Bioinformatic Analysis of 1000 Amphibian Antimicrobial Peptides Uncovers Multiple Length-Dependent Correlations for Peptide Design and Prediction


Abstract
1. Introduction
2. Results
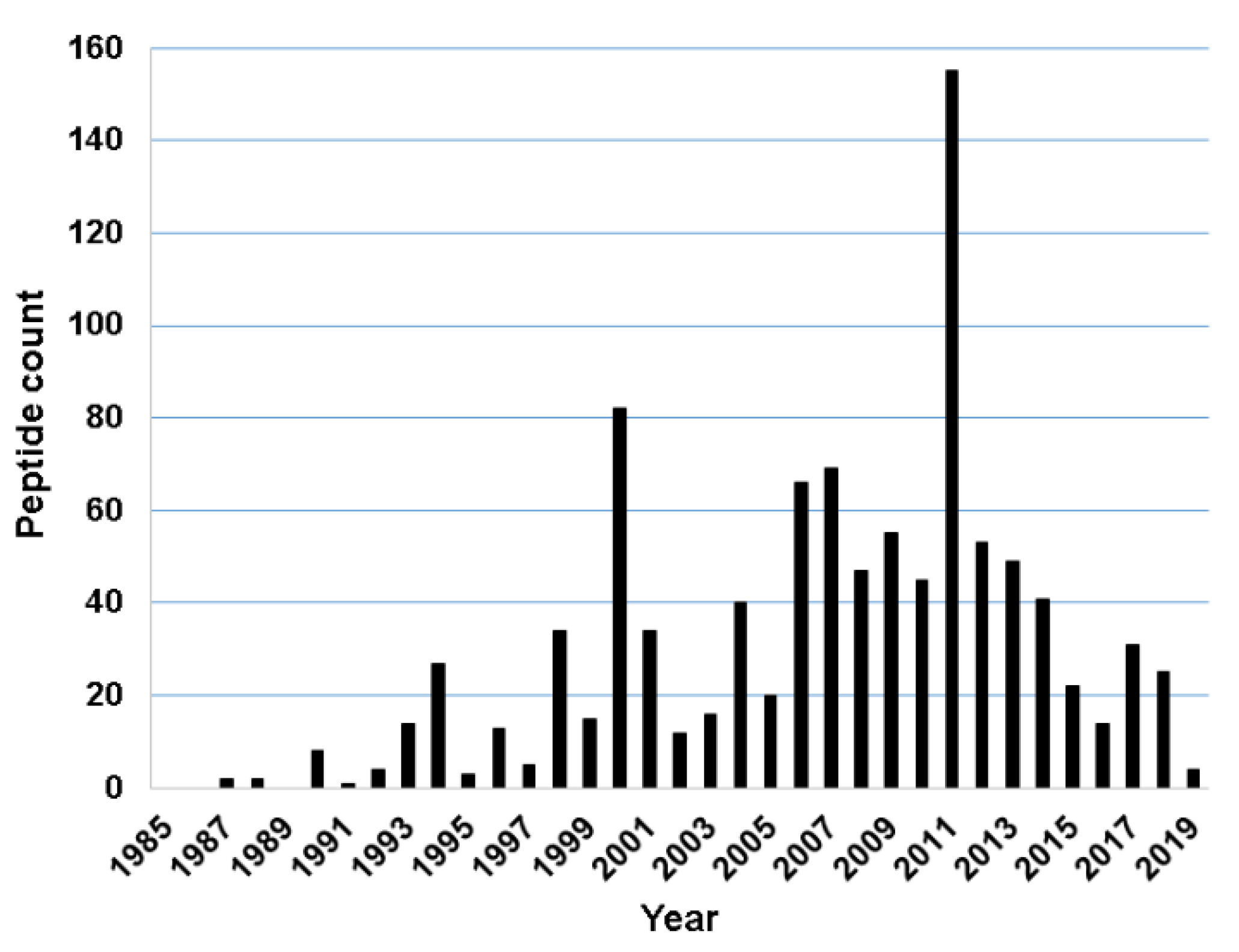
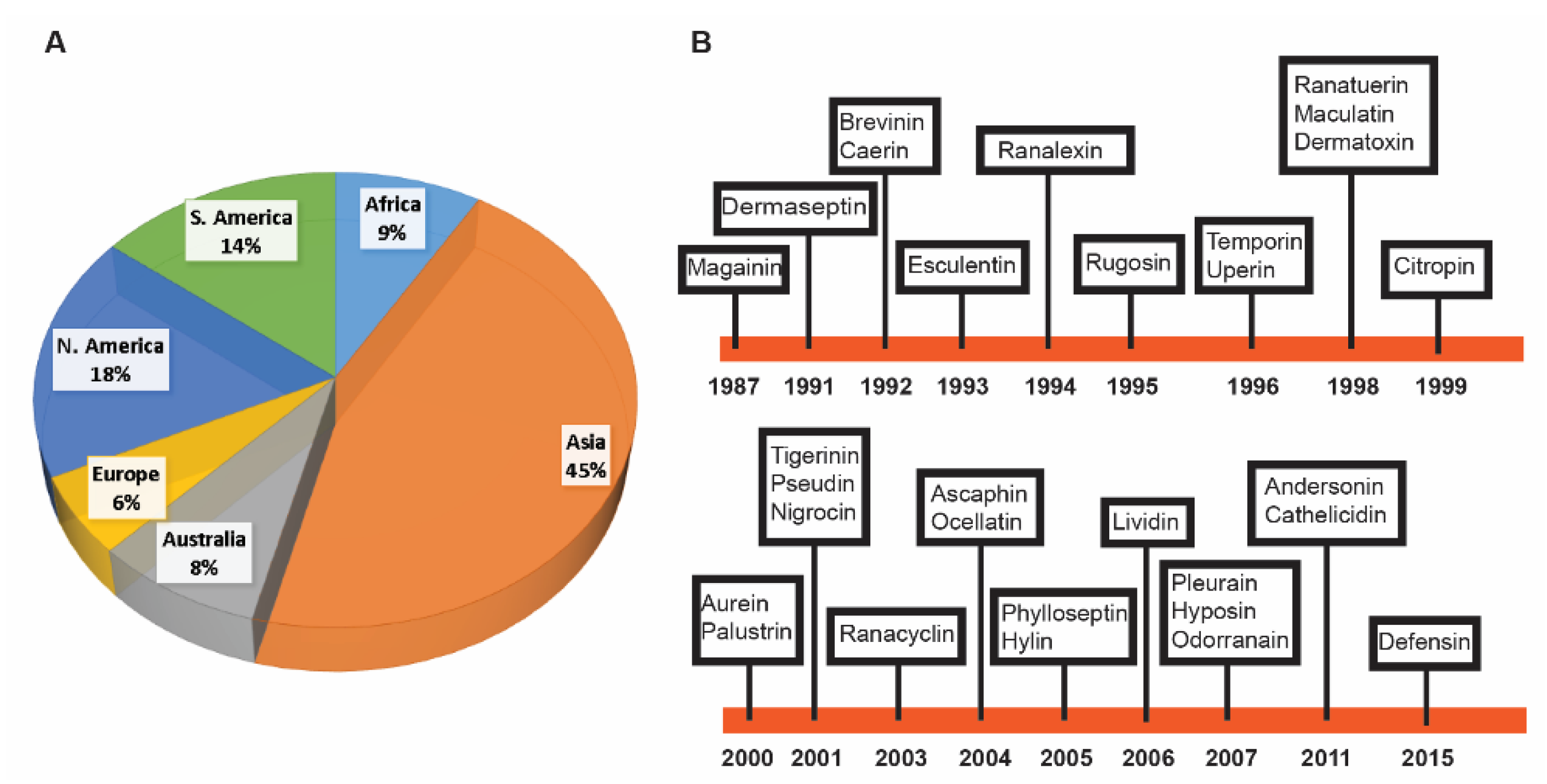
2.1. Discovery Timeline of Frog Antimicrobial Peptides
2.2. Major Families of Frog Peptides
2.3. Biological Activity of Amphibian Peptides
2.4. Toxicity of Amphibian Peptides
2.5. Peptide Sequence Signatures Modulate Activity Spectrum of Frog Host Defense Peptides
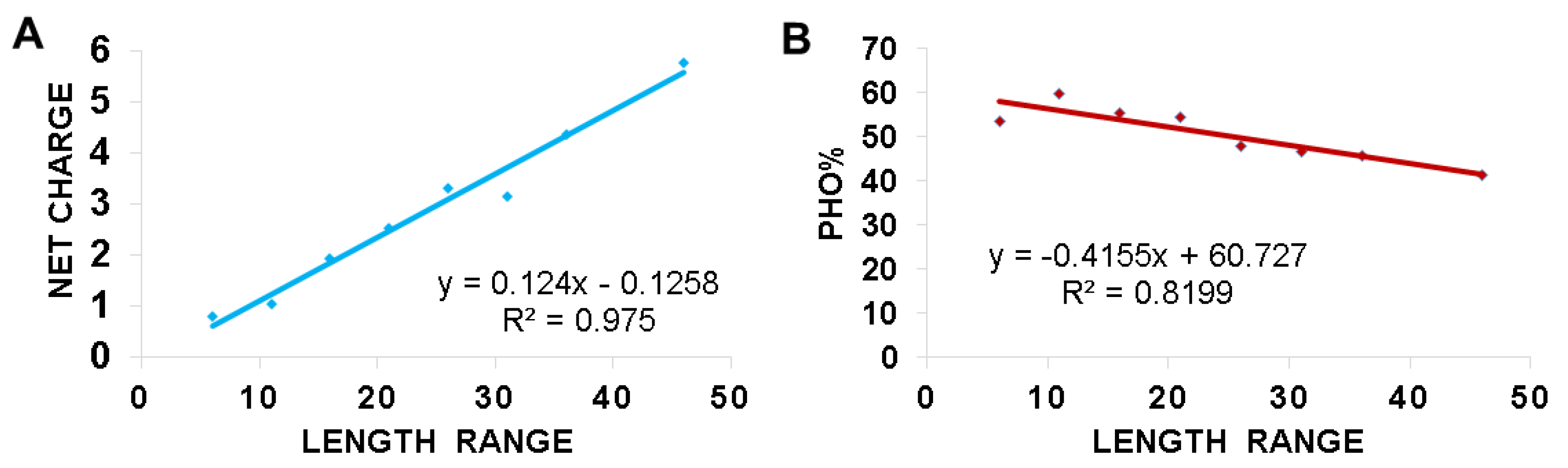
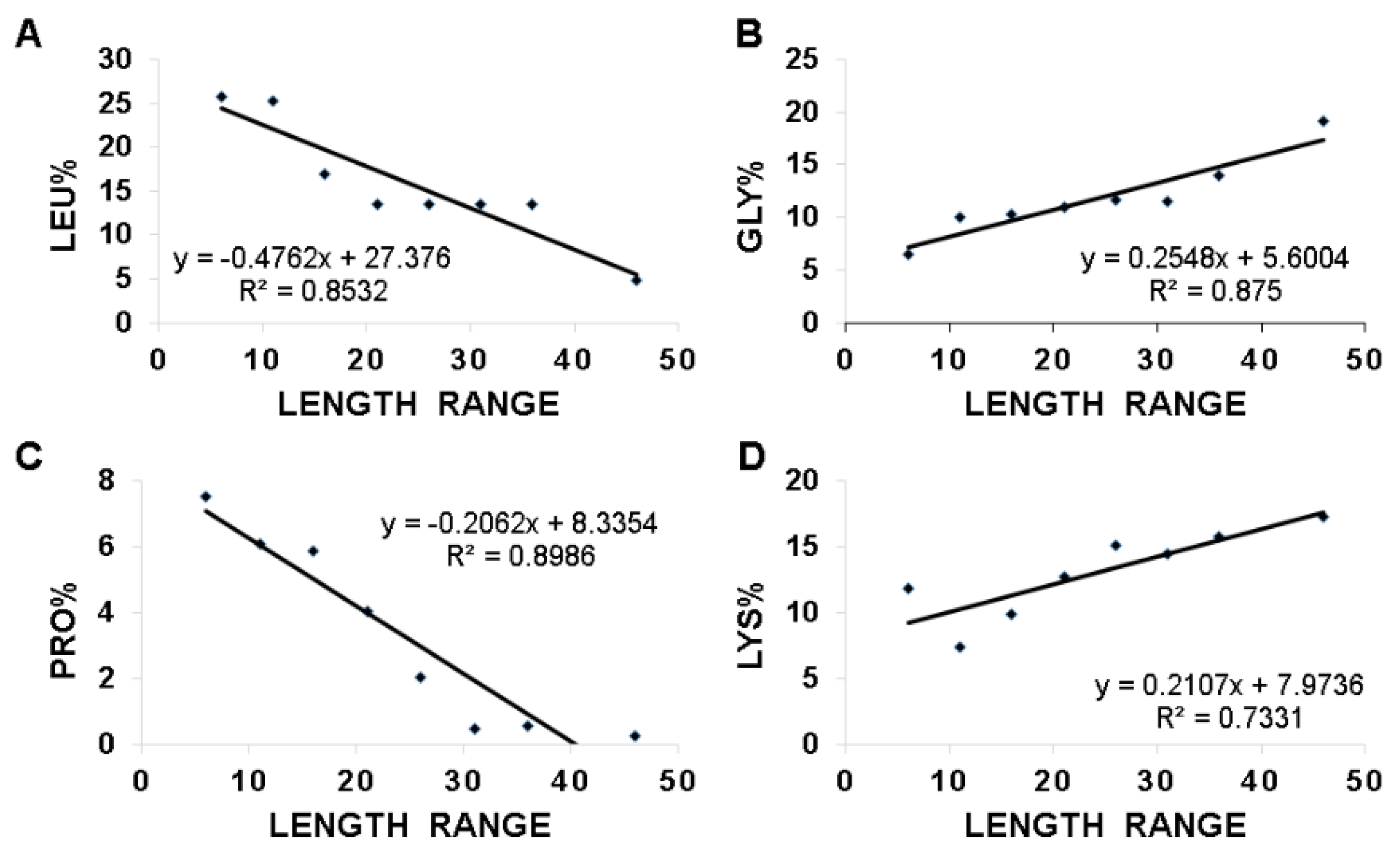
2.6. Length-Dependent Correlations in Frog Antimicrobial Peptides
3. Discussion
3.1. Peptide Length
3.2. The Ratio of Hydrophobic and Basic Amino Acids Determines Peptide Activity spEctrum
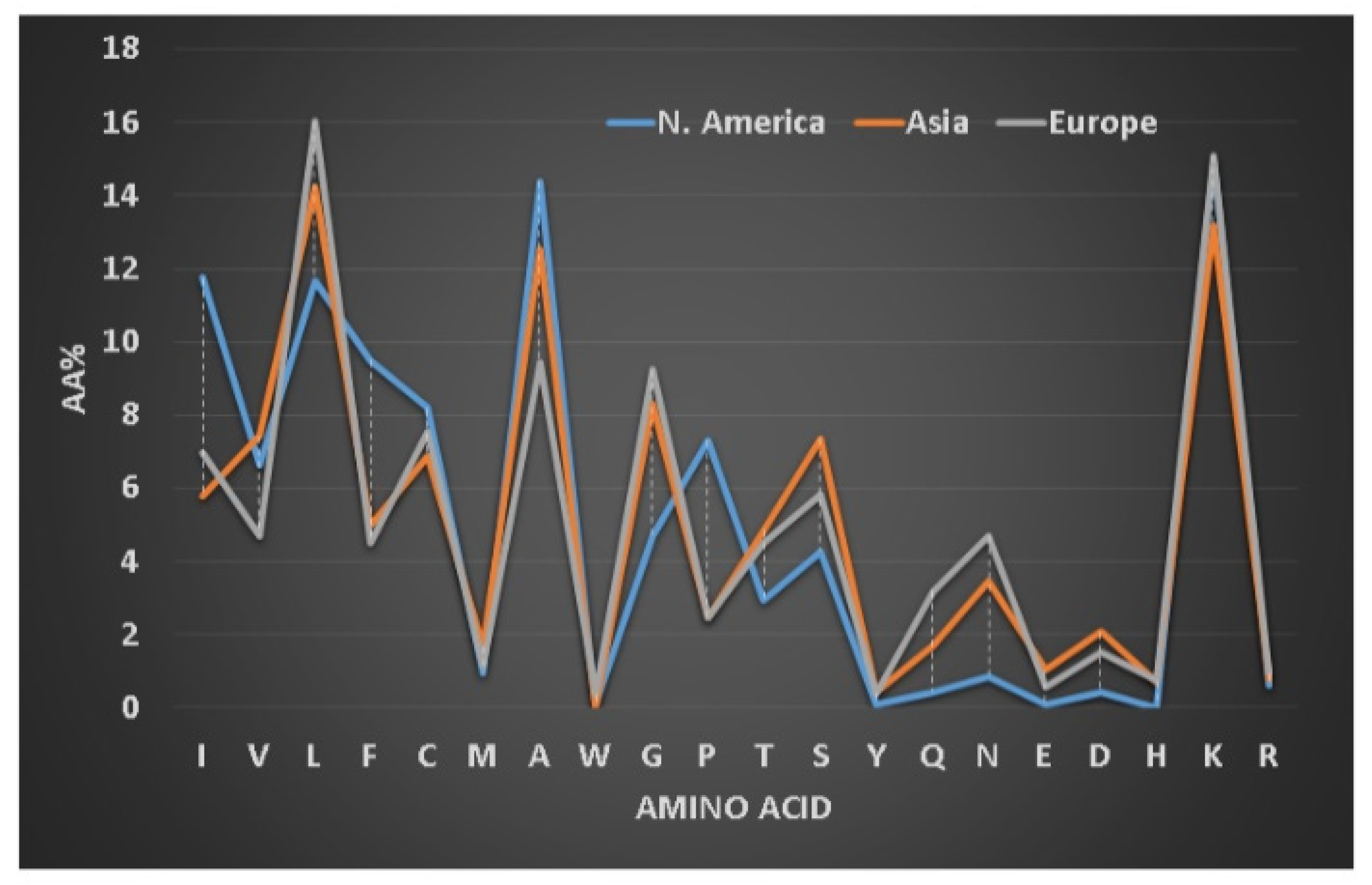
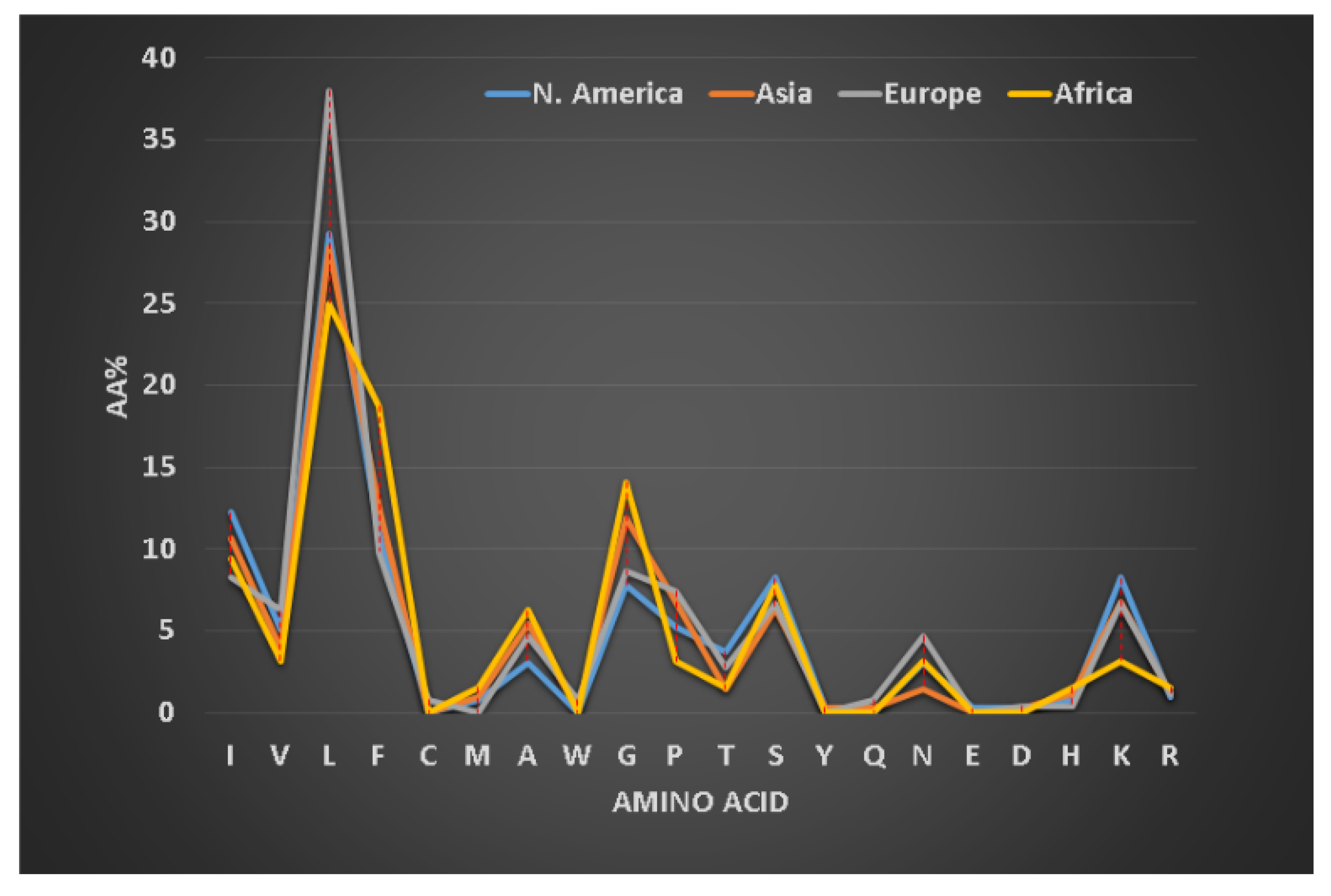
3.3. Hydrophobic and Basic Amino Acids Change with Peptide Length
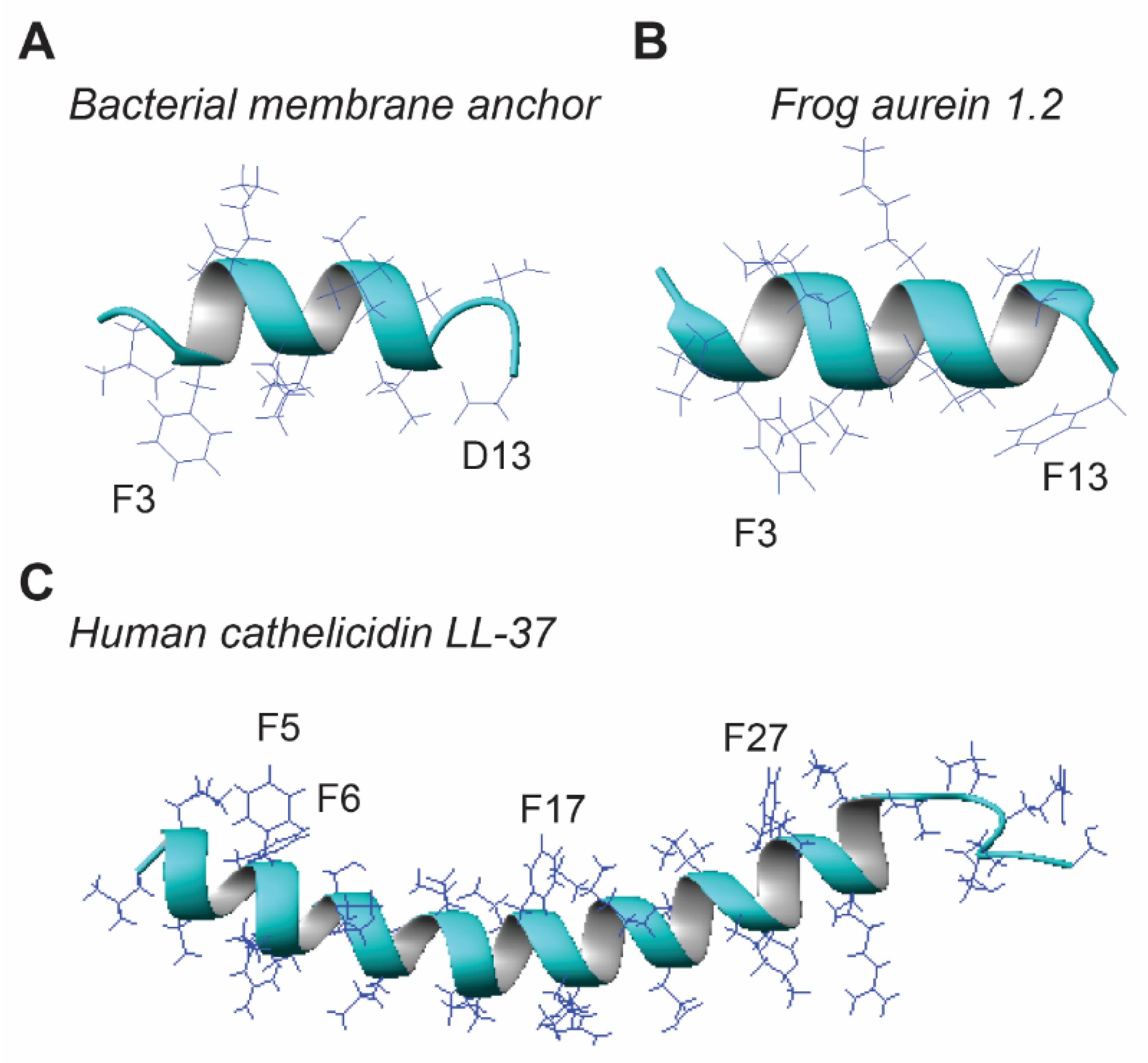
3.4. Post-Translational Modification Improves Helix Stability and Activity
3.5. Peptide Design
3.6. Peptide Mimicries
4. Methods: Database, Data Collection Criteria and Analysis
4.1. Database Versions and Update
4.2. Activity and Structural Informational Annotations
4.3. Peptide Classification
4.4. Data Registration Criteria and Analysis
5. Conclusions
Funding
Conflicts of Interest
References
- Pabijan, M.; Palomar, G.; Antunes, B.; Antoł, W.; Zieliński, P.; Babik, W. Evolutionary principles guiding amphibian conservation. Evol. Appl. 2020, 13, 857–878. [Google Scholar] [CrossRef]
- Nolan, M.W.; Smith, S.A. Amphibian resources on the internet. ILAR J. 2007, 48, 290–296. [Google Scholar] [CrossRef][Green Version]
- O’Rourke, D.P. Amphibians used in research and teaching. ILAR J. 2007, 48, 183–187. [Google Scholar] [CrossRef][Green Version]
- Bevins, C.L.; Zasloff, M. Peptides from frog skin. Annu. Rev. Biochem. 1990, 59, 395–414. [Google Scholar] [CrossRef]
- Bartels, E.J.H.; Dekker, D.; Amiche, M. Dermaseptins, Multifunctional Antimicrobial Peptides: A Review of Their Pharmacology, Effectivity, Mechanism of Action, and Possible Future Directions. Front. Pharmacol. 2019, 10, 1421. [Google Scholar] [CrossRef]
- Mangoni, M.L. Temporins, anti-infective peptides with expanding properties. Cell Mol. Life Sci. 2006, 63, 1060–1069. [Google Scholar] [CrossRef]
- Apponyi, M.A.; Pukala, T.L.; Brinkworth, C.S.; Maselli, V.M.; Bowie, J.H.; Tyler, M.J.; Booker, G.W.; Wallace, J.C.; Carver, J.A.; Separovic, F.; et al. Host-defence peptides of Australian anurans: Structure, mechanism of action and evolutionary significance. Peptides 2004, 25, 1035–1054. [Google Scholar] [CrossRef]
- Xu, X.; Lai, R. The chemistry and biological activities of peptides from amphibian skin secretions. Chem. Rev. 2015, 115, 1760–1846. [Google Scholar] [CrossRef]
- Ladram, A.; Nicolas, P. Antimicrobial peptides from frog skin: Biodiversity and therapeutic promises. Front. Biosci. 2016, 21, 1341–1371. [Google Scholar] [CrossRef]
- Rinaldi, A.C. Antimicrobial peptides from amphibian skin: An expanding scenario. Curr. Opin. Chem. Biol. 2002, 6, 799–804. [Google Scholar] [CrossRef]
- Conlon, J.M. Structural diversity and species distribution of host-defense peptides in frog skin secretions. Cell Mol. Life Sci. 2011, 68, 2303–2315. [Google Scholar] [CrossRef]
- Simmaco, M.; Kreil, G.; Barra, D. Bombinins, antimicrobial peptides from Bombina species. Biochim. Biophys. Acta 2009, 1788, 1551–1555. [Google Scholar] [CrossRef]
- Gibson, B.W.; Poulter, L.; Williams, D.H. A mass spectrometric assay for novel peptides: Application to Xenopus laevis skin secretions. Peptides 1985, 6 (Suppl. 3), 23–27. [Google Scholar] [CrossRef]
- Soravia, E.; Martini, G.; Zasloff, M. Antimicrobial properties of peptides from Xenopus granular gland secretions. FEBS Lett. 1988, 228, 337–340. [Google Scholar] [CrossRef]
- Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus skin: Isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc. Natl. Acad. Sci. USA 1987, 84, 5449–5453. [Google Scholar] [CrossRef]
- Mor, A.; Nguyen, V.H.; Delfour, A.; Migliore-Samour, D.; Nicolas, P. Isolation, amino acid sequence, and synthesis of dermaseptin, a novel antimicrobial peptide of amphibian skin. Biochemistry 1991, 30, 8824–8830. [Google Scholar] [CrossRef]
- Morikawa, N.; Hagiwara, K.; Nakajima, T. Brevinin-1 and -2, unique antimicrobial peptides from the skin of the frog, Rana brevipoda porsa. Biochem. Biophys. Res. Commun. 1992, 189, 184–190. [Google Scholar] [CrossRef]
- Stone, D.J.M.; Waugh, R.J.; Bowie, J.H.; Wallace, J.C.; Tyler, M.J. Peptides from Australian frogs. Structures of the caerins and caeridin 1 from Litoria splendida. J. Chem. Soc. Perkin Trans. 1992, 1, 3173–3178. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Barra, D.; Bossa, F. Novel antimicrobial peptides from skin secretion of the European frog Rana esculenta. FEBS Lett. 1993, 324, 159–161. [Google Scholar] [CrossRef]
- Clark, D.P.; Durell, S.; Maloy, W.L.; Zasloff, M. Ranalexin, a novel antimicrobial peptide from bullfrog (Rana catesbeiana) skin, structurally related to the bacterial antibiotic, polymyxin. J. Biol. Chem. 1994, 269, 10849–10855. [Google Scholar]
- Suzuki, S.; Ohe, Y.; Okubo, T.; Kakegawa, T.; Tatemoto, K. Isolation and characterization of novel antimicrobial peptides, rugosins A, B and C, from the skin of the frog, Rana rugosa. Biochem. Biophys. Res. Commun. 1995, 212, 249–254. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Canofeni, S.; Miele, R.; Mangoni, M.L.; Barra, D. Temporins, antimicrobial peptides from the European red frog Rana temporaria. Eur. J. Biochem. 1996, 242, 788–792. [Google Scholar] [CrossRef]
- Bradford, A.M.; Raftery, M.J.; Bowie, J.H.; Tyler, M.J.; Wallace, J.C.; Adams, G.W.; Severini, C. Novel uperin peptides from the dorsal glands of the australian floodplain toadlet Uperoleia inundata. Aust. J. Chem. 1996, 49, 475–484. [Google Scholar] [CrossRef]
- Goraya, J.; Knoop, F.C.; Conlon, J.M. Ranatuerins: Antimicrobial peptides isolated from the skin of the American bullfrog, Rana catesbeiana. Biochem. Biophys. Res. Commun. 1998, 250, 589–592. [Google Scholar] [CrossRef]
- Rozek, T.; Waugh, R.J.; Steinborner, S.T.; Bowie, J.H.; Tyler, M.J.; Wallace, J.C. The maculatin peptides from the skin glands of the tree frog Litoria genimaculata: A comparison of the structures and antibacterial activities of maculatin 1.1 and caerin 1.1. J. Pept. Sci. 1998, 4, 111–115. [Google Scholar] [CrossRef]
- Wechselberger, C. Cloning of cDNAs encoding new peptides of the dermaseptin-family. Biochim. Biophys. Acta 1998, 1388, 279–283. [Google Scholar] [CrossRef]
- Wegener, K.L.; Wabnitz, P.A.; Carver, J.A.; Bowie, J.H.; Chia, B.C.; Wallace, J.C.; Tyler, M.J. Host defence peptides from the skin glands of the Australian blue mountains tree-frog Litoria citropa. Solution structure of the antibacterial peptide citropin 1.1. Eur. J. Biochem. 1999, 265, 627–637. [Google Scholar] [CrossRef]
- Rozek, T.; Wegener, K.L.; Bowie, J.H.; Olver, I.N.; Carver, J.A.; Wallace, J.C.; Tyler, M.J. The antibiotic and anticancer active aurein peptides from the Australian Bell Frogs Litoria aurea and Litoria raniformis the solution structure of aurein 1.2. Eur. J. Biochem. 2000, 267, 5330–5341. [Google Scholar] [CrossRef]
- Pierre, T.N.; Seon, A.A.; Amiche, M.; Nicolas, P. Phylloxin, a novel peptide antibiotic of the dermaseptin family of antimicrobial/opioid peptide precursors. Eur. J. Biochem. 2000, 267, 370–378. [Google Scholar] [CrossRef]
- Basir, Y.J.; Knoop, F.C.; Dulka, J.; Conlon, J.M. Multiple antimicrobial peptides and peptides related to bradykinin and neuromedin N isolated from skin secretions of the pickerel frog, Rana palustris. Biochim. Biophys. Acta 2000, 1543, 95–105. [Google Scholar] [CrossRef]
- Mattute, B.; Knoop, F.C.; Conlon, J.M. Kassinatuerin-1: A peptide with broad-spectrum antimicrobial activity isolated from the skin of the hyperoliid frog, Kassina senegalensis. Biochem. Biophys. Res. Commun. 2000, 268, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Sai, K.P.; Jagannadham, M.V.; Vairamani, M.; Raju, N.P.; Devi, A.S.; Nagaraj, R.; Sitaram, N. Tigerinins: Novel antimicrobial peptides from the Indian frog Rana tigerina. J. Biol. Chem. 2001, 276, 2701–2707. [Google Scholar] [CrossRef]
- Olson, L., III; Soto, A.M.; Knoop, F.C.; Conlon, J.M. Pseudin-2: An antimicrobial peptide with low hemolytic activity from the skin of the paradoxical frog. Biochem. Biophys. Res. Commun. 2001, 288, 1001–1005. [Google Scholar] [CrossRef]
- Batista, C.V.; Scaloni, A.; Rigden, D.J. A novel heterodimeric antimicrobial peptide from the tree-frog Phyllomedusa distincta. FEBS Lett. 2001, 494, 85–89. [Google Scholar] [CrossRef]
- Park, S.; Park, S.H.; Ahn, H.C.; Kim, S.; Kim, S.S.; Lee, B.J.; Lee, B.J. Structural study of novel antimicrobial peptides, nigrocins, isolated from Rana nigromaculata. FEBS Lett. 2001, 507, 95–100. [Google Scholar] [CrossRef]
- Wegener, K.L.; Brinkworth, C.S.; Bowie, J.H.; Wallace, J.C.; Tyler, M.J. Bioactive dahlein peptides from the skin secretions of the Australian aquatic frog Litoria dahlii: Sequence determination by electrospray mass spectrometry. Rapid Commun. Mass Spectrom. 2001, 15, 1726–1734. [Google Scholar] [CrossRef]
- Isaacson, T.; Soto, A.; Iwamuro, S.; Knoop, F.C.; Conlon, J.M. Antimicrobial peptides with atypical structural features from the skin of the Japanese brown frog Rana japonica. Peptides 2002, 23, 419–425. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Papo, N.; Mignogna, G.; Andreu, D.; Shai, Y.; Barra, D.; Simmaco, M. Ranacyclins, a new family of short cyclic antimicrobial peptides: Biological function, mode of action, and parameters involved in target specificity. Biochemistry 2003, 42, 14023–14035. [Google Scholar] [CrossRef]
- Nascimento, A.C.; Zanotta, L.C.; Kyaw, C.M.; Schwartz, E.N.; Schwartz, C.A.; Sebben, A.; Sousa, M.V.; Fontes, W.; Castro, M.S. Ocellatins: New antimicrobial peptides from the skin secretion of the South American frog Leptodactylus ocellatus (Anura: Leptodactylidae). Protein J. 2004, 23, 501–508. [Google Scholar] [CrossRef]
- Conlon, J.M.; Sonnevend, A.; Davidson, C.; Smith, D.D.; Nielsen, P.F. The ascaphins: A family of antimicrobial peptides from the skin secretions of the most primitive extant frog, Ascaphus truei. Biochem. Biophys. Res. Commun. 2004, 320, 170–175. [Google Scholar] [CrossRef]
- Leite, J.R.; Silva, L.P.; Rodrigues, M.I.; Prates, M.V.; Brand, G.D.; Lacava, B.M.; Azevedo, R.B.; Bocca, A.L.; Albuquerque, S.; Bloch, C., Jr. Phylloseptins: A novel class of anti-bacterial and anti-protozoan peptides from the Phyllomedusa genus. Peptides 2005, 26, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.S.; Matsushita, R.H.; Sebben, A.; Sousa, M.V.; Fontes, W. Hylins: Bombinins H structurally related peptides from the skin secretion of the Brazilian tree-frog Hyla biobeba. Protein Pept. Lett. 2005, 12, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Chen, T.; Walker, B.; Shaw, C. Lividins: Novel antimicrobial peptide homologs from the skin secretion of the Chinese Large Odorous frog, Rana (Odorrana) livida. Identification by “shotgun” cDNA cloning and sequence analysis. Peptides 2006, 27, 2118–2123. [Google Scholar] [CrossRef]
- Zhou, M.; Chen, T.; Walker, B.; Shaw, C. Pelophylaxins: Novel antimicrobial peptide homologs from the skin secretion of the Fukien gold-striped pond frog, Pelophylax plancyi fukienensis: Identification by “shotgun” cDNA cloning and sequence analysis. Peptides 2006, 27, 36–41. [Google Scholar] [CrossRef]
- Kim, S.S.; Shim, M.S.; Chung, J.; Lim, D.Y.; Lee, B.J. Purification and characterization of antimicrobial peptides from the skin secretion of Rana dybowskii. Peptides 2007, 28, 1532–1539. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, Y.; Li, J.; Liu, H.; Xu, X.; Lai, R.; Zhang, K. A new family of antimicrobial peptides from skin secretions of Rana pleuraden. Peptides 2007, 28, 2069–2074. [Google Scholar] [CrossRef]
- Chen, L.; Li, Y.; Li, J.; Xu, X.; Lai, R.; Zou, Q. An antimicrobial peptide with antimicrobial activity against Helicobacter pylori. Peptides 2007, 28, 1527–1531. [Google Scholar] [CrossRef]
- Thompson, A.H.; Bjourson, A.J.; Orr, D.F.; Shaw, C.; McClean, S. Amphibian skin secretomics: Application of parallel quadrupole time-of-flight mass spectrometry and peptide precursor cDNA cloning to rapidly characterize the skin secretory peptidome of Phyllomedusa hypochondrialis azurea: Discovery of a novel peptide family, the hyposins. J. Proteome Res. 2007, 6, 3604–3613. [Google Scholar]
- Jackway, R.J.; Bowie, J.H.; Bilusich, D.; Musgrave, I.F.; Surinya-Johnson, K.H.; Tyler, M.J.; Eichinger, P.C. The fallaxidin peptides from the skin secretion of the Eastern Dwarf Tree Frog Litoria fallax. Sequence determination by positive and negative ion electrospray mass spectrometry: Antimicrobial activity and cDNA cloning of the fallaxidins. Rapid Commun. Mass Spectrom. 2008, 22, 3207–3216. [Google Scholar] [CrossRef]
- Ma, Y.; Liu, C.; Liu, X.; Wu, J.; Yang, H.; Wang, Y.; Li, J.; Yu, H.; Lai, R. Peptidomics and genomics analysis of novel antimicrobial peptides from the frog, Rana nigrovittata. Genomics 2010, 95, 66–71. [Google Scholar] [CrossRef]
- Yang, X.; Lee, W.H.; Zhang, Y. Extremely Abundant Antimicrobial Peptides Existed in the Skins of Nine Kinds of Chinese Odorous Frogs. J. Proteome Res. 2012, 11, 306–319. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Yang, H.; Wei, L.; Yang, S.; Zhu, W.; Ma, D.; Yu, H.; Lai, R. Amphibian cathelicidin fills the evolutionary gap of cathelicidin in vertebrate. Amino Acids 2012, 43, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Mechkarska, M.; Prajeep, M.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; King, J.D.; Conlon, J.M. The hymenochirins: A family of host-defense peptides from the Congo dwarf clawed frog Hymenochirus boettgeri (Pipidae). Peptides 2012, 35, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Mechkarska, M.; Radosavljevic, G.; Attoub, S.; King, J.D.; Lukic, M.L.; McClean, S. A family of antimicrobial and immunomodulatory peptides related to the frenatins from skin secretions of the Orinoco lime frog Sphaenorhynchus lacteus (Hylidae). Peptides 2014, 56, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Che, H.; Han, Y.; Lv, J.; Mu, L.; Lv, L.; Wu, J.; Yang, H. The first anionic defensin from amphibians. Amino Acids 2015, 47, 1301–1308. [Google Scholar] [CrossRef] [PubMed]
- Boman, H.G. Antibacterial peptides: Basic facts and emerging concepts. J. Intern. Med. 2003, 254, 197–215. [Google Scholar] [CrossRef] [PubMed]
- Ferrandon, D.; Imler, J.L.; Hoffmann, J.A. Sensing infection in Drosophila: Toll and beyond. Semin. Immunol. 2004, 16, 43–53. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Mansour, S.C.; Pena, O.M.; Hancock, R.E. Host defense peptides: Front-line immunomodulators. Trends Immunol. 2014, 35, 443–450. [Google Scholar] [CrossRef]
- Merrifield, R.B.; Merrifield, E.L.; Juvvadi, P.; Andreu, D.; Boman, H.G. Design and synthesis of antimicrobial peptides. Ciba Found. Symp. 1994, 186, 5–26. [Google Scholar]
- Kline, A.D.; Wüthrich, K. Complete sequence-specific 1H nuclear magnetic resonance assignments for the alpha-amylase polypeptide inhibitor tendamistat from Streptomyces tendae. J. Mol. Biol. 1986, 192, 869–890. [Google Scholar] [CrossRef]
- Oschkinat, H.; Griesinger, C.; Kraulis, P.J.; Sorensen, O.W.; Ernst, R.R.; Gronenborn, A.M.; Clore, G.M. Three-dimensional NMR spectroscopy of a protein in solution. Nature 1988, 332, 374–376. [Google Scholar] [CrossRef] [PubMed]
- Uggerhøj, L.E.; Poulsen, T.J.; Munk, J.K.; Fredborg, M.; Sondergaard, T.E.; Frimodt-Moller, N.; Hansen, P.R.; Wimmer, R. Rational design of alpha-helical antimicrobial peptides: Do’s and don’ts. Chembiochem 2015, 16, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Hunter, H.N.; Matsuzaki, K.; Vogel, H.J. Solution NMR studies of amphibian antimicrobial peptides: Linking structure to function? Biochim. Biophys. Acta 2009, 1788, 1639–1655. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, Y.; Li, X. Correlation of three-dimensional structures with the antibacterial activity of a group of peptides designed based on a nontoxic bacterial membrane anchor. J. Biol. Chem. 2005, 280, 5803–5811. [Google Scholar] [CrossRef]
- Duclohier, H.; Molle, G.; Dugast, J.Y.; Spach, G. Prolines are not essential residues in the “barrel-stave” model for ion channels induced by alamethicin analogues. Biophys. J. 1992, 63, 868–873. [Google Scholar] [CrossRef]
- Ketchem, R.R.; Hu, W.; Cross, T.A. High-resolution conformation of gramicidin A in a lipid bilayer by solid-state NMR. Science 1993, 261, 1457–1460. [Google Scholar] [CrossRef]
- Shai, Y. Mode of action of membrane active antimicrobial peptides. Biopolymers 2002, 66, 236–248. [Google Scholar] [CrossRef]
- Bechinger, B.; Lohner, K. Detergent-like actions of linear amphipathic cationic antimicrobial peptides. Biochim. Biophys. Acta 2006, 1758, 1529–1539. [Google Scholar] [CrossRef]
- Wimley, W.C. Describing the mechanism of antimicrobial peptide action with the interfacial activity model. ACS Chem. Biol. 2010, 5, 905–917. [Google Scholar] [CrossRef]
- Epand, R.M. Anionic Lipid Clustering Model. Adv. Exp. Med. Biol. 2019, 1117, 65–71. [Google Scholar]
- Grage, S.L.; Afonin, S.; Kara, S.; Buth, G.; Ulrich, A.S. Membrane Thinning and Thickening Induced by Membrane-Active Amphipathic Peptides. Front. Cell Dev. Biol. 2016, 4, 65. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K. Membrane Permeabilization Mechanisms. Adv. Exp. Med. Biol. 2019, 1117, 9–16. [Google Scholar]
- Ludtke, S.J.; He, K.; Heller, W.T.; Harroun, T.A.; Yang, L.; Huang, H.W. Membrane pores induced by magainin. Biochemistry 1996, 35, 13723–13728. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Epand, R.F.; Mishra, B.; Lushnikova, T.; Thomas, V.C.; Bayles, K.W.; Epand, R.M. Decoding the functional roles of cationic side chains of the major antimicrobial region of human cathelicidin LL-37. Antimicrob. Agents Chemother. 2012, 56, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Patrzykat, A.; Friedrich, C.L.; Zhang, L.; Mendoza, V.; Hancock, R.E. Sublethal concentrations of pleurocidin-derived antimicrobial peptides inhibit macromolecular synthesis in Escherichia coli. Antimicrob. Agents Chemother. 2002, 46, 605–614. [Google Scholar] [CrossRef]
- Verjans, E.T.; Zels, S.; Luyten, W.; Landuyt, B.; Schoofs, L. Molecular mechanisms of LL-37-induced receptor activation: An overview. Peptides 2016, 85, 16–26. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD2: The updated antimicrobial peptide database and its application in peptide design. Nucleic Acids Res. 2009, 37, D933–D937. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, G. APD: The Antimicrobial Peptide Database. Nucleic Acids Res. 2004, 32, D590–D592. [Google Scholar] [CrossRef]
- Halverson, T.; Basir, Y.J.; Knoop, F.C.; Conlon, J.M. Purification and characterization of antimicrobial peptides from the skin of the North American green frog Rana clamitans. Peptides 2000, 21, 469–476. [Google Scholar] [CrossRef]
- Goraya, J.; Wang, Y.; Li, Z.; O’Flaherty, M.; Knoop, F.C.; Platz, J.E.; Conlon, J.M. Peptides with antimicrobial activity from four different families isolated from the skins of the North American frogs Rana luteiventris, Rana berlandieri and Rana pipiens. Eur. J. Biochem. 2000, 267, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Matutte, B.; Storey, K.B.; Knoop, F.C.; Conlon, J.M. Induction of synthesis of an antimicrobial peptide in the skin of the freeze-tolerant frog, Rana sylvatica, in response to environmental stimuli. FEBS Lett. 2000, 483, 135–138. [Google Scholar] [CrossRef]
- Li, J.; Xu, X.; Xu, C.; Zhou, W.; Zhang, K.; Yu, H.; Zhang, Y.; Zheng, Y.; Rees, H.H.; Lai, R.; et al. Anti-infection peptidomics of amphibian skin. Mol Cell Proteom. 2007, 6, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Proaño-Bolaños, C.; Zhou, M.; Wang, L.; Coloma, L.A.; Chen, T.; Shaw, C. Peptidomic approach identifies cruzioseptins, a new family of potent antimicrobial peptides in the splendid leaf frog Cruziohyla calcarifer. J. Proteom. 2016, 146, 1–13. [Google Scholar] [CrossRef]
- Conlon, J.M.; Al-Ghafari, N.; Coquet, L.; Leprince, J.; Jouenne, T.; Vaudry, H.; Davidson, C. Evidence from peptidomic analysis of skin secretions that the red-legged frogs, Rana aurora draytonii and Rana aurora aurora, are distinct species. Peptides. 2006, 27, 1305–1312. [Google Scholar] [CrossRef]
- Conlon, J.M.; Mechkarska, M.; Leprince, J. Peptidomic analysis in the discovery of therapeutically valuable peptides in amphibian skin secretions. Expert Rev. Proteom. 2019, 16, 897–908. [Google Scholar] [CrossRef]
- Conlon, J.M. A proposed nomenclature for antimicrobial peptides from frogs of the genus Leptodactylus. Peptides 2008, 29, 1631–1632. [Google Scholar] [CrossRef]
- Lehrer, R.I.; Ganz, T. Defensins of vertebrate animals. Curr. Opin. Immunol. 2002, 14, 96–102. [Google Scholar] [CrossRef]
- Zanetti, M.; Gennaro, R.; Scocchi, M.; Skerlavaj, B. Structure and biology of cathelicidins. Adv. Exp. Med. Biol. 2000, 479, 203–218. [Google Scholar]
- Shi, Y.; Li, C.; Wang, M.; Chen, Z.; Luo, Y.; Xia, X.; Song, Y.; Sun, Y.; Zhang, A. Cathelicidin-DM is an antimicrobial peptide from Duttaphrynus melanostictus and Has Wound-Healing Therapeutic Potential. ACS Omega 2020, 5, 9301–9310. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L.; McDermott, A.M.; Zasloff, M. Antimicrobial peptides and wound healing: Biological and therapeutic considerations. Exp. Dermatol. 2016, 25, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Casciaro, B.; Moros, M.; Rivera-Fernández, S.; Bellelli, A.; de la Fuente, J.M.; Mangoni, M.L. Gold-nanoparticles coated with the antimicrobial peptide esculentin-1a(1–21)NH2 as a reliable strategy for antipseudomonal drugs. Acta Biomater. 2017, 47, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Hasim, S.; Coleman, J.J. Targeting the fungal cell wall: Current therapies and implications for development of alternative antifungal agents. Future Med. Chem. 2019, 11, 869–883. [Google Scholar] [CrossRef] [PubMed]
- Berger, L.; Speare, R.; Daszak, P.; Green, D.E.; Cunningham, A.A.; Goggin, C.L.; Slocombe, R.; Ragan, M.A.; Hyatt, A.D.; McDonald, K.R.; et al. Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America. Proc. Natl. Acad. Sci. USA 1998, 95, 9031–9036. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A. The role of amphibian antimicrobial peptides in protection of amphibians from pathogens linked to global amphibian declines. Biochim. Biophys. Acta 2009, 1788, 1593–1599. [Google Scholar] [CrossRef]
- Bondaryk, M.; Staniszewska, M.; Zielińska, P.; Urbańczyk-Lipkowska, Z. Natural Antimicrobial Peptides as Inspiration for Design of a New Generation Antifungal Compounds. J. Fungi. 2017, 3, 46. [Google Scholar] [CrossRef]
- Wang, G.; Narayana, J.L.; Mishra, B.; Zhang, Y.; Wang, F.; Wang, C.; Zarena, D.; Lushnikova, T.; Wang, X. Design of Antimicrobial Peptides: Progress Made with Human Cathelicidin LL-37. Adv. Exp. Med. Biol. 2019, 1117, 215–240. [Google Scholar]
- Vilas Boas, L.C.P.; Campos, M.L.; Berlanda, R.L.A.; de Carvalho Neves, N.; Franco, O.L. Antiviral peptides as promising therapeutic drugs. Cell Mol. Life Sci. 2019, 76, 3525–3542. [Google Scholar] [CrossRef]
- VanCompernolle, S.; Smith, P.B.; Bowie, J.H.; Tyler, M.J.; Unutmaz, D.; Rollins-Smith, L.A. Inhibition of HIV infection by caerin 1 antimicrobial peptides. Peptides 2015, 71, 296–303. [Google Scholar] [CrossRef]
- Wang, G. Natural antimicrobial peptides as promising anti-HIV candidates. Curr. Top. Pept. Protein Res. 2012, 13, 93–110. [Google Scholar] [PubMed]
- Wang, G.; Watson, K.M.; Peterkofsky, A.; Buckheit, R.W., Jr. Identification of novel human immunodeficiency virus type 1-inhibitory peptides based on the antimicrobial peptide database. Antimicrob. Agents Chemother. 2010, 54, 1343–1346. [Google Scholar] [CrossRef]
- Holthausen, D.J.; Lee, S.H.; Kumar, V.T.; Bouvier, N.M.; Krammer, F.; Ellebedy, A.H.; Wrammert, J.; Lowen, A.C.; George, S.; Pillai, M.R.; et al. An Amphibian Host Defense Peptide Is Virucidal for Human H1 Hemagglutinin-Bearing Influenza Viruses. Immunity 2017, 46, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Marcocci, M.E.; Amatore, D.; Villa, S.; Casciaro, B.; Aimola, P.; Franci, G.; Grieco, P.; Galdiero, M.; Palamara, A.T.; Mangoni, M.L.; et al. The Amphibian Antimicrobial Peptide Temporin B Inhibits In Vitro Herpes Simplex Virus 1 Infection. Antimicrob. Agents Chemother. 2018, 62, e02367-17. [Google Scholar] [CrossRef] [PubMed]
- Rivas, L.; Luque-Ortega, J.R.; Andreu, D. Amphibian antimicrobial peptides and Protozoa: Lessons from parasites. Biochim. Biophys. Acta 2009, 1788, 1570–1581. [Google Scholar] [CrossRef]
- Brand, G.D.; Leite, J.R.; Silva, L.P.; Albuquerque, S.; Prates, M.V.; Azevedo, R.B.; Carregaro, V.; Silva, J.S.; Sá, V.C.; Brandão, R.A.; et al. Dermaseptins from Phyllomedusa oreades and Phyllomedusa distincta. Anti-Trypanosoma cruzi activity without cytotoxicity to mammalian cells. J. Biol. Chem. 2002, 277, 49332–49340. [Google Scholar] [CrossRef]
- Krugliak, M.; Feder, R.; Zolotarev, V.Y.; Gaidukov, L.; Dagan, A.; Ginsburg, H.; Mor, A. Antimalarial activities of dermaseptin S4 derivatives. Antimicrob. Agents Chemother. 2000, 44, 2442–2451. [Google Scholar] [CrossRef]
- Vale, N.; Aguiar, L.; Gomes, P. Antimicrobial peptides: A new class of antimalarial drugs? Front. Pharmacol. 2014, 5, 275. [Google Scholar] [CrossRef]
- Baker, M.A.; Maloy, W.L.; Zasloff, M.; Jacob, L.S. Anticancer efficacy of Magainin2 and analogue peptides. Cancer Res. 1993, 53, 3052–3057. [Google Scholar]
- Lu, C.X.; Nan, K.J.; Lei, Y. Agents from amphibians with anticancer properties. Anti-Cancer Drugs 2008, 19, 931–939. [Google Scholar] [CrossRef]
- Mishra, B.; Wang, X.; Lushnikova, T.; Golla, R.M.; Narayana, J.L.; Wang, C.; McGuire, T.R.; Wang, G. Antibacterial, antifungal, anticancer activities and structural bioinformatics analysis of six naturally occurring temporins. Peptides 2018, 106, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Li, L.; Wang, H.; Li, M.; Wang, L.; Zhou, M.; Su, Q.; Chen, T.; Wu, Y. Novel peptide dermaseptin-PS1 exhibits anticancer activity via induction of intrinsic apoptosis signalling. J. Cell Mol. Med. 2019, 23, 1300–1312. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wu, Y.; Wang, T.; Chen, X.; Zhou, M.; Ma, C.; Xi, X.; Zhang, Y.; Chen, T.; Shaw, C.; et al. Brevinin-1GHd: A novel Hylarana guentheri skin secretion-derived Brevinin-1 type peptide with antimicrobial and anticancer therapeutic potential. Biosci. Rep. 2020, 40, BSR20200019. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, C.; Hamadat, S.; Le Saux, K.; Newton, C.; Mazouni, M.; Zargarian, L.; Miro-Padovani, M.; Zadigue, P.; Delbé, J.; Hamma-Kourbali, Y.; et al. Studies of the antitumor mechanism of action of dermaseptin B2, a multifunctional cationic antimicrobial peptide, reveal a partial implication of cell surface glycosaminoglycans. PLoS ONE 2017, 12, e0182926. [Google Scholar] [CrossRef] [PubMed]
- Musale, V.; Abdel-Wahab, Y.H.A.; Flatt, P.R.; Conlon, J.M.; Mangoni, M.L. Insulinotropic, glucose-lowering, and beta-cell anti-apoptotic actions of peptides related to esculentin-1a(1–21).NH2. Amino Acids 2018, 50, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Moffett, R.C.; Leprince, J.; Flatt, P.R. Identification of components in frog skin secretions with therapeutic potential as antidiabetic agents. Methods Mol Biol. 2018, 1719, 319–333. [Google Scholar]
- Ponti, D.; Mangoni, M.L.; Mignogna, G.; Simmaco, M.; Barra, D. An amphibian antimicrobial peptide variant expressed in Nicotiana tabacum confers resistance to phytopathogens. Biochem. J. 2003, 370, 121–127. [Google Scholar] [CrossRef]
- Lai, R.; Zheng, Y.T.; Shen, J.H.; Liu, G.J.; Liu, H.; Lee, W.H.; Tang, S.Z.; Zhang, Y. Antimicrobial peptides from skin secretions of Chinese red belly toad Bombina maxima. Peptides 2002, 23, 427–435. [Google Scholar] [CrossRef]
- Zairi, A.; Belaïd, A.; Gahbiche, A.; Hani, K. Spermicidal activity of dermaseptins. Contraception 2005, 72, 447–453. [Google Scholar] [CrossRef]
- Tanphaichitr, N.; Srakaew, N.; Alonzi, R.; Kiattiburut, W.; Kongmanas, K.; Zhi, R.; Li, W.; Baker, M.; Wang, G.; Hickling, D. Potential Use of Antimicrobial Peptides as Vaginal Spermicides/Microbicides. Pharmaceuticals 2016, 9, 13. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Shai, Y. Temporins and their synergism against Gram-negative bacteria and in lipopolysaccharide detoxification. Biochim. Biophys. Acta 2009, 1788, 1610–1619. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Yang, J.; He, X.; Mo, G.; Hong, J.; Yan, X.; Lin, D.; Lai, R. Structure and function of a potent lipopolysaccharide-binding antimicrobial and anti-inflammatory peptide. J. Med. Chem. 2013, 56, 3546–3556. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, A.N.; Liu, Y.; Wang, T.; Musgrave, I.F.; Pukala, T.L.; Tabor, R.F.; Martin, L.L.; Carver, J.A.; Bowie, J.H. The Amyloid Fibril-Forming Properties of the Amphibian Antimicrobial Peptide Uperin 3.5. Chembiochem 2016, 17, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Aisenbrey, C.; Amaro, M.; Pospíšil, P.; Hof, M.; Bechinger, B. Highly synergistic antimicrobial activity of magainin 2 and PGLa peptides is rooted in the formation of supramolecular complexes with lipids. Sci. Rep. 2020, 10, 11652. [Google Scholar] [CrossRef]
- Mishra, B.; Lushnikova, T.; Golla, R.M.; Wang, X.; Wang, G. Design and surface immobilization of short anti-biofilm peptides. Acta Biomater. 2017, 49, 316–328. [Google Scholar] [CrossRef]
- Lakshmaiah Narayana, J.; Mishra, B.; Lushinikova, T.; Wu, Q.; Chhonker, Y.S.; Zhang, Y.; Zarena, D.; Salnikov, E.; Dang, X.; Wang, F.; et al. Two distinct amphipathic peptide antibiotics with systemic efficacy. Proc. Natl. Acad. Sci. USA 2020, 117. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Han, H.; Miller, D.W.; Wang, G. Solution structures of human LL-37 fragments and NMR-based identification of a minimal membrane-targeting antimicrobial and anticancer region. J. Am. Chem. Soc. 2006, 128, 5776–5785. [Google Scholar] [CrossRef]
- Mishra, B.; Wang, G. The importance of amino acid composition in natural antimicrobial peptides (AMPs): An evolutional, structural, and functional perspective. Front. Immunol. 2012, 3, 221. [Google Scholar] [CrossRef]
- Wang, G. The Antimicrobial Peptide Database provides a platform for decoding the design principles of naturally occurring antimicrobial peptides. Protein Sci. 2020, 29, 8–18. [Google Scholar] [CrossRef]
- Wabnitz, P.A.; Bowie, J.H.; Tyler, M.J.; Wallace, J.C.; Smith, B.P. Differences in the skin peptides of the male and female Australian tree frog Litoria splendida. The discovery of the aquatic male sex pheromone splendipherin, together with phe8 caerulein and a new antibiotic peptide caerin 1.10. Eur. J. Biochem. 2000, 267, 269–275. [Google Scholar] [CrossRef]
- Wabnitz, P.A.; Walters, H.; Tyler, M.J.; Wallace, J.C.; Bowie, J.H. First record of host defence peptides in tadpoles. The magnificent tree frog Litoria splendida. J. Pept. Res. 1998, 52, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Xu, W.F.; Wang, M.M.; Wang, X.J.; Qian, Y.C. Characteristics of cathelicidin-Bg, a novel gene expressed in the ear-side gland of Bufo gargarizans. Genet. Mol. Res. 2016, 15, gmr15038481. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Ji, S.; Liu, W.; Yu, X.; Meng, Q.; Lai, R. Different expression profiles of bioactive peptides in Pelophylax nigromaculatus from distinct regions. Biosci. Biotechnol. Biochem. 2013, 77, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Steinborner, S.T.; Waugh, R.J.; Bowie, J.H.; Wallace, J.C.; Tyler, M.J.; Ramsay, S.L. New caerin antibacterial peptides from the skin glands of the Australian tree frog Litoria xanthomera. J. Pept. Sci. 1997, 3, 181–185. [Google Scholar] [CrossRef]
- Zhou, X.; Shi, D.; Zhong, R.; Ye, Z.; Ma, C.; Zhou, M.; Xi, X.; Wang, L.; Chen, T.; Kwok, H.F. Bioevaluation of Ranatuerin-2Pb from the Frog Skin Secretion of Rana pipiens and its Truncated Analogues. Biomolecules 2019, 9, 249. [Google Scholar] [CrossRef]
- Wang, G. Structures of human host defense cathelicidin LL-37 and its smallest antimicrobial peptide KR-12 in lipid micelles. J. Biol. Chem. 2008, 283, 32637–32643. [Google Scholar] [CrossRef]
- Wang, G.; Peterkofsky, A.; Clore, G.M. A novel membrane anchor function for the N-terminal amphipathic sequence of the signal-transducing protein IIAGlucose of the Escherichia coli phosphotransferase system. J. Biol. Chem. 2000, 275, 39811–39814. [Google Scholar] [CrossRef]
- Toke, O.; Bánóczi, Z.; Király, P.; Heinzmann, R.; Bürck, J.; Ulrich, A.S.; Hudecz, F. A kinked antimicrobial peptide from Bombina maxima. I. Three-dimensional structure determined by NMR in membrane-mimicking environments. Eur. Biophys. J. 2011, 40, 447–462. [Google Scholar] [CrossRef]
- Turner, J.; Cho, Y.; Dinh, N.N.; Waring, A.J.; Lehrer, R.I. Activities of LL-37, a cathelin-associated antimicrobial peptide of human neutrophils. Antimicrob. Agents Chemother. 1998, 42, 2206–2214. [Google Scholar] [CrossRef]
- Raaymakers, C.; Verbrugghe, E.; Hernot, S.; Hellebuyck, T.; Betti, C.; Peleman, C.; Claeys, M.; Bert, W.; Caveliers, V.; Ballet, S.; et al. Antimicrobial peptides in frog poisons constitute a molecular toxin delivery system against predators. Nat. Commun. 2017, 8, 1495. [Google Scholar] [CrossRef]
- Wang, G.; Keifer, P.A.; Peterkofsky, A. Solution structure of the N-terminal amphitropic domain of Escherichia coli glucose-specific enzyme IIA in membrane-mimetic micelles. Protein Sci. 2003, 12, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Abbassi, F.; Lequin, O.; Piesse, C.; Goasdoué, N.; Foulon, T.; Nicolas, P.; Ladram, A. Temporin-SHf, a new type of phe-rich and hydrophobic ultrashort antimicrobial peptide. J. Biol. Chem. 2010, 285, 16880–16892. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Mishra, B.; Lushnikova, T.; Narayana, J.L.; Wang, G. Amino Acid Composition Determines Peptide Activity Spectrum and Hot-Spot-Based Design of Merecidin. Adv. Biosyst. 2018, 2, 1700259. [Google Scholar] [CrossRef] [PubMed]
- Park, C.B.; Yi, K.S.; Matsuzaki, K.; Kim, M.S.; Kim, S.C. Structure-activity analysis of buforin II, a histone H2A-derived antimicrobial peptide: The proline hinge is responsible for the cell-penetrating ability of buforin II. Proc. Natl. Acad. Sci. USA 2000, 97, 8245–8250. [Google Scholar] [CrossRef]
- Lee, D.G.; Hahm, K.S.; Shin, S.Y. Structure and fungicidal activity of a synthetic antimicrobial peptide, P18, and its truncated peptides. Biotechnol. Lett. 2004, 26, 337–341. [Google Scholar] [CrossRef]
- Craik, D.J.; Cemazar, M.; Wang, C.K.; Daly, N.L. The cyclotide family of circular miniproteins: Nature’s combinatorial peptide template. Biopolymers 2006, 84, 250–266. [Google Scholar] [CrossRef]
- Wang, G. Improved methods for classification, prediction, and design of antimicrobial peptides. Methods Mol. Biol. 2015, 1268, 43–66. [Google Scholar]
- Wang, G. Post-translational Modifications of Natural Antimicrobial Peptides and Strategies for Peptide Engineering. Curr. Biotechnol. 2012, 1, 72–79. [Google Scholar] [CrossRef]
- Zhang, F.; Guo, Z.L.; Chen, Y.; Li, L.; Yu, H.N.; Wang, Y.P. Effects of C-terminal amidation and heptapeptide ring on the biological activities and advanced structure of amurin-9KY, a novel antimicrobial peptide identified from the brown frog, Rana kunyuensis. Zool. Res. 2019, 40, 198–204. [Google Scholar] [CrossRef]
- Muñoz, V.; Blanco, F.J.; Serrano, L. The hydrophobic-staple motif and a role for loop-residues in alpha-helix stability and protein folding. Nat. Struct. Biol. 1995, 2, 380–385. [Google Scholar] [CrossRef]
- Wang, G. Structural Analysis of Amphibian, Insect and Plant Host Defense Peptides Inspires the Design of Novel Therapeutic Molecules. In Host Defense Peptides and Their Potential as Therapeutic Agents; Epand, R.M., Ed.; Springer: Cham, Switzerland, 2016; pp. 229–252. [Google Scholar]
- Casciaro, B.; Cappiello, F.; Loffredo, M.R.; Ghirga, F.; Mangoni, M.L. The Potential of Frog Skin Peptides for Anti-Infective Therapies: The Case of Esculentin-1a(1–21)NH2. Curr. Med. Chem. 2020, 27, 1405–1419. [Google Scholar] [CrossRef] [PubMed]
- Jacob, L.; Zasloff, M. Potential therapeutic applications of magainins and other antimicrobial agents of animal origin. Ciba. Found. Symp. 1994, 186, 197–223. [Google Scholar] [PubMed]
- Uccelletti, D.; Zanni, E.; Marcellini, L.; Palleschi, C.; Barra, D.; Mangoni, M.L. Anti-Pseudomonas activity of frog skin antimicrobial peptides in a Caenorhabditis elegans infection model: A plausible mode of action in vitro and in vivo. Antimicrob. Agents Chemother. 2010, 54, 3853–3860. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L.; Luca, V.; McDermott, A.M. Fighting microbial infections: A lesson from amphibian skin-derived esculentin-1 peptides. Peptides 2015, 71, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Juretić, D.; Vukičević, D.; Petrov, D.; Novković, M.; Bojović, V.; Lučić, B.; Ilić, N.; Tossi, A. Knowledge-based computational methods for identifying or designing novel, non-homologous antimicrobial peptides. Eur. Biophys. J. 2011, 40, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Brito-Sánchez, Y.; Trimble, M.J.; Mansour, S.C.; Cherkasov, A.; Hancock, R.E.W. Computer-aided Discovery of Peptides that Specifically Attack Bacterial Biofilms. Sci. Rep. 2018, 8, 1871. [Google Scholar] [CrossRef]
- Cardoso, M.H.; Orozco, R.Q.; Rezende, S.B.; Rodrigues, G.; Oshiro, K.G.N.; Cândido, E.S.; Franco, O.L. Computer-Aided Design of Antimicrobial Peptides: Are We Generating Effective Drug Candidates? Front. Microbiol. 2020, 10, 3097. [Google Scholar] [CrossRef]
- Wang, G. Database-guided discovery of potent peptides to combat HIV-1 or Superbugs. Pharmaceuticals 2013, 6, 728–758. [Google Scholar] [CrossRef]
- Menousek, J.; Mishra, B.; Hanke, M.L.; Heim, C.E.; Kielian, T.; Wang, G. Database screening and in vivo efficacy of antimicrobial peptides against methicillin-resistant Staphylococcus aureus USA300. Int. J. Antimicrob. Agents 2012, 39, 402–406. [Google Scholar] [CrossRef]
- Mishra, B.; Wang, G. Ab initio design of potent anti-MRSA peptides based on database filtering technology. J. Am. Chem. Soc. 2012, 134, 12426–12429. [Google Scholar] [CrossRef]
- Mishra, B.; Narayana, J.L.; Lushinikova, T.; Wang, X.; Wang, G. Low cationicity is important for systemic in vivo efficacy of database-derived peptides against drug-resistant Gram-positive pathogens. Proc. Natl. Acad. Sci. USA 2019, 116, 13517–13522. [Google Scholar] [CrossRef] [PubMed]
- Rotem, S.; Mor, A. Antimicrobial peptide mimics for improved therapeutic properties. Biochim. Biophys. Acta 2009, 1788, 1582–1592. [Google Scholar] [CrossRef] [PubMed]
- Mensa, B.; Kim, Y.H.; Choi, S.; Scott, R.; Caputo, G.A.; DeGrado, W.F. Antibacterial mechanism of action of arylamide foldamers. Antimicrob. Agents Chemother. 2011, 55, 5043–5053. [Google Scholar] [CrossRef] [PubMed]
- Czyzewski, A.M.; Jenssen, H.; Fjell, C.D.; Waldbrook, M.; Chongsiriwatana, N.P.; Yuen, E.; Hancock, R.E.; Barron, A.E. In Vivo, In Vitro, and In Silico Characterization of Peptoids as Antimicrobial Agents. PLoS ONE 2016, 11, e0135961. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.A.; Weisblum, B.; Gellman, S.H. Interplay among folding, sequence, and lipophilicity in the antibacterial and hemolytic activities of alpha/beta-peptides. J. Am. Chem. Soc. 2007, 129, 417–428. [Google Scholar] [CrossRef]
- Chakraborty, S.; Liu, R.; Hayouka, Z.; Chen, X.; Ehrhardt, J.; Lu, Q.; Burke, E.; Yang, Y.; Weisblum, B.; Wong, G.C.; et al. Ternary nylon-3 copolymers as host-defense peptide mimics: Beyond hydrophobic and cationic subunits. J. Am. Chem. Soc. 2014, 136, 14530–14535. [Google Scholar] [CrossRef]
- Dong, Y.; Lushnikova, T.; Golla, R.M.; Wang, X.; Wang, G. Small molecule mimics of DFTamP1, a database designed anti-Staphylococcal peptide. Bioorg. Med. Chem. 2017, 25, 864–869. [Google Scholar] [CrossRef]
- Mishra, B.; Reiling, S.; Zarena, D.; Wang, G. Host defense antimicrobial peptides as antibiotics: Design and application strategies. Curr. Opin. Chem. Biol. 2017, 38, 87–96. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Koradi, R.; Billeter, M.; Wüthrich, K. MOLMOL: A program for display and analysis of macromolecular structures. J. Mol. Graph. 1996, 14, 32–51. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Peptide | Source Species | Continent | Count | Ref 2 |
|---|---|---|---|---|---|
| 1987 | Magainin | Xenopus laevis | Africa | 8 | [15] |
| 1988 | XPF | Xenopus laevis | Africa | 13 | [14] |
| 1991 | Dermaseptin | Phyllomedusa bicolor | S. America | 62 | [16] |
| 1992 | Brevinin | Rana brevipoda porsa | Asia | 221 | [17] |
| Caerin | Litoria splendida, | Australia | 29 | [18] | |
| 1993 | Esculentin | Rana esculenta | Europe | 58 | [19] |
| 1994 | Ranalexin | Rana catesbeiana | N. America | 4 | [20] |
| 1995 | Rugosin | Rana rugosa | Asia | 8 | [21] |
| 1996 | Temporin | Rana temporaria | Europe | 119 | [22] |
| Uperin | Litoria ewingi | Australia | 12 | [23] | |
| 1998 | Ranatuerin | Rana catesbeiana | N. America | 51 | [24] |
| Maculatin | Litoria eucnemis | Australia | 7 | [25] | |
| Dermatoxin | Agalychnis annae | S. America | 4 | [26] | |
| 1999 | Citropin | Litoria citropa | Australia | 6 | [27] |
| 2000 | Aurein | Litoria aurea | Australia | 12 | [28] |
| Phylloxin | Phyllomedusa bicolor | S. America | 2 | [29] | |
| Palustrin | Rana palustris | N. America | 27 | [30] | |
| Kassinatuerin | Kassina senegalensis | Africa | 5 | [31] | |
| 2001 | Tigerinin | Rana tigerina | Asia | 4 | [32] |
| Pseudin | Pseudis paradoxa | S. America | 4 | [33] | |
| Distinctin | Phyllomedusa distincta | S. America | 2 | [34] | |
| Nigrocin | Pelophylax nigromaculatus | Asia | 35 | [35] | |
| Dahlein | Litoria dahlii | Australia | 11 | [36] | |
| 2002 | Japonicin | Rana japonica | Asia | 7 | [37] |
| 2003 | Ranacyclin | Rana esculenta | Europe | 13 | [38] |
| 2004 | Ocellatin | Leptodactylus ocellatus | S. America | 18 | [39] |
| Ascaphin | Ascaphus truei | N. America | 13 | [40] | |
| 2005 | Phylloseptin | Phylllomedusa hypochondrialis | S. America | 30 | [41] |
| Hylin | Hyla biobeba | S. America | 3 | [42] | |
| 2006 | Lividin | Odorrana livida | Asia | 5 | [43] |
| Pelophylaxin | Pelophylax plancyi fukienensis | Asia | 6 | [44] | |
| 2007 | Dybowskin | Rana dybowskii | Asia | 8 | [45] |
| Pleurain | Rana pleuraden | Asia | 13 | [46] | |
| Odorranain | Odorrana graham | Asia | 31 | [47] | |
| Hyposin | Phyllomedusa hypochondrialis | S. America | 5 | [48] | |
| 2008 | Fallaxidin | Litoria fallax | Australia | 3 | [49] |
| 2010 | Nigroain | Rana nigrovittata | Asia | 8 | [50] |
| 2011 | Andersonin | Odorrana andersonii | Asia | 6 | [51] |
| Cathelicidin | Amolops loloensis | Asia | 9 | [52] | |
| 2012 | Hymenochirin | Hymenochirus boettgeri | Africa | 16 | [53] |
| 2014 | Frenatin | Sphaenorhynchus lacteus | S. America | 6 | [54] |
| 2015 | Defensin | Theloderma kwangsiensis | Asia | 2 | [55] |
| Activity | APD Count | Examples |
|---|---|---|
| Antibacterial | 982 | Magainin 2, Brevinin-1, Temporin-SHf |
| AntiGram+ | 169 | Temporin-1Ola, Aurein 5.2, Phylloseptin-S3 |
| AntiGram− | 84 | XT-2, Dermaseptin DA4, Leptoglycin |
| Anti-MRSA | 54 | Ranalexin, Temporin-1GHd, Temporin-1OLa |
| Antifungal | 490 | Dermaseptin-S1, Temporin-1GHd, PGLa |
| Candidacidal | 373 | Maximin 1, Esculentin-1, Temporin A |
| Antivirus | 45 | Brevinin-1, Urumin, Temporin B, |
| Anti-HIV | 36 | Caerin 1.1., Maculentin 1.1, Dermaseptin-S1 |
| Anti-parasitic | 53 | Dermaseptin-S1, Temporin L, Phylloseptin-S4 |
| Anticancer | 93 | Magainin 2, Aurein 1.2, Dermaseptin |
| Anti-diabetes | 15 | Esculentin-1, Amolopin, Dermaseptin-B4 |
| Anti-endotoxin | 13 | Temporin L, Cathelicidin-PP, Pseudin-2 |
| Insecticidal | 2 | Escculentin-1, Magainin 2 |
| Spermicidal | 5 | Magainin 2, Dermaseptin-S1, Dermaseptin-S4 |
| Chemotactic | 4 | Temporin-A, Temproin B, Dermaseptin-S9 |
| Wound healing | 4 | Magainin 2, Temporin A, Brevinin-2Ta |
| Antioxidant | 19 | Temproin-TP1, Pleurain-A1, Brevinin-1TP3 |
| Protease inhibitor | 4 | Odorranain-B1, P2-Hp-1935 |
| Anti-inflammatory | 3 | Cathelicidin-PY, Cathelicidin-PP, Esculentin-1GN |
| Activity Spectrum 1 | Count | Hydrophobic Content (Pho) 2 | Net Charge | Lysine% |
|---|---|---|---|---|
| Hemolytic | 163 | 55.6% | +2.67 | 12.6% |
| G+/G− | 982 | 51.1% | +2.55 | 12.9% |
| G− | 84 | 47.8% | +2.74 | 14.5% |
| G+ | 169 | 55.1% | +1.62 | 9.5% |
| Length Range | Peptide Count | Average Net Charge | L% | A% | G% | K% |
|---|---|---|---|---|---|---|
| 21–25 | 17 | +1.8 | 13.3 | 19.4 | 13.3 | 12.6 |
| 26–30 | 25 | +2.6 | 12.8 | 21.0 | 11.5 | 13.4 |
| 31–35 | 20 | +3.1 | 9.9 | 28.5 | 10.5 | 14.2 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G. Bioinformatic Analysis of 1000 Amphibian Antimicrobial Peptides Uncovers Multiple Length-Dependent Correlations for Peptide Design and Prediction. Antibiotics 2020, 9, 491. https://doi.org/10.3390/antibiotics9080491
Wang G. Bioinformatic Analysis of 1000 Amphibian Antimicrobial Peptides Uncovers Multiple Length-Dependent Correlations for Peptide Design and Prediction. Antibiotics. 2020; 9(8):491. https://doi.org/10.3390/antibiotics9080491
Chicago/Turabian StyleWang, Guangshun. 2020. "Bioinformatic Analysis of 1000 Amphibian Antimicrobial Peptides Uncovers Multiple Length-Dependent Correlations for Peptide Design and Prediction" Antibiotics 9, no. 8: 491. https://doi.org/10.3390/antibiotics9080491
APA StyleWang, G. (2020). Bioinformatic Analysis of 1000 Amphibian Antimicrobial Peptides Uncovers Multiple Length-Dependent Correlations for Peptide Design and Prediction. Antibiotics, 9(8), 491. https://doi.org/10.3390/antibiotics9080491