
Borreliae Part 2: Borrelia Relapsing Fever Group and Unclassified Borrelia

 , ,
, , 
Simple Summary
Abstract
1. Introduction
2. Endemic Relapsing Fever Borreliosis (STBRF)
2.1. Epidemiology
2.1.1. Old World Strains

{kind=link}
| Borrelia | Ornithodoros Tick | Geographical Areas | Disease (Reservoirs) | References |
|---|---|---|---|---|
| B. armenica | O. verrucosus | Ukraine | Mouse infection (mouse, guinea pigs) | [34] |
| B. babylonensis | O. verrucosus | Russia, Ukraine | Mouse infection | [35] |
| B. baltazardii | O. tholozani | Iran | STBRF | [36,37] |
| B. caucasica | O. verrucosus | Ukraine, Caucasus, southeast Europe | STBRF | [34,38] |
| B. crocidurae | O. marocanus | Western and Northern Africa | STBRF mild symptoms | [39] |
| B duttonii | O. moubata | Central, Eastern, and Southern Africa | STBRF neurological signs, neonatal infection | [40,41] |
| B. graingeri | O.graingeri | Kenya | Flu-like syndrome | [42] |
| B. harveyi | unknown | Kenya, East Africa | Bacteremia of monkeys (Cercopithecus aethiops) | [43] |
| B. hispanica | O. erraticus | Iberian Peninsula and Northern Africa | STBRF Ocular and neurological symptoms | [32] |
| B. kalaharica | O. savignyi | Kalahari desert (Botswana and Namibia) | STBRF | [44] |
| B. latyschewii | O. tartakovsky | Iran, Middle East | STBRF | [45] |
| B. mazzottii | O. talaje | Mexico, Central America, and western USA | STBRF | [46] |
| B. merionesi | O. costalis and O. merionesi | Morocco and Atlantic coastal areas of the Sahara desert | Non-infectious to humans (rodent, monkeys) | [24,47] |
| B. microtti | O. erraticus | Iran, Afghanistan, Eastern Africa | STBRF | [48] |
| B. persica | O. tholozani | Central Asia, Middle East, Egypt, and India | STBRF, infectious for dogs and cats | [49,50] |
| B. tillae | O. zumpti | Southern Africa | Rodents | [51] |
Geographical Distribution
2.1.2. New World Strains
| Borrelia | Ornithodoros Tick | Geographical Areas | Hosts and Disease | Ref. |
|---|---|---|---|---|
| B. braziliensis | O. braziliensis | Plateau of southern Brazil | STBRF (dogs, armadillos) | [54] |
| B. coriaceae | O.coriaceus | Western North America, northwest California | Deer bacteremia, dogs, bovine epizootic abortion | [58] |
| B. dugesii | O. dugesii | Mexico | (Rodents, Neotoma micropus) | [59] |
| B. hermsii | O. hermsi | Western North, White Mountains (Arizona, US), British Columbia (Canada) | STBRF (rodents, squirrels) | [60] |
| B. mazzottii | O. talaje | Mexico, Central America, and western US | STBRF | [46] |
| B. parkeri | O. parkeri | Western US | STBRF | [61,62] |
| B. queenslandica | O. gurneyi | Australia | Bacteremia with relapse (Rodents, Rattus villosissimus, Kangaroo) | [63] |
| B turicatae | O. turicata | British Columbia (Canada), southwestern and south-central US and Mexico | STBRF | [64,65] |
| B. venezualensis | O. rudis | Central America and northern South America, Venezuela, Brazil, Colombia, Panama | STBRF | [66] |
| B. johnsonii | Carios kelleyi | Canada, US, China, Mexico, Cuba, Costa Rica, Chile | Bats, tick-borne illness to be defined | [56,67,68] |
Geographical Distribution
2.2. Microbiology
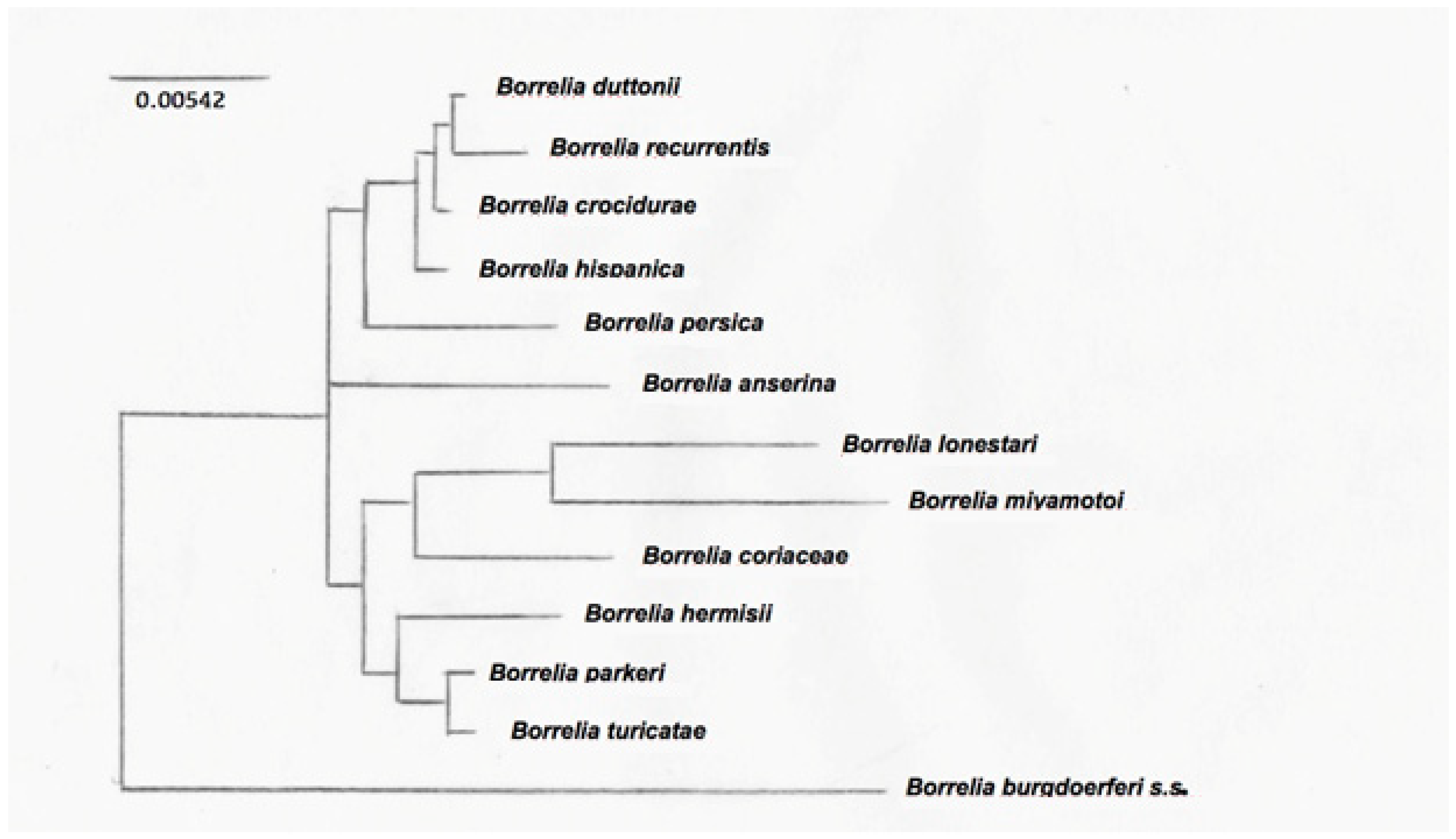
2.2.1. Phylogenetic Analysis
2.2.2. Genome
2.2.3. Species Identification of the STBRF
2.2.4. Antigens
2.2.5. Clinical Aspects
3. Hard-Tick-Borne Relapsing Fever (HTBRF)
3.1. Epidemiology
3.2. Hosts and Reservoirs
3.3. Clinical Aspects
4. Louse-Borne Relapsing Fever (LBRF)
4.1. Epidemiology
4.2. Transmission
4.3. Microbiology
4.4. Clinical Aspects
5. Avian Relapsing Fever Borreliae
6. Unclassified Borreliae
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cutler, S.J.; Abdissa, A.; Trape, J.F. New concepts for the old challenge of African relapsing fever borreliosis. Clin. Microbiol. Infect. 2009, 15, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, G.E.R.; Chadwick, J.; Mann, W.N. Hippocratic Writings; Harmondsworth: New York, NY, USA, 1983. [Google Scholar]
- Warrell, D.A. Louse-borne relapsing fever (Borrelia recurrentis infection). Epidemiol. Infect. 2019, 147, e106. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, W. Historical notes on some epidemic diseases associated with jaundice. Br. Med. Bull. 1957, 13, 146–149. [Google Scholar]
- Rutty, J. A Chronological History of the Weather and Seasons, and of the Prevailing Diseases in Dublin; Robinson and Roberts: London, UK, 1770. [Google Scholar]
- Raoult, D.; Dutour, O.; Houhamdi, L.; Jankauskas, R.; Fournier, P.E.; Ardagna, Y.; Drancourt, M.; Signoli, M.; La, V.D.; Macia, Y.; et al. Evidence for louse-transmitted diseases in soldiers of Napoleon’s Grand Army in Vilnius. J. Infect. Dis. 2006, 193, 112–120. [Google Scholar] [CrossRef]
- Craige, D. Notice of a febrile disorder which has prevailed at Edinburgh during the Summer of 1843. Edinb. Med. Surg. J. 1843, 60, 410–418. [Google Scholar]
- Southern, P.M.; Sanford, J.P. Relapsing fever: A clinical and microbiological review. Medicine 1969, 48, 129–149. [Google Scholar] [CrossRef]
- Wright, D.J.; Maria, B. Ich bin ein Berliner*. Clin. Microbiol. Infect. 2011, 17, 484–486. [Google Scholar] [CrossRef]
- Dschunkowsky, D. Das Rückfallfieber in Persien. Dtsch. Med. Wochenschr. 1913, 39, 419–420. [Google Scholar] [CrossRef][Green Version]
- Weldon, E.D. World Distribution of Spirochetal Diseases. 2, Relapsing Fevers, Louse-Borne and Tick-Borne; American Geographical Society of New York: New York, NY, USA, 1955. [Google Scholar]
- Elbir, H.; Gimenez, G.; Sokhna, C.; Bilcha, K.D.; Ali, J.; Barker, S.C.; Cutler, S.J.; Raoult, D.; Drancourt, M. Multispacer sequence typing relapsing fever Borreliae in Africa. PLoS Negl. Trop. Dis. 2012, 6, e1652. [Google Scholar] [CrossRef]
- Dworkin, M.S.; Anderson, D.E., Jr.; Schwan, T.G.; Shoemaker, P.C.; Banerjee, S.N.; Kassen, B.O.; Burgdorfer, W. Tick-borne relapsing fever in the northwestern United States and southwestern Canada. Clin. Infect. Dis. 1998, 26, 122–131. [Google Scholar] [CrossRef]
- Elbir, H.; Raoult, D.; Drancourt, M. Relapsing fever borreliae in Africa. Am. J. Trop. Med. Hyg. 2013, 89, 288–292. [Google Scholar] [CrossRef]
- Margos, G.; Vollmer, S.A.; Ogden, N.H.; Fish, D. Population genetics, taxonomy, phylogeny and evolution of Borrelia burgdorferi sensu lato. Infect. Genet. Evol. 2011, 11, 1545–1563. [Google Scholar] [CrossRef]
- Schotthoefer, A.M.; Frost, H.M. Ecology and Epidemiology of Lyme Borreliosis. Clin. Lab. Med. 2015, 35, 723–743. [Google Scholar] [CrossRef]
- Pritt, B.S.; Respicio-Kingry, L.B.; Sloan, L.M.; Schriefer, M.E.; Replogle, A.J.; Bjork, J.; Liu, G.; Kingry, L.C.; Mead, P.S.; Neitzel, D.F.; et al. Borrelia mayonii sp. nov., a member of the Borrelia burgdorferi sensu lato complex, detected in patients and ticks in the upper midwestern United States. Int. J. Syst. Evol. Microbiol. 2016, 66, 4878–4880. [Google Scholar] [CrossRef]
- Talagrand-Reboul, E.; Boyer, P.H.; Bergstrom, S.; Vial, L.; Boulanger, N. Relapsing Fevers: Neglected Tick-Borne Diseases. Front. Cell Infect. Microbiol. 2018, 8, 98. [Google Scholar] [CrossRef]
- Lopez, J.E.; Krishnavahjala, A.; Garcia, M.N.; Bermudez, S. Tick-Borne relapsing fever spirochetes in the Americas. Vet. Sci. 2016, 3, 16. [Google Scholar] [CrossRef]
- Felsenfeld, O. Chapter IV Borrelia. In Methods in Microbiology; Norris, J.R., Ribbons, D.W., Eds.; Academic Press: Cambridge, MA, USA, 1973; Volume 8, pp. 75–94. [Google Scholar]
- Ribeiro, J.C.; Endris, T.M.; Endris, R. Saliva of the soft tick, ornithodoros moubata, contains anti-platelet and apyrase activities. Comp. Biochem. Physiol.—Part A Physiol. 1991, 100, 109–112. [Google Scholar] [CrossRef]
- Oleaga, A.; Escudero-Poblacion, A.; Camafeita, E.; Perez-Sanchez, R. A proteomic approach to the identification of salivary proteins from the argasid ticks Ornithodoros moubata and Ornithodoros erraticus. Insect. Biochem. Mol. Biol. 2007, 37, 1149–1159. [Google Scholar] [CrossRef]
- Francischetti, I.M.; Sa-Nunes, A.; Mans, B.J.; Santos, I.M.; Ribeiro, J.M. The role of saliva in tick feeding. Front. Biosci. (Landmark Ed. India) 2009, 14, 2051–2088. [Google Scholar] [CrossRef]
- Trape, J.F.; Diatta, G.; Arnathau, C.; Bitam, I.; Sarih, M.; Belghyti, D.; Bouattour, A.; Elguero, E.; Vial, L.; Mane, Y.; et al. The epidemiology and geographic distribution of relapsing fever borreliosis in West and North Africa, with a review of the Ornithodoros erraticus complex (Acari: Ixodida). PLoS ONE 2013, 8, e78473. [Google Scholar] [CrossRef]
- Rebaudet, S.; Parola, P. Epidemiology of relapsing fever borreliosis in Europe. FEMS Immunol. Med. Microbiol. 2006, 48, 11–15. [Google Scholar] [CrossRef]
- Assous, M.V.; Wilamowski, A. Relapsing fever borreliosis in Eurasia--forgotten, but certainly not gone! Clin. Microbiol. Infect. 2009, 15, 407–414. [Google Scholar] [CrossRef]
- Safdie, G.; Farrah, I.Y.; Yahia, R.; Marva, E.; Wilamowski, A.; Sawalha, S.S.; Wald, N.; Schmiedel, J.; Moter, A.; Gobel, U.B.; et al. Molecular characterization of Borrelia persica, the agent of tick borne relapsing fever in Israel and the Palestinian Authority. PLoS ONE 2010, 5, e14105. [Google Scholar] [CrossRef]
- Palma, M.; Lopes de Carvalho, I.; Osorio, H.; Ze-Ze, L.; Cutler, S.J.; Nuncio, M.S. Portuguese hosts for Ornithodoros erraticus ticks. Vector Borne Zoonotic Dis. 2013, 13, 775–777. [Google Scholar] [CrossRef]
- Palma, M.; Lopes de Carvalho, I.; Figueiredo, M.; Amaro, F.; Boinas, F.; Cutler, S.J.; Nuncio, M.S. Borrelia hispanica in Ornithodoros erraticus, Portugal. Clin. Mictobiol. Infect. 2012, 18, 696–701. [Google Scholar] [CrossRef]
- Pineda Cantero, A.; Perez de Pedro, I.; Martin Tellez, S.; Costo Muriel, C.; Caballero Martinez, L.F.; Gomez Huelgas, R. Borrelia hispanica as a cause of recurrent fever. Med. Clin. (Barc.) 2020, 154, 380. [Google Scholar] [CrossRef]
- Wyplosz, B.; Mihaila-Amrouche, L.; Baixench, M.T.; Bigel, M.L.; Berardi-Grassias, L.; Fontaine, C.; Hornstein, M.; Izri, A.; Baranton, G.; Postic, D. Imported tickborne relapsing fever, France. Emerg. Infect. Dis. 2005, 11, 1801–1803. [Google Scholar] [CrossRef]
- Heida, J.; Van Arkel, A.; Verweij, J.J.; Tijssen, C.C. Meningitis due to infection with Borrelia hispanica. Ned. Tijdschr. Voor Geneeskd. 2019, 163, D3859. [Google Scholar]
- Anda, P.; Sanchez-Yebra, W.; del Mar Vitutia, M.; Perez Pastrana, E.; Rodriguez, I.; Miller, N.S.; Backenson, P.B.; Benach, J.L. A new Borrelia species isolated from patients with relapsing fever in Spain. Lancet 1996, 348, 162–165. [Google Scholar] [CrossRef]
- Filatov, S.; Krishnavajhala, A.; Armstrong, B.A.; Kneubehl, A.R.; Nieto, N.C.; Perez De Leon, A.A.; Lopez, J.E. Isolation and Molecular Characterization of Tick-Borne Relapsing Fever Borrelia Infecting Ornithodoros (Pavlovskyella) verrucosus Ticks Collected in Ukraine. J. Infect. Dis. 2020, 221, 804–811. [Google Scholar] [CrossRef]
- Goubau, P.F. Relapsing fevers. A review. Ann. Soc. Belg. Med. Trop. 1984, 64, 335–364. [Google Scholar] [PubMed]
- Moradi-Asl, E.; Jafari, S. The habitat suitability model for the potential distribution of Ornithodoros tholozani (Laboulbene et Megnin, 1882) and Ornithodoros lahorensis (Neumann, 1908) (Acari: Argasidae): The main vectors of tick-borne relapsing fever in Iran. Ann. Parasitol. 2020, 66, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Karimi, Y.; Hovind-Hougen, K.; Birch-Andersen, A.; Asmar, M. Borrelia persica and B. baltazardi sp. nov.: Experimental pathogenicity for some animals and comparison of the ultrastructure. Annales de Microbiologie 1979, 130 B, 157-168. Ann. Microbiol. 1979, 130 B, 157–168. [Google Scholar]
- Kandelaki, S.P. On the Relapsing fever transmitted by ticks in Transcaucasia. Med. Parasitol. 1935, 4, 65–66. [Google Scholar]
- Souidi, Y.; Boudebouch, N.; Ezikouri, S.; Belghyti, D.; Trape, J.F.; Sarih, M. Borrelia crocidurae in Ornithodoros ticks from northwestern Morocco: A range extension in relation to climatic change? J. Vector. Ecol. 2014, 39, 316–320. [Google Scholar] [CrossRef]
- van Holten, J.; Tiems, J.; Jongen, V.H. Neonatal Borrelia duttoni infection: A report of three cases. Trop. Doct. 1997, 27, 115–116. [Google Scholar] [CrossRef]
- Obolo-Mvoulouga, P.; Oleaga, A.; Manzano-Roman, R.; Perez-Sanchez, R. Evaluation of the protective efficacy of Ornithodoros moubata midgut membrane antigens selected using omics and in silico prediction algorithms. Ticks Tick Borne Dis. 2018, 9, 1158–1172. [Google Scholar] [CrossRef]
- Heisch, R.B. On a spirochaete isolated from ornithodoros graingeri. Parasitology 1953, 43, 133–135. [Google Scholar] [CrossRef]
- Garnham, P.C. A new blood spirochaete in the grivet monkey; Ceropithecus aethiops. East Afr. Med. J. 1947, 24, 47–51. [Google Scholar]
- Cutler, S.J.; Idris, J.M.; Ahmed, A.O.; Elelu, N. Ornithodoros savignyi, the Tick Vector of “Candidatus Borrelia kalaharica” in Nigeria. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef]
- Baltazard, M.; Pournaki, R.; Bahmanyar, M.; Chamsa, M. Ornithodorus tartakovskyi Olenev 1931 and Borrelia (Spirochaeta) latychevii Sofiev 1941; supplementary note. Ann. Parasitol. Hum. Comp. 1955, 30, 225–242. [Google Scholar] [CrossRef]
- Davis, G.E. A relapsing fever spirochete, Borrelia mazzottii (sp. nov.) from Ornithodoros talaje from Mexico. Am. J. Hyg. 1956, 63, 13–17. [Google Scholar] [CrossRef]
- Diatta, G.; Souidi, Y.; Granjon, L.; Arnathau, C.; Durand, P.; Chauvancy, G.; Mane, Y.; Sarih, M.; Belghyti, D.; Renaud, F.; et al. Epidemiology of tick-borne borreliosis in Morocco. PLoS Negl. Trop. Dis. 2012, 6, e1810. [Google Scholar] [CrossRef]
- Assmar, M.; Soleimani, M.; Oreiz, F.; Piazak, N.; Hossini, S.M.; Saghiri, R.; Zamani, Z. Purification of periplasmic flagellar antigen from Borrelia microtti. Scand. J. Infect. Dis. 2002, 34, 267–272. [Google Scholar] [CrossRef]
- Colin de Verdiere, N.; Hamane, S.; Assous, M.V.; Sertour, N.; Ferquel, E.; Cornet, M. Tickborne relapsing fever caused by Borrelia persica, Uzbekistan and Tajikistan. Emerg. Infect. Dis. 2011, 17, 1325–1327. [Google Scholar] [CrossRef]
- Baneth, G.; Nachum-Biala, Y.; Halperin, T.; Hershko, Y.; Kleinerman, G.; Anug, Y.; Abdeen, Z.; Lavy, E.; Aroch, I.; Straubinger, R.K. Borrelia persica infection in dogs and cats: Clinical manifestations, clinicopathological findings and genetic characterization. Parasites Vectors 2016, 9, 244. [Google Scholar] [CrossRef]
- Geigy, R.; Aeschlimann, A. Comparative Study of the Biology of Borrelia duttoni and Borrelia tillae. Rev. Suisse Zool. 1965, 72, 87–98. [Google Scholar] [CrossRef]
- Fingerle, V.; Pritsch, M.; Wachtler, M.; Margos, G.; Ruske, S.; Jung, J.; Loscher, T.; Wendtner, C.; Wieser, A. “Candidatus Borrelia kalaharica” Detected from a Febrile Traveller Returning to Germany from Vacation in Southern Africa. PLoS Negl. Trop. Dis. 2016, 10, e0004559. [Google Scholar] [CrossRef]
- Morel, P.-C. Les Tiques d’Afrique et du Bassin Méditerranéen; CIRAD-EMVT: Montpellier, France, 2003. [Google Scholar]
- Reck, J.; Marks, F.S.; Guimaraes, J.A.; Termignoni, C.; Martins, J.R. Epidemiology of Ornithodoros brasiliensis (mouro tick) in the southern Brazilian highlands and the description of human and animal retrospective cases of tick parasitism. Ticks Tick Borne Dis. 2013, 4, 101–109. [Google Scholar] [CrossRef]
- Gill, J.S.; Ullmann, A.J.; Loftis, A.D.; Schwan, T.G.; Raffel, S.J.; Schrumpf, M.E.; Piesman, J. Novel relapsing fever spirochete in bat tick. Emerg. Infect. Dis. 2008, 14, 522–523. [Google Scholar] [CrossRef]
- Kingry, L.C.; Anacker, M.; Pritt, B.; Bjork, J.; Respicio-Kingry, L.; Liu, G.; Sheldon, S.; Boxrud, D.; Strain, A.; Oatman, S.; et al. Surveillance for and Discovery of Borrelia Species in US Patients Suspected of Tickborne Illness. Clin. Infect. Dis. 2018, 66, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Schwan, T.G.; Raffel, S.J.; Schrumpf, M.E.; Gill, J.S.; Piesman, J. Characterization of a novel relapsing fever spirochete in the midgut, coxal fluid, and salivary glands of the bat tick Carios kelleyi. Vector Borne Zoonotic Dis. 2009, 9, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.C.; Burgdorfer, W.; Lane, R.S. Borrelia coriaceae sp. nov.: Putative agent of epizootic bovine abortion. Int. J. Syst. Bacteriol. 1987, 37, 72–74. [Google Scholar] [CrossRef]
- Guzman-Cornejo, C.; Herrera-Mares, A.; Robbins, R.G.; Rebollo-Hernandez, A. The soft ticks (Parasitiformes: Ixodida: Argasidae) of Mexico: Species, hosts, and geographical distribution. Zootaxa 2019, 4623, 485–525. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.; Artsob, H.; Margos, G.; Tsao, J.I. Other Tick-borne Bacterial Diseases (including Lyme disease, relapsing feverand Tularemia). In Biology of Ticks; Sonenshine, D., Roe, M.R., Eds.; Oxford University Press: New York, NY, USA, 2014; Volume 2, pp. 278–312. [Google Scholar]
- Davis, G.E. Species unity or plurality of the relapsing fever spirochetes. In A Symposium on Relapsing Fever in Americas; Moulton, F.R., Ed.; American Association for the Advancement of Science: Washington, DC, USA, 1942; pp. 41–47. [Google Scholar]
- Gage, K.L.; Eggleston, M.E.; Gilmore, R.D., Jr.; Dolan, M.C.; Montenieri, J.A.; Tanda, D.T.; Piesman, J. Isolation and characterization of Borrelia parkeri in Ornithodoros parkeri (Ixodida: Argasidae) collected in Colorado. J. Med. Entomol. 2001, 38, 665–674. [Google Scholar] [CrossRef]
- Hussain-Yusuf, H.; Stenos, J.; Vincent, G.; Shima, A.; Abell, S.; Preece, N.D.; Tadepalli, M.; Hii, S.F.; Bowie, N.; Mitram, K.; et al. Screening for Rickettsia, Coxiella and Borrelia Species in Ticks from Queensland, Australia. Pathogens 2020, 9, 1016. [Google Scholar] [CrossRef]
- Donaldson, T.G.; Perez de Leon, A.A.; Li, A.Y.; Castro-Arellano, I.; Wozniak, E.; Boyle, W.K.; Hargrove, R.; Wilder, H.K.; Kim, H.J.; Teel, P.D.; et al. Assessment of the Geographic Distribution of Ornithodoros turicata (Argasidae): Climate Variation and Host Diversity. PLoS Negl. Trop. Dis. 2016, 10, e0004383. [Google Scholar] [CrossRef]
- Krishnavajhala, A.; Armstrong, B.A.; Lopez, J.E. Vector Competence of Geographical Populations of Ornithodoros turicata for the Tick-Borne Relapsing Fever Spirochete Borrelia turicatae. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef]
- Munoz-Leal, S.; Faccini-Martinez, A.A.; Costa, F.B.; Marcili, A.; Mesquita, E.; Marques, E.P., Jr.; Labruna, M.B. Isolation and molecular characterization of a relapsing fever Borrelia recovered from Ornithodoros rudis in Brazil. Ticks Tick Borne Dis 2018, 9, 864–871. [Google Scholar] [CrossRef]
- Nadolny, R.M.; Kennedy, A.C.; Rodgers, J.M.; Vincent, Z.T.; Cornman, H.; Haynes, S.A.; Casal, C.; Robbins, R.G.; Richards, A.L.; Jiang, J.; et al. Carios kelleyi (Acari: Ixodida: Argasidae) Infected With Rickettsial Agents Documented Infesting Housing in Kansas, United States. J. Med. Entomol. 2021, tjab069. [Google Scholar] [CrossRef]
- Li, Z.M.; Xiao, X.; Zhou, C.M.; Liu, J.X.; Gu, X.L.; Fang, L.Z.; Liu, B.Y.; Wang, L.R.; Yu, X.J.; Han, H.J. Human-pathogenic relapsing fever Borrelia found in bats from Central China phylogenetically clustered together with relapsing fever borreliae reported in the New World. PLoS Negl. Trop. Dis. 2021, 15, e0009113. [Google Scholar] [CrossRef]
- Fritz, C.L.; Payne, J.R.; Schwan, T.G. Serologic evidence for Borrelia hermsii infection in rodents on federally owned recreational areas in California. Vector Borne Zoonotic Dis. 2013, 13, 376–381. [Google Scholar] [CrossRef]
- Brumpt, E.; Mazzotti, L.; Brumpt, L.C. Étude épidémiologique de la fièvre récurrente endémique des hauts plateaux mexicains. Ann. Parasitol. Hum. Comp. 1939, 17, 275–286. [Google Scholar] [CrossRef]
- Colunga-Salas, P.; Sanchez-Montes, S.; Volkow, P.; Ruiz-Remigio, A.; Becker, I. Lyme disease and relapsing fever in Mexico: An overview of human and wildlife infections. PLoS ONE 2020, 15, e0238496. [Google Scholar] [CrossRef]
- Munoz-Leal, S.; Marcili, A.; Fuentes-Castillo, D.; Ayala, M.; Labruna, M.B. A relapsing fever Borrelia and spotted fever Rickettsia in ticks from an Andean valley, central Chile. Exp. Appl. Acarol. 2019, 78, 403–420. [Google Scholar] [CrossRef]
- van Dam, A.P.; van Gool, T.; Wetsteyn, J.C.; Dankert, J. Tick-borne relapsing fever imported from West Africa: Diagnosis by quantitative buffy coat analysis and in vitro culture of Borrelia crocidurae. J. Clin. Microbiol. 1999, 37, 2027–2030. [Google Scholar] [CrossRef]
- Adeolu, M.; Gupta, R.S. A phylogenomic and molecular marker based proposal for the division of the genus Borrelia into two genera: The emended genus Borrelia containing only the members of the relapsing fever Borrelia, and the genus Borreliella gen. nov. containing the members of the Lyme disease Borrelia (Borrelia burgdorferi sensu lato complex). Antonie Van Leeuwenhoek 2014, 105, 1049–1072. [Google Scholar] [CrossRef]
- Ras, N.M.; Lascola, B.; Postic, D.; Cutler, S.J.; Rodhain, F.; Baranton, G.; Raoult, D. Phylogenesis of relapsing fever Borrelia spp. Int. J. Syst. Bacteriol. 1996, 46, 859–865. [Google Scholar] [CrossRef]
- Wilder, H.K.; Raffel, S.J.; Barbour, A.G.; Porcella, S.F.; Sturdevant, D.E.; Vaisvil, B.; Kapatral, V.; Schmitt, D.P.; Schwan, T.G.; Lopez, J.E. Transcriptional Profiling the 150 kb Linear Megaplasmid of Borrelia turicatae Suggests a Role in Vector Colonization and Initiating Mammalian Infection. PLoS ONE 2016, 11, e0147707. [Google Scholar] [CrossRef]
- Boyle, W.K.; Wilder, H.K.; Lawrence, A.M.; Lopez, J.E. Transmission dynamics of Borrelia turicatae from the arthropod vector. PLoS Negl. Trop. Dis. 2014, 8, e2767. [Google Scholar] [CrossRef]
- Krishnavajhala, A.; Armstrong, B.A.; Lopez, J.E. The impact of in vitro cultivation on the natural life cycle of the tick-borne relapsing fever spirochete Borrelia turicatae. PLoS ONE 2020, 15, e0239089. [Google Scholar] [CrossRef]
- Barbour, A.G. Antigenic variation of a relapsing fever Borrelia species. Ann. Rev. Microbiol. 1990, 44, 155–171. [Google Scholar] [CrossRef]
- Scott, J.C. Typing African relapsing fever spirochetes. Emerg. Infect. Dis. 2005, 11, 1722–1729. [Google Scholar] [CrossRef]
- Mitani, H.; Talbert, A.; Fukunaga, M. New World relapsing fever Borrelia found in Ornithodoros porcinus ticks in central Tanzania. Microbiol. Immunol. 2004, 48, 501–505. [Google Scholar] [CrossRef]
- Cutler, S.J.; Scott, J.C.; Wright, D.J.M. Phylogenetic origins of Borrelia recurrentis. Int. J. Med. Microbiol. 2008, 298, 193–202. [Google Scholar] [CrossRef]
- Miller, S.C.; Porcella, S.F.; Raffel, S.J.; Schwan, T.G.; Barbour, A.G. Large linear plasmids of Borrelia species that cause relapsing fever. J. Bacteriol. 2013, 195, 3629–3639. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.M.; Casjens, S.; Huang, W.M.; Sutton, G.G.; Clayton, R.; Lathigra, R.; White, O.; Ketchum, K.A.; Dodson, R.; Hickey, E.K.; et al. Genomic sequence of a Lyme disease spirochaete, Borrelia burgdorferi. Nature 1997, 390, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Tilly, K.; Elias, A.F.; Errett, J.; Fischer, E.; Iyer, R.; Schwartz, I.; Bono, J.L.; Rosa, P. Genetics and regulation of chitobiose utilization in Borrelia burgdorferi. J. Bacteriol. 2001, 183, 5544–5553. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dobrindt, U.; Hacker, J. Whole genome plasticity in pathogenic bacteria. Curr. Opin. Microbiol. 2001, 4, 550–557. [Google Scholar] [CrossRef]
- Halperin, T.; Orr, N.; Cohen, R.; Hasin, T.; Davidovitch, N.; Klement, E.; Kayouf, R.; Baneth, G.; Cohen, D.; Yavzori, M. Detection of relapsing fever in human blood samples from Israel using PCR targeting the glycerophosphodiester phosphodiesterase (GlpQ) gene. Acta Trop. 2006, 98, 189–195. [Google Scholar] [CrossRef]
- Oshaghi, M.A.; Rafinejad, J.; Choubdar, N.; Piazak, N.; Vatandoost, H.; Telmadarraiy, Z.; Mohtarami, F.; Ravasan, N.M. Discrimination of relapsing fever Borrelia persica and Borrelia microtti by diagnostic species-specific primers and polymerase chain reaction-restriction fragment length polymorphism. Vector Borne Zoonotic Dis. 2011, 11, 201–207. [Google Scholar] [CrossRef]
- Toledo, A.; Anda, P.; Escudero, R.; Larsson, C.; Bergstrom, S.; Benach, J.L. Phylogenetic analysis of a virulent Borrelia species isolated from patients with relapsing fever. J. Clin. Microbiol. 2010, 48, 2484–2489. [Google Scholar] [CrossRef]
- Lopez, J.E.; Wilder, H.K.; Hargrove, R.; Brooks, C.P.; Peterson, K.E.; Beare, P.A.; Sturdevant, D.E.; Nagarajan, V.; Raffel, S.J.; Schwan, T.G. Development of genetic system to inactivate a Borrelia turicatae surface protein selectively produced within the salivary glands of the arthropod vector. PLoS Negl. Trop. Dis. 2013, 7, e2514. [Google Scholar] [CrossRef]
- Trevisan, G.; Cinco, M.; Trevisini, S.; di Meo, N.; Chersi, K.; Ruscio, M.; Forgione, P.; Bonin, S. Borreliae Part 1: Borrelia Lyme Group and Echidna-Reptile Group. Biology 2021, 10, 1036. [Google Scholar] [CrossRef]
- Trevisan, G.; Bonin, S.; Ruscio, M. A Practical Approach to the Diagnosis of Lyme Borreliosis: From Clinical Heterogeneity to Laboratory Methods. Front. Med. (Lausanne) 2020, 7, 265. [Google Scholar] [CrossRef]
- Montgomery, R.R.; Lusitani, D.; De Boisfleury Chevance, A.; Malawista, S.E. Tick saliva reduces adherence and area of human neutrophils. Infect. Immun. 2004, 72, 2989–2994. [Google Scholar] [CrossRef]
- Nordstrand, A.; Shamaei-Tousi, A.; Ny, A.; Bergstrom, S. Delayed invasion of the kidney and brain by Borrelia crocidurae in plasminogen-deficient mice. Infect. Immun. 2001, 69, 5832–5839. [Google Scholar] [CrossRef]
- Gebbia, J.A.; Coleman, J.L.; Benach, J.L. Borrelia spirochetes upregulate release and activation of matrix metalloproteinase gelatinase B (MMP-9) and collagenase 1 (MMP-1) in human cells. Infect. Immun. 2001, 69, 456–462. [Google Scholar] [CrossRef]
- Burman, N.; Shamaei-Tousi, A.; Bergstrom, S. The spirochete Borrelia crocidurae causes erythrocyte rosetting during relapsing fever. Infect. Immun. 1998, 66, 815–819. [Google Scholar] [CrossRef]
- Pettersson, J.; Schrumpf, M.E.; Raffel, S.J.; Porcella, S.F.; Guyard, C.; Lawrence, K.; Gherardini, F.C.; Schwan, T.G. Purine salvage pathways among Borrelia species. Infect. Immun. 2007, 75, 3877–3884. [Google Scholar] [CrossRef]
- Barbour, A.G.; Tessier, S.L.; Stoenner, H.G. Variable major proteins of Borrellia hermsii. J. Exp. Med. 1982, 156, 1312–1324. [Google Scholar] [CrossRef]
- Alugupalli, K.R.; Gerstein, R.M.; Chen, J.; Szomolanyi-Tsuda, E.; Woodland, R.T.; Leong, J.M. The resolution of relapsing fever borreliosis requires IgM and is concurrent with expansion of B1b lymphocytes. J. Immunol. 2003, 170, 3819–3827. [Google Scholar] [CrossRef]
- Hinnebusch, B.J.; Barbour, A.G.; Restrepo, B.I.; Schwan, T.G. Population structure of the relapsing fever spirochete Borrelia hermsii as indicated by polymorphism of two multigene families that encode immunogenic outer surface lipoproteins. Infect. Immun. 1998, 66, 432–440. [Google Scholar] [CrossRef]
- Raffel, S.J.; Battisti, J.M.; Fischer, R.J.; Schwan, T.G. Inactivation of genes for antigenic variation in the relapsing fever spirochete Borrelia hermsii reduces infectivity in mice and transmission by ticks. PLoS Pathog. 2014, 10, e1004056. [Google Scholar] [CrossRef] [PubMed]
- Dai, Q.; Restrepo, B.I.; Porcella, S.F.; Raffel, S.J.; Schwan, T.G.; Barbour, A.G. Antigenic variation by Borrelia hermsii occurs through recombination between extragenic repetitive elements on linear plasmids. Mol. Microbiol. 2006, 60, 1329–1343. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Audic, S.; Robert, C.; Nguyen, T.T.; Blanc, G.; Cutler, S.J.; Wincker, P.; Couloux, A.; Claverie, J.M.; Raoult, D.; et al. The genome of Borrelia recurrentis, the agent of deadly louse-borne relapsing fever, is a degraded subset of tick-borne Borrelia duttonii. PLoS Genet. 2008, 4, e1000185. [Google Scholar] [CrossRef] [PubMed]
- Ras, N.M.; Postic, D.; Ave, P.; Huerre, M.; Baranton, G. Antigenic variation of Borrelia turicatae Vsp surface lipoproteins occurs in vitro and generates novel serotypes. Res. Microbiol. 2000, 151, 5–12. [Google Scholar] [CrossRef]
- Zuckert, W.R.; Kerentseva, T.A.; Lawson, C.L.; Barbour, A.G. Structural conservation of neurotropism-associated VspA within the variable Borrelia Vsp-OspC lipoprotein family. J. Biol. Chem. 2001, 276, 457–463. [Google Scholar] [CrossRef]
- Crowder, C.D.; Ghalyanchi Langeroudi, A.; Shojaee Estabragh, A.; Lewis, E.R.G.; Marcsisin, R.A.; Barbour, A.G. Pathogen and Host Response Dynamics in a Mouse Model of Borrelia hermsii Relapsing Fever. Vet. Sci. 2016, 3, 19. [Google Scholar] [CrossRef]
- Cutler, S.J. Relapsing Fever Borreliae: A Global Review. Clin. Lab. Med. 2015, 35, 847–865. [Google Scholar] [CrossRef]
- Cadavid, D.; Barbour, A.G. Neuroborreliosis during relapsing fever: Review of the clinical manifestations, pathology, and treatment of infections in humans and experimental animals. Clin. Infect. Dis. 1998, 26, 151–164. [Google Scholar] [CrossRef]
- Oza, S.; Lawn, J.E.; Hogan, D.R.; Mathers, C.; Cousens, S.N. Neonatal cause-of-death estimates for the early and late neonatal periods for 194 countries: 2000-2013. Bull. World Health Organ. 2015, 93, 19–28. [Google Scholar] [CrossRef]
- Larsson, C.; Andersson, M.; Guo, B.P.; Nordstrand, A.; Hagerstrand, I.; Carlsson, S.; Bergstrom, S. Complications of pregnancy and transplacental transmission of relapsing-fever borreliosis. J. Infect. Dis. 2006, 194, 1367–1374. [Google Scholar] [CrossRef]
- UNICEF. Levels and Trends in Child Mortalit 2017; United Nations Interagency Group for Child Mortality: New York, NY, USA, 2017. [Google Scholar]
- Goubau, P.F.; Munyangeyo, C. Tick-borne relapsing fever and pregnancy. A clinical study in Rwanda. Ann. Soc. Belg. Med. Trop. 1983, 63, 347–355. [Google Scholar]
- Piccione, J.; Levine, G.J.; Duff, C.A.; Kuhlman, G.M.; Scott, K.D.; Esteve-Gassent, M.D. Tick-Borne Relapsing Fever in Dogs. J. Vet. Intern. Med. 2016, 30, 1222–1228. [Google Scholar] [CrossRef]
- Elelu, N. Tick-borne relapsing fever as a potential veterinary medical problem. Vet. Med. Sci. 2018, 4, 271–279. [Google Scholar] [CrossRef]
- Theiler, A. Spirillosis of cattle. J. Comp. Pathol. Ther. 1904, 17, 47–55. [Google Scholar] [CrossRef]
- Barbour, A.G.; Maupin, G.O.; Teltow, G.J.; Carter, C.J.; Piesman, J. Identification of an uncultivable Borrelia species in the hard tick Amblyomma americanum: Possible agent of a Lyme disease-like illness. J. Infect. Dis. 1996, 173, 403–409. [Google Scholar] [CrossRef]
- Hue, F.; Ghalyanchi Langeroudi, A.; Barbour, A.G. Chromosome Sequence of Borrelia miyamotoi, an Uncultivable Tick-Borne Agent of Human Infection. Genome Announc. 2013, 1. [Google Scholar] [CrossRef]
- Barbour, A.G. Phylogeny of a relapsing fever Borrelia species transmitted by the hard tick Ixodes scapularis. Infect. Genet. Evol. 2014, 27, 551–558. [Google Scholar] [CrossRef]
- Nakao, R.; Kasama, K.; Boldbaatar, B.; Ogura, Y.; Kawabata, H.; Toyoda, A.; Hayashi, T.; Takano, A.; Maeda, K. The evolution of hard tick-borne relapsing fever borreliae is correlated with vector species rather than geographical distance. BMC Ecol. Evol. 2021, 21, 105. [Google Scholar] [CrossRef]
- Allan, B.F.; Goessling, L.S.; Storch, G.A.; Thach, R.E. Blood meal analysis to identify reservoir hosts for Amblyomma americanum ticks. Emerg. Infect. Dis. 2010, 16, 433–440. [Google Scholar] [CrossRef]
- Moyer, P.L.; Varela, A.S.; Luttrell, M.P.; Moore, V.A.t.; Stallknecht, D.E.; Little, S.E. White-tailed deer (Odocoileus virginianus) develop spirochetemia following experimental infection with Borrelia lonestari. Vet. Microbiol. 2006, 115, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Krishnavajhala, A.; Wilder, H.K.; Boyle, W.K.; Damania, A.; Thornton, J.A.; Perez de Leon, A.A.; Teel, P.D.; Lopez, J.E. Imaging of Borrelia turicatae Producing the Green Fluorescent Protein Reveals Persistent Colonization of the Ornithodoros turicata Midgut and Salivary Glands from Nymphal Acquisition through Transmission. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef]
- Gugliotta, J.L.; Goethert, H.K.; Berardi, V.P.; Telford, S.R., 3rd. Meningoencephalitis from Borrelia miyamotoi in an immunocompromised patient. N. Engl. J. Med. 2013, 368, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Guglielmone, A.A.; Robbins, R.G.; Apanaskevich, D.A.; Petney, T.N.; Estrada-Peña, A.; Horak, I.G. The Hard Ticks of the World, 1st ed.; Springer: Dordrecht, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Laveran, A. Sur la spirillose des bovidés. CR Acad. Sci. Paris 1903, 136, 939–941. [Google Scholar]
- Theiler, A.; Bruce, D. Transmission and inoculability of Spirillum theileri (Laveran). Proc. R. Soc. Lond. 1905, 76, 504–506. [Google Scholar] [CrossRef]
- Callow, L.L. Observations on tick-transmitted spirochaetes of cattle in Australia and South Africa. Br. Vet. J. 1967, 123, 492–497. [Google Scholar] [CrossRef]
- Krause, P.J.; Fish, D.; Narasimhan, S.; Barbour, A.G. Borrelia miyamotoi infection in nature and in humans. Clin. Microbiol. Infect. 2015, 21, 631–639. [Google Scholar] [CrossRef]
- Fukunaga, M.; Takahashi, Y.; Tsuruta, Y.; Matsushita, O.; Ralph, D.; McClelland, M.; Nakao, M. Genetic and phenotypic analysis of Borrelia miyamotoi sp. nov., isolated from the ixodid tick Ixodes persulcatus, the vector for Lyme disease in Japan. Int. J. Syst. Bacteriol. 1995, 45, 804–810. [Google Scholar] [CrossRef]
- Piesman, J.; Donahue, J.G.; Mather, T.N.; Spielman, A. Transovarially acquired Lyme disease spirochetes (Borrelia burgdorferi) in field-collected larval Ixodes dammini (Acari: Ixodidae). J. Med. Entomol. 1986, 23, 219. [Google Scholar] [CrossRef]
- Rollend, L.; Fish, D.; Childs, J.E. Transovarial transmission of Borrelia spirochetes by Ixodes scapularis: A summary of the literature and recent observations. Ticks Tick Borne Dis. 2013, 4, 46–51. [Google Scholar] [CrossRef]
- Platonov, A.E.; Karan, L.S.; Kolyasnikova, N.M.; Makhneva, N.A.; Toporkova, M.G.; Maleev, V.V.; Fish, D.; Krause, P.J. Humans infected with relapsing fever spirochete Borrelia miyamotoi, Russia. Emerg. Infect. Dis. 2011, 17, 1816–1823. [Google Scholar] [CrossRef]
- Schwan, T.G.; Piesman, J. Vector interactions and molecular adaptations of lyme disease and relapsing fever spirochetes associated with transmission by ticks. Emerg. Infect. Dis. 2002, 8, 115–121. [Google Scholar] [CrossRef]
- Franck, M.; Ghozzi, R.; Pajaud, J.; Lawson-Hogban, N.E.; Mas, M.; Lacout, A.; Perronne, C. Borrelia miyamotoi: 43 Cases Diagnosed in France by Real-Time PCR in Patients With Persistent Polymorphic Signs and Symptoms. Front. Med. (Lausanne) 2020, 7, 55. [Google Scholar] [CrossRef]
- Szekeres, S.; Lakos, A.; Foldvari, G. Borrelia miyamotoi: A recently identified human pathogenic tick-borne relapsing fever spirochete. Orv. Hetil. 2017, 158, 1124–1130. [Google Scholar] [CrossRef]
- Ravagnan, S.; Tomassone, L.; Montarsi, F.; Krawczyk, A.I.; Mastrorilli, E.; Sprong, H.; Milani, A.; Rossi, L.; Capelli, G. First detection of Borrelia miyamotoi in Ixodes ricinus ticks from northern Italy. Parasites Vectors 2018, 11, 130. [Google Scholar] [CrossRef]
- Cutler, S.; Vayssier-Taussat, M.; Estrada-Pena, A.; Potkonjak, A.; Mihalca, A.D.; Zeller, H. A new Borrelia on the block: Borrelia miyamotoi—A human health risk? Eurosurveillance 2019, 24. [Google Scholar] [CrossRef]
- Magnarelli, L.A.; Anderson, J.F.; Johnson, R.C. Cross-reactivity in serological tests for Lyme disease and other spirochetal infections. J. Infect. Dis. 1987, 156, 183–188. [Google Scholar] [CrossRef]
- Jordan, B.E.; Onks, K.R.; Hamilton, S.W.; Hayslette, S.E.; Wright, S.M. Detection of Borrelia burgdorferi and Borrelia lonestari in birds in Tennessee. J. Med. Entomol. 2009, 46, 131–138. [Google Scholar] [CrossRef]
- Barbour, A.G.; Bunikis, J.; Travinsky, B.; Hoen, A.G.; Diuk-Wasser, M.A.; Fish, D.; Tsao, J.I. Niche partitioning of Borrelia burgdorferi and Borrelia miyamotoi in the same tick vector and mammalian reservoir species. Am. J. Trop. Med. Hyg. 2009, 81, 1120–1131. [Google Scholar] [CrossRef]
- Hamsikova, Z.; Coipan, C.; Mahrikova, L.; Minichova, L.; Sprong, H.; Kazimirova, M. Borrelia miyamotoi and Co-Infection with Borrelia afzelii in Ixodes ricinus Ticks and Rodents from Slovakia. Microb. Ecol. 2017, 73, 1000–1008. [Google Scholar] [CrossRef]
- Wagemakers, A.; Jahfari, S.; de Wever, B.; Spanjaard, L.; Starink, M.V.; de Vries, H.J.C.; Sprong, H.; Hovius, J.W. Borrelia miyamotoi in vectors and hosts in The Netherlands. Ticks Tick Borne Dis. 2017, 8, 370–374. [Google Scholar] [CrossRef]
- Hamer, S.A.; Hickling, G.J.; Keith, R.; Sidge, J.L.; Walker, E.D.; Tsao, J.I. Associations of passerine birds, rabbits, and ticks with Borrelia miyamotoi and Borrelia andersonii in Michigan, U.S.A. Parasites Vectors 2012, 5, 231. [Google Scholar] [CrossRef]
- Takano, A.; Toyomane, K.; Konnai, S.; Ohashi, K.; Nakao, M.; Ito, T.; Andoh, M.; Maeda, K.; Watarai, M.; Sato, K.; et al. Tick surveillance for relapsing fever spirochete Borrelia miyamotoi in Hokkaido, Japan. PLoS ONE 2014, 9, e104532. [Google Scholar] [CrossRef]
- Crowder, C.D.; Carolan, H.E.; Rounds, M.A.; Honig, V.; Mothes, B.; Haag, H.; Nolte, O.; Luft, B.J.; Grubhoffer, L.; Ecker, D.J.; et al. Prevalence of Borrelia miyamotoi in Ixodes ticks in Europe and the United States. Emerg. Infect. Dis. 2014, 20, 1678–1682. [Google Scholar] [CrossRef]
- Geller, J.; Nazarova, L.; Katargina, O.; Jarvekulg, L.; Fomenko, N.; Golovljova, I. Detection and genetic characterization of relapsing fever spirochete Borrelia miyamotoi in Estonian ticks. PLoS ONE 2012, 7, e51914. [Google Scholar] [CrossRef]
- Wagemakers, A.; Koetsveld, J.; Narasimhan, S.; Wickel, M.; Deponte, K.; Bleijlevens, B.; Jahfari, S.; Sprong, H.; Karan, L.S.; Sarksyan, D.S.; et al. Variable Major Proteins as Targets for Specific Antibodies against Borrelia miyamotoi. J. Immunol. 2016, 196, 4185–4195. [Google Scholar] [CrossRef]
- Koetsveld, J.; Kolyasnikova, N.M.; Wagemakers, A.; Toporkova, M.G.; Sarksyan, D.S.; Oei, A.; Platonov, A.E.; Hovius, J.W. Development and optimization of an in vitro cultivation protocol allows for isolation of Borrelia miyamotoi from patients with hard tick-borne relapsing fever. Clin. Microbiol. Infect. 2017, 23, 480–484. [Google Scholar] [CrossRef]
- James, A.M.; Liveris, D.; Wormser, G.P.; Schwartz, I.; Montecalvo, M.A.; Johnson, B.J. Borrelia lonestari infection after a bite by an Amblyomma americanum tick. J. Infect. Dis. 2001, 183, 1810–1814. [Google Scholar] [CrossRef] [PubMed]
- Masters, E.J.; Grigery, C.N.; Masters, R.W. STARI, or Masters disease: Lone Star tick-vectored Lyme-like illness. Infect. Dis. Clin. N. Am. 2008, 22, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Goddard, J.; Varela-Stokes, A.S. Role of the lone star tick, Amblyomma americanum (L.), in human and animal diseases. Vet. Parasitol. 2009, 160, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Mackie, F.P. The Part Played by Pediculus Corporis in the Transmission of Relapsing Fever. Br. Med. J. 1907, 2, 1706–1709. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hoch, M.; Wieser, A.; Loscher, T.; Margos, G.; Purner, F.; Zuhl, J.; Seilmaier, M.; Balzer, L.; Guggemos, W.; Rack-Hoch, A.; et al. Louse-borne relapsing fever (Borrelia recurrentis) diagnosed in 15 refugees from northeast Africa: Epidemiology and preventive control measures, Bavaria, Germany, July to October 2015. Eurosurveillance 2015, 20. [Google Scholar] [CrossRef]
- Antinori, S.; Mediannikov, O.; Corbellino, M.; Raoult, D. Louse-borne relapsing fever among East African refugees in Europe. Travel Med. Infect. Dis. 2016, 14, 110–114. [Google Scholar] [CrossRef]
- Amanzougaghene, N.; Akiana, J.; Mongo Ndombe, G.; Davoust, B.; Nsana, N.S.; Parra, H.J.; Fenollar, F.; Raoult, D.; Mediannikov, O. Head Lice of Pygmies Reveal the Presence of Relapsing Fever Borreliae in the Republic of Congo. PLoS Negl. Trop. Dis. 2016, 10, e0005142. [Google Scholar] [CrossRef]
- Cutler, S.J.; Moss, J.; Fukunaga, M.; Wright, D.J.; Fekade, D.; Warrell, D. Borrelia recurrentis characterization and comparison with relapsing-fever, Lyme-associated, and other Borrelia spp. Int. J. Syst. Bacteriol. 1997, 47, 958–968. [Google Scholar] [CrossRef]
- Cutler, S.J.; Fekade, D.; Hussein, K.; Knox, K.A.; Melka, A.; Cann, K.; Emilianus, A.R.; Warrell, D.A.; Wright, D.J. Successful in-vitro cultivation of Borrelia recurrentis. Lancet 1994, 343, 242. [Google Scholar] [CrossRef]
- Marosevic, D.; Margos, G.; Wallich, R.; Wieser, A.; Sing, A.; Fingerle, V. First insights in the variability of Borrelia recurrentis genomes. PLoS Negl. Trop. Dis. 2017, 11, e0005865. [Google Scholar] [CrossRef]
- Ruzic-Sabljic, E.; Maraspin, V.; Stupica, D.; Rojko, T.; Bogovic, P.; Strle, F.; Cerar, T. Comparison of MKP and BSK-H media for the cultivation and isolation of Borrelia burgdorferi sensu lato. PLoS ONE 2017, 12, e0171622. [Google Scholar] [CrossRef]
- Guellil, M.; Kersten, O.; Namouchi, A.; Bauer, E.L.; Derrick, M.; Jensen, A.O.; Stenseth, N.C.; Bramanti, B. Genomic blueprint of a relapsing fever pathogen in 15th century Scandinavia. Proc. Natl. Acad. Sci. USA 2018, 115, 10422–10427. [Google Scholar] [CrossRef]
- Salih, S.Y.; Mustafa, D.; Abdel Wahab, S.M.; Ahmed, M.A.; Omer, A. Louse-borne relapsing fever: I. A clinical and laboratory study of 363 cases in the Sudan. Trans. R. Soc. Trop. Med. Hyg. 1977, 71, 43–48. [Google Scholar] [CrossRef]
- Bryceson, A.D.; Parry, E.H.; Perine, P.L.; Warrell, D.A.; Vukotich, D.; Leithead, C.S. Louse-borne relapsing fever. Q. J. Med. 1970, 39, 129–170. [Google Scholar]
- Vidal, V.; Scragg, I.G.; Cutler, S.J.; Rockett, K.A.; Fekade, D.; Warrell, D.A.; Wright, D.J.; Kwiatkowski, D. Variable major lipoprotein is a principal TNF-inducing factor of louse-borne relapsing fever. Nat. Med. 1998, 4, 1416–1420. [Google Scholar] [CrossRef]
- Sakharoff, M.N. Spirochaeta anserina et la septicémie des oies. Ann. Inst. Pasteur Paris 1891, 564–566. [Google Scholar]
- Hovind-Hougen, K. A morphological characterization of Borrelia anserina. Microbiology (Reading) 1995, 141 Pt 1, 79–83. [Google Scholar] [CrossRef][Green Version]
- Thomas, N.J.; Bunikis, J.; Barbour, A.G.; Wolcott, M.J. Fatal spirochetosis due to a relapsing fever-like Borrelia sp. in a northern spotted owl. J. Wildl. Dis. 2002, 38, 187–193. [Google Scholar] [CrossRef]
- DaMassa, A.J.; Adler, H.E. Avian spirochetosis: Natural transmission by Argas (Persicargas) sanchezi (Ixodoidea: Argasidae) and existence of different serologic and immunologic types of Borrelia anserina in the United States. Am. J. Vet. Res. 1979, 40, 154–157. [Google Scholar]
- McNeil, E.; Hinshaw, W.R.; Kissling, R.E. A Study of Borrelia Anserina Infection (Spirochetosis) in Turkeys. J. Bacteriol. 1949, 57, 191–206. [Google Scholar] [CrossRef]
- Reeves, W.K.; Loftis, A.D.; Sanders, F.; Spinks, M.D.; Wills, W.; Denison, A.M.; Dasch, G.A. Borrelia, Coxiella, and Rickettsia in Carios capensis (Acari: Argasidae) from a brown pelican (Pelecanus occidentalis) rookery in South Carolina, USA. Exp. Appl. Acarol. 2006, 39, 321–329. [Google Scholar] [CrossRef]
- Han, S.W.; Chae, J.B.; Jo, Y.S.; Cho, Y.K.; Kang, J.G.; Shin, N.S.; Youn, H.J.; Youn, H.Y.; Nam, H.M.; Kim, H.J.; et al. First detection of Borrelia and Rickettsia species from Ornithodoros ticks in the Republic of Korea. Ticks Tick Borne Dis. 2021, 12, 101689. [Google Scholar] [CrossRef]
- Ferdows, M.S.; Serwer, P.; Griess, G.A.; Norris, S.J.; Barbour, A.G. Conversion of a linear to a circular plasmid in the relapsing fever agent Borrelia hermsii. J. Bacteriol. 1996, 178, 793–800. [Google Scholar] [CrossRef]
- Elbir, H.; Sitlani, P.; Bergstrom, S.; Barbour, A.G. Chromosome and Megaplasmid Sequences of Borrelia anserina (Sakharoff 1891), the Agent of Avian Spirochetosis and Type Species of the Genus. Genome Announc. 2017, 5. [Google Scholar] [CrossRef]
- Lin, T.; Gao, L.; Seyfang, A.; Oliver, J.H. ‘Candidatus Borrelia texasensis’, from the American dog tick Dermacentor variabilis. Int. J. Syst. Evol. Microbiol. 2005, 55, 685–693. [Google Scholar] [CrossRef]
- Mediannikov, O.; Abdissa, A.; Socolovschi, C.; Diatta, G.; Trape, J.F.; Raoult, D. Detection of a new Borrelia species in ticks taken from cattle in Southwest Ethiopia. Vector Borne Zoonotic Dis. 2013, 13, 266–269. [Google Scholar] [CrossRef]
- Ehounoud, C.B.; Yao, K.P.; Dahmani, M.; Achi, Y.L.; Amanzougaghene, N.; Kacou N’Douba, A.; N’Guessan, J.D.; Raoult, D.; Fenollar, F.; Mediannikov, O. Multiple Pathogens Including Potential New Species in Tick Vectors in Cote d’Ivoire. PLoS Negl. Trop. Dis. 2016, 10, e0004367. [Google Scholar] [CrossRef]
- Kumsa, B.; Socolovschi, C.; Raoult, D.; Parola, P. New Borrelia species detected in ixodid ticks in Oromia, Ethiopia. Ticks Tick Borne Dis 2015, 6, 401–407. [Google Scholar] [CrossRef]
- Evans, N.J.; Bown, K.; Timofte, D.; Simpson, V.R.; Birtles, R.J. Fatal borreliosis in bat caused by relapsing fever spirochete, United Kingdom. Emerg. Infect. Dis. 2009, 15, 1331–1333. [Google Scholar] [CrossRef] [PubMed]

| Sub-Groups | Main Symptoms in Humans | Reservoirs | Vectors | ||
|---|---|---|---|---|---|
| Headache | Fever | Erythema Migrans | |||
| STBRF endemic RF | Yes | Yes | No | Rodents birds, bats, insectivorous Ornithodoros moubata | Ornithodoros sp. Carios kalleyi |
| HTBRF | Yes | Yes | No | Rodents, birds, deer (Odocoileus virginiatus) | Ixodes sp., Amblyomma sp. |
| LBRF epidemic RF | Yes | Yes | No | No reservoirs | Pediculus humanus humanus |
| Avian worldwide RF | No | Birds | Argas sp. | ||
| Borreliae | Hard Ticks | Geographical Areas | Disease (Hosts) | References |
|---|---|---|---|---|
| B. lonestari | Amblyomma americanum (lone star tick) | USA (Missouri, California) | Southern tick-associated rash illness (STARI) (?), (deer (Odocoileus virginianus), migratory birds, wild turkeys (Meleagris gallopavo silvestris)) | [120,121,122] |
| B. miyamotoi | Ixodes persulcatus, I. ricinus, I. scapularis | Asia, Europe, USA | HTBRF (rodents, birds) | [123] |
| B. theileri | Rhipicephalus sp., Margaropus australis | Africa, Australia, Brazil, northern South America | Bovine spirochaestosis (cattle, sheep, goats, horses) | [124,125,126,127] |
| Borrelia | Soft Ticks Vectors | Geographical Areas | Reservoirs/Hosts |
|---|---|---|---|
| B. anserina | Argas sp. | Worldwide | Birds |
| Borrelia | Vectors | Geographical Areas | Reservoirs |
|---|---|---|---|
| B. texasensis | Dermacentor variabilis | USA (Texas) | Unknown |
| Candidatus Borrelia africana Candidatus Borrelia ivorensis | Amblyomma sp. Rhipicephalus sp. | Ivory Coast | |
| Oromia’s Borrelia | Amblyomma sp., Rhipicephalus sp. | Ethiopia | Domestic animals |
| UK pipistrelle bat associated spirochaete | Argas vespertilionis | UK | Bats |
| Borrelia sp. Cachapoal | Ornithodoros sp. Cachapoal | Chile | Rodents |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trevisan, G.; Cinco, M.; Trevisini, S.; di Meo, N.; Ruscio, M.; Forgione, P.; Bonin, S. Borreliae Part 2: Borrelia Relapsing Fever Group and Unclassified Borrelia. Biology 2021, 10, 1117. https://doi.org/10.3390/biology10111117
Trevisan G, Cinco M, Trevisini S, di Meo N, Ruscio M, Forgione P, Bonin S. Borreliae Part 2: Borrelia Relapsing Fever Group and Unclassified Borrelia. Biology. 2021; 10(11):1117. https://doi.org/10.3390/biology10111117
Chicago/Turabian StyleTrevisan, Giusto, Marina Cinco, Sara Trevisini, Nicola di Meo, Maurizio Ruscio, Patrizia Forgione, and Serena Bonin. 2021. "Borreliae Part 2: Borrelia Relapsing Fever Group and Unclassified Borrelia" Biology 10, no. 11: 1117. https://doi.org/10.3390/biology10111117
APA StyleTrevisan, G., Cinco, M., Trevisini, S., di Meo, N., Ruscio, M., Forgione, P., & Bonin, S. (2021). Borreliae Part 2: Borrelia Relapsing Fever Group and Unclassified Borrelia. Biology, 10(11), 1117. https://doi.org/10.3390/biology10111117