Antimicrobial Drug-Resistant Gram-Negative Saprophytic Bacteria Isolated from Ambient, Near-Shore Sediments of an Urbanized Estuary: Absence of β-Lactamase Drug-Resistance Genes

 ,
, 
Abstract
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
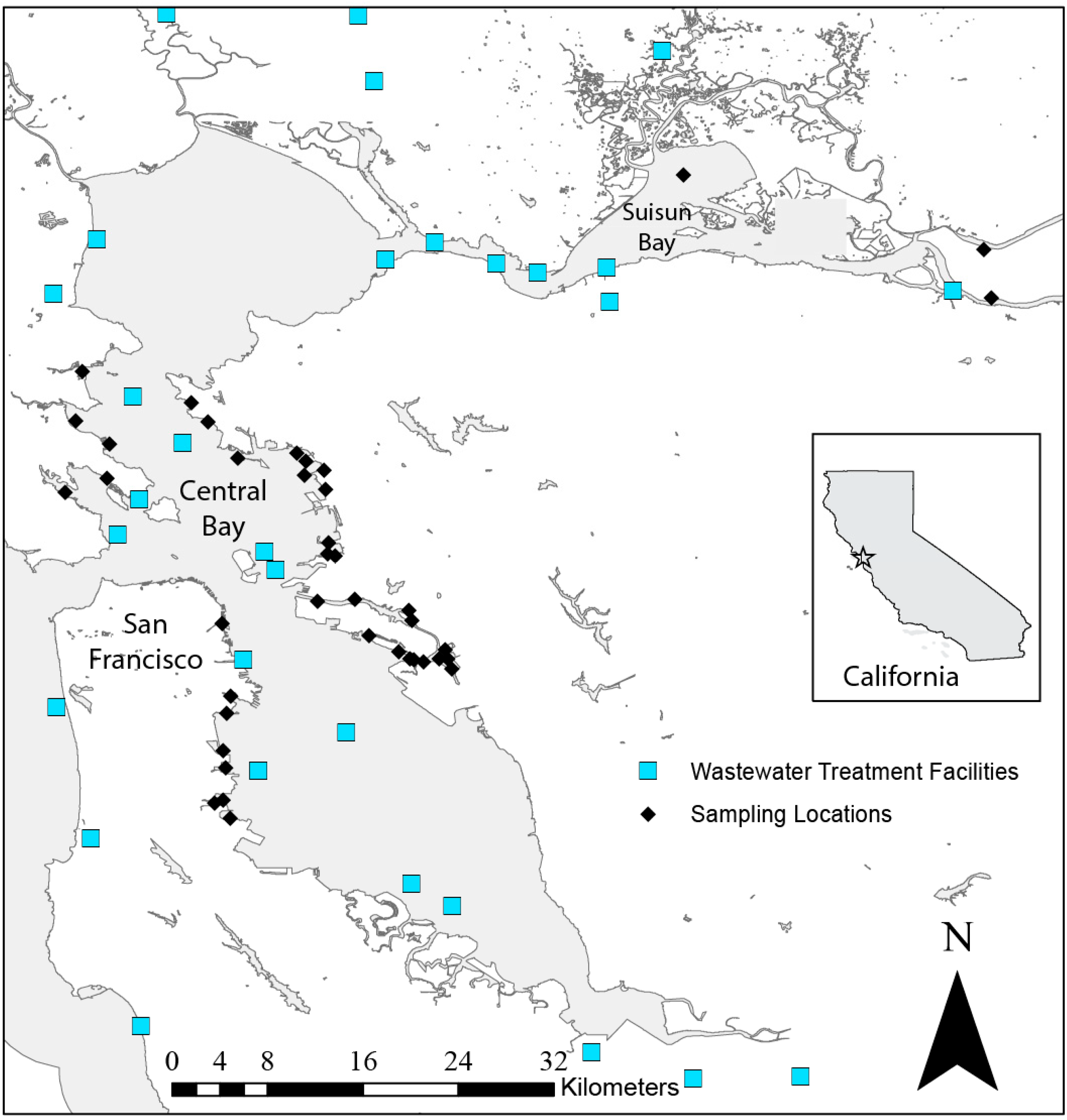
4.1. Sample Collection and Processing
4.2. Gram-Negative Bacteria Isolation
4.3. Bacterial Species Identification and Drug-Susceptibility Tests
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Korzeniewska, E.; Harnisz, M. Extended-Spectrum Beta-Lactamase (ESBL)-Positive Enterobacteriaceae in Municipal Sewage and Their Emission to the Environment. J. Environ. Manage. 2013, 128, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Goñi-Urriza, M.; Capdepuy, M.; Arpin, C.; Raymond, N.; Pierre Caumette, C.Q. Impact of an Urban Effluent on Antibiotic Resistance of Riverine Enterobacteriaceae and Aeromonas Spp. Appl. Environ. Microbiol. 2000, 66, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Tello, A.; Austin, B.; Telfer, T.C. Selective Pressure of Antibiotic Pollution on Bacteria of Importance to Public Health. Environ. Health Perspect. 2012, 120, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Amos, G.C.A.; Hawkey, P.M.; Gaze, W.H.; Wellington, E.M. Waste Water Effluent Contributes to the Dissemination of CTX-M-15 in the Natural Environment. J. Antimicrob. Chemother. 2014, 69, 1785–1791. [Google Scholar] [CrossRef]
- Fleming-Dutra, K.E.; Hersh, A.L.; Shapiro, D.J.; Bartoces, M.; Enns, E.A.; File, T.M.; Finkelstein, J.A.; Gerber, J.S.; Hyun, D.Y.; Linder, J.A.; et al. Prevalence of Inappropriate Antibiotic Prescriptions among Us Ambulatory Care Visits, 2010–2011. JAMA-J. Am. Med. Assoc. 2016, 315, 1864–1873. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Zhao, Y.; Li, B.; Huang, C.L.; Zhang, S.Y.; Yu, S.; Chen, Y.S.; Zhang, T.; Gillings, M.R.; Su, J.Q. Continental-Scale Pollution of Estuaries with Antibiotic Resistance Genes. Nat. Microbiol. 2017, 2, 16270. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.M.; Wertheim, H.F.L.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic Resistance-the Need for Global Solutions. Lancet Infect. Dis. 2013, 1057–1098. [Google Scholar] [CrossRef]
- Nagvekar, V.; Sawant, S.; Amey, S. Prevalence of Multi Drug Resistant Gram-Negative Bacteria Cases at Admission in Multispecialty Hospital. J. Glob. Antimicrob. Resist. 2020. [Google Scholar] [CrossRef]
- Fasciana, T.; Gentile, B.; Aquilina, M.; Ciammaruconi, A.; Mascarella, C.; Anselmo, A.; Fortunato, A.; Fillo, S.; Petralito, G.; Lista, F.; et al. Co-Existence of Virulence Factors and Antibiotic Resistance in New Klebsiella Pneumoniae Clones Emerging in South of Italy. BMC Infect. Dis. 2019, 19, 928. [Google Scholar] [CrossRef]
- Adams-Sapper, S.; Sergeevna-Selezneva, J.; Tartof, S.; Raphael, E.; Diep, B.A.; Perdreau-Remington, F.; Riley, L.W. Globally Dispersed Mobile Drug-Resistance Genes in Gram-Negative Bacterial Isolates from Patients with Bloodstream Infections in a US Urban General Hospital. J. Med. Microbiol. 2012, 61, 968–974. [Google Scholar] [CrossRef][Green Version]
- Center for Disease Control and Prevention. Antibiotic Resistance Threats in the United States; Center for Disease Control and Prevention: Atlanta, GA, USA, 2013. [Google Scholar]
- Berglund, B. Environmental Dissemination of Antibiotic Resistance Genes and Correlation to Anthropogenic Contamination with Antibiotics. Infect. Ecol. Epidemiol. 2015, 5, 28564. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef]
- Woodford, N.; Wareham, D.W.; Guerra, B.; Teale, C. Carbapenemase-Producing Enterobacteriaceae and Non-Enterobacteriaceae from Animals and the Environment: An Emerging Public Health Risk of Our Own Making? J. Antimicrob. Chemother. 2014, 69, 287–291. [Google Scholar] [CrossRef]
- Czekalski, N.; Sigdel, R.; Birtel, J.; Matthews, B.; Bürgmann, H. Does Human Activity Impact the Natural Antibiotic Resistance Background? Abundance of Antibiotic Resistance Genes in 21 Swiss Lakes. Environ. Int. 2015, 81, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Raphael, E.; Wong, L.K.; Riley, L.W. Extended-Spectrum Beta-Lactamase Gene Sequences in Gram-Negative Saprophytes on Retail Organic and Nonorganic Spinach. Appl. Environ. Microbiol. 2011, 77, 1601–1607. [Google Scholar] [CrossRef]
- Matyar, F.; Kaya, A.; Dinçer, S. Antibacterial Agents and Heavy Metal Resistance in Gram-Negative Bacteria Isolated from Seawater, Shrimp and Sediment in Iskenderun Bay, Turkey. Sci. Total Environ. 2008, 407, 279–285. [Google Scholar] [CrossRef]
- Griffin, D.W.; Banks, K.; Gregg, K.; Shedler, S.; Walker, B.K. Antibiotic Resistance in Marine Microbial Communities Proximal to a Florida Sewage Outfall System. Antibiotics 2020, 118. [Google Scholar] [CrossRef] [PubMed]
- Tacão, M.; Correia, A.; Henriques, I.S. Low Prevalence of Carbapenem-Resistant Bacteria in River Water: Resistance Is Mostly Related to Intrinsic Mechanisms. Microb. Drug Resist. 2015, 21, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Bradford, P.A. β-Lactams and β-Lactamase Inhibitors: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- Bush, K. Past and Present Perspectives on β-Lactamases. Antimicrob. Agents Chemother. 2018, 62, e01076-18. [Google Scholar] [CrossRef]
- Adams-Sapper, S.; Diep, B.A.; Perdreau-Remington, F.; Riley, L.W. Clonal Composition and Community Clustering of Drug-Susceptible and -Resistant Escherichia Coli Isolates from Bloodstream Infections. Antimicrob. Agents Chemother. 2013, 57, 490–497. [Google Scholar] [CrossRef]
- Papp-Wallace, K.M.; Endimiani, A.; Taracila, M.A.; Bonomo, R.A. Carbapenems: Past, Present, and Future. Antimicrob. Agents Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Dortet, L.; Poirel, L. Carbapenem Resistance in Enterobacteriaceae: Here Is the Storm! Trends Mol. Med. 2012, 18, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Scotta, C.; Juan, C.; Cabot, G.; Oliver, A.; Lalucat, J.; Bennasar, A.; Albertí, S. Environmental Microbiota Represents a Natural Reservoir for Dissemination of Clinically Relevant Metallo-β-Lactamases. Antimicrob. Agents Chemother. 2011, 55, 5376–5379. [Google Scholar] [CrossRef] [PubMed]
- Montezzi, L.F.; Campana, E.H.; Corrêa, L.L.; Justo, L.H.; Paschoal, R.P.; Da Silva, I.L.V.D.; Souza, M.D.C.M.; Drolshagen, M.; Picão, R.C. Occurrence of Carbapenemase-Producing Bacteria in Coastal Recreational Waters. Int. J. Antimicrob. Agents 2015, 45, 174–177. [Google Scholar] [CrossRef]
- Lu, S.Y.; Zhang, Y.L.; Geng, S.N.; Li, T.Y.; Ye, Z.M.; Zhang, D.S.; Zou, F.; Zhou, H.W. High Diversity of Extended-Spectrum Beta-Lactamase-Producing Bacteria in an Urban River Sediment Habitat. Appl. Environ. Microbiol. 2010, 76, 5972–5976. [Google Scholar] [CrossRef] [PubMed]
- Novick, E.; Senn, D.B. External Nutrient Loads to San Francisco Bay; San Francisco Estuary Institute: Richmond, CA, USA, 2014. [Google Scholar]
- Seto, E.Y.; Konnan, J.; Olivieri, A.W.; Danielson, R.E.; Gray, D.M.D. A Quantitative Microbial Risk Assessment of Wastewater Treatment Plant Blending: Case Study in San Francisco Bay. Environ. Sci. Water Res. Technol. 2015, 2, 134–145. [Google Scholar] [CrossRef]
- Trowbridge, P.R.; Davis, J.A.; Mumley, T.; Taberski, K.; Feger, N.; Valiela, L.; Ervin, J.; Arsem, N.; Olivieri, A.; Carroll, P.; et al. The Regional Monitoring Program for Water Quality in San Francisco Bay, California, USA: Science in Support of Managing Water Quality. Reg. Stud. Mar. Sci. 2016, 4, 21–33. [Google Scholar] [CrossRef]
- Shellenbarger, G.G.; Athearn, N.D.; Takekawa, J.Y.; Boehm, A.B. Fecal Indicator Bacteria and Salmonella in Ponds Managed as Bird Habitat, San Francisco Bay, California, USA. Water Res. 2008, 42, 2921–2930. [Google Scholar] [CrossRef]
- Heal the Bay. 2018–2019 Beach Report Card; Heal the Bay: Santa Monica, CA, USA, 2019. [Google Scholar]
- San Francisco Estuary Partnership. The State of the Estuary 2015; San Francisco Estuary Partnership: Oakland, CA, USA, 2015. [Google Scholar]
- Picão, R.C.; Cardoso, J.P.; Campana, E.H.; Nicoletti, A.G.; Petrolini, F.V.B.; Assis, D.M.; Juliano, L.; Gales, A.C. The Route of Antimicrobial Resistance from the Hospital Effluent to the Environment: Focus on the Occurrence of KPC-Producing Aeromonas Spp. and Enterobacteriaceae in Sewage. Diagn. Microbiol. Infect. Dis. 2013, 76, 80–85. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, X.-X.; Huang, K.; Miao, Y.; Shi, P.; Liu, B.; Long, C.; Li, A. Metagenomic Profiling of Antibiotic Resistance Genes and Mobile Genetic Elements in a Tannery Wastewater Treatment Plant. PLoS ONE 2013, 8, e76079. [Google Scholar] [CrossRef] [PubMed]
- Yee, D.; Wong, A.; Shimabuku, I.; Trowbridge, P. Characterization of Sediment Contamination in Central Bay Margin Areas; San Francisco Estuary Institute: Richmond, CA, USA, 2017. [Google Scholar]
- Baker-Austin, C.; Stockley, L.; Rangdale, R.; Martinez-Urtaza, J. Environmental Occurrence and Clinical Impact of Vibrio Vulnificus and Vibrio Parahaemolyticus: A European Perspective. Environ. Microbiol. Rep. 2010, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. The Genus Shewanella: From the Briny Depths below to Human Pathogen. Crit. Rev. Microbiol. 2014, 40, 293–312. [Google Scholar] [CrossRef] [PubMed]
- Holt, H.M.; Gahrn-Hansen, B.; Bruun, B. Shewanella Algae and Shewanella Putrefaciens: Clinical and Microbiological Characteristics. Clin. Microbiol. Infect. 2005, 11, 347–352. [Google Scholar] [CrossRef]
- Maravić, A.; Skočibušić, M.; Šamanić, I.; Fredotović, Ž.; Cvjetan, S.; Jutronić, M.; Puizina, J. Aeromonas Spp. Simultaneously Harbouring BlaCTX-M-15, BlaSHV-12, BlaPER-1 and BlaFOX-2, in Wild-Growing Mediterranean Mussel (Mytilus Galloprovincialis) from Adriatic Sea, Croatia. Int. J. Food Microbiol. 2013, 166, 301–308. [Google Scholar] [CrossRef]
- Parkins, M.D.; Gregson, D.B.; Pitout, J.D.D.; Ross, T.; Laupland, K.B. Population-Based Study of the Epidemiology and the Risk Factors for Pseudomonas Aeruginosa Bloodstream Infection. Infection 2010, 38, 25–32. [Google Scholar] [CrossRef]
- Poirel, L.; Héritier, C.; Nordmann, P. Chromosome-Encoded Ambler Class D β-Lactamase of Shewanella Oneidensis as a Progenitor of Carbapenem-Hydrolyzing Oxacillinase. Antimicrob. Agents Chemother. 2004, 48, 348–351. [Google Scholar] [CrossRef]
- Pellegrini, C.; Mercuri, P.S.; Celenza, G.; Galleni, M.; Segatore, B.; Sacchetti, E.; Volpe, R.; Amicosante, G.; Perilli, M. Identification of BlaIMP-22 in Pseudomonas Spp. in Urban Wastewater and Nosocomial Environments: Biochemical Characterization of a New IMP Metallo-Enzyme Variant and Its Genetic Location. J. Antimicrob. Chemother. 2009, 63, 901–908. [Google Scholar] [CrossRef][Green Version]
- Rossolini, G.M.; Condemi, M.A.; Pantanella, F.; Docquier, J.D.; Amicosante, G.; Thaller, M.C. Metallo-β-Lactamase Producers in Environmental Microbiota: New Molecular Class B Enzyme in Janthinobacterium Lividum. Antimicrob. Agents Chemother. 2001, 45, 837–844. [Google Scholar] [CrossRef]
- Shaw, K.S.; Rosenberg Goldstein, R.E.; He, X.; Jacobs, J.M.; Crump, B.C.; Sapkota, A.R. Antimicrobial Susceptibility of Vibrio Vulnificus and Vibrio Parahaemolyticus Recovered from Recreational and Commercial Areas of Chesapeake Bay and Maryland Coastal Bays. PLoS ONE 2014, 9, e89616. [Google Scholar] [CrossRef] [PubMed]
- Héritier, C.; Poirel, L.; Nordmann, P. Genetic and Biochemical Characterization of a Chromosome-Encoded Carbapenem-Hydrolyzing Ambler Class D β-Lactamase from Shewanella Algae. Antimicrob. Agents Chemother. 2004, 48, 1670–1675. [Google Scholar] [CrossRef] [PubMed]
- Girlich, D.; Poirel, L.; Nordmann, P. Novel Ambler Class A Carbapenem-Hydrolyzing β-Lactamase from a Pseudomonas Fluorescens Isolate from the Seine River, Paris, France. Antimicrob. Agents Chemother. 2010, 54, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Quinteira, S.; Ferreira, H.; Peixe, L. First Isolation of BlaVIM-2 in an Environmental Isolate of Pseudomonas Pseudoalcaligenes. Antimicrob. Agents Chemother. 2005, 49, 2140–2141. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Bao, X.; Ji, L.; Chen, L.; Liu, J.; Miao, J.; Chen, D.; Bian, H.; Li, Y.; Yu, G. Resistance Integrons: Class 1, 2 and 3 Integrons. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 45. [Google Scholar] [CrossRef]
- Stevens, D.L., Jr.; Olsen, A.R. Spatially Balanced Sampling of Natural Resources. J. Am. Stat. Assoc. 2004, 99, 262–278. [Google Scholar] [CrossRef]
- SFEI. Regional Monitoring Program for Water Quality in San Francisco Bay 2015 Bay Margins Sediment Study Cruise Plan; San Francisco Estuary Institute: Richmond, CA, USA, 2015. [Google Scholar]
- Coastal Contamination & Research. Contaminant Concentrations in Central Bay Margins Sediment Cruise Report; Coastal Contamination & Research: Moss Landing, CA, USA, 2015. [Google Scholar]
- CLSI. M100-S25 Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Dallenne, C.; da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a Set of Multiplex PCR Assays for the Detection of Genes Encoding Important β-Lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef]


{kind=link}
| Antibiotic a | Sites with Growth (N, %) | CFU/g b | Sites with Isolates Obtained (N) | Morphologically Distinct Isolates Obtained (N) c |
|---|---|---|---|---|
| No antimicrobial agent | 40 (100%) | 3513 | ||
| Ampicillin | 34 (85%) | 1280 | 32 | 87 |
| Cefotaxime | 9 (22.5%) | 16 | 8 | 11 |
| Imipenem | 15 (37.5%) | 106 | 15 | 37 |
| Gentamicin | 27 (67.5%) | 196 | 13 | 39 |
| Antibiotic | Species | Isolates (N) | Antibiotic | Species | Isolates (N) |
|---|---|---|---|---|---|
| Ampicillin (16 µg mL−1) | Total | 21 | Imipenem (1 µg mL−1) | Total | 35 |
| Vibrio alginolyticus | 6 | Aeromonas australiensis | 1 | ||
| Vibrio parahaemolyticus | 6 | Aeromonas hydrophila | 1 | ||
| Vibrio alginolyticus/parahaemolyticusa | 7 | Aeromonas veronii | 1 | ||
| Vibrio alginolyticus/azureusa | 2 | Castellaniella defragrans | 1 | ||
| Cefotaxime (1 µg mL−1) | Total | 11 | Pseudomonas sp. b | 1 | |
| Acinetobacter venetianus | 1 | Shewanella algae | 7 | ||
| Gallaecimonas xiamenensis | 1 | Shewanella algae/haliotisa | 11 | ||
| Pseudomonas fluorescens | 2 | Shewanella loihica | 8 | ||
| Pseudomonas oleovorans | 1 | Stenotrophomonas maltophilia | 2 | ||
| Pseudomonas putida | 3 | Vibrio diazotrophicus | 1 | ||
| Pseudomonas stutzeri | 1 | Vibrio fluvialis | 1 | ||
| Rhizobium sp. b | 1 | Gentamicin (10 µg mL−1) | Vibrio parahaemolyticus | 5 | |
| Vibrio fluvialis | 1 |
| Species | Isolates (N) | Plate | Resistance (Disc Diffusion) a | Intermediate Resistance (Disc Diffusion) a |
|---|---|---|---|---|
| Acinetobacter venetianus | 1 | CTX | CTX | |
| Aeromonas australiensis | 1 | IPM | AMC | |
| Aeromonas hydrophila | 1 | IPM | FOX | AMC |
| Aeromonas veronii | 1 | IPM | None | |
| Shewanella algae | 4 | IPM | None | |
| Shewanella algae | 3 | IPM | IPM | |
| Shewanella algae/halitosisb | 11 | IPM | None | |
| Shewanella loihica | 8 | IPM | None | |
| Vibrio alginolyticus | 3 | AMP | AMP | |
| Vibrio alginolyticus | 1 | AMP | AMP, CTX, GEN, FOX | |
| Vibrio alginolyticus | 1 | AMP | AMP, GEN | |
| Vibrio alginolyticus/parahaemolyticusb | 4 | AMP | AMP | |
| Vibrio diazotrophicus | 1 | IPM | None | |
| Vibrio fluvialis | 1 | CTX | AMP, CTX, AMC | FOX |
| Vibrio fluvialis | 1 | IPM | AMP | AMC |
| Vibrio parahaemolyticus | 5 | AMP | AMP | |
| Vibrio parahaemolyticus | 1 | AMP | AMP | CIP |
| Vibrio parahaemolyticus | 5 | GEN | AMP |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moritz, C.F.; Snyder, R.E.; Riley, L.W.; Immke, D.W.; Greenfield, B.K. Antimicrobial Drug-Resistant Gram-Negative Saprophytic Bacteria Isolated from Ambient, Near-Shore Sediments of an Urbanized Estuary: Absence of β-Lactamase Drug-Resistance Genes. Antibiotics 2020, 9, 400. https://doi.org/10.3390/antibiotics9070400
Moritz CF, Snyder RE, Riley LW, Immke DW, Greenfield BK. Antimicrobial Drug-Resistant Gram-Negative Saprophytic Bacteria Isolated from Ambient, Near-Shore Sediments of an Urbanized Estuary: Absence of β-Lactamase Drug-Resistance Genes. Antibiotics. 2020; 9(7):400. https://doi.org/10.3390/antibiotics9070400
Chicago/Turabian StyleMoritz, Charles F., Robert E. Snyder, Lee W. Riley, Devin W. Immke, and Ben K. Greenfield. 2020. "Antimicrobial Drug-Resistant Gram-Negative Saprophytic Bacteria Isolated from Ambient, Near-Shore Sediments of an Urbanized Estuary: Absence of β-Lactamase Drug-Resistance Genes" Antibiotics 9, no. 7: 400. https://doi.org/10.3390/antibiotics9070400
APA StyleMoritz, C. F., Snyder, R. E., Riley, L. W., Immke, D. W., & Greenfield, B. K. (2020). Antimicrobial Drug-Resistant Gram-Negative Saprophytic Bacteria Isolated from Ambient, Near-Shore Sediments of an Urbanized Estuary: Absence of β-Lactamase Drug-Resistance Genes. Antibiotics, 9(7), 400. https://doi.org/10.3390/antibiotics9070400