Antimicrobial Resistance Profiles of Adherent Invasive Escherichia coli Show Increased Resistance to β-Lactams

 , , , , and
, , , , and
Abstract
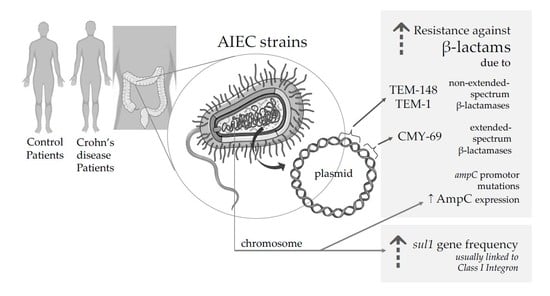
1. Introduction
2. Results
2.1. Susceptibility Profiles in AIEC and Non-AIEC Strains
2.2. Susceptibility Profiles in E. coli From Crohn’s Disease and Control Subjects
2.3. Mechanism of Resistance Associated with Strains Resistant to β-Lactams
2.4. Genomic Analysis of Antimicrobial Resistance Genes of Internal and External Strain Collections
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Antimicrobial Susceptibility Determination
4.3. Detection of AmpC β-Lactamases
4.4. Genome Sequencing of AIEC Strains
4.5. Genomic Analysis for the Detection of Antimicrobial Resistance Genes and ampC Variants
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Boudeau, J.; Glasser, A.L.; Masseret, E.; Joly, B.; Darfeuille-Michaud, A. Invasive Ability of an Escherichia coli Strain Isolated from the Ileal Mucosa of a Patient with Crohn’s Disease. Infect. Immun. 1999, 6, 4499–4509. [Google Scholar] [CrossRef]
- Baumgart, M.; Dogan, B.; Rishniw, M.; Weitzman, G.; Bosworth, B.; Yantiss, R.; Orsi, R.H.; Wiedmann, M.; McDonough, P.; Kim, S.G.; et al. Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of Clostridiales in Crohn’s disease involving the ileum. ISME J. 2007, 1, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Céspedes, S.; Saitz, W.; Del Canto, F.; De la Fuente, M.; Quera, R.; Hermoso, M.; Muñoz, R.; Ginard, D.; Khorrami, S.; Girón, J.; et al. Genetic Diversity and Virulence Determinants of Escherichia coli Strains Isolated from Patients with Crohn’s Disease in Spain and Chile. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Conte, M.; Longhi, C.; Marazzato, M.; Conte, A.L.; Aleandri, M.; Lepanto, M.S.; Zagaglia, C.; Nicoletti, M.; Aloi, M.; Totino, V.; et al. Adherent-invasive Escherichia coli (AIEC) in pediatric Crohn’s disease patients: Phenotypic and genetic pathogenic features. BMC Res. Notes 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Dogan, B.; Scherl, E.; Bosworth, B.; Yantiss, R.; Altier, C.; McDonough, P.L.; Jiang, Z.D.; DuPont, H.L.; Garneau, P.; Harel, J.; et al. Multidrug Resistance is Common in Escherichia coli Associated with Ileal Crohnʼs Disease. Inflamm. Bowel Dis. 2013, 19, 141–150. [Google Scholar] [CrossRef]
- Martinez-Medina, M.; Aldeguer, X.; Lopez-Siles, M.; González-Huix, F.; López-Oliu, C.; Dahbi, G.; Blanco, J.E.; Blanco, J.; Garcia-Gil, L.J.; Darfeuille-Michaud, A. Molecular diversity of Escherichia coli in the human gut: New ecological evidence supporting the role of adherent-invasive E. coli (AIEC) in Crohnʼs disease. Inflamm. Bowel Dis. 2009, 15, 872–882. [Google Scholar] [CrossRef]
- Negroni, A.; Costanzo, M.; Vitali, R.; Superti, F.; Bertuccini, L.; Tinari, A.; Minelli, F.; Di Nardo, G.; Nuti, F.; Pierdomenico, M.; et al. Characterization of adherent-invasive Escherichia coli isolated from pediatric patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2012, 18, 913–924. [Google Scholar] [CrossRef]
- Raso, T.; Crivellaro, S.; Chirillo, M.G.; Pais, P.; Gaia, E.; Savoia, D. Analysis of Escherichia coli Isolated from Patients Affected by Crohn’s Disease. Curr. Microbiol. 2011, 63, 131–137. [Google Scholar] [CrossRef]
- Sasaki, M.; Sitaraman, S.V.; Babbin, B.A. Invasive Escherichia coli are a feature of Crohn’s disease. Lab. Investig. 2007, 87, 1042–1054. [Google Scholar] [CrossRef]
- Čurová, K.; Kmeťová, M.; Sabol, M.; Gombošová, L.; Lazúrová, I.; Siegfried, L. Enterovirulent E. coli in inflammatory and noninflammatory bowel diseases. Folia Microbiol. 2009, 54, 81–86. [Google Scholar] [CrossRef]
- Prorok-Hamon, M.; Friswell, M.K.; Alswied, A.; Roberts, C.L.; Song, F.; Flanagan, P.K.; Knight, P.; Codling, C.; Marchesi, J.R.; Winstanley, C.; et al. Colonic mucosa-associated diffusely adherent afaC+ Escherichia coli expressing lpfA and pks are increased in inflammatory bowel disease and colon cancer. Gut 2014, 63, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Sobieszczańska, B.; Duda-Madej, A.; Turniak, M.; Franiczek, R.; Kasprzykowska, U.; Duda, A.K.; Rzeszutko, M.; Iwańczak, B. Invasive Properties, Adhesion Patterns and Phylogroup Profiles Among Escherichia coli Strains Isolated from Children with Inflammatory Bowel Disease. Adv. Clin. Exp. Med. 2012, 21, 591–599. [Google Scholar] [PubMed]
- Darfeuille-Michaud, A.; Boudeau, J.; Bulois, P.; Neut, C.; Glasser, A.L.; Barnich, N.; Bringer, M.A.; Swidsinski, A.; Beaugerie, L.; Colombel, J.F. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology 2004, 127, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Palmela, C.; Chevarin, C.; Xu, Z.; Torres, J.; Sevrin, G.; Hirten, R.; Barnich, N.; Siew, C.; Colombel, J.F. Adherent-invasive Escherichia coli in inflammatory bowel disease. Gut 2017, 67, 574–587. [Google Scholar] [CrossRef]
- Baumgart, D.C.; Sandborn, W.J. Crohn’s disease. Lancet 2012, 380, 1590–1605. [Google Scholar] [CrossRef]
- Hansen, J.J.; Sartor, R.B. Therapeutic Manipulation of the Microbiome in IBD: Current Results and Future Approaches. Curr. Treat. Options Gastroenterol. 2015, 13, 105–120. [Google Scholar] [CrossRef]
- Fedorak, R.N.; Ismond, K.P. Practical Considerations and the Intestinal Microbiome in Disease: Antibiotics for IBD Therapy. Dig. Dis. 2016, 34, 112–121. [Google Scholar] [CrossRef]
- Subramanian, S.; Roberts, C.L.; Hart, C.A.; Martin, H.M.; Edwards, S.W.; Rhodes, J.M.; Campbell, B.J. Replication of Colonic Crohn’s Disease Mucosal Escherichia coli Isolates within Macrophages and Their Susceptibility to Antibiotics. Antimicrob. Agents Chemother. 2008, 52, 427–434. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Miquel, S.; Peyretaillade, E.; Claret, L.; de Vallée, A.; Dossat, C.; Vacherie, B.; Zineb, H.; Segurens, B.; Barbe, V.; Sauvanet, P.; et al. Complete Genome Sequence of Crohn’s Disease-Associated Adherent-Invasive E. coli Strain LF82. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Caroff, N.; Espaze, E.; Gautreau, D.; Richet, H.; Reynaud, A. Analysis of the effects of -42 and -32 ampC promoter mutations in clinical isolates of Escherichia coli hyperproducing AmpC. J. Antimicrob. Chemother. 2000, 45, 783–788. [Google Scholar] [CrossRef] [PubMed]
- Clarke, B.; Hiltz, M.; Musgrave, H.; Forward, K.R. Cephamycin Resistance in Clinical Isolates and Laboratory-derived Strains of Escherichia coli, Nova Scotia, Canada. Emerg. Infect. Dis. 2003, 9, 1254–1259. [Google Scholar] [CrossRef] [PubMed]
- Forward, K.R.; Willey, B.M.; Low, D.E.; McGeer, A.; Kapala, M.A.; Kapala, M.M.; Burrows, L.L. Molecular mechanisms of cefoxitin resistance in Escherichia coli from the Toronto area hospitals. Diagn. Microbiol. Infect. Dis. 2001, 41, 57–63. [Google Scholar] [CrossRef]
- Tracz, D.M.; Boyd, D.A.; Bryden, L.; Hizon, R.; Giercke, S.; Van Caeseele, P.; Mulvey, M.R. Increase in ampC promoter strength due to mutations and deletion of the attenuator in a clinical isolate of cefoxitin-resistant Escherichia coli as determined by RT–PCR. J. Antimicrob. Chemother. 2005, 55, 768–772. [Google Scholar] [CrossRef][Green Version]
- Perna, A.; Hay, E.; Contieri, M.; De Luca, A.; Guerra, G.; Lucariello, A. Adherent-invasive Escherichia coli (AIEC): Cause or consequence of inflammation, dysbiosis, and rupture of cellular joints in patients with IBD? J. Cell. Physiol. 2020, 235, 5041–5049. [Google Scholar] [CrossRef]
- Shaler, C.R.; Elhenawy, W.; Coombes, B.K. The Unique Lifestyle of Crohn’s Disease-Associated Adherent-Invasive Escherichia coli. J. Mol. Biol. 2019, 431, 2970–2981. [Google Scholar] [CrossRef]
- Barrios-Villa, E.; Cortés-Cortés, G.; Lozano-Zaraín, P.; Arenas-Hernández, M.M.P.; Martínez de la Peña, C.F.; Martínez-Laguna, Y.; Torres, C.; Rocha-Gracia, R.D.C. Adherent/invasive Escherichia coli (AIEC) isolates from asymptomatic people: New E. coli ST131 O25:H4/H30-Rx virotypes. Ann. Clin. Microbiol. Antimicrob. 2018, 17. [Google Scholar] [CrossRef]
- Gündoğdu, A.; Long, Y.B.; Vollmerhausen, T.L.; Katouli, M. Antimicrobial resistance and distribution of sul genes and integron-associated intI genes among uropathogenic Escherichia coli in Queensland, Australia. J. Med. Microbiol. 2011, 60, 1633–1642. [Google Scholar] [CrossRef]
- Koczura, R.; Mokracka, J.; Barczak, A.; Krysiak, N.; Kaznowski, A. Association between the Presence of Class 1 Integrons, Virulence Genes, and Phylogenetic Groups of Escherichia coli Isolates from River Water. Microb. Ecol. 2013, 65, 84–90. [Google Scholar] [CrossRef]
- Elliott, T.R.; Hudspith, B.N.; Wu, G. Quantification and Characterization of Mucosa-Associated and Intracellular Escherichia coli in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2013, 19, 2326–2338. [Google Scholar] [CrossRef]
- Dogan, B.; Fu, J.; Zhang, S.; Scherl, E.J.; Simpson, K.W. Rifaximin decreases virulence of Crohn’s disease-associated Escherichia coli and epithelial inflammatory responses. J. Antibiot. 2018, 71, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Virta, L.; Auvinen, A.; Helenius, H.; Huovinen, P.; Kolho, K.L. Association of Repeated Exposure to Antibiotics With the Development of Pediatric Crohn’s Disease-A Nationwide, Register-based Finnish Case-Control Study. Am. J. Epidemiol. 2012, 175, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Oberc, A.M.; Fiebig-Comyn, A.A.; Tsai, C.N.; Elhenawy, W.; Coombes, B.K. Antibiotics Potentiate Adherent-Invasive E. coli Infection and Expansion. Inflamm. Bowel Dis. 2019, 25, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Mydock-McGrane, L.K.; Hannan, T.J.; Janetka, J.W. Rational design strategies for FimH antagonists: New drugs on the horizon for urinary tract infection and Crohn’s disease. Expert Opin. Drug Discov. 2017, 12, 711–731. [Google Scholar] [CrossRef]
- Brown, C.L.; Smith, K.; Wall, D.M.; Walker, D. Activity of Species-specific Antibiotics Against Crohnʼs Disease–Associated Adherent-invasive Escherichia coli. Inflamm. Bowel Dis. 2015, 1. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard, Ninth Edition. In CLSI Document M07-A9; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing; 29th informational supplement. In CLSI Document M100-S29; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- European Society of Clinical Microbiology and Infectious Diseases. EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance Version 1.0. 2013. EUCAST. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Resistance_mechanisms/EUCAST_detection_of_resistance_mechanisms_v1.0_20131211.pdf (accessed on 10 March 2020).
- Perez-Perez, F.J.; Hanson, N.D. Detection of Plasmid-Mediated AmpC -Lactamase Genes in Clinical Isolates by Using Multiplex PCR. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef]
- Camprubí-Font, C.; Lopez-Siles, M.; Ferrer-Guixeras, M.; Niubó-Carulla, L.; Abellà-Ametller, C.; Garcia-Gil, L.J.; Martinez-Medina, M. Comparative genomics reveals new single-nucleotide polymorphisms that can assist in identification of adherent-invasive Escherichia coli. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Clarke, D.J.; Chaudhuri, R.R.; Martin, H.M.; Campbell, B.J.; Rhodes, J.M.; Constantinidou, C.; Pallen, M.J.; Loman, N.J.; Cunningham, A.F.; Browning, D.F.; et al. Complete Genome Sequence of the Crohn’s Disease-Associated Adherent-Invasive Escherichia coli Strain HM605. J. Bacteriol. 2011, 193. [Google Scholar] [CrossRef]
- Dogan, B.; Suzuki, H.; Herlekar, D.; Sartor, R.B.; Campbell, B.J.; Roberts, C.L.; Stewart, K.; Scherl, E.J.; Araz, Y.; Bitar, P.P.; et al. Inflammation-associated Adherent-invasive Escherichia coli are Enriched in Pathways for Use of Propanediol and Iron and M-cell Translocation. Inflamm. Bowel Dis. 2014, 20, 1919–1932. [Google Scholar] [CrossRef]
- Krause, D.O.; Little, A.C.; Dowd, S.E.; Bernstein, C.N. Complete genome sequence of adherent invasive Escherichia coli UM146 Isolated from Ileal Crohn’s disease biopsy tissue. J. Bacteriol. 2011, 193. [Google Scholar] [CrossRef]
- Nash, J.H.; Villegas, A.; Kropinski, A.M.; Aguilar-Valenzuela, R.; Konczy, P.; Mascarenhas, M.; Ziebell, K.; Torres, A.G.; Karmali, M.A.; Coombes, B.K. Genome sequence of adherent-invasive Escherichia coli and comparative genomic analysis with other E. coli pathotypes. BMC Genom. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- National Institute of Health. Human Microbial Project. Available online: http://www.hmpdacc.org/HMRGD/#data (accessed on 10 March 2020).
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11. [Google Scholar] [CrossRef]
- Arango-Argoty, G.; Garner, E.; Pruden, A.; Heath, L.S.; Vikesland, P.; Zhang, L. DeepARG: A deep learning approach for predicting antibiotic resistance genes from metagenomic data. Microbiome 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Cock, P.J.; Antao, T.; Chang, J.T.; Chapman, B.A.; Cox, C.J.; Dalke, A.; Friedberg, I.; Hamelryck, T.; Kauff, F.; Wilczynski, B.; et al. Biopython: Freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 2009, 25, 1422–1423. [Google Scholar] [CrossRef] [PubMed]
- Di Tommaso, P.; Chatzou, M.; Floden, E.W.; Barja, P.P.; Palumbo, E.; Notredame, C. Nextflow enables reproducible computational workflows. Nat. Biotechnol. 2017, 35, 316–319. [Google Scholar] [CrossRef] [PubMed]


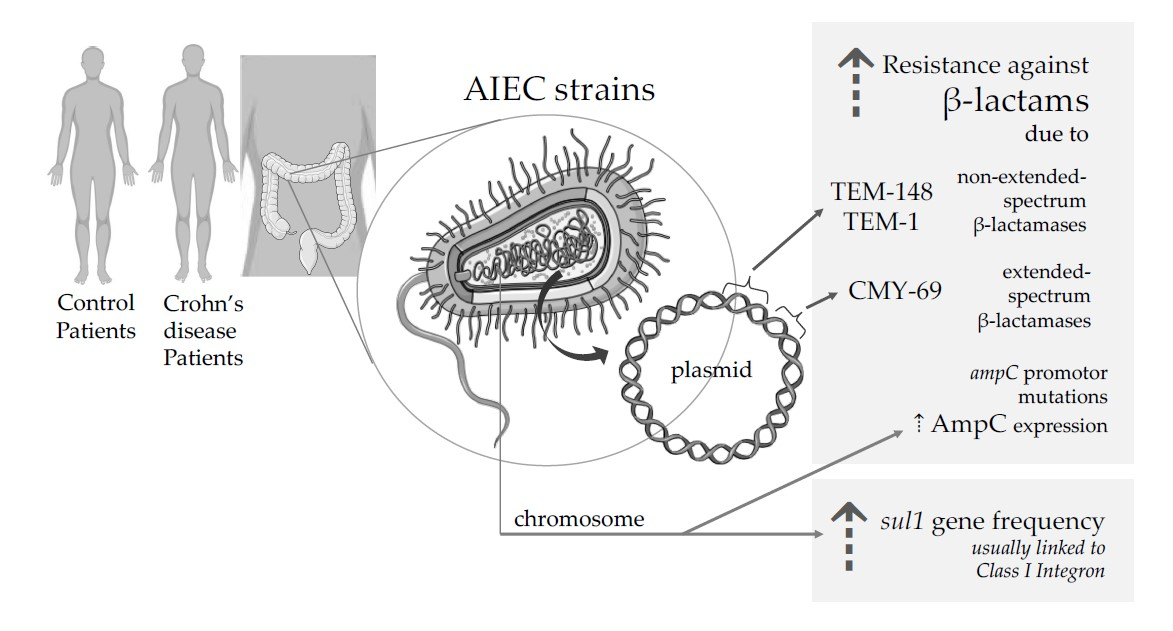{kind=link}
{kind=link}
| Strain | Pathotype | Phylogroup | Disease | Ampicillin | Ticarcillin | Amoxicillin-clavulanate | Piperacillin-tazobactam | Ticarcillin-clavulanate | Cefazolin | Cefuroxime | Cefotaxime | Ceftazidime | Cefpodoxime | Cefovecin | Ceftiofur | Cefoxitin | Tetracycline | Doxycycline | Kanamycin | Streptomycin | Nalidixic acid | Ciprofloxacin | Enrofloxacin | Marbofloxacin | TMP/SMX | Chloramphenicol | Rifampin§ |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AIEC10 | AIEC | A | C | R | R | S | S | S | S | R | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | >2 |
| AIEC19 | AIEC | A | C | R | R | S | S | S | S | S | S | S | S | S | S | S | R | I | S | R | S | S | S | S | S | S | >2 |
| AIEC23 | AIEC | A | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | R | R | S | S | S | S | S | S | S | S | >2 |
| AIEC24 | AIEC | A | CD | R | R | R | R | R | I | S | S | S | S | S | S | S | R | R | S | S | R | R | R | R | R | R | >2 |
| AIEC07 | AIEC | B1 | C | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | >2 |
| AIEC04 | AIEC | B2 | C | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | 2 |
| AIEC06 | AIEC | B2 | C | S | S | S | S | S | S | S | S | S | S | S | S | S | R | R | S | I | S | S | S | S | S | S | <1 |
| AIEC08 | AIEC | B2 | C | R | I | R | S | I | I | R | R | R | R | R | R | R | S | S | S | I | S | S | S | S | S | S | 2 |
| AIEC01 | AIEC | B2 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | >2 |
| AIEC02 | AIEC | B2 | CD | R | R | R | S | I | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | R | >2 |
| AIEC05 | AIEC | B2 | CD | R | R | S | R | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | S | S | S | 2 |
| AIEC09 | AIEC | B2 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | >2 |
| AIEC11 | AIEC | B2 | CD | R | R | R | R | I | I | R | S | S | S | S | S | S | R | R | S | R | S | S | S | S | R | S | >2 |
| AIEC12 | AIEC | B2 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | >2 |
| AIEC14-1 | AIEC | B2 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | >2 |
| AIEC15-1 | AIEC | B2 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | >2 |
| AIEC16-2 | AIEC | B2 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | >2 |
| AIEC21 | AIEC | B2 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | >2 |
| AIEC25 | AIEC | B2 | CD | R | R | R | S | I | S | S | S | S | S | S | S | S | R | R | S | R | S | S | S | S | S | S | >2 |
| LF82 | AIEC | B2 | CD | R | R | R | R | R | R | R | R | R | R | R | I | R | S | S | S | S | S | S | S | S | S | S | >2 |
| AIEC17 | AIEC | D | CD | R | R | S | S | S | S | S | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | 2 |
| AIEC20 | AIEC | D | CD | R | R | S | S | I | S | S | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | 2 |
| ECG16 | NON AIEC | A | C | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | R | S | 2 |
| ECG22 | NON AIEC | A | C | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | I | S | S | S | S | S | S | 2 |
| ECG18 | NON AIEC | A | CD | R | R | S | S | S | S | S | S | S | S | S | S | S | S | S | S | R | R | R | R | S | S | S | >2 |
| ECG19 | NON AIEC | A | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | >2 |
| ECG65 | NON AIEC | A | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | >2 |
| ECG23 | NON AIEC | At. | C | R | R | S | S | S | S | S | S | S | S | S | S | S | R | I | S | R | S | S | S | S | R | S | >2 |
| ECG04 | NON AIEC | B1 | C | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | >2 |
| ECG46 | NON AIEC | B1 | C | S | S | S | S | S | S | S | S | S | S | S | S | S | R | I | S | S | R | S | S | S | S | S | 2 |
| ECG02 | NON AIEC | B1 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | I | S | S | S | S | S | S | S | S | >2 |
| ECG21 | NON AIEC | B1 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | R | I | S | R | S | S | S | S | S | S | >2 |
| ECG63 | NON AIEC | B1 | CD | S | S | S | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | S | S | S | S | >2 |
| ECG64 | NON AIEC | B1 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | >2 |
| ECG08 | NON AIEC | B2 | C | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | 2 |
| ECG12 | NON AIEC | B2 | C | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | >2 |
| ECG13 | NON AIEC | B2 | C | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S | S | >2 |
| ECG17 | NON AIEC | B2 | C | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | 2 |
| ECG41 | NON AIEC | B2 | C | R | R | R | S | I | S | S | S | S | S | S | S | S | S | S | S | I | S | S | S | S | S | S | 2 |
| ECG43 | NON AIEC | B2 | C | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | 2 |
| ECG49 | NON AIEC | B2 | C | S | S | S | S | S | S | S | S | S | S | S | S | S | R | R | I | R | S | S | S | S | S | S | 2 |
| ECG01 | NON AIEC | B2 | CD | R | R | R | S | I | S | S | S | S | S | S | S | S | R | R | S | R | S | S | S | S | S | S | 2 |
| ECG05 | NON AIEC | B2 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | 2 |
| ECG09 | NON AIEC | B2 | CD | R | R | S | S | S | S | S | S | S | S | S | S | S | R | I | S | R | S | S | S | S | R | R | 2 |
| ECG15 | NON AIEC | B2 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | 2 |
| ECG26 | NON AIEC | B2 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | R | S | S | S | S | S | >2 |
| ECG42 | NON AIEC | B2 | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | S | 2 |
| ECG34 | NON AIEC | D | CD | S | S | S | S | S | S | S | S | S | S | S | S | S | R | I | S | S | S | S | S | S | S | S | >2 |
| ECG57 | NON AIEC | D | CD | R | R | S | S | S | S | S | S | S | S | S | S | S | R | R | S | R | S | S | S | S | R | S | 2 |
| Antimicrobial Category | Antimicrobial | All Strains (n = 49) | Non-AIEC (n = 27) | AIEC (n = 22) | p | C-E. coli (n = 18) | CD-E. coli (n = 31) | p |
|---|---|---|---|---|---|---|---|---|
| β-lactams | ||||||||
| Penicillins | Ampicillin | 17 | 22.2 | 50 | 0.042 | 27.8 | 38.7 | ns |
| Ticarcillin | 16 | 22.2 | 50 | 0.042 | 27.8 | 38.7 | ns | |
| Penicillins+ β-lactamase inhibitors | Amoxicillin-clavulanate | 8 | 7.4 | 31.8 | 0.034 | 11.1 | 22.6 | ns |
| Piperacillin-tazobactam | 4 | 0 | 18.2 | 0.035 | 0 | 12.9 | ns | |
| Ticarcillin-clavulanate | 9 | 7.4 | 31.8 | 0.034 | 11.1 | 22.6 | ns | |
| Non expanded spectrum cephalosporins (1st and 2nd gen.) | Cefazolin | 4 | 0 | 18.2 | 0.035 | 5.6 | 9.7 | ns |
| Cefuroxime | 4 | 0 | 18.2 | 0.035 | 11.1 | 6.5 | ns | |
| Expanded spectrum cephalosporines (3d and 4th gen.) | Cefotaxime | 2 | 0 | 9.1 | ns | 5.6 | 3.2 | ns |
| Ceftazidime | 2 | 0 | 9.1 | ns | 5.6 | 3.2 | ns | |
| Cefpodoxime | 2 | 0 | 9.1 | ns | 5.6 | 3.2 | ns | |
| Cefovecin | 2 | 0 | 9.1 | ns | 5.6 | 3.2 | ns | |
| Ceftiofur | 2 | 0 | 9.1 | ns | 5.6 | 3.2 | ns | |
| Cefepime | 0 | 0 | 0 | ns | 0 | 0 | ns | |
| Cephamycins | Cefoxitin | 4 | 0 | 13.6 | 0.084 | 5.6 | 6.5 | ns |
| Carbapenems | Ertapenem | 0 | 0 | 0 | ns | 0 | 0 | ns |
| Imipenem | 0 | 0 | 0 | ns | 0 | 0 | ns | |
| Tetracyclines | Tetracycline | 15 | 29.6 | 31.8 | ns | 27.8 | 32.3 | ns |
| Doxycycline | 15 | 33.3 | 27.3 | ns | 27.8 | 32.3 | ns | |
| Aminoglycosides | Amikacin | 0 | 0 | 0 | ns | 0 | 0 | ns |
| Gentamicin | 0 | 0 | 0 | ns | 0 | 0 | ns | |
| Kanamycin | 1 | 3.7 | 0 | ns | 5.6 | 0 | ns | |
| Streptomycin | 22 | 40.7 | 50 | ns | 55.6 | 38.7 | ns | |
| Quinolones | Nalidixic acid | 4 | 11.1 | 4.5 | ns | 5.6 | 9.7 | ns |
| Fluoroquinolones | Ciprofloxacin | 2 | 3.7 | 4.5 | ns | 0 | 6.5 | ns |
| Enrofloxacin | 2 | 3.7 | 4.5 | ns | 0 | 6.5 | ns | |
| Marbofloxacin | 1 | 3.7 | 4.5 | ns | 0 | 6.5 | ns | |
| Folate Pathway Inhibitors | Trimethoprim-sulfamethoxazole | 6 | 14.8 | 9.1 | ns | 11.1 | 12.9 | ns |
| Phenicols | Chloramphenicol | 3 | 3.7 | 9.1 | ns | 0 | 9.7 | ns |
| Glycylcyclines | Tigecycline | 0 | 0 | 0 | ns | 0 | 0 | ns |
| Positions at the ampC Promotor with SNPs | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | Resistance Profile to β-Lactams | Mechanism of Resistance | −88 | −82 | −73 | −32 | −28 | −18 | −1 | +17 | +23 | +58 | Variant |
| K12 | C | A | C | T | G | G | C | C | G | C | 1 | ||
| LF82 | expanded-spectrum cephalosporins | Mutations in ampC promotor | C | A | T | A | A | G | C | T | A | C | 2 |
| AIEC08 | expanded-spectrum cephalosporins | BLEE (CMY-69) | - | - | - | - | - | - | C | C | G | C | nr |
| AIEC02 | not expanded-spectrum cephalosporins | Mechanism not deciphered | C | A | C | T | G | G | C | C | G | C | 1 |
| AIEC11 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-148) | C | A | C | T | G | G | C | C | G | C | 1 |
| AIEC19 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-148) | C | A | C | T | G | G | C | C | G | C | 1 |
| AIEC20 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-148) | C | A | C | T | G | G | C | C | G | C | 1 |
| AIEC24 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-148) | C | A | C | T | G | G | C | C | G | C | 1 |
| ECG18 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-148) | C | A | C | T | G | G | C | C | G | C | 1 |
| ECG57 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-1) | C | A | C | T | G | G | C | C | G | C | 1 |
| ECG09 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-148) | C | A | T | T | A | G | C | C | G | C | 3 |
| AIEC05 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-148) | C | A | T | T | A | G | C | T | G | C | 4 |
| ECG41 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-148) | C | A | T | T | G | G | C | C | G | C | 5 |
| AIEC17_E1 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-148) | C | A | T | T | G | G | C | C | G | T | 6 |
| AIEC25 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-148) | C | A | T | T | G | G | C | C | G | T | 6 |
| ECG01 | not expanded-spectrum cephalosporins | TEM β-lactamase (TEM-148) | C | A | T | T | G | G | C | C | G | T | 6 |
| AIEC10 | not expanded-spectrum cephalosporins | Mutations in ampC promotor | T | G | C | T | G | A | T | C | G | T | 7 |
| AIEC12 | not resistant to β-lactams | none | C | A | C | T | G | G | C | C | G | C | 1 |
| AIEC23 | not resistant to β-lactams | none | C | A | C | T | G | G | C | C | G | C | 1 |
| ECG16 | not resistant to β-lactams | none | C | A | C | T | G | G | C | C | G | C | 1 |
| ECG19 | not resistant to β-lactams | none | C | A | C | T | G | G | C | C | G | C | 1 |
| ECG34 | not resistant to β-lactams | none | C | A | C | T | G | G | C | C | G | C | 1 |
| ECG42 | not resistant to β-lactams | none | C | A | C | T | G | G | C | C | G | C | 1 |
| ECG46 | not resistant to β-lactams | none | C | A | C | T | G | G | C | C | G | C | 1 |
| ECG65 | not resistant to β-lactams | none | C | A | C | T | G | G | C | C | G | C | 1 |
| ECG26 | not resistant to β-lactams | none | C | A | T | T | A | G | C | T | G | C | 4 |
| ECG43 | not resistant to β-lactams | none | C | A | T | T | A | G | C | T | G | C | 4 |
| AIEC01_E4 | not resistant to β-lactams | none | C | A | T | T | G | G | C | C | G | C | 5 |
| AIEC04 | not resistant to β-lactams | none | C | A | T | T | G | G | C | C | G | C | 5 |
| AIEC06 | not resistant to β-lactams | none | C | A | T | T | G | G | C | C | G | C | 5 |
| AIEC09 | not resistant to β-lactams | none | C | A | T | T | G | G | C | C | G | C | 5 |
| AIEC14_1 | not resistant to β-lactams | none | C | A | T | T | G | G | C | C | G | C | 5 |
| AIEC15_1 | not resistant to β-lactams | none | C | A | T | T | G | G | C | C | G | C | 5 |
| AIEC16_2 | not resistant to β-lactams | none | C | A | T | T | G | G | C | C | G | C | 5 |
| AIEC21 | not resistant to β-lactams | none | C | A | T | T | G | G | C | C | G | C | 5 |
| ECG17 | not resistant to β-lactams | none | C | A | T | T | G | G | C | C | G | C | 5 |
| ECG05 | not resistant to β-lactams | none | C | A | T | T | G | G | C | C | G | T | 6 |
| ECG15 | not resistant to β-lactams | none | C | A | T | T | G | G | C | C | G | T | 6 |
| AIEC07_E6 | not resistant to β-lactams | none | T | G | C | T | G | A | T | C | G | T | 7 |
| ECG02 | not resistant to β-lactams | none | T | G | C | T | G | A | T | C | G | T | 7 |
| ECG04 | not resistant to β-lactams | none | T | G | C | T | G | A | T | C | G | T | 7 |
| ECG21 | not resistant to β-lactams | none | T | G | C | T | G | A | T | C | G | T | 7 |
| ECG63 | not resistant to β-lactams | none | T | G | C | T | G | A | T | C | G | T | 7 |
| ECG64 | not resistant to β-lactams | none | T | G | C | T | G | A | T | C | G | T | 7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinez-Medina, M.; Strozzi, F.; Ruiz Del Castillo, B.; Serrano-Morillas, N.; Ferrer Bustins, N.; Martínez-Martínez, L. Antimicrobial Resistance Profiles of Adherent Invasive Escherichia coli Show Increased Resistance to β-Lactams. Antibiotics 2020, 9, 251. https://doi.org/10.3390/antibiotics9050251
Martinez-Medina M, Strozzi F, Ruiz Del Castillo B, Serrano-Morillas N, Ferrer Bustins N, Martínez-Martínez L. Antimicrobial Resistance Profiles of Adherent Invasive Escherichia coli Show Increased Resistance to β-Lactams. Antibiotics. 2020; 9(5):251. https://doi.org/10.3390/antibiotics9050251
Chicago/Turabian StyleMartinez-Medina, Margarita, Francesco Strozzi, Belén Ruiz Del Castillo, Natalia Serrano-Morillas, Nuria Ferrer Bustins, and Luis Martínez-Martínez. 2020. "Antimicrobial Resistance Profiles of Adherent Invasive Escherichia coli Show Increased Resistance to β-Lactams" Antibiotics 9, no. 5: 251. https://doi.org/10.3390/antibiotics9050251
APA StyleMartinez-Medina, M., Strozzi, F., Ruiz Del Castillo, B., Serrano-Morillas, N., Ferrer Bustins, N., & Martínez-Martínez, L. (2020). Antimicrobial Resistance Profiles of Adherent Invasive Escherichia coli Show Increased Resistance to β-Lactams. Antibiotics, 9(5), 251. https://doi.org/10.3390/antibiotics9050251