
Isorhodopsin: An Undervalued Visual Pigment Analog

Abstract
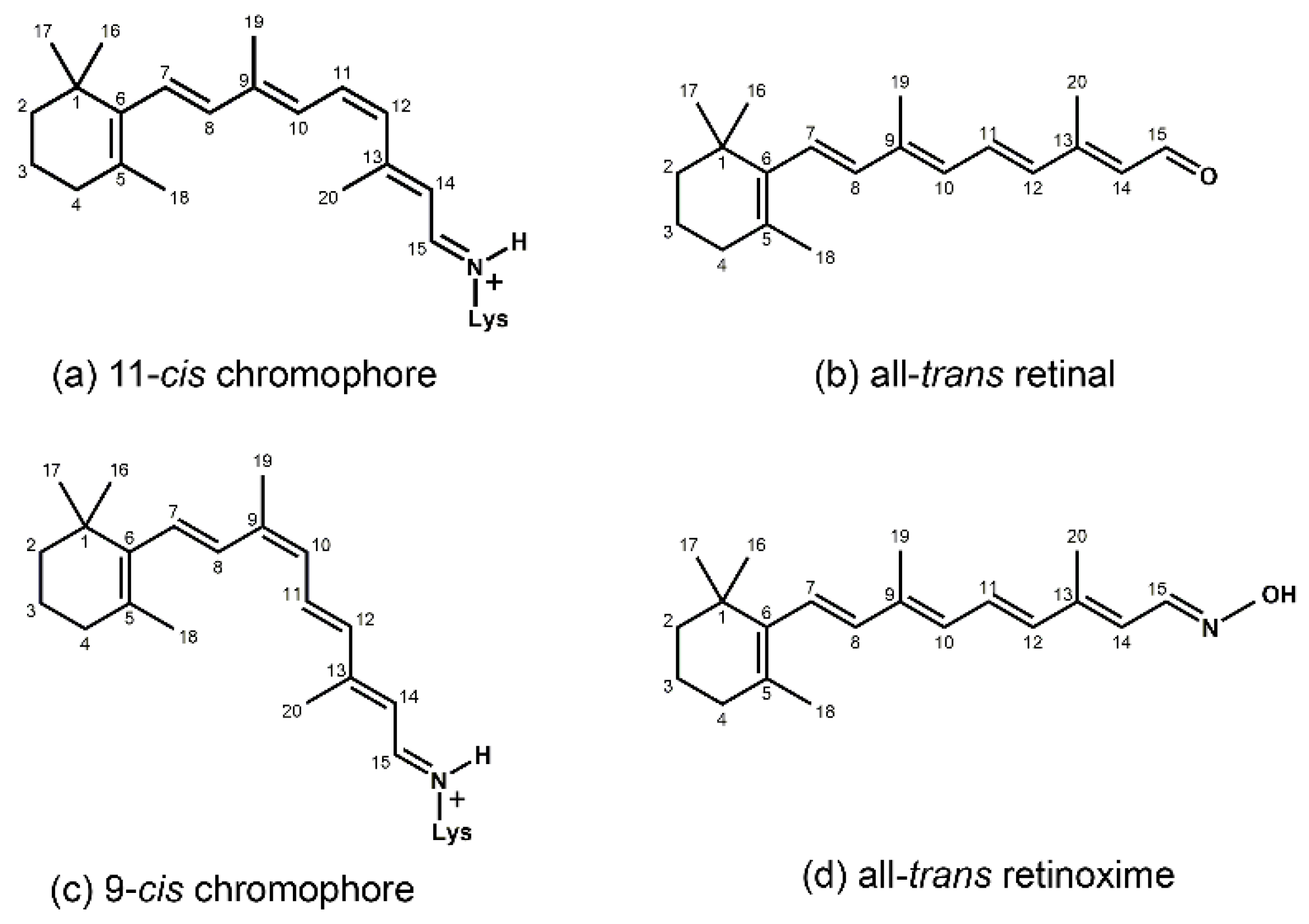
1. Introduction
2. Isorhodopsin—General Aspects
3. Analog Pigments
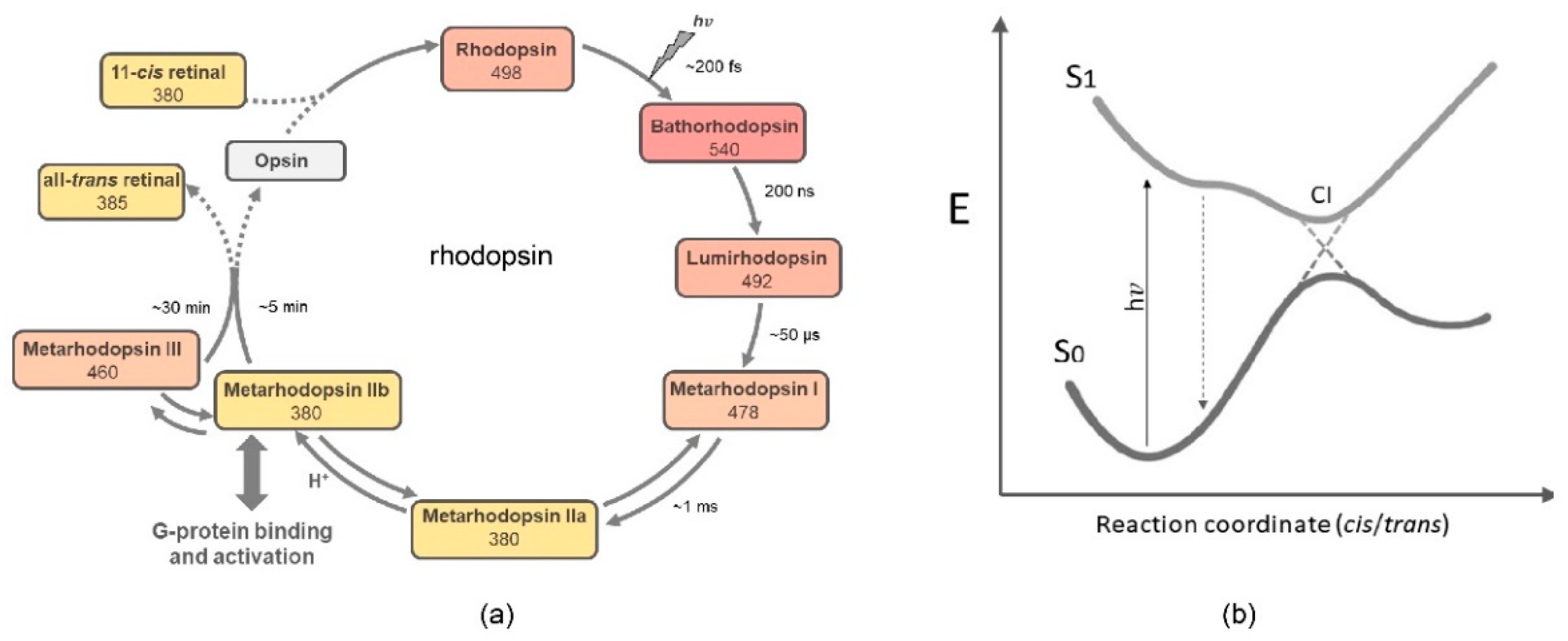
4. Photochemistry

{kind=link}
{kind=link}
| Retinal * | Isomer | λmax Pigment (nm ± 0.02) | Photochemistry | References | |
|---|---|---|---|---|---|
| Time for Batho to Max | Quantum Yield | ||||
| Native (A1) | 11-cis 9-cis | 498 486 | 200 fs 600 fs | 0.65 ± 0.02 0.25 ± 0.04 | [72,99,158,169,170,172,208,209,213,226] |
| 7,8-Dihydro A1 | 11-cis 9-cis | 426 428 | - - | 0.68 ± 0.06 0.39 ± 0.04 | [105,127,134,228,229] |
| 9-Cyclopropyl A1 | 11-cis 9-cis | 492 504 | - - | 0.08 ± 0.04 0.39 ± 0.04 | [105,154] |
| 10-Methyl A1 | 11-cis 9-cis | 506 498 | - - | 0.55 ± 0.07 <0.2 | [130,198,230] |
| 11,12-D2 A1 | 11-cis 9-cis | 498 - | 200 fs - | 0.69 ± 0.02 - | [179] |
| 12-D A1 | 11-cis 9-cis | 498 - | 200 fs - | 0.48 ± 0.03 - | [179] |
| 13-Desmethyl A1 | 11-cis 9-cis | 496 486 | 400 fs - | 0.46 ± 0.04 - | [72,137,221,231,232,233,234] |
| 14-Fluoro A1 | 11-cis 9-cis | 528 510 | - - | 0.55 ± 0.10 0.40 ± 0.06 | [149,198,235,236] |
| 3,4-Dehydro A1 (A2) | 11-cis 9-cis | 518 500 | - - | 0.63 ± 0.04 0.10 ± 0.02 | [155,208,237,238] |
5. Physiological and Medical Relevance
6. Overview and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BR | Bacteriorhodopsin |
| CHAPSO | 3-[(3-Cholamidopropyl)dimethylammonio]-2-hydroxy-1-propanesulfonate |
| Cryo-EM | Cryo-electron microscopy |
| DDM | Dodecylmaltoside |
| DFT | Density functional theory |
| FTIR | Fourier-transform infrared |
| Glu | Glutamate residue in the protein |
| HOOP | Hydrogen out-of-plane vibration |
| LRAT | Lecithin retinol acyltransferase |
| NMR | Nuclear magnetic resonance |
| OG | Octyl glucoside |
| QM/MM | Quantum-mechanical/molecular-mechanical |
| XFEL | X-ray free-electron laser |
References
- Tansley, K. The regeneration of visual purple: Its relation to dark adaptation andnight blindness. J. Physiol. 1931, 71, 442–458. [Google Scholar] [CrossRef] [PubMed]
- Wald, G. Carotenoids and the visual cycle. J. Gen. Physiol. 1935, 19, 351–371. [Google Scholar] [CrossRef] [PubMed]
- Honig, B.; Ebrey, T.G. Structure and spectra of chromophore of visual pigments. Annu. Rev. Biophys. Bioeng. 1974, 3, 151–177. [Google Scholar] [CrossRef] [PubMed]
- Morton, R.A.; Goodwin, T.W. Preparation of Retinene in Vitro. Nature 1944, 153, 405–406. [Google Scholar] [CrossRef]
- Morton, R.A.; Pitt, G.A.J. Visual pigments. Fortschr. Chem. Organ. Nat. 1957, 14, 244–316. [Google Scholar]
- Hubbard, R.; Wald, G. Cis-trans isomers of vitamin A and retinene in the rhodopsin system. J. Gen. Physiol. 1952, 36, 269–315. [Google Scholar] [CrossRef]
- Wald, G. The biochemistry of vision. Annu. Rev. Biochem. 1953, 22, 497–526. [Google Scholar] [CrossRef]
- Baylor, D.A. Photoreceptor signals and vision. Invest. Ophthalmol. Vis. Sci. 1987, 28, 34–49. [Google Scholar]
- Bloomfield, S.A.; Dacheux, R.F. Rod vision: Pathways and processing in the mammalian retina. Prog. Retin. Eye Res. 2001, 20, 351–384. [Google Scholar] [CrossRef]
- Porciatti, V.; Pizzorusso, T.; Maffei, L. The visual physiology of the wild type mouse determined with pattern VEPs. Vis. Res. 1999, 39, 3071–3081. [Google Scholar] [CrossRef]
- McConnell, S.K. The generation of neuronal diversity in the central nervous system. Annu. Rev. Neurosci. 1991, 14, 269–300. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, K.P. Late photoproducts and signaling states of bovine rhodopsin. In Molecular Mechanisms in Visual Transduction; Stavenga, D.G., DeGrip, W.J., Pugh, E.N., Jr., Eds.; Elsevier Science Pub.: Amsterdam, The Netherlands, 2000; Volume 3, pp. 91–142. [Google Scholar]
- Rothschild, K.J.; Gillespie, J.; DeGrip, W.J. Evidence for rhodopsin refolding during the decay of meta II. Biophys. J. 1987, 51, 345–350. [Google Scholar] [CrossRef]
- Park, J.H.; Scheerer, P.; Hofmann, K.P.; Choe, H.-W.; Ernst, O.P. Crystal structure of the ligand-free G-protein-coupled receptor opsin. Nature 2008, 454, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Jäger, S.; Palczewski, K.; Hofmann, K.P. Opsin/all-trans-retinal complex activates transducin by different mechanisms than photolyzed rhodopsin. Biochemistry 1996, 35, 2901–2908. [Google Scholar] [CrossRef]
- Sato, S.; Jastrzebska, B.; Engel, A.; Palczewski, K.; Kefalov, V.J. Apo-Opsin Exists in Equilibrium Between a Predominant Inactive and a Rare Highly Active State. J. Neurosci. 2019, 39, 212–223. [Google Scholar] [CrossRef]
- Hubbard, R.; Brown, P.K.; Bownds, M.D. Methodology of vitamin A and visual pigments. Methods Enzym. 1971, 18C, 615–653. [Google Scholar]
- Kiser, P.D.; Golczak, M.; Maeda, A.; Palczewski, K. Key enzymes of the retinoid (visual) cycle in vertebrate retina. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2012, 1821, 137–151. [Google Scholar] [CrossRef]
- Choi, E.H.; Daruwalla, A.; Suh, S.; Leinonen, H.; Palczewski, K. Retinoids in the visual cycle: Role of the retinal G protein-coupled receptor. J. Lipid Res. 2021, 62, 100040. [Google Scholar] [CrossRef]
- Hecht, S.; Shlaer, S.; Pirenne, M.H. Energy, quanta, and vision. J. Gen. Physiol. 1942, 25, 819–840. [Google Scholar] [CrossRef]
- Musilova, Z.; Cortesi, F.; Matschiner, M.; Davies, W.I.L.; Patel, J.S.; Stieb, S.M.; de Busserolles, F.; Malmstrom, M.; Torresen, O.K.; Brown, C.J.; et al. Vision using multiple distinct rod opsins in deep-sea fishes. Science 2019, 364, 588–592. [Google Scholar] [CrossRef]
- Dartnall, H.J.A. The Identity and Distribution of Visual Pigments in the Animal Kingdom. In The Visual Process, 1st ed.; Davson, H., Ed.; Academic Press: New York, NY, USA, 1962; Volume 2, pp. 367–426. [Google Scholar]
- Crescitelli, F. The natural history of visual pigments: 1990. Prog. Retin. Res. 1991, 11, 1–32. [Google Scholar] [CrossRef]
- Imamoto, Y.; Shichida, Y. Cone visual pigments. Biochim. Biophys. Acta-Bioenerg. 2014, 1837, 664–673. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, S.; Yokoyama, R. Comparative Molecular Biology of Visual Pigments. In Molecular Mechanisms in Visual Transduction; Stavenga, D.G., DeGrip, W.J., Pugh, E.N., Jr., Eds.; Elsevier Science Pub.: Amsterdam, The Netherlands, 2000; Volume 3, pp. 257–296. [Google Scholar]
- Gärtner, W. Invertebrate visual pigments. In Molecular Mechanisms in Visual Transduction; Stavenga, D.G., DeGrip, W.J., Pugh, E.N., Jr., Eds.; Elsevier Science Pub.: Amsterdam, The Netherlands, 2000; Volume 3, pp. 298–388. [Google Scholar]
- Tsukamoto, H.; Terakita, A. Diversity and functional properties of bistable pigments. Photochem. Photobiol. Sci. 2010, 9, 1435–1443. [Google Scholar] [CrossRef]
- Stavenga, D.G. Insect retinal pigments: Spectral characteristics and physiological functions. Prog. Retin. Eye Res. 1995, 15, 231–259. [Google Scholar] [CrossRef]
- Bertolucci, C.; Foà, A. Extraocular photoreception and circadian entrainment in nonmammalian vertebrates. Chronobiol. Int. 2004, 21, 501–519. [Google Scholar] [CrossRef]
- Peirson, S.N.; Halford, S.; Foster, R.G. The evolution of irradiance detection: Melanopsin and the non-visual opsins. Philos. Trans. R. Soc. B—Biol. Sci. 2009, 364, 2849–2865. [Google Scholar]
- Koyanagi, M.; Terakita, A. Diversity of animal opsin-based pigments and their optogenetic potential. Biochim. Biophys. Acta-Bioenerg. 2014, 1837, 710–716. [Google Scholar] [CrossRef]
- Davies, W.I.L.; Tamai, T.K.; Zheng, L.; Fu, J.K.; Rihel, J.; Foster, R.G.; Whitmore, D.; Hankins, M.W. An extended family of novel vertebrate photopigments is widely expressed and displays a diversity of function. Genome Res. 2015, 25, 1666–1679. [Google Scholar] [CrossRef]
- Palczewski, K. G protein-coupled receptor rhodopsin. Annu. Rev. Biochem. 2006, 75, 743–767. [Google Scholar] [CrossRef]
- Kühn, H. Interactions between photoexcited rhodopsin and light-activated enzymes in rods. Prog. Retin. Res. 1984, 3, 123–156. [Google Scholar] [CrossRef]
- Oesterhelt, D.; Stoeckenius, W. Rhodopsin-like Protein from the Purple Membrane of Halobacterium halobium. Nat. New Biol. 1971, 233, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Oesterhelt, D. The structure and mechanism of the family of retinal proteins from halophilic archaea. Curr. Opin. Struct. Biol. 1998, 8, 489–500. [Google Scholar] [CrossRef]
- Nagata, T.; Inoue, K. Rhodopsins at a glance. J. Cell Sci. 2022, 134, jcs258989. [Google Scholar] [CrossRef] [PubMed]
- Rozenberg, A.; Inoue, K.; Kandori, H.; Béjà, O. Microbial Rhodopsins: The Last Two Decades. Annu. Rev. Microbiol. 2021, 75, 427–447. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Hegemann, P.; Broser, M. Enzymerhodopsins: Novel photoregulated catalysts for optogenetics. Curr. Opin. Struct. Biol. 2019, 57, 118–126. [Google Scholar] [CrossRef]
- Govorunova, E.G.; Sineshchekov, O.A.; Li, H.; Spudich, J.L. Microbial Rhodopsins: Diversity, Mechanisms, and Optogenetic Applications. Annu. Rev. Biochem. 2017, 86, 845–872. [Google Scholar] [CrossRef]
- Koenekoop, R.K.; Sui, R.F.; Sallum, J.; van den Born, L.I.; Ajlan, R.; Khan, A.; den Hollander, A.I.; Cremers, F.P.M.; Mendola, J.D.; Bittner, A.K.; et al. Oral 9-cis retinoid for childhood blindness due to Leber congenital amaurosis caused by RPE65 or LRAT mutations: An open-label phase 1b trial. Lancet 2014, 384, 1513–1520. [Google Scholar] [CrossRef]
- Maeda, T.; Maeda, A.; Casadesus, G.; Palczewski, K.; Margaron, P. Evaluation of 9-cis-retinyl acetate therapy in RPE65−/− mice. Invest. Ophthalmol. Vis. Sci. 2009, 50, 4368–4378. [Google Scholar] [CrossRef]
- Fan, J.; Rohrer, B.; Moiseyev, G.; Ma, J.-X.; Crouch, R.K. Isorhodopsin rather than rhodopsin mediates rod function in RPE65 knock-out mice. Proc. Natl. Acad. Sci. USA 2003, 100, 13662–13667. [Google Scholar] [CrossRef]
- Honig, B.; Karplus, M. Implications of Torsional Potential of Retinal Isomers for Visual Excitation. Nature 1971, 229, 558–560. [Google Scholar] [CrossRef]
- Nakanishi, K.; Crouch, R.K. Application of artificial pigments to structure determination and study of photoinduced transformations of retinal proteins. Isr. J. Chem. 1995, 35, 253–272. [Google Scholar] [CrossRef]
- Ebrey, T.G.; Govindjee, R.; Honig, B.; Pollock, E.; Chan, W.; Crouch, R.K.; Yudd, A.; Nakanishi, K. Properties of several sterically modified retinal analogs and their photosensitive pigments. Biochemistry 1975, 14, 3933–3941. [Google Scholar] [CrossRef]
- Huang, L.; Deng, H.; Weng, G.; Koutalos, Y.; Ebrey, T.G.; Groesbeek, M.; Lugtenburg, J.; Tsuda, M.; Callender, R.H. A resonance raman study of the C=N configurations of octopus rhodopsin, bathorhodopsin, and isorhodopsin. Biochemistry 1996, 35, 8504–8510. [Google Scholar] [CrossRef] [PubMed]
- Bagley, K.A.; Balogh-Nair, V.; Croteau, A.A.; Dollinger, G.; Ebrey, T.G.; Eisenstein, L.; Hong, M.K.; Nakanishi, K.; Vittitow, J. Fourier-transform infrared difference spectroscopy of rhodopsin and its photoproducts at low temperature. Biochemistry 1985, 24, 6055–6071. [Google Scholar] [CrossRef]
- Deng, H.; Huang, L.; Groesbeek, M.; Lugtenburg, J.; Callender, R.H. Vibrational analysis of a retinal protonated schiff-base analog. J. Phys. Chem. 1994, 98, 4776–4779. [Google Scholar] [CrossRef]
- Kandori, H.; Matuoka, S.; Shichida, Y.; Yoshizawa, T.; Ito, M.; Tsukida, K.; Balogh-Nair, V.; Nakanishi, K. Mechanism of isomerization of rhodopsin studied by use of 11-cis-locked rhodopsin analogs excited with a picosecond laser pulse. Biochemistry 1989, 28, 6460–6467. [Google Scholar] [CrossRef]
- Ito, M.; Hiroshima, T.; Tsukida, K.; Shichida, Y.; Yoshizawa, T. A novel rhodopsin analog possessing the conformationally 6-s-cis-fixed retinylidene chromophore. J. Chem. Soc. Chem. Commun. 1985, 1443–1444. [Google Scholar] [CrossRef]
- Ito, M.; Mantani, Y.; Tsukida, K.; Shichida, Y.; Ioshida, S.; Fukada, Y.; Yoshizawa, T. A novel rhodopsin analog with bicyclic retinal involving the 8–18 bonded structure in the chromophore. J. Nutr. Sci. Vitam. 1988, 34, 641–646. [Google Scholar] [CrossRef]
- Ito, M.; Katsuta, Y.; Imamoto, Y.; Shichida, Y.; Yoshizawa, T. Conformational analysis of the rhodopsin chromophore using bicyclic retinal analogues. Photochem. Photobiol. 1992, 56, 915–919. [Google Scholar] [CrossRef]
- Fukada, Y.; Shichida, Y.; Yoshizawa, T.; Ito, M.; Kodama, A.; Tsukida, K. Studies on structure and function of rhodopsin by use of cyclopentatrienylidene 11-cis-locked-rhodopsin. Biochemistry 1984, 23, 5826–5832. [Google Scholar] [CrossRef]
- Mollevanger, L.C.P.J.; Kentgens, A.P.M.; Pardoen, J.A.; Courtin, J.M.L.; Veeman, W.S.; Lugtenburg, J.; DeGrip, W.J. High-resolution solid-state 13C-NMR study of carbons C-5 and C-12 of the chromophore of bovine rhodopsin: Evidence for a 6-S-cis conformation with negative-charge perturbation near C-12. Eur. J. Biochem. 1987, 163, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Jäger, F.; Jäger, S.; Kräutle, O.; Friedman, N.; Sheves, M.; Hofmann, K.P.; Siebert, F. Interactions of the β-ionone ring with the protein in the visual pigment rhodopsin control the activation mechanism. An FTIR and fluorescence study on artificial vertebrate rhodopsins. Biochemistry 1994, 33, 7389–7397. [Google Scholar] [CrossRef] [PubMed]
- Nakamichi, H.; Buss, V.; Okada, T. Photoisomerization mechanism of rhodopsin and 9-cis-rhodopsin revealed by X-ray crystallography. Biophys. J. 2007, 92, L106–L108. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vogel, R.; Lüdeke, S.; Siebert, F.; Sakmar, T.P.; Hirshfeld, A.; Sheves, M. Agonists and partial agonists of rhodopsin: Retinal polyene methylation affects receptor activation. Biochemistry 2006, 45, 1640–1652. [Google Scholar] [CrossRef]
- Ganter, U.M.; Schmid, E.D.; Perez-Sala, D.; Rando, R.R.; Siebert, F. Removal of the 9-methyl group of retinal inhibits signal transduction in the visual process. A Fourier-transform infrared and biochemical investigation. Biochemistry 1989, 28, 5954–5962. [Google Scholar] [CrossRef]
- Khorana, H.G. Rhodopsin, photoreceptor of the rod cell—An emerging pattern for structure and function. J. Biol. Chem. 1992, 267, 1–4. [Google Scholar] [CrossRef]
- DeGrip, W.J.; Bonting, S.L.; Daemen, F.J.M. The binding site of retinaldehyde in cattle rhodopsin. Biochim. Biophys. Acta 1973, 303, 189–193. [Google Scholar] [CrossRef]
- Sekharan, S.; Sugihara, M.; Weingart, O.; Okada, T.; Buss, V. Protein assistance in the photoisomerization of rhodopsin and 9-cis-rhodopsin-Insights from experiment and theory. J. Am. Chem. Soc. 2007, 129, 1052–1054. [Google Scholar] [CrossRef]
- Vogel, R.; Fan, G.-B.; Sheves, M.; Siebert, F. The molecular origin of the inhibition of transducin activation in rhodopsin lacking the 9-methyl group of the retinal chromophore: A UV-Vis and FTIR spectroscopic study. Biochemistry 2000, 39, 8895–8908. [Google Scholar] [CrossRef]
- Ganter, U.M.; Gärtner, W.; Siebert, F. The influence of the 13-methyl group of the retinal on the photoreaction of rhodopsin revealed by FTIR difference spectroscopy. Eur. Biophys. J. 1990, 18, 295–299. [Google Scholar] [CrossRef]
- Ebrey, T.G.; Tsuda, M.; Sassenrath, G.; West, J.L.; Waddell, W.H. Light activation of bovine rod phosphodiesterase by non-physiological visual pigments. FEBS Lett. 1980, 116, 217–219. [Google Scholar] [CrossRef]
- Okada, T.; Sugihara, M.; Bondar, A.-N.; Elstner, M.; Entel, P.; Buss, V. The retinal conformation and its environment in rhodopsin in light of a new 2.2 Å crystal structure. J. Mol. Biol. 2004, 342, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Edwards, P.C.; Burghammer, M.; Villa, C.; Schertler, G.F.X. Structure of bovine rhodopsin in a trigonal crystal form. J. Mol. Biol. 2004, 343, 1409–1438. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-J.; Bovee-Geurts, P.H.M.; Lugtenburg, J.; DeGrip, W.J. Constraints of the 9-methyl group binding pocket of the rhodopsin chromophore probed by 9-halogeno substitution. Biochemistry 2004, 43, 14802–14810. [Google Scholar] [CrossRef]
- Smith, S.O.; Palings, I.; Miley, M.E.; Courtin, J.M.L.; de Groot, H.J.M.; Lugtenburg, J.; Mathies, R.A.; Griffin, R.G. Solid-state NMR studies of the mechanism of the opsin shift in the visual pigment rhodopsin. Biochemistry 1990, 29, 8158–8164. [Google Scholar] [CrossRef]
- Rothschild, K.J.; Cantore, W.A.; Marrero, H. Fourier transform infrared difference spectra of intermediates in rhodopsin bleaching. Science 1983, 219, 1333–1335. [Google Scholar] [CrossRef]
- Eyring, G.; Curry, B.; Broek, A.; Lugtenburg, J.; Mathies, R.A. Assignment and interpretation of hydrogen out-of-plane vibrations in the resonance Raman spectra of rhodopsin and bathorhodopsin. Biochemistry 1982, 21, 384–393. [Google Scholar] [CrossRef]
- Kochendoerfer, G.G.; Verdegem, P.J.E.; Van der Hoef, I.; Lugtenburg, J.; Mathies, R.A. Retinal analog study of the role of steric interactions in the excited state isomerization dynamics of rhodopsin. Biochemistry 1996, 35, 16230–16240. [Google Scholar] [CrossRef]
- Verdegem, P.J.E.; Bovee-Geurts, P.H.M.; DeGrip, W.J.; Lugtenburg, J.; de Groot, H.J.M. Retinylidene ligand structure in bovine rhodopsin, metarhodopsin I, and 10-methylrhodopsin from internuclear distance measurements using 13C-labeling and 1-D rotational resonance MAS NMR. Biochemistry 1999, 38, 11316–11324. [Google Scholar] [CrossRef]
- Feng, X.; Verdegem, P.J.E.; Lee, Y.K.; Sandström, D.; Edén, M.; Bovee-Geurts, P.H.M.; DeGrip, W.J.; Lugtenburg, J.; de Groot, H.J.M.; Levitt, M.H. Direct determination of a molecular torsional angle in the membrane protein rhodopsin by solid-state NMR. J. Am. Chem. Soc. 1997, 119, 6853–6857. [Google Scholar] [CrossRef]
- Gröbner, G.; Choi, G.; Burnett, I.J.; Glaubitz, C.; Verdegem, P.J.E.; Lugtenburg, J.; Watts, A. Photoreceptor rhodopsin: Structural and conformational study of its chromophore 11-cis retinal in oriented membranes by deuterium solid state NMR. FEBS Lett. 1998, 422, 201–204. [Google Scholar] [CrossRef]
- Carravetta, M.; Zhao, X.; Johannessen, O.G.; Lai, W.C.; Verhoeven, M.A.; Bovee-Geurts, P.H.M.; Verdegem, P.J.E.; Kiihne, S.R.; Luthman, H.; de Groot, H.J.M.; et al. Protein-induced bonding perturbation of the rhodopsin chromophore detected by double-quantum solid-state NMR. J. Am. Chem. Soc. 2004, 126, 3948–3953. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brown, M.F.; Heyn, M.P.; Job, C.; Kim, S.; Moltke, S.; Nakanishi, K.; Nevzorov, A.A.; Struts, A.V.; Salgado, G.F.J.; Wallat, I. Solid-State 2H NMR spectroscopy of retinal proteins in aligned membranes. Biochim. Biophys. Acta-Biomembr. 2007, 1768, 2979–3000. [Google Scholar] [CrossRef] [PubMed]
- Palings, I.; Pardoen, J.A.; VanDenBerg, E.M.M.; Winkel, C.; Lugtenburg, J.; Mathies, R.A. Assignment of fingerprint vibrations in the resonance Raman spectra of rhodopsin, isorhodopsin, and bathorhodopsin: Implications for chromophore structure and environment. Biochemistry 1987, 26, 2544–2556. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.C.; Nanbu, S.; Ishida, T. QM/MM trajectory surface hopping approach to photoisomerization of rhodopsin and isorhodopsin: The origin of faster and more efficient isomerization for rhodopsin. J. Phys. Chem. B 2012, 116, 8009–8023. [Google Scholar] [CrossRef]
- Sugihara, M.; Buss, V.; Entel, P.; Hafner, J. The nature of the complex counterion of the chromophore in rhodopsin. J. Phys. Chem. B 2004, 108, 3673–3680. [Google Scholar] [CrossRef]
- Sekharan, S.; Sugihara, M.; Buss, V. Origin of spectral tuning in rhodopsin—It is not the binding pocket. Angew. Chem. Int. Ed. 2007, 46, 269–271. [Google Scholar] [CrossRef]
- Ernst, O.P.; Lodowski, D.T.; Elstner, M.; Hegemann, P.; Brown, L.S.; Kandori, H. Microbial and animal rhodopsins: Structures, functions, and molecular mechanisms. Chem. Rev. 2014, 114, 126–163. [Google Scholar] [CrossRef]
- Lesca, E.; Panneels, V.; Schertler, G.F.X. The role of water molecules in phototransduction of retinal proteins and G protein-coupled receptors. Faraday Discuss. 2018, 207, 27–37. [Google Scholar] [CrossRef]
- Nikolaev, D.M.; Shtyrov, A.A.; Mereshchenko, A.S.; Panov, M.S.; Tveryanovich, Y.S.; Ryazantsev, M.N. An assessment of water placement algorithms in quantum mechanics/molecular mechanics modeling: The case of rhodopsins’ first spectral absorption band maxima. Phys. Chem. Chem. Phys. 2020, 22, 18114–18123. [Google Scholar] [CrossRef]
- Siebert, F.; Mäntele, W.; Gerwert, K. Fourier-transform infrared spectroscopy applied to rhodopsin. The problem of the protonation state of the retinylidene Schiff base re-investigated. Eur. J. Biochem. 1983, 136, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, M.A.; Creemers, A.F.L.; Bovee-Geurts, P.H.M.; DeGrip, W.J.; Lugtenburg, J.; de Groot, H.J.M. Ultra-high-field MAS NMR assay of a multispin labeled ligand bound to its G-protein receptor target in the natural membrane environment: Electronic structure of the retinylidene chromophore in rhodopsin. Biochemistry 2001, 40, 3282–3288. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Fujiyoshi, Y.; Silow, M.; Navarro, J.; Landau, E.M.; Shichida, Y. Functional role of internal water molecules in rhodopsin revealed by X-ray crystallography. Proc. Natl. Acad. Sci. USA 2002, 99, 5982–5987. [Google Scholar] [CrossRef] [PubMed]
- Sakmar, T.P.; Franke, R.R.; Khorana, H.G. Glutamic acid-113 serves as the retinylidene Schiff base counterion in bovine rhodopsin. Proc. Natl. Acad. Sci. USA 1989, 86, 8309–8313. [Google Scholar] [CrossRef]
- Creemers, A.F.L.; Klaassen, C.H.W.; Bovee-Geurts, P.H.M.; Kelle, R.; Kragl, U.; Raap, J.; DeGrip, W.J.; Lugtenburg, J.; de Groot, H.J.M. 15N Solid state NMR evidence for a compex Schiff base counterion in the visual G-protein coupled receptor rhodopsin. Biochemistry 1999, 38, 7195–7199. [Google Scholar] [CrossRef]
- Deng, H.; Huang, L.W.; Callender, R.H.; Ebrey, T.G. Evidence for a bound water molecule next to the retinal Schiff base in bacteriorhodopsin and rhodopsin: A resonance raman study of the Schiff base hydrogen/deuterium exchange. Biophys. J. 1994, 66, 1129–1136. [Google Scholar] [CrossRef][Green Version]
- Nagata, T.; Terakita, A.; Kandori, H.; Kojima, D.; Shichida, Y.; Maeda, A. Water and peptide backbone structure in the active center of bovine rhodopsin. Biochemistry 1997, 36, 6164–6170. [Google Scholar] [CrossRef]
- Nagata, T.; Terakita, A.; Kandori, H.; Shichida, Y.; Maeda, A. The hydrogen-bonding network of water molecules and the peptide backbone in the region connecting Asp83, Gly120, and Glu113 in bovine rhodopsin. Biochemistry 1998, 37, 17216–17222. [Google Scholar] [CrossRef]
- Fujimoto, K.J. Electronic Couplings and Electrostatic Interactions Behind the Light Absorption of Retinal Proteins. Front. Mol. Biosci. 2021, 8, 752700. [Google Scholar] [CrossRef]
- Shtyrov, A.A.; Nikolaev, D.M.; Mironov, V.N.; Vasin, A.V.; Panov, M.S.; Tveryanovich, Y.S.; Ryazantsev, M.N. Simple Models to Study Spectral Properties of Microbial and Animal Rhodopsins: Evaluation of the Electrostatic Effect of Charged and Polar Residues on the First Absorption Band Maxima. Int. J. Mol. Sci. 2021, 22, 3029. [Google Scholar] [CrossRef]
- Honig, B.; Dinur, U.; Nakanishi, K.; Balogh-Nair, V.; Gawinowicz, M.A.; Arnaboldi, M.; Motto, M.G. An external point-charge model for wavelength regulation in visual pigments. J. Am. Chem. Soc. 1979, 101, 7084–7086. [Google Scholar] [CrossRef]
- Demoulin, B.; Maiuri, M.; Berbasova, T.; Geiger, J.H.; Borhan, B.; Garavelli, M.; Cerullo, G.; Rivalta, I. Control of Protonated Schiff Base Excited State Decay within Visual Protein Mimics: A Unified Model for Retinal Chromophores. Chem. Eur. J. 2021, 27, 16389–16400. [Google Scholar] [CrossRef] [PubMed]
- Agathangelou, D.; Roy, P.P.; Del Carmen Marín, M.; Ferré, N.; Olivucci, M.; Buckup, T.; Léonard, J.; Haacke, S. Sub-picosecond C=C bond photo-isomerization: Evidence for the role of excited state mixing. Compt. Rendus. Phys. 2021, 22, 1–28. [Google Scholar] [CrossRef]
- Curry, B.; Palings, I.; Broek, A.D.; Pardoen, J.A.; Lugtenburg, J.; Mathies, R.A. Vibrational Analysis of the Retinal Isomers. In Advances in Infrared and Raman Spectroscopy; Clark, R.J.H., Hester, R.E., Eds.; Wiley, Heyden: Chichester, UK, 1985; Volume 12, pp. 115–178. [Google Scholar]
- Birge, R.R.; Einterz, C.M.; Knapp, H.M.; Murray, L.P. The nature of the primary photochemical events in rhodopsin and isorhodopsin. Biophys. J. 1988, 53, 367–385. [Google Scholar] [CrossRef][Green Version]
- Cooper, A. Energetics of rhodopsin and isorhodopsin. FEBS Lett. 1979, 100, 382–384. [Google Scholar] [CrossRef]
- Makino, C.L.; Kraft, T.W.; Mathies, R.A.; Lugtenburg, J.; Miley, M.E.; van der Steen, R.; Baylor, D.A. Effects of modified chromophores on the spectral sensitivity of salamander, squirrel and macaque cones. J. Physiol. 1990, 424, 545–560. [Google Scholar] [CrossRef]
- Creemers, A.F.L.; Bovee-Geurts, P.H.M.; DeGrip, W.J.; Lugtenburg, J.; de Groot, H.J.M. Solid-state NMR analysis of ligand-receptor interactions reveals an induced misfit in the binding site of isorhodopsin. Biochemistry 2004, 43, 16011–16018. [Google Scholar] [CrossRef]
- Mirzadegan, T.; Liu, R.S.H. Probing the visual pigment rhodopsin and its analogs by molecular modeling analysis and computer graphics. Prog. Retin. Res. 1991, 11, 57–74. [Google Scholar] [CrossRef]
- Nakamichi, H.; Okada, T. X-ray crystallographic analysis of 9-cis-rhodopsin, a model analogue visual pigment. Photochem. Photobiol. 2007, 83, 232–235. [Google Scholar] [CrossRef]
- Bovee-Geurts, P.H.M.; Lugtenburg, J.; DeGrip, W.J. Coupled HOOP signature correlates with quantum yield of isorhodopsin and analog pigments. Biochim. Biophys. Acta—Bioenerg. 2017, 1858, 118–125. [Google Scholar] [CrossRef]
- Mathies, R.A.; Smith, S.O.; Palings, I. Determination of retinal chromophore structure in rhodopsins. In Resonance Raman Spectra of Polyenes and Aromatics; Spiro, T.G., Ed.; John Wiley & Sons: New York, NY, USA, 1987; Volume 2, pp. 59–108. [Google Scholar]
- Palings, I.; Van den Berg, E.M.M.; Lugtenburg, J.; Mathies, R.A. Complete assignment of the hydrogen out-of-plane wagging vibrations of bathorhodopsin: Chromophore structure and energy storage in the primary photoproduct of vision. Biochemistry 1989, 28, 1498–1507. [Google Scholar] [CrossRef] [PubMed]
- Eyring, G.; Curry, B.; Mathies, R.A.; Fransen, R.; Palings, I.; Lugtenburg, J. Interpretation of the resonance Raman spectrum of bathorhodopsin based on visual pigment analogues. Biochemistry 1980, 19, 2410–2418. [Google Scholar] [CrossRef] [PubMed]
- Mathies, R.A.; Freedman, T.B.; Stryer, L. Resonance Raman studies of conformation of retinal in rhodopsin and isorhodopsin. J. Mol. Biol. 1977, 109, 367–372. [Google Scholar] [CrossRef]
- Fahmy, K.; Siebert, F.; Grossjean, M.F.; Tavan, P. Photoisomerization in bacteriorhodopsin studied by FTIR, linear dichroism and photoselection experiments combined with quantum chemical theoretical-analysis. J. Mol. Struct. 1989, 214, 257–288. [Google Scholar] [CrossRef]
- Warshel, A.; Barboy, N. Energy storage and reaction pathways in the first step of the vision process. J. Am. Chem. Soc. 1982, 104, 1469–1476. [Google Scholar] [CrossRef]
- Dawadi, P.B.S.; Lugtenburg, J. Synthesis and use of stable isotope enriched retinals in the field of vitamin A. Molecules 2010, 15, 1825–1872. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.S.H.; Asato, A.E. The binding site of opsin based on analog studies with isomeric, fluorinated, alkylated, and other modified retinals. In Chemistry and Biology of Synthetic Retinoids; Dawson, M.I., Okamura, W.H., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 1990; pp. 52–75. [Google Scholar]
- Lugtenburg, J.; Mathies, R.A.; Griffin, R.G.; Herzfeld, J. Structure and function of rhodopsins from solid state NMR and resonance Raman spectroscopy of isotopic retinal derivatives. Trends. Biochem. Sci. 1988, 13, 388–393. [Google Scholar] [CrossRef]
- DeGrip, W.J.; DeLange, F.; Klaassen, C.H.W.; Verdegem, P.J.E.; Wallace-Williams, S.E.; Creemers, A.F.L.; Bergo, V.B.; Bovee-Geurts, P.H.M.; Raap, J.; Rothschild, K.J.; et al. Photoactivation of rhodopsin: Interplay between protein and chromophore. In Rhodopsins and Phototransduction; Goode, J.A., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 1999; Volume 224, pp. 102–118. [Google Scholar]
- Creemers, A.F.L.; Kiihne, S.R.; Bovee-Geurts, P.H.M.; DeGrip, W.J.; Lugtenburg, J.; de Groot, H.J.M. 1H and 13C MAS NMR evidence for pronounced ligand-protein interactions involving the ionone ring of the retinylidene chromophore in rhodopsin. Proc. Natl. Acad. Sci. USA 2002, 99, 9101–9106. [Google Scholar] [CrossRef]
- Kiihne, S.R.; Creemers, A.F.L.; DeGrip, W.J.; Bovee-Geurts, P.H.M.; Lugtenburg, J.; de Groot, H.J.M. Selective interface detection: Mapping binding site contacts in membrane proteins by NMR spectroscopy. J. Am. Chem. Soc. 2005, 127, 5734–5735. [Google Scholar] [CrossRef]
- Brinkmann, A.; Sternberg, U.; Bovee-Geurts, P.H.M.; Fernández Fernández, I.; Lugtenburg, J.; Kentgens, A.P.M.; DeGrip, W.J. Insight into the chromophore of rhodopsin and its Meta-II photointermediate by 19F solid-state NMR and chemical shift tensor calculations. Phys. Chem. Chem. Phys. 2018, 20, 30174–30188. [Google Scholar] [CrossRef]
- Carravetta, M.; Edén, M.; Johannessen, O.G.; Luthman, H.; Verdegem, P.J.E.; Lugtenburg, J.; Sebald, A.; Levitt, M.H. Estimation of carbon-carbon bond lengths and medium-range internuclear distances by solid-state nuclear magnetic resonance. J. Am. Chem. Soc. 2001, 123, 10628–10638. [Google Scholar] [CrossRef] [PubMed]
- Peersen, O.B.; Smith, S.O. Rotational resonance NMR of biological membranes. Concepts Magn. Reson. 1993, 5, 303–317. [Google Scholar] [CrossRef]
- Spooner, P.J.R.; Sharples, J.M.; Verhoeven, M.A.; Lugtenburg, J.; Glaubitz, C.; Watts, A. Relative orientation between the β-ionone ring and the polyene chain for the chromophore of rhodopsin in native membranes. Biochemistry 2002, 41, 7549–7555. [Google Scholar] [CrossRef] [PubMed]
- Spooner, P.J.R.; Sharples, J.M.; Goodall, S.C.; Bovee-Geurts, P.H.M.; Verhoeven, M.A.; Lugtenburg, J.; Pistorius, A.M.A.; DeGrip, W.J.; Watts, A. The ring of the rhodopsin chromophore in a hydrophobic activation switch within the binding pocket. J. Mol. Biol. 2004, 343, 719–730. [Google Scholar] [CrossRef]
- Yoshizawa, T. Photophysiological functions of visual pigments. Adv. Biophys. 1984, 17, 5–67. [Google Scholar] [CrossRef]
- Nonaka, Y.; Hanai, S.; Katayama, K.; Imai, H.; Kandori, H. Unique Retinal Binding Pocket of Primate Blue-Sensitive Visual Pigment. Biochemistry 2020, 59, 2602–2607. [Google Scholar] [CrossRef]
- Fasick, J.I.; Lee, N.; Oprian, D.D. Spectral tuning in the human blue cone pigment. Biochemistry 1999, 38, 11593–11596. [Google Scholar] [CrossRef]
- Kropf, A.; Whittenberger, B.P.; Goff, S.P.; Waggoner, A.S. The spectral properties of some visual pigment analogs. Exp. Eye Res. 1973, 17, 591–606. [Google Scholar] [CrossRef]
- Derguini, F.; Nakanishi, K. Synthetic rhodopsin analogs. Photobiochem. Photobiophys. 1986, 13, 259–283. [Google Scholar]
- Álvarez, R.; Vaz, B.; Gronemeyer, H.; de Lera, A.R. Functions, therapeutic applications, and synthesis of retinoids and carotenoids. Chem. Rev. 2014, 114, 1–125. [Google Scholar] [CrossRef]
- Kahremany, S.; Sander, C.L.; Tochtrop, G.P.; Kubas, A.; Palczewski, K. Z-isomerization of retinoids through combination of monochromatic photoisomerization and metal catalysis. Org. Biomol. Chem. 2019, 17, 8125–8139. [Google Scholar] [CrossRef] [PubMed]
- DeLange, F.; Bovee-Geurts, P.H.M.; VanOostrum, J.; Portier, M.D.; Verdegem, P.J.E.; Lugtenburg, J.; DeGrip, W.J. An additional methyl group at the 10-position of retinal dramatically slows down the kinetics of the rhodopsin photocascade. Biochemistry 1998, 37, 1411–1420. [Google Scholar] [PubMed]
- Trehan, A.; Liu, R.S.H.; Shichida, Y.; Imamoto, Y.; Nakamura, K.; Yoshizawa, T. On retention of chromophore configuration of rhodopsin isomers derived from three dicis retinal isomers. Bioorg. Chem. 1990, 18, 30–40. [Google Scholar]
- DeGrip, W.J.; VanOostrum, J.; Bovee-Geurts, P.H.M.; van der Steen, R.; VanAmsterdam, L.J.P.; Groesbeek, M.; Lugtenburg, J. 10,20-Methanorhodopsins: (7E,9E,13E)-10,20-methanorhodopsin and (7E,9Z,13Z)-10,20-methanorhodopsin-11-cis-Locked rhodopsin analog pigments with unusual thermal and photo-stability. Eur. J. Biochem. 1990, 191, 211–220. [Google Scholar]
- Wang, Y.-J.; Bovee-Geurts, P.H.M.; Lugtenburg, J.; DeGrip, W.J. Alpha-retinals as rhodopsin chromophores—Preference for the 9-Z configuration and partial agonist activity. Photochem. Photobiol. 2008, 84, 889–894. [Google Scholar] [CrossRef] [PubMed]
- DeGrip, W.J.; Bovee-Geurts, P.H.M.; Van der Hoef, I.; Lugtenburg, J. 7,8-Dihydro-retinals outperform the native retinals in conferring photosensitivity to visual opsin. J. Am. Chem. Soc. 2007, 129, 13265–13269. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, R.S.H. Divergent pathways in photobleaching of 7,9-dicis-rhodopsin and 9,11-dicis-12-fluororhodopsin: One-photon-two-bond and one-photon- one-bond isomerization. Biochemistry 1993, 32, 10233–10238. [Google Scholar] [CrossRef]
- Shichida, Y.; Nakamura, K.; Yoshizawa, T.; Trehan, A.; Denny, M.; Liu, R.S.H. 9,13-dicis-Rhodopsin and Its One-Photon-One-Double-Bond Isomerization. Biochemistry 1988, 27, 6495–6499. [Google Scholar] [CrossRef]
- Liu, R.S.H.; Mirzadegan, T. The shape of a three-dimensional binding-site of rhodopsin based on molecular modeling analyses of isomeric and other visual pigment analogs. Bioorganic studies of visual pigments. 11. J. Am. Chem. Soc. 1988, 110, 8617–8623. [Google Scholar] [CrossRef]
- Groenendijk, G.W.T.; DeGrip, W.J.; Daemen, F.J.M. Identification and characterization of syn-isomers and anti-isomers of retinaloximes. Anal. Biochem. 1979, 99, 304–310. [Google Scholar] [CrossRef]
- Groenendijk, G.W.T.; DeGrip, W.J.; Daemen, F.J.M. Quantitative determination of retinals with complete retention of their geometric configuration. Biochim. Biophys. Acta 1980, 617, 430–438. [Google Scholar] [CrossRef]
- Kini, A.; Matsumoto, H.; Liu, R.S.H. 9,11-Di-cis-retinal and 9,11-Di-cis-rhodopsin. Bioorg. Chem. 1980, 9, 406–410. [Google Scholar] [CrossRef]
- Zhu, Y.; Ganapathy, S.; Trehan, A.; Asato, A.E.; Liu, R.S.H. FT-IR spectra of all sixteen isomers of retinal, their isolation, and other spectroscopic properties. Tetrahedron 1992, 48, 10061–10074. [Google Scholar] [CrossRef]
- Groenendijk, G.W.T.; Jacobs, C.W.M.; Bonting, S.L.; Daemen, F.J.M. Dark isomerization of retinals in the presence of phosphatidylethanolamine. Eur. J. Biochem. 1980, 106, 119–128. [Google Scholar] [CrossRef]
- Braiman, M.S.; Rothschild, K.J. Fourier transform infrared techniques for probing membrane protein structure. Annu. Rev. Biophys. Biophys. Chem. 1988, 17, 541–570. [Google Scholar] [CrossRef]
- Kochendoerfer, G.G.; Lin, S.W.; Sakmar, T.P.; Mathies, R.A. How color visual pigments are tuned. Trends. Biochem. Sci. 1999, 24, 300–305. [Google Scholar] [CrossRef]
- Hirano, T.; Fujioka, N.; Imai, H.; Kandori, H.; Wada, A.; Ito, M.; Shichida, Y. Assignment of the vibrational modes of the chromophores of iodopsin and bathoiodopsin: Low-temperature Fourier transform infrared spectroscopy of 13C and 2H-labeled iodopsins. Biochemistry 2006, 45, 1285–1294. [Google Scholar] [CrossRef]
- Lin, S.W.; Imamoto, Y.; Fukada, Y.; Shichida, Y.; Yoshizawa, T.; Mathies, R.A. What makes red visual pigments red? A resonance Raman microprobe study of retinal chromophore structure in iodopsin. Biochemistry 1994, 33, 2151–2160. [Google Scholar] [CrossRef]
- Pande, C.; Pande, A.; Yue, K.T.; Callender, R.H.; Ebrey, T.G.; Tsuda, M. Resonance Raman spectroscopy of octopus rhodopsin and its photoproducts. Biochemistry 1987, 26, 4941–4947. [Google Scholar] [CrossRef]
- Bagley, K.A.; Eisenstein, L.; Ebrey, T.G.; Tsuda, M. A comparative study of the infrared difference spectra for octopus and bovine rhodopsins and their bathorhodopsin photointermediates. Biochemistry 1989, 28, 3366–3373. [Google Scholar] [CrossRef]
- Bovee-Geurts, P.H.M.; Fernández Fernández, I.; Liu, R.S.H.; Mathies, R.A.; Lugtenburg, J.; DeGrip, W.J. Fluoro derivatives of retinal illuminate the decisive role of the C12-H element in photoisomerization and rhodopsin activation. J. Am. Chem. Soc. 2009, 131, 17933–17942. [Google Scholar] [CrossRef] [PubMed]
- Oseroff, A.R.; Callender, R.H. Resonance Raman spectroscopy of rhodopsin in retinal disk membranes. Biochemistry 1974, 13, 4243–4248. [Google Scholar] [CrossRef] [PubMed]
- Popp, A.; Ujj, L.; Atkinson, G.H. Vibrational spectra of room-temperature rhodopsin: Concentration dependence in picosecond resonance coherent anti-stokes Raman scattering. Biophys. Chem. 1995, 56, 129–135. [Google Scholar] [CrossRef]
- Jäger, F.; Ujj, L.; Atkinson, G.H. Vibrational spectrum of bathorhodopsin in the room-temperature rhodopsin photoreaction. J. Am. Chem. Soc. 1997, 119, 12610–12618. [Google Scholar] [CrossRef]
- Lin, S.W.; Groesbeek, M.; Van der Hoef, I.; Verdegem, P.J.E.; Lugtenburg, J.; Mathies, R.A. Vibrational assignment of torsional normal modes of rhodopsin: Probing excited-state isomerization dynamics along the reactive C11=C12 torsion coordinate. J. Phys. Chem. B 1998, 102, 2787–2806. [Google Scholar] [CrossRef]
- DeGrip, W.J.; Bovee-Geurts, P.H.M.; Wang, Y.-J.; Verhoeven, M.A.; Lugtenburg, J. Cyclopropyl and isopropyl derivatives of 11-cis and 9-cis retinals at C-9 and C-13: Subtle steric differences with major effects on ligand efficacy in rhodopsin. J. Nat. Prod. 2011, 74, 383–390. [Google Scholar] [CrossRef]
- Barry, B.; Mathies, R.A.; Pardoen, J.A.; Lugtenburg, J. Raman microscope and quantum yield studies on the primary photochemistry of A2-visual pigments. Biophys. J. 1987, 52, 603–610. [Google Scholar] [CrossRef]
- Bridges, C.D.B. The rhodopsin-porphyropsin visual system. In Photochemistry of Vision; Dartnall, H.J.A., Ed.; Springer: Berlin/Heidelberg, Germany, 1972; Volume VII/1, pp. 417–480. [Google Scholar]
- Imai, H.; Hirano, T.; Terakita, A.; Shichida, Y.; Muthyala, R.S.; Chen, R.-L.; Colmenares, L.U.; Liu, R.S.H. Probing for the threshold energy for visual transduction: Red-shifted visual pigment analogs from 3-methoxy-3-dehydroretinal and related compounds. Photochem. Photobiol. 1999, 70, 111–115. [Google Scholar] [CrossRef]
- Polli, D.; Rivalta, I.; Nenov, A.; Weingart, O.; Garavelli, M.; Cerullo, G. Tracking the primary photoconversion events in rhodopsins by ultrafast optical spectroscopy. Photochem. Photobiol. Sci. 2015, 14, 213–228. [Google Scholar] [CrossRef]
- Panneels, V.; Wu, W.T.; Tsai, C.-J.; Nogly, P.; Rheinberger, J.; Jaeger, K.; Cicchetti, G.; Gati, C.; Kick, L.M.; Sala, L.; et al. Time-resolved structural studies with serial crystallography: A new light on retinal proteins. Struct. Dyn. 2015, 2, 041718. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Kandori, H. Primary photochemical events in the rhodopsin molecule. Prog. Retin. Res. 1991, 11, 33–55. [Google Scholar] [CrossRef]
- Farrens, D.L. What site-directed labeling studies tell us about the mechanism of rhodopsin activation and G-protein binding. Photochem. Photobiol. Sci. 2010, 9, 1466–1474. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Smith, S.O. Structure and activation of the visual pigment rhodopsin. Annu. Rev. Biophys. 2010, 39, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.S.; Ladizhansky, V. Membrane proteins in their native habitat as seen by solid-state NMR spectroscopy. Protein Sci. 2015, 24, 1333–1346. [Google Scholar] [CrossRef] [PubMed]
- Mathies, R.A.; Lugtenburg, J. The primary photoreaction of rhodopsin. In Molecular Mechanisms in Visual Transduction; Stavenga, D.G., DeGrip, W.J., Pugh, E.N., Jr., Eds.; Elsevier Science Pub.: Amsterdam, The Netherlands, 2000; Volume 3, pp. 55–90. [Google Scholar]
- DeGrip, W.J.; Rothschild, K.J. Structure and mechanism of vertebrate visual pigments. In Molecular Mechanisms in Visual Transduction; Stavenga, D.G., DeGrip, W.J., Pugh, E.N., Jr., Eds.; Elsevier Science Pub.: Amsterdam, The Netherlands, 2000; Volume 3, pp. 1–54. [Google Scholar]
- Weingart, O.; Schapiro, I.; Buss, V. Photochemistry of visual pigment chromophore models by ab initio molecular dynamics. J. Phys. Chem. B 2007, 111, 3782–3788. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, T.; Wald, G. Pre-lumirhodopsin and the bleaching of visual pigments. Nature 1963, 197, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, T. The behaviour of visual pigments at low temperatures. In Photochemistry of Vision; Dartnall, H.J.A., Ed.; Springer: Berlin/Heidelberg, Germany, 1972; Volume VII/I, pp. 146–179. [Google Scholar]
- Hurley, J.B.; Ebrey, T.G.; Honig, B.; Ottolenghi, M. Temperature and wavelength effects on photochemistry of rhodopsin, isorhodopsin, bacteriorhodopsin and their photoproducts. Nature 1977, 270, 540–542. [Google Scholar] [CrossRef]
- Kim, J.E.; Tauber, M.J.; Mathies, R.A. Wavelength dependent cis-trans isomerization in vision. Biochemistry 2001, 40, 13774–13778. [Google Scholar] [CrossRef]
- Kim, J.E.; Tauber, M.J.; Mathies, R.A. Analysis of the mode-specific excited-state energy distribution and wavelength-dependent photoreaction quantum yield in rhodopsin. Biophys. J. 2003, 84, 2492–2501. [Google Scholar] [CrossRef]
- Hubbard, R.; Kropf, A. The action of light on rhodopsin. Proc. Natl. Acad. Sci. USA 1958, 44, 130–139. [Google Scholar] [CrossRef]
- Kropf, A.; Hubbard, R. The photoisomerization of retinal. Photochem. Photobiol. 1970, 12, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Becker, R.S.; Freedman, K. A comprehensive investigation of the mechanism and photophysics of isomerization of a protonated and unprotonated Schiff-base of 11-cis-retinal. J. Am. Chem. Soc. 1985, 107, 1477–1485. [Google Scholar] [CrossRef]
- Becker, R.S.; Freedman, K.; Hutchinson, J.A.; Noe, L.J. Kinetic study of the photoisomerization of a protonated Schiff-base of 11-cis-retinal over the picosecond-to-second time regimes. J. Am. Chem. Soc. 1985, 107, 3942–3944. [Google Scholar] [CrossRef]
- Lewis, J.W.; Hug, S.J.; Wallace-Williams, S.E.; Kliger, D.S. Direct evidence for an equilibrium between early photolysis intermediates of rhodopsin. J. Am. Chem. Soc. 1990, 112, 6711–6712. [Google Scholar] [CrossRef]
- Birge, R.R. Nature of the primary photochemical events in rhodopsin and bacteriorhodopsin. Biochim. Biophys. Acta 1990, 1016, 293–327. [Google Scholar] [CrossRef]
- Yang, X.; Manathunga, M.; Gozem, S.; Léonard, J.; Andruniów, T.; Olivucci, M. Quantum-classical simulations of rhodopsin reveal excited-state population splitting and its effects on quantum efficiency. Nat. Chem. 2022, 14, 441–449. [Google Scholar] [CrossRef]
- Schnedermann, C.; Yang, X.; Liebel, M.; Spillane, K.M.; Lugtenburg, J.; Fernández, I.; Valentini, A.; Schapiro, I.; Olivucci, M.; Kukura, P.; et al. Evidence for a vibrational phase-dependent isotope effect on the photochemistry of vision. Nat. Chem. 2018, 10, 449–455. [Google Scholar] [CrossRef]
- Johnson, P.J.M.; Halpin, A.; Morizumi, T.; Prokhorenko, V.I.; Ernst, O.P.; Miller, R.J.D. Local vibrational coherences drive the primary photochemistry of vision. Nat. Chem. 2015, 7, 980–986. [Google Scholar] [CrossRef]
- Schick, G.A.; Cooper, T.M.; Holloway, R.A.; Murray, L.P.; Birge, R.R. Energy-storage in the primary photochemical events of rhodopsin and isorhodopsin. Biochemistry 1987, 26, 2556–2562. [Google Scholar] [CrossRef]
- Cooper, A. Energy uptake in the first step of visual excitation. Nature 1979, 282, 531–533. [Google Scholar] [CrossRef]
- Birge, R.R.; Vought, B.W. Energetics of rhodopsin photobleaching: Photocalorimetric studies of energy storage in early and later intermediates. Methods Enzym. 2000, 315, 143–163. [Google Scholar]
- Nakamichi, H.; Okada, T. Local peptide movement in the photoreaction intermediate of rhodopsin. Proc. Natl. Acad. Sci. USA 2006, 103, 12729–12734. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.W.; Jäger, S.; Kliger, D.S. Absorbance changes by aromatic amino acid side chains in early rhodopsin photointermediates. Photochem. Photobiol. 1997, 66, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Doukas, A.G.; Aton, B.; Callender, R.H.; Ebrey, T.G. Resonance Raman studies of bovine metarhodopsin I and metarhodopsin II. Biochemistry 1978, 17, 2430–2435. [Google Scholar] [CrossRef]
- Furutani, Y.; Kandori, H.; Shichida, Y. Structural changes in lumirhodopsin and metarhodopsin I studied by their photoreactions at 77 K. Biochemistry 2003, 42, 8494–8500. [Google Scholar] [CrossRef]
- Schertler, G.F.X. Structure of rhodopsin and the metarhodopsin I photointermediate. Curr. Opin. Struct. Biol. 2005, 15, 408–415. [Google Scholar] [CrossRef]
- Vogel, R.; Sakmar, T.P.; Sheves, M.; Siebert, F. Coupling of protonation switches during rhodopsin activation. Photochem. Photobiol. 2007, 83, 286–292. [Google Scholar] [CrossRef]
- Pope, A.L.; Sanchez-Reyes, O.B.; South, K.; Zaitseva, E.; Ziliox, M.; Vogel, R.; Reeves, P.J.; Smith, S.O. A Conserved Proline Hinge Mediates Helix Dynamics and Activation of Rhodopsin. Structure 2020, 28, 1004–1013. [Google Scholar] [CrossRef]
- Vogel, R.; Mahalingam, M.; Lüdeke, S.; Huber, T.; Siebert, F.; Sakmar, T.P. Functional role of the “Ionic Lock”—An interhelical hydrogen-bond network in family a heptahelical receptors. J. Mol. Biol. 2008, 380, 648–655. [Google Scholar] [CrossRef]
- Jäger, F.; Fahmy, K.; Sakmar, T.P.; Siebert, F. Identification of glutamic acid 113 as the Schiff base proton acceptor in the metarhodopsin II photointermediate of rhodopsin. Biochemistry 1994, 33, 10878–10882. [Google Scholar] [CrossRef]
- Maeda, A. Internal water molecules as mobile polar groups for light-induced proton translocation in bacteriorhodopsin and rhodopsin as studied by difference FTIR spectroscopy. Biochemistry 2001, 66, 1256–1268. [Google Scholar] [PubMed]
- Furutani, Y.; Shichida, Y.; Kandori, H. Structural changes of water molecules during the photoactivation processes in bovine rhodopsin. Biochemistry 2003, 42, 9619–9625. [Google Scholar] [CrossRef] [PubMed]
- Choe, H.-W.; Kim, Y.J.; Park, J.H.; Morizumi, T.; Pai, E.F.; Krauß, N.; Hofmann, K.P.; Scheerer, P.; Ernst, O.P. Crystal structure of metarhodopsin II. Nature 2011, 471, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Jastrzebska, B.; Palczewski, K.; Golczak, M. Role of bulk water in hydrolysis of the rhodopsin chromophore. J. Biol. Chem. 2011, 286, 18930–18937. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, R.; Stevens, C.F. Arrestin binding determines the rate of inactivation of the G protein-coupled receptor rhodopsin in vivo. Cell 1995, 81, 841–848. [Google Scholar] [CrossRef]
- Liu, R.S.H.; Crescitelli, F.; Denny, M.; Matsumoto, H.; Asato, A.E. Photosensitivity of 10-substituted visual pigment analogues: Detection of a specific secondary opsin-retinal interaction. Biochemistry 1986, 25, 7026–7030. [Google Scholar] [CrossRef]
- Fan, J.; Woodruff, M.L.; Cilluffo, M.C.; Crouch, R.K.; Fain, G.L. Opsin activation of transduction in the rods of dark-reared Rpe65 knockout mice. J. Physiol. 2005, 568, 83–95. [Google Scholar] [CrossRef]
- Concistrè, M.; Gansmüller, A.; McLean, N.; Johannessen, O.G.; Marín-Montesinos, I.; Bovee-Geurts, P.H.M.; Verdegem, P.J.E.; Lugtenburg, J.; Brown, R.C.D.; DeGrip, W.J.; et al. Double-quantum 13C nuclear magnetic resonance of bathorhodopsin, the first photointermediate in mammalian vision. J. Am. Chem. Soc. 2008, 130, 10490–10491. [Google Scholar] [CrossRef]
- Nakamichi, H.; Okada, T. Crystallographic analysis of primary visual photochemistry. Angew. Chem. Int. Ed. 2006, 45, 4270–4273. [Google Scholar] [CrossRef]
- Kukura, P.; McCamant, D.W.; Yoon, S.; Wandschneider, D.B.; Mathies, R.A. Structural observation of the primary isomerization in vision with femtosecond-stimulated Raman. Science 2005, 310, 1006–1009. [Google Scholar] [CrossRef]
- DeLange, F.; Bovee-Geurts, P.H.M.; Pistorius, A.M.A.; Rothschild, K.J.; DeGrip, W.J. Probing intramolecular orientations in rhodopsin and metarhodopsin II by polarized infrared difference spectroscopy. Biochemistry 1999, 38, 13200–13209. [Google Scholar] [CrossRef] [PubMed]
- Popp, A.; Ujj, L.; Atkinson, G.H. Bathorhodopsin structure in the room-temperature rhodopsin photosequence: Picosecond time-resolved coherent anti-Stokes Raman scattering. Proc. Natl. Acad. Sci. USA 1996, 93, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Ebrey, T.G.; Crouch, R. Bathoproducts of rhodopsin, isorhodopsin-i, and isorhodopsin-ii. Biophys. J. 1980, 29, 247–256. [Google Scholar] [CrossRef]
- Lewis, J.W.; Liang, J.; Ebrey, T.G.; Sheves, M.; Livnah, N.; Kuwata, O.; Jäger, S.; Kliger, D.S. Early photolysis intermediates of gecko and bovine artificial visual pigments. Biochemistry 1997, 36, 14593–14600. [Google Scholar] [CrossRef]
- Polli, D.; Weingart, O.; Brida, D.; Poli, E.; Maiuri, M.; Spillane, K.M.; Bottoni, A.; Kukura, P.; Mathies, R.A.; Cerullo, G.; et al. Wavepacket splitting and two-pathway deactivation in the photoexcited visual pigment isorhodopsin. Angew. Chem. Int. Ed. 2014, 53, 2504–2507. [Google Scholar] [CrossRef]
- Dartnall, H.J.A. The photosensitivities of visual pigments in presence of hydroxylamine. Vis. Res. 1968, 8, 339–358. [Google Scholar] [CrossRef]
- Schoenlein, R.W.; Peteanu, L.A.; Mathies, R.A.; Shank, C.V. The first step in vision: Femtosecond isomerization of rhodopsin. Science 1991, 254, 412–415. [Google Scholar] [CrossRef]
- Wang, Q.; Schoenlein, R.W.; Peteanu, L.A.; Mathies, R.A.; Shank, C.V. Vibrationally coherent photochemistry in the femtosecond primary event of vision. Science 1994, 266, 422–424. [Google Scholar] [CrossRef]
- McCamant, D.W.; Kukura, P.; Mathies, R.A. Femtosecond stimulated Raman study of excited-state evolution in bacteriorhodopsin. J. Phys. Chem. B 2005, 109, 10449–10457. [Google Scholar] [CrossRef]
- Kim, J.E.; McCamant, D.W.; Zhu, L.Y.; Mathies, R.A. Resonance Raman structural evidence that the cis-to-trans isomerization in rhodopsin occurs in femtoseconds. J. Phys. Chem. B 2001, 105, 1240–1249. [Google Scholar] [CrossRef]
- Peteanu, L.A.; Schoenlein, R.W.; Wang, Q.; Mathies, R.A.; Shank, C.V. The first step in vision occurs in femtoseconds: Complete blue and red spectral studies. Proc. Natl. Acad. Sci. USA 1993, 90, 11762–11766. [Google Scholar] [CrossRef] [PubMed]
- Weingart, O.; Altoè, P.; Stenta, M.; Bottoni, A.; Orlandi, G.; Garavelli, M. Product formation in rhodopsin by fast hydrogen motions. Phys. Chem. Chem. Phys. 2011, 13, 3645–3648. [Google Scholar] [CrossRef] [PubMed]
- Schapiro, I.; Ryazantsev, M.N.; Frutos, L.M.; Ferré, N.; Lindh, R.; Olivucci, M. The ultrafast photoisomerizations of rhodopsin and bathorhodopsin are modulated by bond length alternation and HOOP driven electronic effects. J. Am. Chem. Soc. 2011, 133, 3354–3364. [Google Scholar] [CrossRef] [PubMed]
- Strambi, A.; Coto, P.B.; Frutos, L.M.; Ferré, N.; Olivucci, M. Relationship between the excited state relaxation paths of rhodopsin and isorhodopsin. J. Am. Chem. Soc. 2008, 130, 3382–3388. [Google Scholar] [CrossRef] [PubMed]
- Polli, D.; Altoè, P.; Weingart, O.; Spillane, K.M.; Manzoni, C.; Brida, D.; Tomasello, G.; Orlandi, G.; Kukura, P.; Mathies, R.A.; et al. Conical intersection dynamics of the primary photoisomerization event in vision. Nature 2010, 467, 440–443. [Google Scholar] [CrossRef]
- Weingart, O. The twisted C11=C12 bond of the rhodopsin chromophore—A photochemical hot spot. J. Am. Chem. Soc. 2007, 129, 10618–10619. [Google Scholar] [CrossRef]
- Gozem, S.; Luk, H.L.; Schapiro, I.; Olivucci, M. Theory and Simulation of the Ultrafast Double-Bond lsomerization of Biological Chromophores. Chem. Rev. 2017, 117, 13502–13565. [Google Scholar] [CrossRef]
- Weingart, O.; Garavelli, M. Modelling vibrational coherence in the primary rhodopsin photoproduct. J. Chem. Phys. 2012, 137, 22A523. [Google Scholar] [CrossRef]
- Verhoeven, M.A.; Bovee-Geurts, P.H.M.; de Groot, H.J.M.; Lugtenburg, J.; DeGrip, W.J. Methyl substituents at the 11- or 12-position of retinal profoundly and differentially affect photochemistry and signalling activity of rhodopsin. J. Mol. Biol. 2006, 363, 98–113. [Google Scholar] [CrossRef]
- Nogly, P.; Weinert, T.; James, D.; Carbajo, S.; Ozerov, D.; Furrer, A.; Gashi, D.; Borin, V.; Skopintsev, P.; Jaeger, K.; et al. Retinal isomerization in bacteriorhodopsin captured by a femtosecond X-ray laser. Science 2018, 361, 145–151. [Google Scholar] [CrossRef]
- Skopintsev, P.; Ehrenberg, D.; Weinert, T.; James, D.; Kar, R.K.; Johnson, P.J.M.; Ozerov, D.; Furrer, A.; Martiel, I.; Dworkowski, F.; et al. Femtosecond-to-millisecond structural changes in a light-driven sodium pump. Nature 2020, 583, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Oda, K.; Nomura, T.; Nakane, T.; Yamashita, K.; Inoue, K.; Ito, S.; Vierock, J.; Hirata, K.; Maturana, A.D.; Katayama, K.; et al. Time-resolved serial femtosecond crystallography reveals early structural changes in channelrhodopsin. eLife 2021, 10, 62389. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.S.H.; Asato, A.E. Photochemistry of polyenes 22. The primary process of vision and the structure of bathorhodopsin: A mechanism for photoisomerization of polyenes. Proc. Natl. Acad. Sci. USA 1985, 82, 259–263. [Google Scholar] [CrossRef]
- Schoenlein, R.W.; Peteanu, L.A.; Wang, Q.; Mathies, R.A.; Shank, C.V. Femtosecond Dynamics of Cis-Trans Isomerization in a Visual Pigment Analog: Isorhodopsin. J. Phys. Chem. 1993, 97, 12087–12092. [Google Scholar] [CrossRef]
- Albeck, A.; Friedman, N.; Ottolenghi, M.; Sheves, M.; Einterz, C.M.; Hug, S.J.; Lewis, J.W.; Kliger, D.S. Photolysis intermediates of the artificial visual pigment cis-5,6-dihydro-isorhodopsin. Biophys. J. 1989, 55, 233–241. [Google Scholar] [CrossRef]
- Muto, O.; Tokunaga, F.; Yoshizawa, T.; Kamat, V.; Blatchly, H.A.; Balogh-Nair, V.; Nakanishi, K. Photochemical reaction of 7,8-dihydrorhodopsin at low temperatures. Biochim. Biophys. Acta-Lipids Lipid Metab. 1984, 766, 597–602. [Google Scholar] [CrossRef]
- Koutalos, Y.; Ebrey, T.G.; Tsuda, M.; Odashima, K.; Lien, T.; Park, M.H.; Shimizu, N.; Derguini, F.; Nakanishi, K.; Gilson, H.R.; et al. Regeneration of bovine and octopus opsins in situ with natural and artificial retinals. Biochemistry 1989, 28, 2732–2739. [Google Scholar] [CrossRef]
- Asato, A.E.; Denny, M.; Matsumoto, H.; Mirzadegan, T.; Ripka, W.C.; Crescitelli, F.; Liu, R.S.H. Study of the shape of the binding site of bovine opsin using 10-substituted retinal isomers. Biochemistry 1986, 25, 7021–7026. [Google Scholar] [CrossRef]
- Wang, Q.; Kochendoerfer, G.G.; Schoenlein, R.W.; Verdegem, P.J.E.; Lugtenburg, J.; Mathies, R.A.; Shank, C.V. Femtosecond spectroscopy of a 13-demethylrhodopsin visual pigment analogue: The role of non-bonded interactions in the isomerization process. J. Phys. Chem. B 1996, 100, 17388–17394. [Google Scholar] [CrossRef]
- Shichida, Y.; Kropf, A.; Yoshizawa, T. Photochemical reactions of 13-demethyl visual pigment analogues at low temperatures. Biochemistry 1981, 20, 1962–1968. [Google Scholar] [CrossRef]
- Nelson, R.; Deriel, J.K.; Kropf, A. 13-Desmethyl rhodopsin and 13-desmethyl isorhodopsin—Visual pigment analogues. Proc. Natl. Acad. Sci. USA 1970, 66, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Blatz, P.E.; Lin, M.; Balasubramaniyan, P.; Balasubramaniyan, V.; Dewhurst, P.B. A new series of synthetic visual pigments from cattle opsin and homologs of retinal. J. Am. Chem. Soc. 1969, 91, 5930–5931. [Google Scholar] [CrossRef] [PubMed]
- Colmenares, L.U.; Asato, A.E.; Denny, M.; Mead, D.; Zingoni, J.P.; Liu, R.S.H. NMR studies of fluorinated visual pigment analogs. Biochem. Biophys. Res. Commun. 1991, 179, 1337–1343. [Google Scholar] [CrossRef]
- Asato, A.E.; Matsumoto, H.; Denny, M.; Liu, R.S.H. Fluorinated rhodopsin analogs from 10-fluororetinal and 14-fluororetinal. J. Am. Chem. Soc. 1978, 100, 5957–5960. [Google Scholar] [CrossRef]
- de Grip, W.J. (Leiden University, Leiden, The Netherlands); Bovee-Geurts, P.H.M. (Radboudumc, Nijmegen, The Netherlands); Wada, A. (Kobe Pharmaceutical University, Kobe, Japan); Lugtenburg, J. (Leiden University, Leiden, The Netherlands). Photosensitivity and Spectral Properties of 11-cis and 9-cis 3,4-Dehydrorhodopsin. Personal Communication. 2022. [Google Scholar]
- Azuma, M.; Azuma, K.; Kito, Y. Circular dichroism of visual pigment analogues containing 3-dehydroretinal and 5,6-epoxy-3-dehydroretinal as the chromophore. Biochim. Biophys. Acta 1973, 295, 520–527. [Google Scholar] [CrossRef]
- Chung, W.C.; Nanbu, S.; Ishida, T. Nonadiabatic ab initio dynamics of a model protonated Schiff base of 9-cis retinal. J. Phys. Chem. A 2010, 114, 8190–8201. [Google Scholar] [CrossRef]
- Popp, A.; Wolperdinger, M.; Hampp, N.; Bräuchle, C.; Oesterhelt, D. Photochemical conversion of the O-intermediate to 9-cis-retinal- containing products in bacteriorhodopsin films. Biophys. J. 1993, 65, 1449–1459. [Google Scholar] [CrossRef]
- Mei, G.X.; Cavini, C.M.; Mamaeva, N.; Wang, P.; DeGrip, W.J.; Rothschild, K.J. Optical switching between long-lived states of opsin transmembrane voltage sensors. Photochem. Photobiol. 2021, 97, 1001–1015. [Google Scholar] [CrossRef]
- Mori, A.; Yagasaki, J.; Homma, M.; Reissig, L.; Sudo, Y. Investigation of the chromophore binding cavity in the 11-cis acceptable microbial rhodopsin MR. Chem. Phys. 2013, 419, 23–29. [Google Scholar] [CrossRef]
- DeGrip, W.J.; Daemen, F.J.M.; Bonting, S.L. Isolation and purification of bovine rhodopsin. Methods Enzym. 1980, 67, 301–320. [Google Scholar]
- Yoshizawa, T.; Wald, G. Photochemistry of iodopsin. Nature 1967, 214, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Okano, T.; Fukada, Y.; Artamonov, I.D.; Yoshizawa, T. Purification of cone visual pigments from chicken retina. Biochemistry 1989, 28, 8848–8856. [Google Scholar] [CrossRef] [PubMed]
- Kropf, A. A New Detergent for the Study of Visual Pigments. Vis. Res. 1982, 22, 495–497. [Google Scholar] [CrossRef]
- Stubbs, G.W.; Smith, H.G.; Litman, B.J. Alkyl glucosides as effective solubilizing agents for bovine rhodopsin—A comparison with several commonly used detergents. Biochim. Biophys. Acta 1976, 426, 46–56. [Google Scholar] [CrossRef]
- DeGrip, W.J.; Bovee-Geurts, P.H.M. Synthesis and properties of alkylglucosides with mild detergent action: Improved synthesis and purification of β-1-octyl, -nonyl- and -decyl-glucose. Synthesis of β-1-undecylglucose and β-1-dodecylmaltose. Chem. Phys. Lipids 1979, 23, 321–335. [Google Scholar] [CrossRef]
- Oprian, D.D.; Molday, R.S.; Kaufman, R.J.; Khorana, H.G. Expression of a synthetic bovine rhodopsin gene in monkey kidney cells. Proc. Natl. Acad. Sci. USA 1987, 84, 8874–8878. [Google Scholar] [CrossRef]
- Nathans, J.; Weitz, C.J.; Agarwal, N.; Nir, I.; Papermaster, D.S. Production of bovine rhodopsin by mammalian cell lines expressing cloned c-DNA: Spectrophotometry and subcellular localization. Vis. Res. 1989, 29, 907–914. [Google Scholar] [CrossRef]
- Reeves, P.J.; Callewaert, N.; Contreras, R.; Khorana, H.G. Structure and function in rhodopsin: High-level expression of rhodopsin with restricted and homogeneous N-glycosylation by a tetracycline-inducible N-acetylglucosaminyltransferase I-negative HEK293S stable mammalian cell line. Proc. Natl. Acad. Sci. USA 2002, 99, 13419–13424. [Google Scholar] [CrossRef]
- Janssen, J.J.M.; VanDeVen, W.J.M.; VanGroningen-Luyben, W.A.H.M.; Roosien, J.; Vlak, J.M.; DeGrip, W.J. Synthesis of functional bovine opsin in insect cells under control of the baculovirus polyhedrin promotor. Mol. Biol. Rep. 1988, 13, 65–71. [Google Scholar] [CrossRef]
- Klaassen, C.H.W.; Bovee-Geurts, P.H.M.; DeCaluwé, G.L.J.; DeGrip, W.J. Large-scale production and purification of functional recombinant bovine rhodopsin using the baculovirus expression system. Biochem. J. 1999, 342, 293–300. [Google Scholar] [CrossRef]
- Groenendijk, G.W.T.; Jansen, P.A.A.; Daemen, F.J.M.; Bonting, S.L. Analysis of geometrically isomeric vitamin A compounds. Methods Enzym. 1980, 67, 203–220. [Google Scholar]
- Ganapathy, S.; Liu, R.S.H. Photoisomerization of sixteen isomers of retinal. Initial product distribution in direct and sensitized irradiation. Photochem. Photobiol. 1992, 56, 959–964. [Google Scholar] [CrossRef]
- Davies, W.I.L.; Hankins, M.W.; Foster, R.G. Vertebrate ancient opsin and melanopsin: Divergent irradiance detectors. Photochem. Photobiol. Sci. 2010, 9, 1444–1457. [Google Scholar] [CrossRef] [PubMed]
- Varma, N.; Mutt, E.; Mühle, J.; Panneels, V.; Terakita, A.; Deupi, X.; Nogly, P.; Schertler, G.F.X.; Lesca, E. Crystal structure of jumping spider rhodopsin-1 as a light sensitive GPCR. Proc. Natl. Acad. Sci. USA 2019, 116, 14547–14556. [Google Scholar] [CrossRef] [PubMed]
- Bailes, H.J.; Zhuang, L.-Y.; Lucas, R.J. Reproducible and sustained regulation of Gαs signalling using a metazoan opsin as an optogenetic tool. PLoS ONE 2012, 7, e30774. [Google Scholar] [CrossRef]
- Church, J.R.; Haugaard Olsen, J.M.; Schapiro, I. The Impact of Retinal Configuration on the Protein-Chromophore Interactions in Bistable Jumping Spider Rhodopsin-1. Molecules 2022, 27, 71. [Google Scholar] [CrossRef]
- Davies, W.I.L.; Tay, B.-H.; Zheng, L.; Danks, J.A.; Brenner, S.; Foster, R.G.; Collin, S.P.; Hankins, M.W.; Venkatesh, B.; Hunt, D.M. Evolution and functional characterisation of melanopsins in a deep-sea chimaera (Elephant shark, Callorhinchus milii). PLoS ONE 2012, 7, e51276. [Google Scholar] [CrossRef]
- Herrera-Hernández, M.G.; Ramon, E.; Lupala, C.S.; Tena-Campos, M.; Pérez, J.J.; Garriga, P. Flavonoid allosteric modulation of mutated visual rhodopsin associated with retinitis pigmentosa. Sci. Rep. 2017, 7, 11167. [Google Scholar] [CrossRef]
- Gragg, M.; Park, P.S.-H. Misfolded rhodopsin mutants display variable aggregation properties. Biochim. Biophys. Acta-Mol. Basis Dis. 2018, 1864, 2938–2948. [Google Scholar] [CrossRef]
- Pasqualetto, G.; Pileggi, E.; Schepelmann, M.; Varricchio, C.; Rozanowska, M.; Brancale, A.; Bassetto, M. Ligand-based rational design, synthesis and evaluation of novel potential chemical chaperones for opsin. Eur. J. Med. Chem. 2021, 226, 113841. [Google Scholar] [CrossRef]
- Zhang, T.; Cao, L.-H.; Kumar, S.; Enemchukwu, N.O.; Zhang, N.; Lambert, A.; Zhao, X.C.; Jones, A.; Wang, S.X.; Dennis, E.M.; et al. Dimerization of visual pigments in vivo. Proc. Natl. Acad. Sci. USA 2016, 113, 9093–9098. [Google Scholar] [CrossRef] [PubMed]
- Behnen, P.; Felline, A.; Comitato, A.; Di Salvo, M.T.; Raimondi, F.; Gulati, S.; Kahremany, S.; Palczewski, K.; Marigo, V.; Fanelli, F. A Small Chaperone Improves Folding and Routing of Rhodopsin Mutants Linked to Inherited Blindness. iScience 2018, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.X.; Sun, W.; Kramp, K.; Zheng, M.; Salom, D.; Jastrzebska, B.; Jin, H.; Palczewski, K.; Feng, Z.Y. Light-sensitive coupling of rhodopsin and melanopsin to Gi/o and Gq signal transduction in Caenorhabditis elegans. FASEB J. 2012, 26, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Chen, Y.; Jastrzebska, B.; Golczak, M.; Gulati, S.; Tang, H.; Seibel, W.; Li, X.Y.; Jin, H.; Han, Y.; et al. A novel small molecule chaperone of rod opsin and its potential therapy for retinal degeneration. Nat. Commun. 2018, 9, 1976. [Google Scholar] [CrossRef]
- Kaya, K.D.; Chen, H.Y.; Brooks, M.J.; Kelley, R.A.; Shimada, H.; Nagashima, K.; de Val, N.; Drinnan, C.T.; Gieser, L.; Kruczek, K.; et al. Transcriptome-based molecular staging of human stem cell-derived retinal organoids uncovers accelerated photoreceptor differentiation by 9-cis retinal. Mol. Vis. 2019, 25, 663–678. [Google Scholar]
- Ohgane, K.; Dodo, K.; Hashimoto, Y. Structural development study of a novel pharmacological chaperone for folding-defective rhodopsin mutants responsible for retinitis pigmentosa. Yakugaku Zasshi 2011, 131, 325–334. [Google Scholar] [CrossRef]
- Kiser, P.D.; Palczewski, K. Retinoids and Retinal Diseases. Annu. Rev. Vis. Sci. 2016, 2, 197–234. [Google Scholar] [CrossRef]
- Sullivan, L.S.; Daiger, S.P. Inherited retinal degeneration: Exceptional genetic and clinical heterogeneity. Mol. Med. Today 1996, 2, 380–386. [Google Scholar] [CrossRef]
- Rattner, A.; Sun, H.; Nathans, J. Molecular genetics of human retinal disease. Annu. Rev. Genet. 1999, 33, 89–131. [Google Scholar] [CrossRef]
- Fan, J.; Crouch, R.K.; Kono, M. Light prevents exogenous 11-cis retinal from maintaining cone photoreceptors in chromophore-deficient mice. Invest. Ophthalmol. Vis. Sci. 2011, 52, 2412–2416. [Google Scholar] [CrossRef][Green Version]
- VanHooser, J.P.; Alemán, T.S.; He, Y.-G.; Cideciyan, A.V.; Kuksa, V.; Pittler, S.J.; Stone, E.M.; Jacobson, S.G.; Palczewski, K. Rapid restoration of visual pigment and function with oral retinoid in a mouse model of childhood blindness. Proc. Natl. Acad. Sci. USA 2000, 97, 8623–8628. [Google Scholar] [CrossRef] [PubMed]
- VanHooser, J.P.; Liang, Y.; Maeda, T.; Kuksa, V.; Jang, G.-F.; He, Y.-G.; Rieke, F.; Fong, H.K.W.; Detwiler, P.B.; Palczewski, K. Recovery of visual functions in a mouse model of Leber congenital amaurosis. J. Biol. Chem. 2002, 277, 19173–19182. [Google Scholar] [CrossRef] [PubMed]
- Maeda, A.; Maeda, T.; Palczewski, K. Improvement in rod and cone function in mouse model of Fundus albipunctatus after pharmacologic treatment with 9-cis-retinal. Invest. Ophthalmol. Vis. Sci. 2006, 47, 4540–4546. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Dong, Z.Q.; Jin, H.; Sawada, O.; Gao, S.Q.; Utkhede, D.; Monk, W.; Palczewska, G.; Palczewski, K. QLT091001, a 9-cis-retinal analog, is well-tolerated by retinas of mice with impaired visual cycles. Invest. Ophthalmol. Vis. Sci. 2013, 54, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Maeda, A.; Leahy, P.; Saperstein, D.A.; Palczewski, K. Effects of long-term administration of 9-cis-retinyl acetate on visual function in mice. Invest. Ophthalmol. Vis. Sci. 2009, 50, 322–333. [Google Scholar] [CrossRef]
- Scholl, H.P.N.; Moore, A.T.; Koenekoop, R.K.; Wen, Y.Q.; Fishman, G.A.; van den Born, L.I.; Bittner, A.; Bowles, K.; Fletcher, E.C.; Collison, F.T.; et al. Safety and Proof-of-Concept Study of Oral QLT091001 in Retinitis Pigmentosa Due to Inherited Deficiencies of Retinal Pigment Epithelial 65 Protein (RPE65) or Lecithin: Retinol Acyltransferase (LRAT). PLoS ONE 2015, 10, e0143846. [Google Scholar] [CrossRef]
- Gao, S.Q.; Kahremany, S.; Zhang, J.Y.; Jastrzebska, B.; Querubin, J.; Petersen-Jones, S.M.; Palczewski, K. Retinal-chitosan Conjugates Effectively Deliver Active Chromophores to Retinal Photoreceptor Cells in Blind Mice and Dogs. Mol. Pharm. 2018, 93, 438–452. [Google Scholar] [CrossRef]
- Dowling, J.E. Vitamin A: Its many roles—From vision and synaptic plasticity to infant mortality. J. Comp. Physiol. A 2020, 206, 389–399. [Google Scholar] [CrossRef]
- Sokolov, M.; Yadav, R.P.; Brooks, C.; Artemyev, N.O. Chaperones and retinal disorders. In Molecular Chaperones in Human Disorders; Donev, R., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 85–117. [Google Scholar]
- Veritti, D.; Sarao, V.; Samassa, F.; Danese, C.; Lowenstein, A.; Schmidt-Erfurth, U.; Lanzetta, P. State-of-the art pharmacotherapy for non-neovascular age-related macular degeneration. Expert Opin. Pharm. 2020, 21, 773–784. [Google Scholar] [CrossRef]
- Gao, S.Q.; Parmar, T.; Palczewska, G.; Dong, Z.Q.; Golczak, M.; Palczewski, K.; Jastrzebska, B. Protective Effect of a Locked Retinal Chromophore Analog against Light-Induced Retinal Degeneration. Mol. Pharm. 2018, 94, 1132–1144. [Google Scholar] [CrossRef]
- Hirano, T.; Lim, I.T.; Kim, D.M.; Zheng, X.-G.; Yoshihara, K.; Oyama, Y.; Imai, H.; Shichida, Y.; Ishiguro, M. Constraints of opsin structure on the ligand-binding site: Studies with ring-fused retinals. Photochem. Photobiol. 2002, 76, 606–615. [Google Scholar] [CrossRef]
- Yun, J.-H.; Li, X.X.; Park, J.-H.; Wang, Y.; Ohki, M.; Jin, Z.Y.; Lee, W.; Park, S.-Y.; Hu, H.; Li, C.F.; et al. Non-cryogenic structure of a chloride pump provides crucial clues to temperature-dependent channel transport efficiency. J. Biol. Chem. 2019, 294, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Gui, M.; Wang, Z.-F.; Gorgulla, C.; Yu, J.J.; Wu, H.; Sun, Z.-Y.J.; Klenk, C.; Merklinger, L.; Morstein, L.; et al. Cryo-EM structure of an activated GPCR-G protein complex in lipid nanodiscs. Nat. Struct. Mol. Biol. 2021, 28, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, I.R.; Pei, J.M.; Baek, M.; Krishnakumar, A.; Anishchenko, I.; Ovchinnikov, S.; Zhang, J.; Ness, T.J.; Banjade, S.; Bagde, S.R.; et al. Computed structures of core eukaryotic protein complexes. Science 2021, 374, 1340. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Pedraza-González, L.; Barneschi, L.; Padula, D.; De Vico, L.; Olivucci, M. Evolution of the Automatic Rhodopsin Modeling (ARM) Protocol. Top Curr. Chem. 2022, 380, 21. [Google Scholar] [CrossRef] [PubMed]
- El-Tahawy, M.M.T.; Conti, I.; Bonfanti, M.; Nenov, A.; Garavelli, M. Tailoring Spectral and Photochemical Properties of Bioinspired Retinal Mimics by in Silico Engineering. Angew. Chem. Int. Ed. 2020, 59, 20619–20627. [Google Scholar] [CrossRef]
- Tan, P.; He, L.; Huang, Y.; Zhou, Y. Optophysiology: Illuminating cell physiology with optogenetics. Physiol. Rev. 2022, 102, 1263–1325. [Google Scholar] [CrossRef]
- Gilhooley, M.J.; Lindner, M.; Palumaa, T.; Hughes, S.; Peirson, S.N.; Hankins, M.W. A systematic comparison of optogenetic approaches to visual restoration. Mol. Ther. Methods Clin. Dev. 2022, 25, 111–123. [Google Scholar] [CrossRef]


| Retinal | 11-cis Pigments | 9-cis Pigments | References | |||
|---|---|---|---|---|---|---|
| Modification | Chemical Structure * | HOOP Frequency (cm−1) # | HOOP Frequency (cm−1) # | |||
| 7=8 | 11=12 | 7=8 | 11=12 | |||
| - |  | u | 969 | 959 | 969 (w) | [48,71,78,105,108,144,149,150,151,152] |
| 7,8-Dihydro |  | - | 971 | - | 967 (w) | [56,105,134] |
| 13-Desmethyl |  | u | 975 (w) | n.a. | n.a. | [64,153] |
| 10-Methyl |  | u | 954 (s) | n.a. | n.a. | [130] |
| 9-Cyclopropyl |  | 977 (w) | 988 (w) | u | 974 | [105,154] |
| 3,4-Dehydro (retinal A2) |  | n.a. | n.a. | 960 (s) | 970 (w) | [155] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Grip, W.J.; Lugtenburg, J. Isorhodopsin: An Undervalued Visual Pigment Analog. Colorants 2022, 1, 256-279. https://doi.org/10.3390/colorants1030016
de Grip WJ, Lugtenburg J. Isorhodopsin: An Undervalued Visual Pigment Analog. Colorants. 2022; 1(3):256-279. https://doi.org/10.3390/colorants1030016
Chicago/Turabian Stylede Grip, Willem J., and Johan Lugtenburg. 2022. "Isorhodopsin: An Undervalued Visual Pigment Analog" Colorants 1, no. 3: 256-279. https://doi.org/10.3390/colorants1030016
APA Stylede Grip, W. J., & Lugtenburg, J. (2022). Isorhodopsin: An Undervalued Visual Pigment Analog. Colorants, 1(3), 256-279. https://doi.org/10.3390/colorants1030016