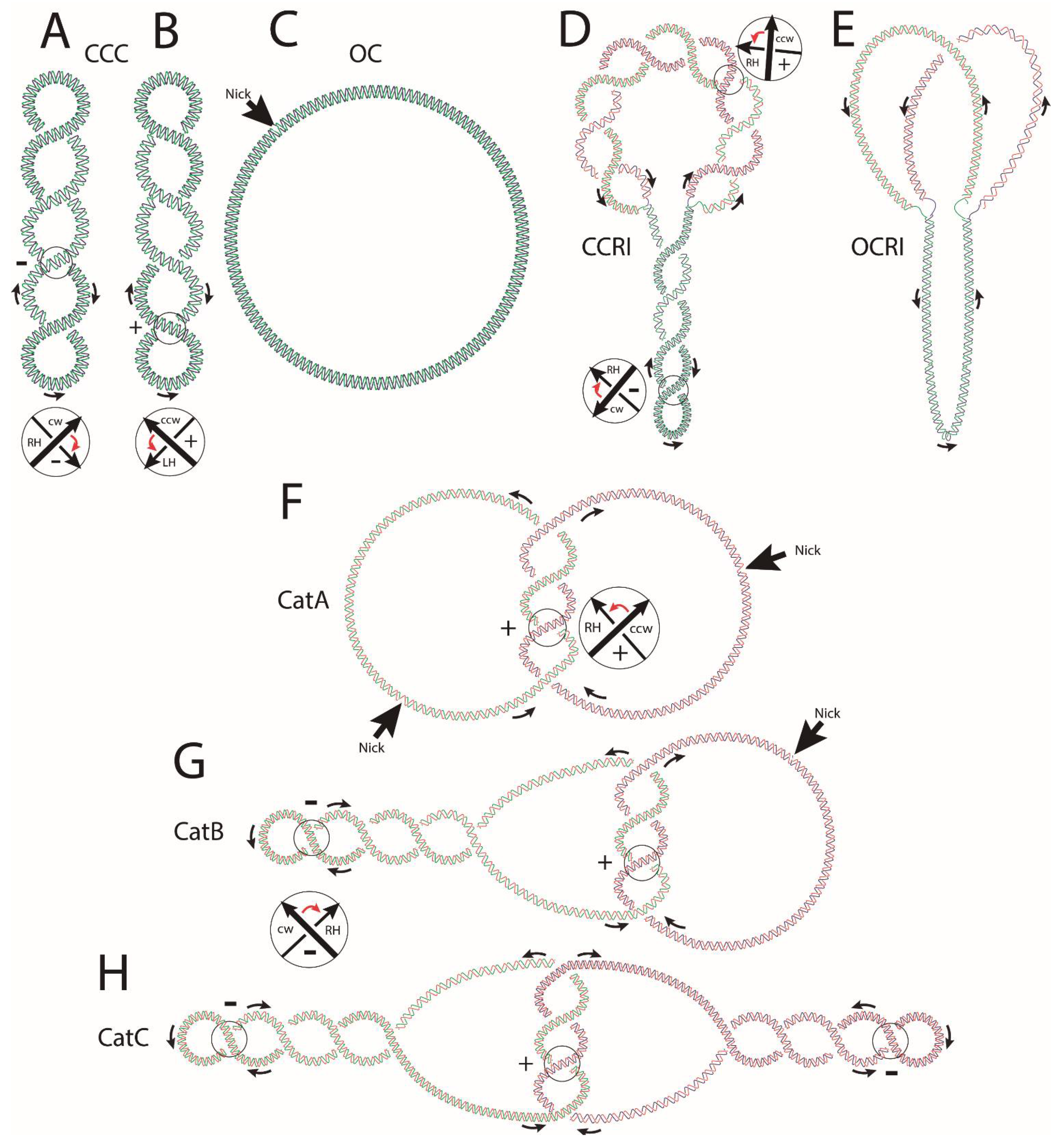{kind=link}
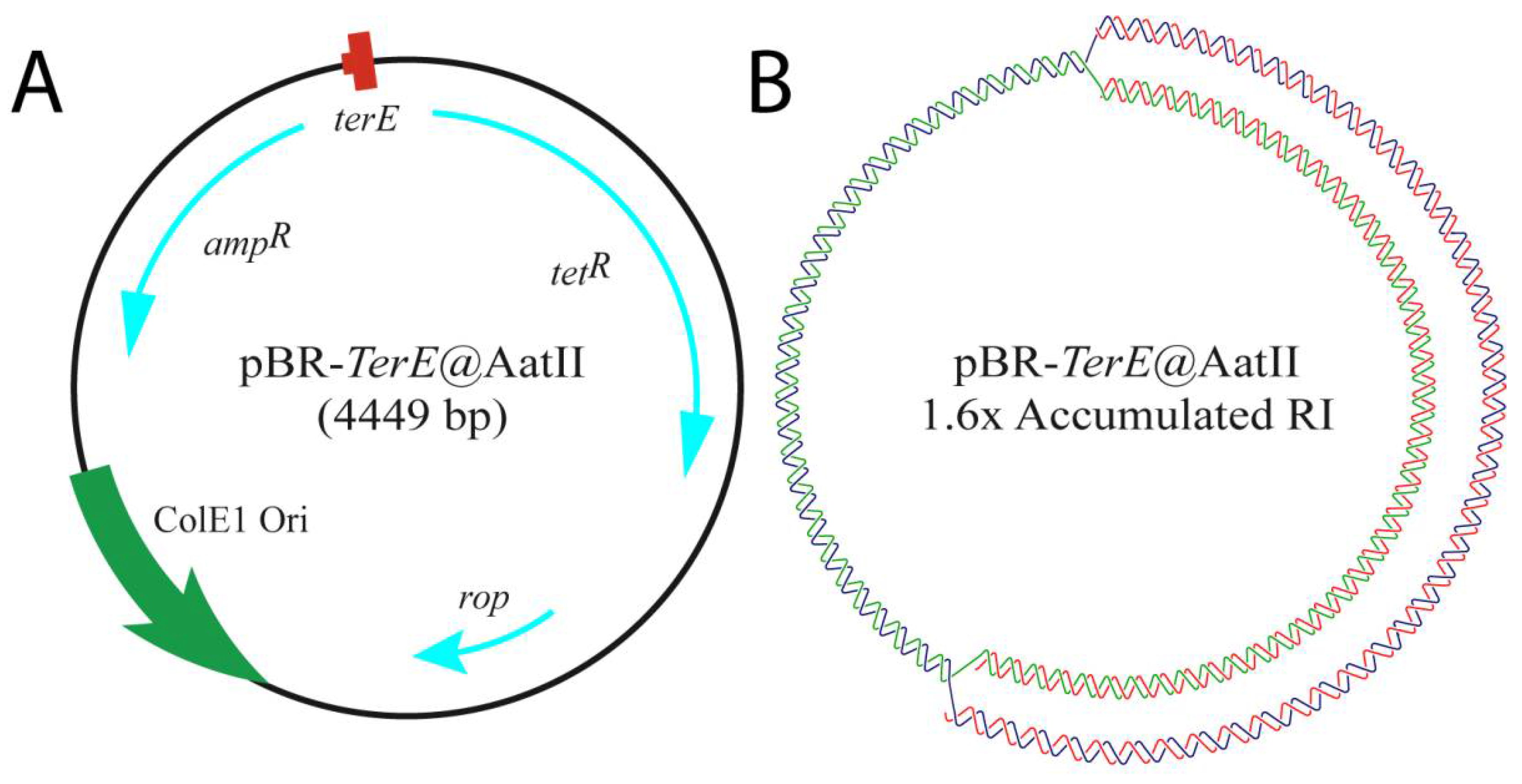{kind=link}
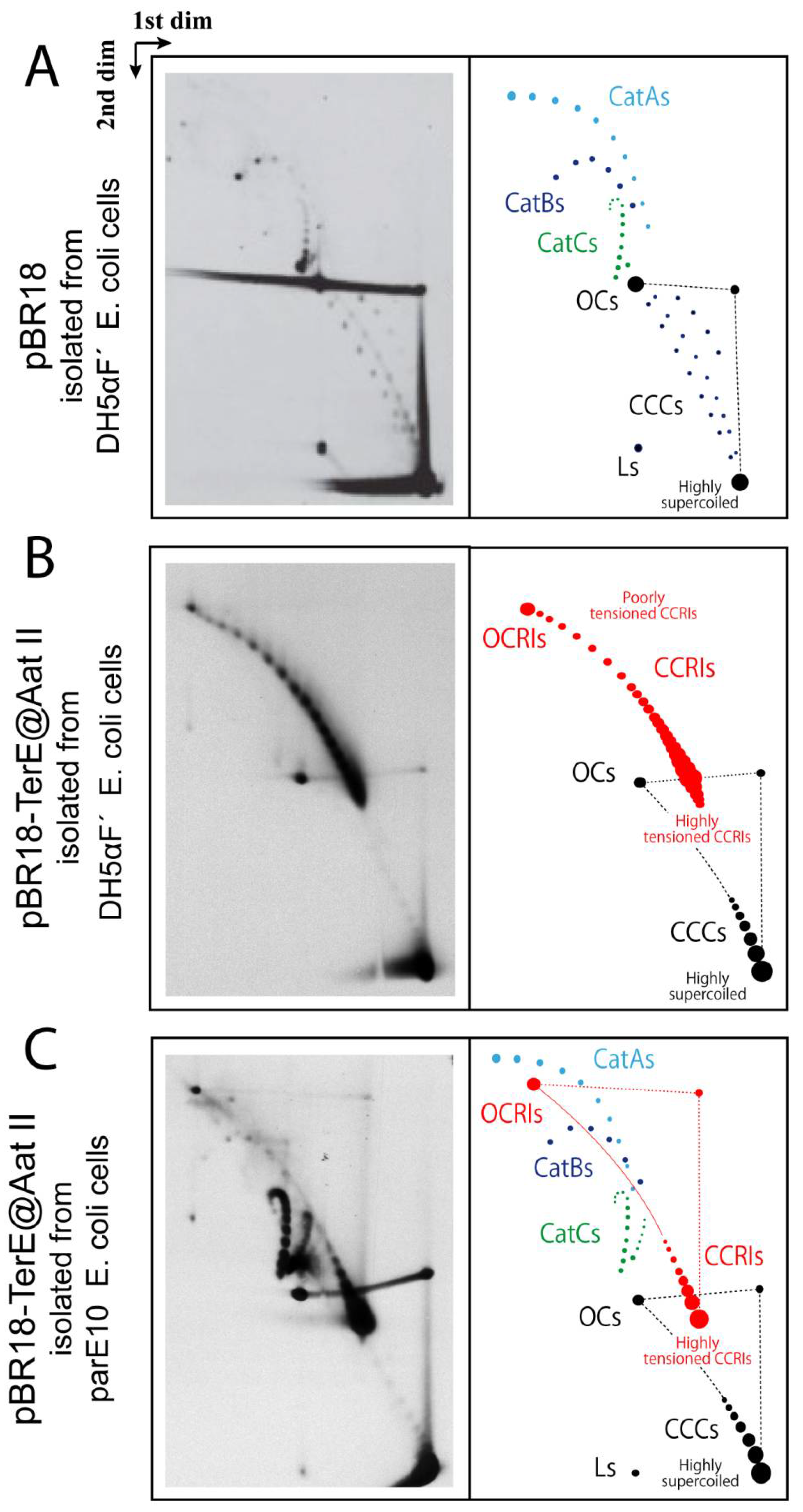{kind=link}
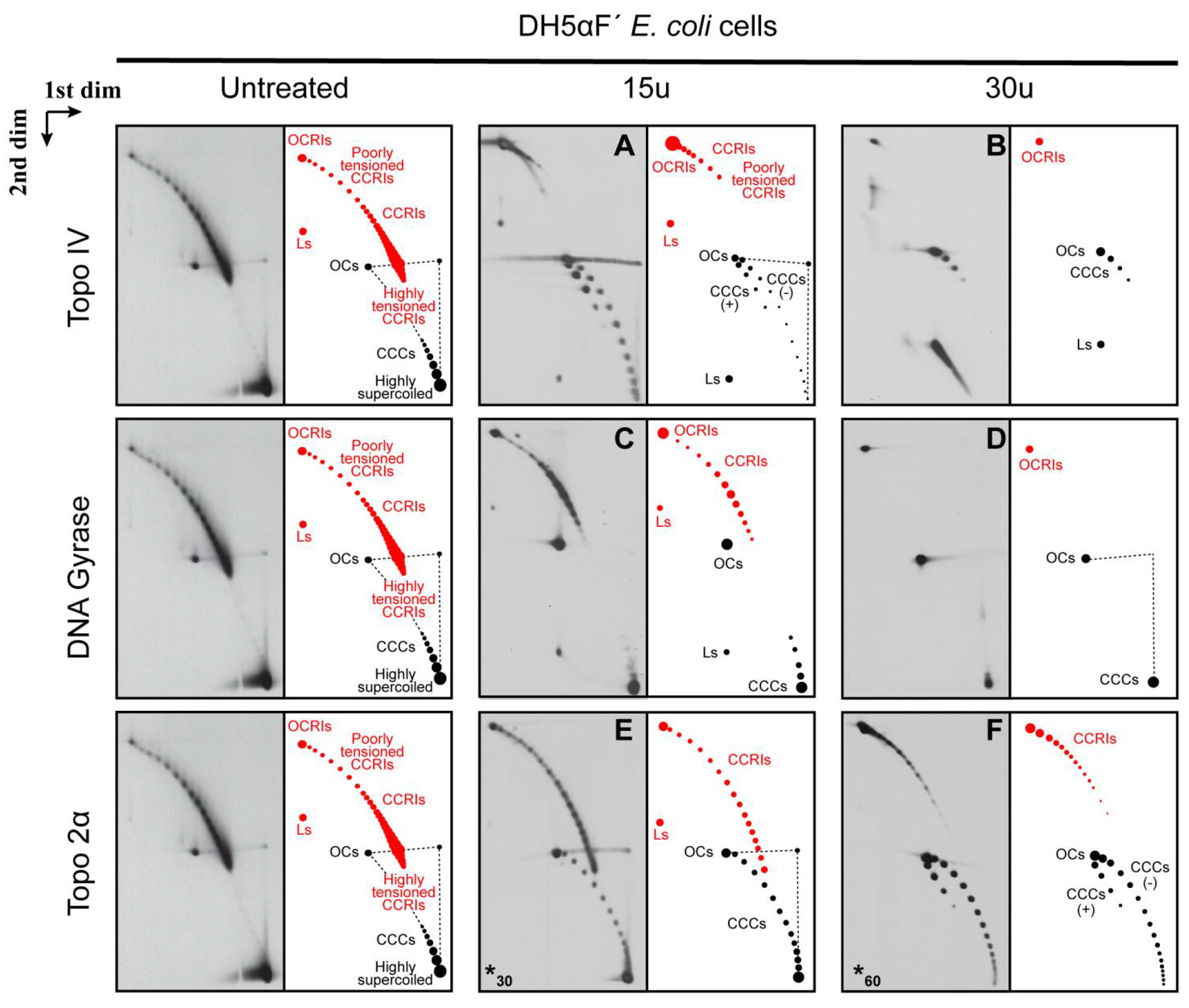{kind=link}
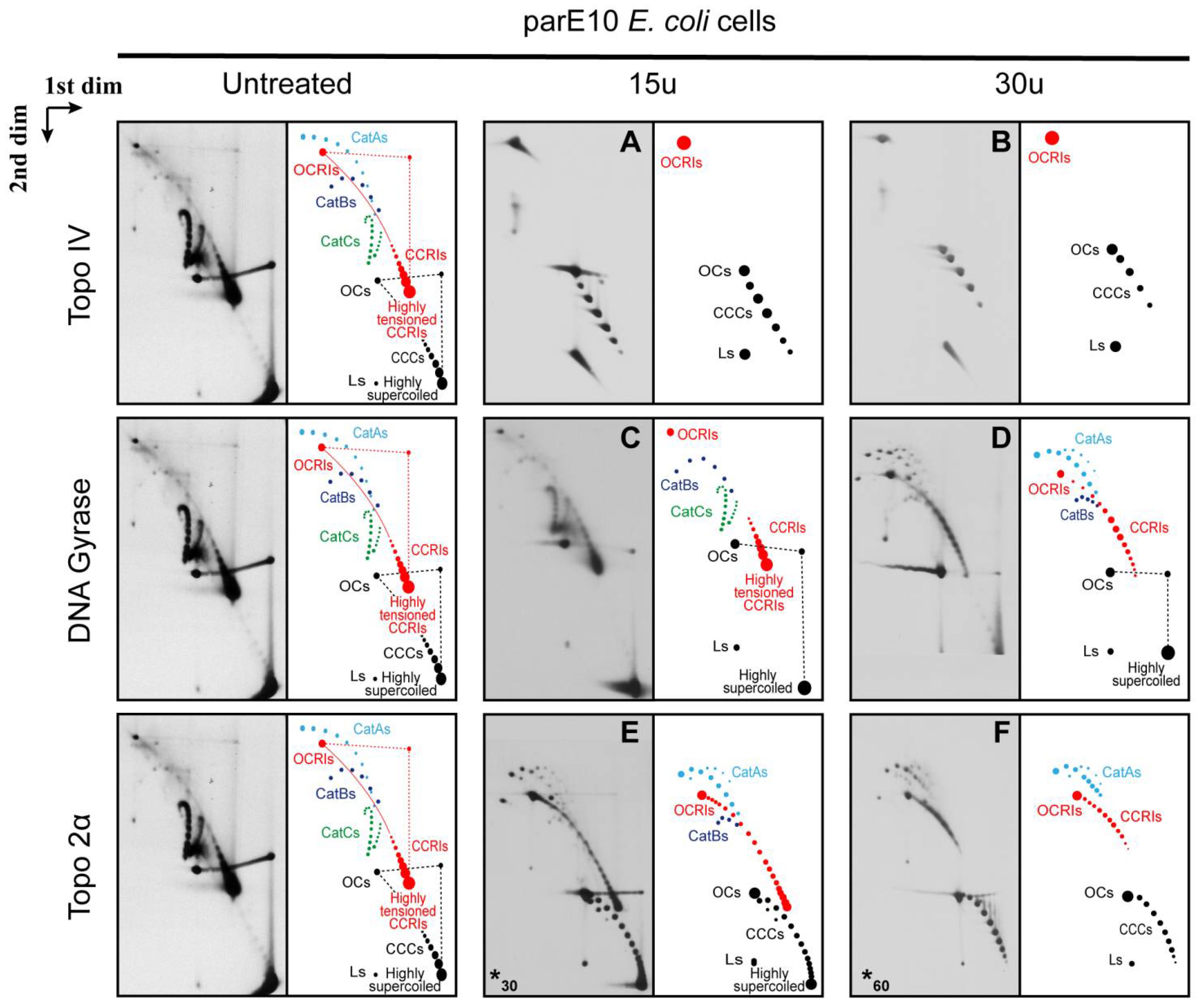{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids and Culture Medium
2.2. DNA Treatments
2.3. Two-Dimensional Agarose Gel Electrophoresis and Southern Transfer
2.4. Non-Radioactive Hybridization
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Champoux, J.J.; Been, M.D. Topoisomerases and the swivel problem. In Mechanistic Studies of DNA Replication and Genetic Recombination; Alberts, B., Ed.; ICN-UCLA Symposia on Molecular and Cellular Biology; Academic Press: New York, NY, USA, 1980; pp. 809–815. [Google Scholar]
- Forterre, P.; Gadelle, D. Phylogenomics of DNA topoisomerases: Their origin and putative roles in the emergence of modern organisms. Nucleic Acids Res. 2009, 37, 679–692. [Google Scholar] [CrossRef]
- Watson, J.D.; Crick, F.H.C. Molecular structure of nucleic acids. Nature 1953, 161, 737–738. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, J.W.; Greenall, R.J.; Probert, M.I.J.; Noy, A.; Leake, M.C. The emergence of sequence-dependent structural motifs in stretched, torsionally constrained DNA. Nucleic Acids Res. 2020, 48, 1748–1763. [Google Scholar] [CrossRef] [PubMed]
- Bates, A.D.; Maxwell, A. DNA Topology; Oxford University Press: Oxford, UK, 2005; p. 216. [Google Scholar]
- Schvartzman, J.B.; Hernandez, P.; Krimer, D.B.; Dorier, J.; Stasiak, A. Closing the DNA replication cycle: From simple circular molecules to supercoiled and knotted DNA catenanes. Nucleic Acids Res. 2019, 47, 7182–7198. [Google Scholar] [CrossRef]
- Lilley, D.M. DNA supercoiling and DNA structure. Biochem. Soc. Trans. 1986, 14, 211–213. [Google Scholar] [CrossRef] [PubMed]
- Ullsperger, C.; Vologodskii, A.A.; Cozzarelli, N.R. Unlinking of DNA by topoisomerases during DNA replication. In Nucleic Acids and Molecular Biology; Lilley, D.M.J., Eckstein, F., Eds.; Springer: Berlin, Germany, 1995; pp. 115–142. [Google Scholar]
- Schvartzman, J.B.; Stasiak, A. A topological view of the replicon. EMBO Rep. 2004, 5, 256–261. [Google Scholar] [CrossRef]
- Wang, J. Untangling the Double Helix; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009; p. 233. [Google Scholar]
- Martinez-Robles, M.L.; Witz, G.; Hernandez, P.; Schvartzman, J.B.; Stasiak, A.; Krimer, D.B. Interplay of DNA supercoiling and catenation during the segregation of sister duplexes. Nucleic Acids Res. 2009, 37, 5126–5137. [Google Scholar] [CrossRef]
- Kornberg, A.; Baker, T.A. DNA Replication, 2nd ed.; W.H. Freeman and Co.: New York, NY, USA, 1992; p. 850. [Google Scholar]
- Neuman, K.C. Evolutionary twist on topoisomerases: Conversion of gyrase to topoisomerase IV. Proc. Natl. Acad. Sci. USA 2010, 107, 22363–22364. [Google Scholar] [CrossRef]
- Tretter, E.M.; Berger, J.M. Mechanisms for defining supercoiling set point of DNA gyrase orthologs: I. A nonconserved acidic C-terminal tail modulates Escherichia coli gyrase activity. J. Biol. Chem. 2012, 287, 18636–18644. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C. Interaction between DNA and an Escherichia coli protein omega. J. Mol. Biol. 1971, 55, 523–533. [Google Scholar] [CrossRef]
- Gellert, M.; Mizuuchi, K.; O’Dea, M.H.; Nash, H.A. DNA gyrase: An enzyme that introduces superhelical turns into DNA. Proc. Natl. Acad. Sci. USA 1976, 73, 3872–3876. [Google Scholar] [CrossRef]
- Dean, F.; Krasnow, M.A.; Otter, R.; Matzuk, M.M.; Spengler, S.J.; Cozzarelli, N.R. Escherichia coli type-1 topoisomerases: Identification, mechanism, and role in recombination. Cold Spring Harb. Symp. Quant. Biol. 1983, 47, 769–777. [Google Scholar] [CrossRef]
- Kato, J.; Nishimura, Y.; Imamura, R.; Niki, H.; Hiraga, S.; Suzuki, H. New topoisomerase essential for chromosome segregation in E. coli. Cell 1990, 63, 393–404. [Google Scholar] [CrossRef]
- Kato, J.; Suzuki, H.; Ikeda, H. Purification and characterization of DNA topoisomerase IV in Escherichia coli. J. Biol. Chem. 1992, 267, 25676–25684. [Google Scholar] [CrossRef]
- Drlica, K. Control of bacterial DNA supercoiling. Mol. Microbiol. 1992, 6, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Kellner, U.; Rudolph, P.; Parwaresch, R. Human DNA-Topoisomerases—Diagnostic and Therapeutic Implications for Cancer. Onkologie 2000, 23, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Zechiedrich, E.L.; Cozzarelli, N.R. Roles of topoisomerase IV and DNA gyrase in DNA unlinking during replication in Escherichia coli. Genes Dev. 1995, 9, 2859–2869. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.E.; Shekhtman, E.M.; Zechiedrich, E.L.; Schmid, M.B.; Cozzarelli, N.R. The role of topoisomerase IV in partitioning bacterial replicons and the structure of catenated intermediates in DNA replication. Cell 1992, 71, 277–288. [Google Scholar] [CrossRef]
- Zechiedrich, E.L.; Khodursky, A.B.; Cozzarelli, N.R. Topoisomerase IV, not gyrase, decatenates products of site-specific recombination in Escherichia coli. Genes Dev. 1997, 11, 2580–2592. [Google Scholar] [CrossRef] [PubMed]
- Deibler, R.W.; Rahmati, S.; Zechiedrich, E.L. Topoisomerase IV, alone, unknots DNA in E-coli. Genes Dev. 2001, 15, 748–761. [Google Scholar] [CrossRef]
- Lopez, V.; Martinez-Robles, M.L.; Hernandez, P.; Krimer, D.B.; Schvartzman, J.B. Topo IV is the topoisomerase that knots and unknots sister duplexes during DNA replication. Nucleic Acids Res. 2012, 40, 3563–3573. [Google Scholar] [CrossRef] [PubMed]
- DiNardo, S.; Voelkel, K.A.; Sternglanz, R.; Reynolds, A.E.; Wright, A. Escherichia coli DNA topoisomerase I mutants have compensatory mutations in DNA gyrase genes. Cell 1982, 31, 43–51. [Google Scholar] [CrossRef]
- Drolet, M.; Phoenix, P.; Menzel, R.; Masse, E.; Liu, L.F.; Crouch, R.J. Overexpression of RNase H partially complements the growth defect of an Escherichia coli delta topA mutant: R-loop formation is a major problem in the absence of DNA topoisomerase I. Proc. Natl. Acad. Sci. USA 1995, 92, 3526–3530. [Google Scholar] [CrossRef] [PubMed]
- Reuss, D.R.; Fasshauer, P.; Mroch, P.J.; Ul-Haq, I.; Koo, B.M.; Pohlein, A.; Gross, C.A.; Daniel, R.; Brantl, S.; Stulke, J. Topoisomerase IV can functionally replace all type 1A topoisomerases in Bacillus subtilis. Nucleic Acids Res. 2019, 47, 5231–5242. [Google Scholar] [CrossRef]
- Brochu, J.; Breton, E.V.; Drolet, M. Supercoiling, R-loops, Replication and the Functions of Bacterial Type 1A Topoisomerases. Genes 2020, 11, 249. [Google Scholar] [CrossRef]
- Brochu, J.; Vlachos-Breton, E.; Sutherland, S.; Martel, M.; Drolet, M. Topoisomerases I and III inhibit R-loop formation to prevent unregulated replication in the chromosomal Ter region of Escherichia coli. PLoS Genet 2018, 14, e1007668. [Google Scholar] [CrossRef]
- Lee, C.M.; Wang, G.; Pertsinidis, A.; Marians, K.J. Topoisomerase III Acts at the Replication Fork To Remove Precatenanes. J. Bacteriol. 2019, 201, e00563-18. [Google Scholar] [CrossRef]
- Keller, W. Determination of the number of superhelical turns in simian virus 40 DNA by gel electrophoresis. Proc. Natl. Acad. Sci. USA 1975, 72, 4876–4880. [Google Scholar] [CrossRef]
- Thorne, H.V. Electrophoretic separation of polyoma virus DNA from host cell DNA. Virology 1966, 29, 234–239. [Google Scholar] [CrossRef]
- Sundin, O.; Varshavsky, A. Terminal stages of SV40 DNA replication proceed via multiply intertwined catenated dimers. Cell 1980, 21, 103–114. [Google Scholar] [CrossRef]
- Brewer, B.J.; Fangman, W.L. The localization of replication origins on ARS plasmids in S. cerevisiae. Cell 1987, 51, 463–471. [Google Scholar] [CrossRef]
- Brewer, B.J.; Sena, E.P.; Fangman, W.L. Analysis of replication intermediates by two-dimensional agarose gel electrophoresis. Cancer Cells 1988, 6, 229–234. [Google Scholar]
- Viguera, E.; Hernandez, P.; Krimer, D.B.; Lurz, R.; Schvartzman, J.B. Visualisation of plasmid replication intermediates containing reversed forks. Nucleic Acids Res. 2000, 28, 498–503. [Google Scholar] [CrossRef]
- Fierro-Fernandez, M.; Hernandez, P.; Krimer, D.B.; Stasiak, A.; Schvartzman, J.B. Topological locking restrains replication fork reversal. Proc. Natl. Acad. Sci. USA 2007, 104, 1500–1505. [Google Scholar] [CrossRef] [PubMed]
- Stellwagen, N.C. Electrophoresis of DNA in agarose gels, polyacrylamide gels and in free solution. Electrophoresis 2009, 30 (Suppl. 1), S188–S195. [Google Scholar] [CrossRef]
- Neelsen, K.J.; Lopes, M. Replication fork reversal in eukaryotes: From dead end to dynamic response. Nat. Reviews. Mol. Cell Biol. 2015, 16, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Huberman, J.A.; Riggs, A.D. Autoradiography of chromosomal DNA fibers from Chinese hamster cells. Proc. Natl. Acad. Sci. USA 1966, 55, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Michalet, X.; Ekong, R.; Fougerousse, F.; Rousseaux, S.; Schurra, C.; Hornigold, N.; van Slegtenhorst, M.; Wolfe, J.; Povey, S.; Beckmann, J.S.; et al. Dynamic molecular combing: Stretching the whole human genome for high-resolution studies. Science 1997, 277, 1518–1523. [Google Scholar] [CrossRef]
- Sogo, J.M.; Stahl, H.; Koller, T.; Knippers, R. Structure of replicating Simian Virus 40 minichromosomes. J. Mol. Biol. 1986, 186, 189–204. [Google Scholar] [CrossRef]
- Sogo, J.M.; Stasiak, A.; Martínez-Robles, M.L.; Krimer, D.B.; Hernández, P.; Schvartzman, J.B. Formation of knots in partially replicated DNA molecules. J. Mol. Biol. 1999, 286, 637–643. [Google Scholar] [CrossRef]
- Weber, C.; Stasiak, A.; De Los Rios, P.; Dietler, G. Numerical simulation of gel electrophoresis of DNA knots in weak and strong electric fields. Biophys. J. 2006, 90, 3100–3105. [Google Scholar] [CrossRef]
- Charvin, G.; Bensimon, D.; Croquette, V. Single-molecule study of DNA unlinking by eukaryotic and prokaryotic type-II topoisomerases. Proc. Natl. Acad. Sci. USA 2003, 100, 9820–9825. [Google Scholar] [CrossRef]
- Witz, G.; Stasiak, A. DNA supercoiling and its role in DNA decatenation and unknotting. Nucleic Acids Res. 2010, 38, 2119–2133. [Google Scholar] [CrossRef] [PubMed]
- Rawdon, E.J.; Dorier, J.; Racko, D.; Millett, K.C.; Stasiak, A. How topoisomerase IV can efficiently unknot and decatenate negatively supercoiled DNA molecules without causing their torsional relaxation. Nucleic Acids Res. 2016, 44, 4528–4538. [Google Scholar] [CrossRef] [PubMed]
- O’Donnol, D.; Stasiak, A.; Buck, D. Two convergent pathways of DNA knotting in replicating DNA molecules as revealed by tetha-curve analysis. Nucleic Acids Res. 2018, 17, 9181–9188. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.; Schaerer, C.; Hernandez, P.; Krimer, D.B.; Schvartzman, J.B.; Fernandez-Nestosa, M.J. Distribution of torsional stress between the un-replicated and replicated regions in partially replicated molecules. J. Biomol. Struct. Dyn. 2020, 39, 2266–2277. [Google Scholar] [CrossRef] [PubMed]
- Gubaev, A.; Klostermeier, D. DNA-induced narrowing of the gyrase N-gate coordinates T-segment capture and strand passage. Proc. Natl. Acad. Sci. USA 2011, 108, 14085–14090. [Google Scholar] [CrossRef]
- Gubaev, A.; Weidlich, D.; Klostermeier, D. DNA gyrase with a single catalytic tyrosine can catalyze DNA supercoiling by a nicking-closing mechanism. Nucleic Acids Res. 2016, 44, 10354–10366. [Google Scholar] [CrossRef]
- Lee, I.; Dong, K.C.; Berger, J.M. The role of DNA bending in type IIA topoisomerase function. Nucleic Acids Res. 2013, 41, 5444–5456. [Google Scholar] [CrossRef]
- Wendorff, T.J.; Schmidt, B.H.; Heslop, P.; Austin, C.A.; Berger, J.M. The structure of DNA-bound human topoisomerase II alpha: Conformational mechanisms for coordinating inter-subunit interactions with DNA cleavage. J. Mol. Biol. 2012, 424, 109–124. [Google Scholar] [CrossRef]
- Olavarrieta, L.; Martinez-Robles, M.L.; Sogo, J.M.; Stasiak, A.; Hernandez, P.; Krimer, D.B.; Schvartzman, J.B. Supercoiling, knotting and replication fork reversal in partially replicated plasmids. Nucleic Acids Res. 2002, 30, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, M.; Kobayashi, T.; Horiuchi, T. A Newly Identified DNA Replication Terminus Site, TerE, on the Escherichia-Coli Chromosome. J. Bacteriol. 1991, 173, 391–393. [Google Scholar] [CrossRef]
- Gottlieb, P.A.; Wu, S.; Zhang, X.; Tecklenburg, M.; Kuempel, P.; Hill, T.M. Equilibrium, Kinetic, and Footprinting Studies of the Tus-Ter Protein-DNA Interaction. J. Biol. Chem. 1992, 267, 7434–7443. [Google Scholar] [CrossRef]
- Santamaría, D.; Hernández, P.; Martínez-Robles, M.L.; Krimer, D.B.; Schvartzman, J.B. Premature termination of DNA replication in plasmids carrying two inversely oriented ColE1 origins. J. Mol. Biol. 2000, 300, 75–82. [Google Scholar] [CrossRef]
- Cebrian, J.; Castan, A.; Martinez, V.; Kadomatsu-Hermosa, M.J.; Parra, C.; Fernandez-Nestosa, M.J.; Schaerer, C.; Hernandez, P.; Krimer, D.B.; Schvartzman, J.B. Direct Evidence for the Formation of Precatenanes during DNA Replication. J. Biol. Chem. 2015, 290, 13725–13735. [Google Scholar] [CrossRef]
- Lucas, I.; Germe, T.; Chevrier-Miller, M.; Hyrien, O. Topoisomerase II can unlink replicating DNA by precatenane removal. EMBO J. 2001, 20, 6509–6519. [Google Scholar] [CrossRef] [PubMed]
- Hayama, R.; Marians, K.J. Physical and functional interaction between the condensin MukB and the decatenase topoisomerase IV in Escherichia coli. Proc. Natl. Acad. Sci. USA 2010, 107, 18826–18831. [Google Scholar] [CrossRef]
- Hayama, R.; Bahng, S.; Karasu, M.E.; Marians, K.J. The MukB-ParC Interaction Affects the Intramolecular, Not Intermolecular, Activities of Topoisomerase IV. J. Biol. Chem. 2013, 288, 7653–7661. [Google Scholar] [CrossRef]
- Hill, T.M.; Pelletier, A.J.; Tecklenburg, M.L.; Kuempel, P.L. Identification of the DNA sequence from E. coli terminus region that halts replication forks. Cell 1988, 55, 459–466. [Google Scholar] [CrossRef]
- Santamaría, D.; delaCueva, G.; Martínez-Robles, M.L.; Krimer, D.B.; Hernández, P.; Schvartzman, J.B. DnaB helicase is unable to dissociate RNA-DNA hybrids—Its implication in the polar pausing of replication forks at ColE1 origins. J. Biol. Chem. 1998, 273, 33386–33396. [Google Scholar] [CrossRef]
- Harms, A.; Stanger, F.V.; Scheu, P.D.; de Jong, I.G.; Goepfert, A.; Glatter, T.; Gerdes, K.; Schirmer, T.; Dehio, C. Adenylylation of Gyrase and Topo IV by FicT Toxins Disrupts Bacterial DNA Topology. Cell Rep. 2015, 12, 1497–1507. [Google Scholar] [CrossRef]
- Sandhaus, S.; Chapagain, P.P.; Tse-Dinh, Y.C. Discovery of novel bacterial topoisomerase I inhibitors by use of in silico docking and in vitro assays. Sci. Rep. 2018, 8, 1437. [Google Scholar] [CrossRef]
- Stros, M.; Bacikova, A.; Polanska, E.; Stokrova, J.; Strauss, F. HMGB1 interacts with human topoisomerase IIalpha and stimulates its catalytic activity. Nucleic Acids Res. 2007, 35, 5001–5013. [Google Scholar] [CrossRef]
- Gibson, E.G.; Oviatt, A.A.; Osheroff, N. Two-Dimensional Gel Electrophoresis to Resolve DNA Topoisomers. Methods Mol. Biol. 2020, 2119, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Peter, B.J.; Ullsperger, C.; Hiasa, H.; Marians, K.J.; Cozzarelli, N.R. The structure of supercoiled intermediates in DNA replication. Cell 1998, 94, 819–827. [Google Scholar] [CrossRef]
- Martin-Parras, L.; Lucas, I.; Martinez-Robles, M.L.; Hernandez, P.; Krimer, D.B.; Hyrien, O.; Schvartzman, J.B. Topological complexity of different populations of pBR322 as visualized by two-dimensional agarose gel electrophoresis. Nucleic Acids Res. 1998, 26, 3424–3432. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Crisona, N.J.; Strick, T.R.; Bensimon, D.; Croquette, V.; Cozzarelli, N.R. Preferential relaxation of positively supercoiled DNA by E. coli topoisomerase IV in single-molecule and ensemble measurements. Genes Dev. 2000, 14, 2881–2892. [Google Scholar] [CrossRef] [PubMed]
- Schvartzman, J.B.; Martinez, V.; Hernandez, P.; Krimer, D.B.; Fernandez-Nestosa, M.J. Changes in the topology of DNA replication intermediates: Important discrepancies between in vitro and in vivo. BioEssays News Rev. Mol. Cell. Dev. Biol. 2021, 43, e2000309. [Google Scholar] [CrossRef]
- Bigot, S.; Marians, K.J. DNA chirality-dependent stimulation of topoisomerase IV activity by the C-terminal AAA+ domain of FtsK. Nucleic Acids Res. 2010, 38, 3031–3040. [Google Scholar] [CrossRef]
- Buck, G.R.; Zechiedrich, E.L. DNA disentangling by type-2 topoisomerases. J. Mol. Biol. 2004, 340, 933–939. [Google Scholar] [CrossRef]
- Charvin, G.; Strick, T.R.; Bensimon, D.; Croquette, V. Topoisomerase IV bends and overtwists DNA upon binding. Biophys. J. 2005, 89, 384–392. [Google Scholar] [CrossRef][Green Version]
- Neuman, K.C.; Charvin, G.; Bensimon, D.; Croquette, V. Mechanisms of chiral discrimination by topoisomerase IV. Proc. Natl. Acad. Sci. USA 2009, 106, 6986–6991. [Google Scholar] [CrossRef]
- Stone, M.D.; Bryant, Z.; Crisona, N.J.; Smith, S.B.; Vologodskii, A.; Bustamante, C.; Cozzarelli, N.R. Chirality sensing by Escherichia coli topoisomerase IV and the mechanism of type II topoisomerases. Proc. Natl. Acad. Sci. USA 2003, 100, 8654–8659. [Google Scholar] [CrossRef] [PubMed]
- Hiasa, H.; Digate, R.J.; Marians, K.J. Decatenating activity of escherichia-coli DNA gyrase and Topoisomerase-I and Topoisomerase-III during oriC and pBR322 DNA replication invitro. J. Biol. Chem. 1994, 269, 2093–2099. [Google Scholar] [CrossRef]
- Hiasa, H.; Marians, K.J. Two distinct modes of strand unlinking during theta-type DNA replication. J. Biol. Chem. 1996, 271, 21529–21535. [Google Scholar] [CrossRef] [PubMed]
- Kreuzer, K.N.; Cozzarelli, N.R. Formation and resolution of DNA catenanes by DNA gyrase. Cell 1980, 20, 245–254. [Google Scholar] [CrossRef]
- Levine, C.; Hiasa, H.; Marians, K.J. DNA gyrase and topoisomerase IV: Biochemical activities, physiological roles during chromosome replication, and drug sensitivities. Biochim. Biophys. Acta Gene Struct. Expr. 1998, 1400, 29–43. [Google Scholar] [CrossRef]
- Marians, K.J. DNA gyrase-catalyzed decatenation of multiply linked DNA dimers. J. Biol. Chem. 1987, 262, 10362–10368. [Google Scholar] [CrossRef]
- Ullsperger, C.; Cozzarelli, N.R. Contrasting enzymatic activities of topoisomerase IV and DNA gyrase from Escherichia coli. J. Biol. Chem. 1996, 271, 31549–31555. [Google Scholar] [CrossRef]
- Ashley, R.E.; Dittmore, A.; McPherson, S.A.; Turnbough, C.L., Jr.; Neuman, K.C.; Osheroff, N. Activities of gyrase and topoisomerase IV on positively supercoiled DNA. Nucleic Acids Res. 2017, 45, 9611–9624. [Google Scholar] [CrossRef]
- Nollmann, M.; Stone, M.D.; Bryant, Z.; Gore, J.; Crisona, N.J.; Hong, S.C.; Mitelheiser, S.; Maxwell, A.; Bustamante, C.; Cozzarelli, N.R. Multiple modes of Escherichia coli DNA gyrase activity revealed by force and torque. Nat. Struct. Mol. Biol. 2007, 14, 264–271. [Google Scholar] [CrossRef]
- Roca, J.; Wang, J.C. DNA transport by a type II DNA topoisomerase: Evidence in favor of a two-gate mechanism. Cell 1994, 77, 609–616. [Google Scholar] [CrossRef]
- Rybenkov, V.V.; Ullsperger, C.; Vologodskii, A.V.; Cozzarelli, N.R. Simplification of DNA topology below equilibrium values by type II topoisomerases. Science 1997, 277, 690–693. [Google Scholar] [CrossRef]
- Seol, Y.; Gentry, A.C.; Osheroff, N.; Neuman, K.C. Chiral discrimination and writhe-dependent relaxation mechanism of human topoisomerase IIalpha. J. Biol. Chem. 2013, 288, 13695–13703. [Google Scholar] [CrossRef] [PubMed]
- McClendon, A.K.; Rodriguez, A.C.; Osheroff, N. Human topoisomerase IIalpha rapidly relaxes positively supercoiled DNA: Implications for enzyme action ahead of replication forks. J. Biol. Chem. 2005, 280, 39337–39345. [Google Scholar] [CrossRef] [PubMed]
- McClendon, A.K.; Gentry, A.C.; Dickey, J.S.; Brinch, M.; Bendsen, S.; Andersen, A.H.; Osheroff, N. Bimodal recognition of DNA geometry by human topoisomerase II alpha: Preferential relaxation of positively supercoiled DNA requires elements in the C-terminal domain. Biochemistry 2008, 47, 13169–13178. [Google Scholar] [CrossRef]
- Kramlinger, V.M.; Hiasa, H. The “GyrA-box” is required for the ability of DNA gyrase to wrap DNA and catalyze the supercoiling reaction. J. Biol. Chem. 2006, 281, 3738–3742. [Google Scholar] [CrossRef]
- Han, J.; Wang, Y.; Sahin, O.; Shen, Z.; Guo, B.; Shen, J.; Zhang, Q. A fluoroquinolone resistance associated mutation in gyrA Affects DNA supercoiling in Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2012, 2, 21. [Google Scholar] [CrossRef]
- Kugelberg, E.; Lofmark, S.; Wretlind, B.; Andersson, D.I. Reduction of the fitness burden of quinolone resistance in Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2005, 55, 22–30. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cebrián, J.; Martínez, V.; Hernández, P.; Krimer, D.B.; Fernández-Nestosa, M.-J.; Schvartzman, J.B. Two-Dimensional Gel Electrophoresis to Study the Activity of Type IIA Topoisomerases on Plasmid Replication Intermediates. Biology 2021, 10, 1195. https://doi.org/10.3390/biology10111195
Cebrián J, Martínez V, Hernández P, Krimer DB, Fernández-Nestosa M-J, Schvartzman JB. Two-Dimensional Gel Electrophoresis to Study the Activity of Type IIA Topoisomerases on Plasmid Replication Intermediates. Biology. 2021; 10(11):1195. https://doi.org/10.3390/biology10111195
Chicago/Turabian StyleCebrián, Jorge, Victor Martínez, Pablo Hernández, Dora B. Krimer, María-José Fernández-Nestosa, and Jorge B. Schvartzman. 2021. "Two-Dimensional Gel Electrophoresis to Study the Activity of Type IIA Topoisomerases on Plasmid Replication Intermediates" Biology 10, no. 11: 1195. https://doi.org/10.3390/biology10111195
APA StyleCebrián, J., Martínez, V., Hernández, P., Krimer, D. B., Fernández-Nestosa, M.-J., & Schvartzman, J. B. (2021). Two-Dimensional Gel Electrophoresis to Study the Activity of Type IIA Topoisomerases on Plasmid Replication Intermediates. Biology, 10(11), 1195. https://doi.org/10.3390/biology10111195