
whISOBAXTM Inhibits Bacterial Pathogenesis and Enhances the Effect of Antibiotics

 , ,
, ,
Abstract
1. Introduction
2. Methods
2.1. Bacteria
2.2. Test Formulations
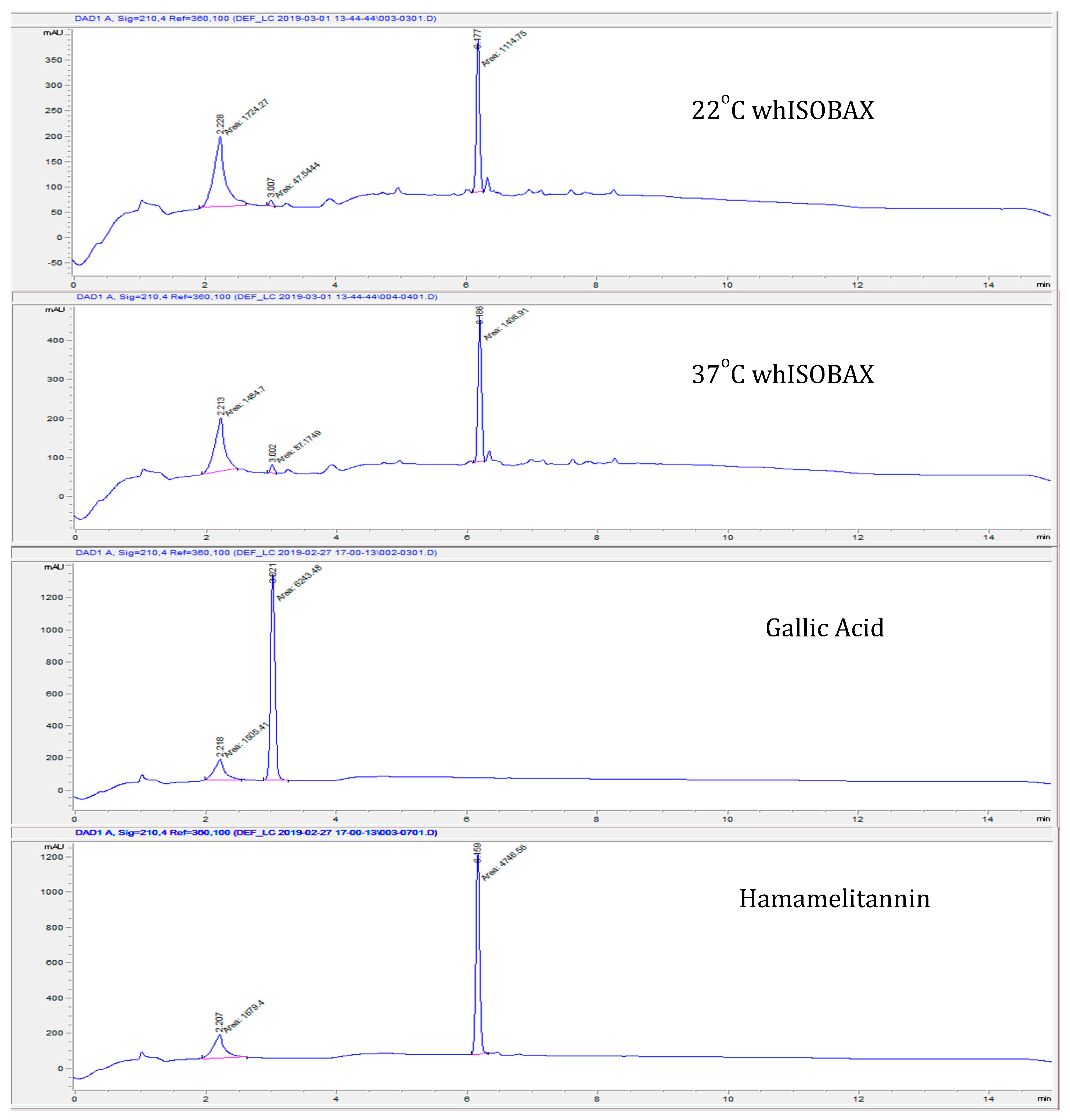
2.3. Stability of Hamamelitannin (HAMA) and Gallic Acid (GA) Content in whISOBAX by Column Chromatography
2.4. Minimal Inhibitory Concentrations (MICs)
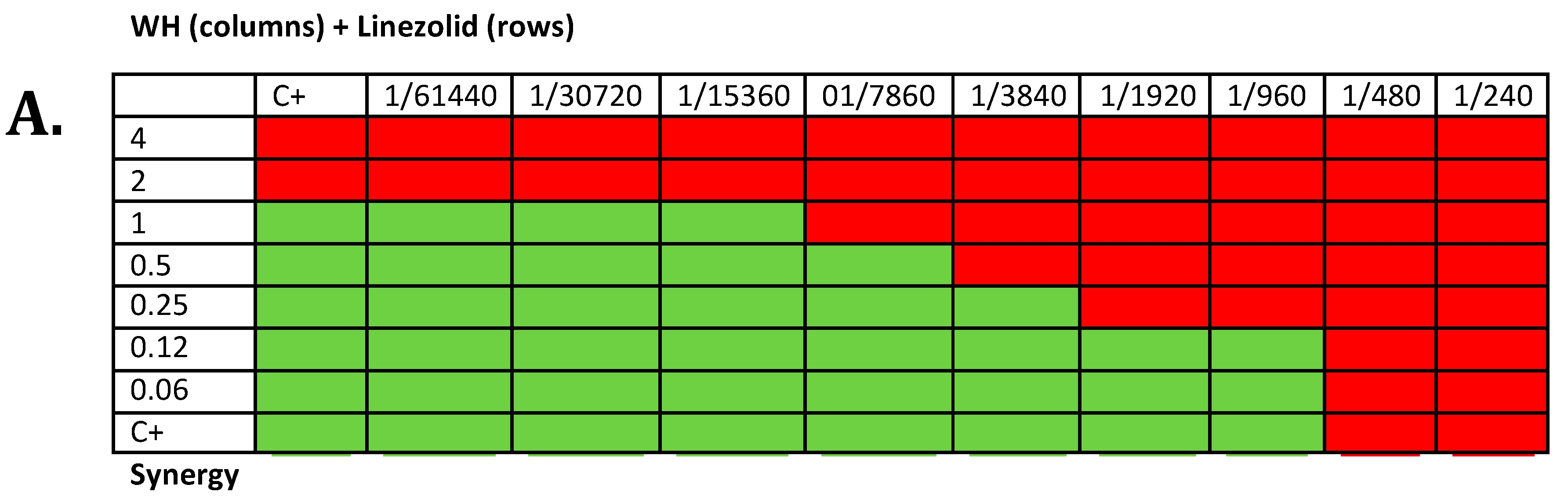
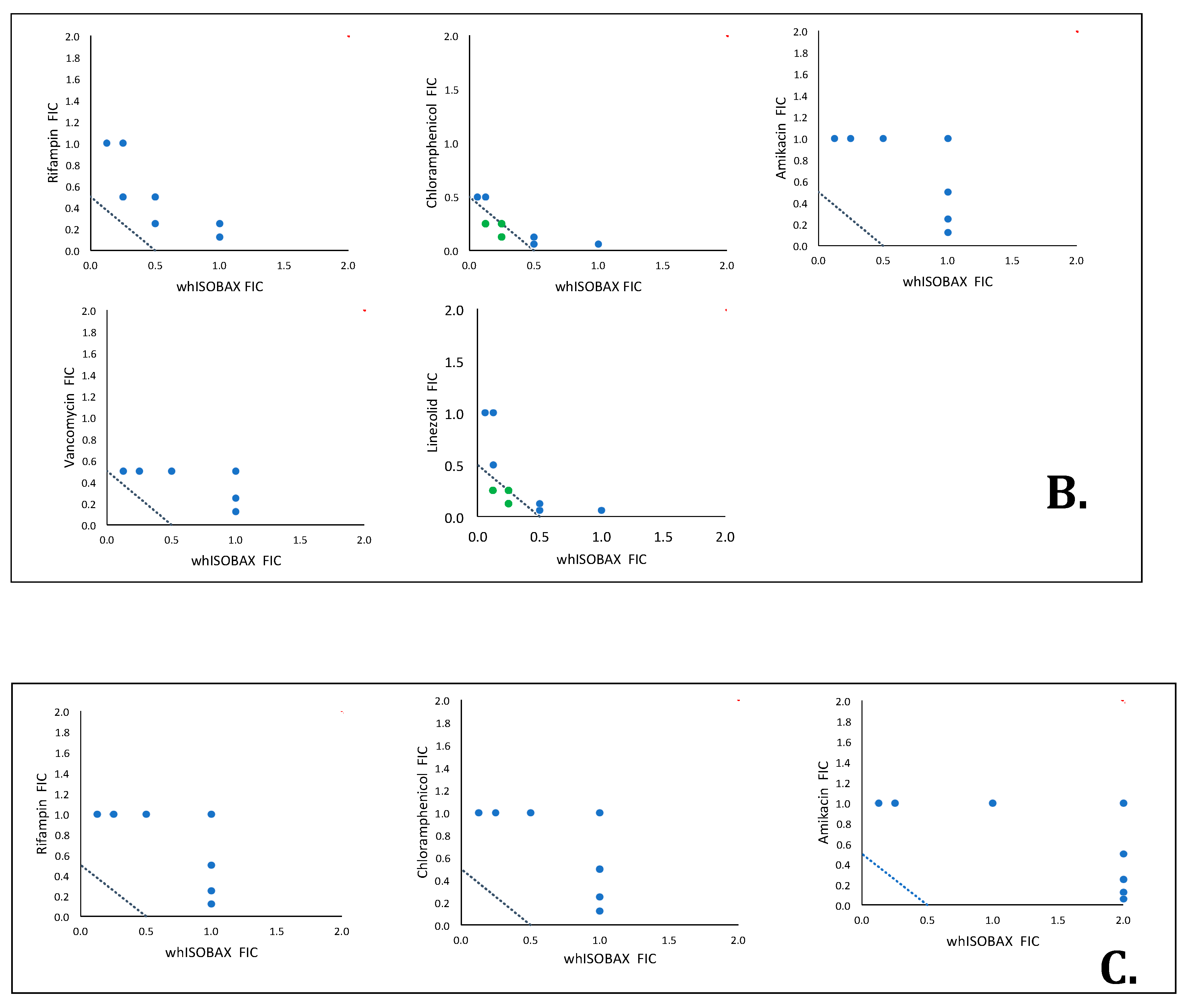
2.5. Synergy Testing
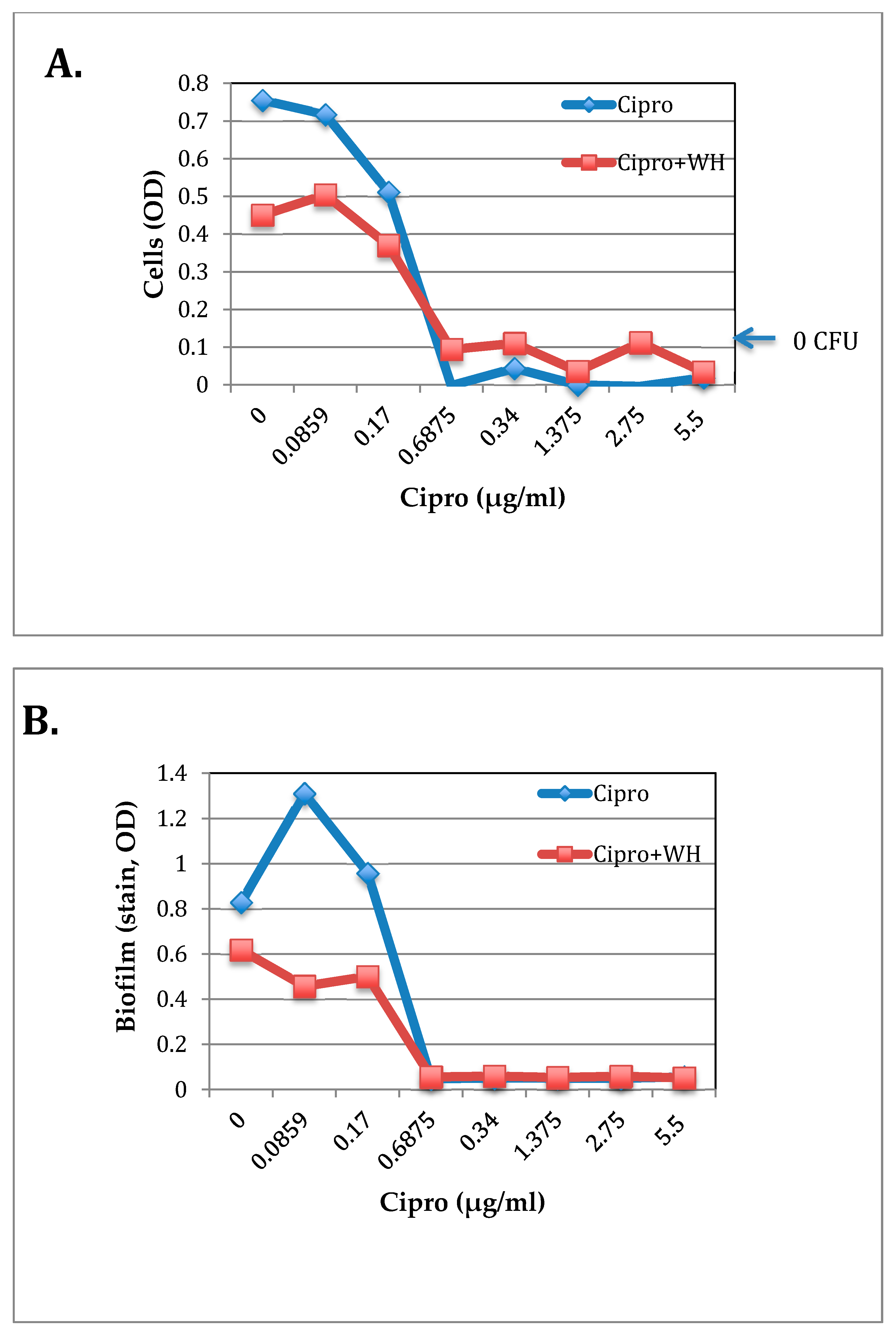
2.6. The Effect of WH and Ciprofloxacin on S. epidermidis Growth and Biofilm Formation
2.7. Staphylococcal Enterotoxin A (SEA) Production
2.8. Statistical Analysis
3. Results
3.1. Stability of Hamamelitannin (HAMA) and Gallic Acid in whISOBAX (WH)
3.2. The Effect of WH on the In Vitro Growth of Clinically Relevant Bacteria
3.3. whISOBAX Can Enhance the Antibacterial Effect of Antibiotics
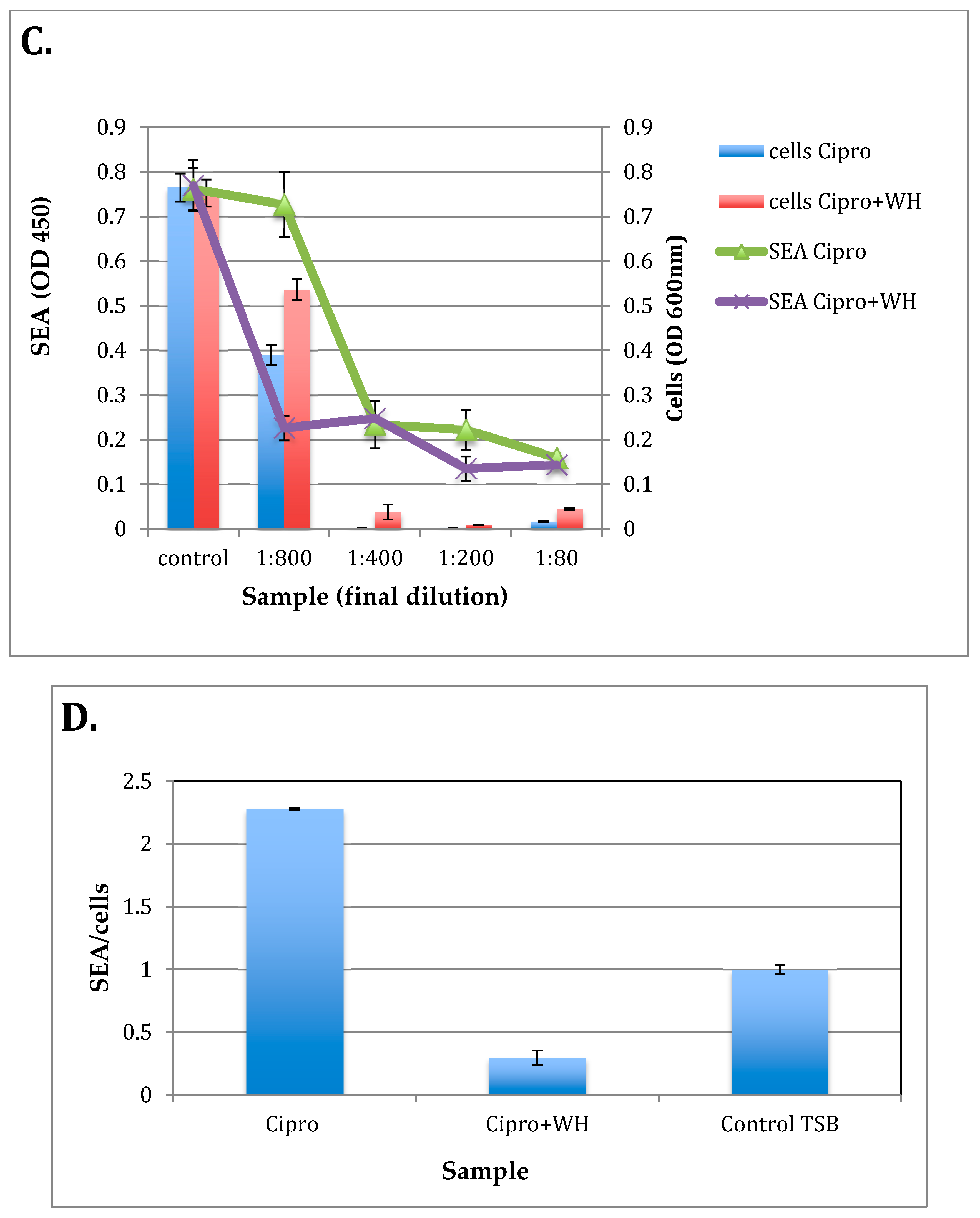
3.4. The Effect of whISOBAX and Ciprofloxacin on Staphylococcal Growth and Pathogenesis (Biofilm Formation and Toxin Production)
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vestergaard, M.; Frees, D.; Ingmer, H. Antibiotic Resistance and the MRSA Problem. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Danna, D.M. Hospital Costs Associated with Sepsis Compared with Other Medical Conditions. Crit. Care Nurs. Clin. North Am. 2018, 30, 389–398. [Google Scholar] [CrossRef] [PubMed]
- DeMellow, J.; Kim, T.Y. Technology-enabled performance monitoring in intensive care: An integrative literature review. Intensive Crit. Care Nurs. 2018, 48, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Kouatly, I.A.; Nassar, N.; Nizam, M.; Badr, L.K. Evidence on Nurse Staffing Ratios and Patient Outcomes in a Low-Income Country: Implications for Future Research and Practice. Worldviews Evid. Based Nurs. 2018, 15, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Monegro, A.F.; Regunath, H. Hospital Acquired Infections; StatPearls: Treasure Island, FL, USA, 2019. [Google Scholar]
- Centers for Disease Control and Prevention. Nearly Half a Million Americans Suffered from Clostridium Difficile Infections in a Single Year; CDC: Atlanta, GA, USA, 2017. [Google Scholar]
- Lindsay, D.; von Holy, A. Bacterial biofilms within the clinical setting: What healthcare professionals should know. J. Hosp. Infect. 2006, 64, 313–325. [Google Scholar] [CrossRef]
- Percival, S.L.; Suleman, L.; Vuotto, C.; Donelli, G. Healthcare-associated infections, medical devices and biofilms: Risk, tolerance and control. J. Med. Microbiol. 2015, 64, 323–334. [Google Scholar] [CrossRef]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- James, G.A.; Swogger, E.; Wolcott, R.; Pulcini, E.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in chronic wounds. Wound Repair Regen. 2008, 16, 37–44. [Google Scholar] [CrossRef]
- Anderson, G.G.; Moreau-Marquis, S.; Stanton, B.A.; O’Toole, G.A. In vitro analysis of tobramycin-treated Pseudomonas aeruginosa biofilms on cystic fibrosis-derived airway epithelial cells. Infect. Immun. 2008, 76, 1423–1433. [Google Scholar] [CrossRef]
- Hoiby, N.; Ciofu, O.; Bjarnsholt, T. Pseudomonas aeruginosa biofilms in cystic fibrosis. Future Microbiol. 2010, 5, 1663–1674. [Google Scholar] [CrossRef]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [PubMed]
- Huber, B.; Eberl, L.; Feucht, W.; Polster, J. Influence of polyphenols on bacterial biofilm formation and quorum-sensing. Z. Nat. C J. Biosci. 2003, 58, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed]
- Sirk, T.W.; Brown, E.F.; Friedman, M.; Sum, A.K. Molecular binding of catechins to biomembranes: Relationship to biological activity. J. Agric. Food Chem. 2009, 57, 6720–6728. [Google Scholar] [CrossRef]
- Sirk, T.W.; Brown, E.F.; Sum, A.K.; Friedman, M. Molecular dynamics study on the biophysical interactions of seven green tea catechins with lipid bilayers of cell membranes. J. Agric. Food Chem. 2008, 56, 7750–7758. [Google Scholar] [CrossRef]
- Brackman, G.; Breyne, K.; De Rycke, R.; Vermote, A.; Van Nieuwerburgh, F.; Meyer, E.; Van Calenbergh, S.; Coenye, T. The Quorum Sensing Inhibitor Hamamelitannin Increases Antibiotic Susceptibility of Staphylococcus aureus Biofilms by Affecting Peptidoglycan Biosynthesis and eDNA Release. Sci. Rep. 2016, 6, 20321. [Google Scholar] [CrossRef]
- Brackman, G.; Cos, P.; Maes, L.; Nelis, H.J.; Coenye, T. Quorum sensing inhibitors increase the susceptibility of bacterial biofilms to antibiotics in vitro and in vivo. Antimicrob. Agents Chemother. 2011, 55, 2655–2661. [Google Scholar] [CrossRef]
- Cobrado, L.; Azevedo, M.M.; Silva-Dias, A.; Ramos, J.P.; Pina-Vaz, C.; Rodrigues, A.G. Cerium, chitosan and hamamelitannin as novel biofilm inhibitors? J. Antimicrob. Chemother. 2012, 67, 1159–1162. [Google Scholar] [CrossRef]
- Cobrado, L.; Silva-Dias, A.; Azevedo, M.M.; Pina-Vaz, C.; Rodrigues, A.G. In vivo antibiofilm effect of cerium, chitosan and hamamelitannin against usual agents of catheter-related bloodstream infections. J. Antimicrob. Chemother. 2013, 68, 126–130. [Google Scholar] [CrossRef]
- Kiran, M.D.; Adikesavan, N.V.; Cirioni, O.; Giacometti, A.; Silvestri, C.; Scalise, G.; Ghiselli, R.; Saba, V.; Orlando, F.; Shoham, M.; et al. Discovery of a quorum-sensing inhibitor of drug-resistant staphylococcal infections by structure-based virtual screening. Mol. Pharmacol. 2008, 73, 1578–1586. [Google Scholar] [CrossRef]
- Vermote, A.; Brackman, G.; Risseeuw, M.D.P.; Coenye, T.; Van Calenbergh, S. Novel hamamelitannin analogues for the treatment of biofilm related MRSA infections-A scaffold hopping approach. Eur. J. Med. Chem. 2017, 127, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Provan, G.J.; Helliwell, K. Determination of hamamelitannin, catechins and gallic acid in witch hazel bark, twig and leaf by HPLC. J. Pharm. Biomed. Anal. 2003, 33, 539–544. [Google Scholar] [CrossRef]
- Pillai, S.K.; Moellering, R.C.; Eliopoulos, G.M. Antimicrobial Combinations. In Antibiotics in Laboratory Medicine; Lorian, V., Ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2005. [Google Scholar]
- Fratini, F.; Mancini, S.; Turchi, B.; Friscia, E.; Pistelli, L.; Giusti, G.; Cerri, D. A novel interpretation of the Fractional Inhibitory Concentration Index: The case Origanum vulgare L. and Leptospermum scoparium J. R. et G. Forst essential oils against Staphylococcus aureus strains. Microbiol. Res. 2017, 195, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.Y.; Pei, L.; Liu, Q.; Chen, S.; Dou, H.; Shu, G.; Yuan, Z.X.; Lin, J.; Peng, G.; Zhang, W.; et al. Isobologram Analysis: A Comprehensive Review of Methodology and Current Research. Front. Pharmacol. 2019, 10, 1222. [Google Scholar] [CrossRef]
- Kuang, H.; Wang, W.; Xu, L.; Ma, W.; Liu, L.; Wang, L.; Xu, C. Monoclonal antibody-based sandwich ELISA for the detection of staphylococcal enterotoxin A. Int. J. Environ. Res. Public Health 2013, 10, 1598–1608. [Google Scholar] [CrossRef]
- Rasooly, R.; Molnar, A.; Choi, H.Y.; Do, P.; Racicot, K.; Apostolidis, E. In-Vitro Inhibition of Staphylococcal Pathogenesis by Witch-Hazel and Green Tea Extracts. Antibiotics 2019, 8, 244. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simoes, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Bielaszewska, M.; Idelevich, E.A.; Zhang, W.; Bauwens, A.; Schaumburg, F.; Mellmann, A.; Peters, G.; Karch, H. Effects of antibiotics on Shiga toxin 2 production and bacteriophage induction by epidemic Escherichia coli O104:H4 strain. Antimicrob. Agents Chemother. 2012, 56, 3277–3282. [Google Scholar] [CrossRef]
- Kimmitt, P.T.; Harwood, C.R.; Barer, M.R. Toxin gene expression by shiga toxin-producing Escherichia coli: The role of antibiotics and the bacterial SOS response. Emerg. Infect. Dis. 2000, 6, 458–465. [Google Scholar] [CrossRef]
- Zhang, X.; McDaniel, A.D.; Wolf, L.E.; Keusch, G.T.; Waldor, M.K.; Acheson, D.W. Quinolone antibiotics induce Shiga toxin-encoding bacteriophages, toxin production, and death in mice. J. Infect. Dis. 2000, 181, 664–670. [Google Scholar] [CrossRef]
- Balaban, N.; Rasooly, A. Staphylococcal enterotoxins. Int. J. Food Microbiol. 2000, 61, 1–10. [Google Scholar] [CrossRef]
- Friedman, M.; Rasooly, R. Review of the inhibition of biological activities of food-related selected toxins by natural compounds. Toxins 2013, 5, 743–775. [Google Scholar] [CrossRef] [PubMed]
- Rasooly, R.; Molnar, A.; Do, P.; Morroni, G.; Brescini, L.; Cirioni, O.; Giacometti, A.; Apostolidis, E. Witch Hazel Significantly Improves the Efficacy of Commercially Available Teat Dips. Pathogens 2020, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.; Goldkorn, T.; Gov, Y.; Hirshberg, M.; Koyfman, N.; Matthews, H.R.; Nhan, R.T.; Singh, B.; Uziel, O. Regulation of Staphylococcus aureus pathogenesis via target of RNAIII-activating Protein (TRAP). J. Biol. Chem. 2001, 276, 2658–2667. [Google Scholar] [CrossRef]
- Balaban, N.; Goldkorn, T.; Nhan, R.T.; Dang, L.B.; Scott, S.; Ridgley, R.M.; Rasooly, A.; Wright, S.C.; Larrick, J.W.; Rasooly, R.; et al. Autoinducer of virulence as a target for vaccine and therapy against Staphylococcus aureus. Science 1998, 280, 438–440. [Google Scholar] [CrossRef]
- Kiran, M.D.; Bala, S.; Hirshberg, M.; Balaban, N. YhgC protects Bacillus anthracis from oxidative stress. Int. J. Artif. Organs 2010, 33, 590–607. [Google Scholar] [CrossRef]
- Kiran, M.D.; Balaban, N. TRAP plays a role in stress response in Staphylococcus aureus. Int. J. Artif. Organs 2009, 32, 592–599. [Google Scholar] [CrossRef]
- Kiran, M.D.; Giacometti, A.; Cirioni, O.; Balaban, N. Suppression of biofilm related, device-associated infections by staphylococcal quorum sensing inhibitors. Int. J. Artif. Organs 2008, 31, 761–770. [Google Scholar] [CrossRef]
- Lopez-Leban, F.; Kiran, M.D.; Wolcott, R.; Balaban, N. Molecular mechanisms of RIP, an effective inhibitor of chronic infections. Int. J. Artif. Organs 2010, 33, 582–589. [Google Scholar] [CrossRef]
- Wolcott, R.; Lopez-Leban, F.; Kiran, M.; Balaban, N. Wound Healing by an Anti-Staphylococcal Biofilm Approach. In Biofilm Highlights; Hans-Curt, F., Wingender, J., Szewzyk, U., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 141–161. [Google Scholar] [CrossRef]
- Cirioni, O.; Simonetti, O.; Morroni, G.; Brescini, L.; Kamysz, W.; Kamysz, E.; Orlando, F.; Pierpaoli, E.; Caffarini, M.; Orciani, M.; et al. Efficacy of Pexiganan Combination with Tigecycline in a Mouse Model of Pseudomonas aeruginosa Sepsis. Curr. Top. Med. Chem. 2018, 18, 2127–2132. [Google Scholar] [CrossRef]
- Gyawali, R.; Ibrahim, S.A. Natural products as antimicrobial agents. Food Control 2014, 46, 412–429. [Google Scholar] [CrossRef]
- Morroni, G.; Simonetti, O.; Brenciani, A.; Brescini, L.; Kamysz, W.; Kamysz, E.; Neubauer, D.; Caffarini, M.; Orciani, M.; Giovanetti, E.; et al. In vitro activity of Protegrin-1, alone and in combination with clinically useful antibiotics, against Acinetobacter baumannii strains isolated from surgical wounds. Med. Microbiol. Immunol. 2019, 208, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Henrick, K.; Hirshberg, M. Structure of the signal transduction protein TRAP (target of RNAIII-activating protein). Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2012, 68, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Jayaswal, R.K.; Wilkinson, B.J. Cell wall-active antibiotic induced proteins of Staphylococcus aureus identified using a proteomic approach. FEMS Microbiol. Lett. 2001, 199, 79–84. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | MIC Mean (µg/mL) | MIC Range (µg/mL) * | Mean Dilutions of WH (50 mg/mL) that Inhibit Growth |
|---|---|---|---|
| Gram Positives | |||
| S. epidermidis RP62A | 26 | 26 | 1/1920 |
| S. aureus ATCC 43300 | 45 | 26–78 | 1/1120 |
| S. aureus ATCC 29213 | 104 | 104 | 1/460 |
| E. faecalis ATCC 29212 | 45 | 39–52 | 1/1120 |
| E. faecium 64/3 | 34 | 19–52 | 1/1000 |
| S. agalactiae 1357 | 3958 | 1250–6667 | 1/23 |
| S. pneumoniae ATCC49619 | 4583 | 2500–6667 | 1/13 |
| Gram Negatives | |||
| A. baumannii ATCC 19606 | 182 | 156–208 | 1/280 |
| K. pneumoniae ATCC 700603 | 572 | 312–833 | 1/110 |
| P. aeruginosa ATCC 27853 | 2916 | 1667–5000 | 1/30 |
| E. coli ATCC25922 | 3657 | 1250–10,000 | 1/25 |
| Antibiotic | Minimum FICI | Maximum FICI | Interaction |
|---|---|---|---|
| S. aureus ATCC 29213 | |||
| Rifampin | 0.625 | 1.250 | Cumulative |
| Chloramphenicol | 0.375 | 1.125 | Synergistic |
| Amikacin | 1.125 | 2.000 | Indifferent |
| Vancomycin | 0.625 | 1.500 | Cumulative |
| Linezolid | 0.375 | 1.250 | Synergistic |
| E. coli ATCC 25922 | |||
| Rifampin | 1.125 | 2.000 | Indifferent |
| Chloramphenicol | 1.125 | 2.000 | Indifferent |
| Amikacin | 1.125 | 3.000 | Indifferent |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasooly, R.; Choi, H.-Y.; Do, P.; Morroni, G.; Brescini, L.; Cirioni, O.; Giacometti, A.; Apostolidis, E. whISOBAXTM Inhibits Bacterial Pathogenesis and Enhances the Effect of Antibiotics. Antibiotics 2020, 9, 264. https://doi.org/10.3390/antibiotics9050264
Rasooly R, Choi H-Y, Do P, Morroni G, Brescini L, Cirioni O, Giacometti A, Apostolidis E. whISOBAXTM Inhibits Bacterial Pathogenesis and Enhances the Effect of Antibiotics. Antibiotics. 2020; 9(5):264. https://doi.org/10.3390/antibiotics9050264
Chicago/Turabian StyleRasooly, Reuven, Hwang-Yong Choi, Paula Do, Gianluca Morroni, Lucia Brescini, Oscar Cirioni, Andrea Giacometti, and Emmanouil Apostolidis. 2020. "whISOBAXTM Inhibits Bacterial Pathogenesis and Enhances the Effect of Antibiotics" Antibiotics 9, no. 5: 264. https://doi.org/10.3390/antibiotics9050264
APA StyleRasooly, R., Choi, H.-Y., Do, P., Morroni, G., Brescini, L., Cirioni, O., Giacometti, A., & Apostolidis, E. (2020). whISOBAXTM Inhibits Bacterial Pathogenesis and Enhances the Effect of Antibiotics. Antibiotics, 9(5), 264. https://doi.org/10.3390/antibiotics9050264