Overview of Evidence of Antimicrobial Use and Antimicrobial Resistance in the Food Chain

 , , ,
, , , 
Abstract
1. Introduction
2. Results
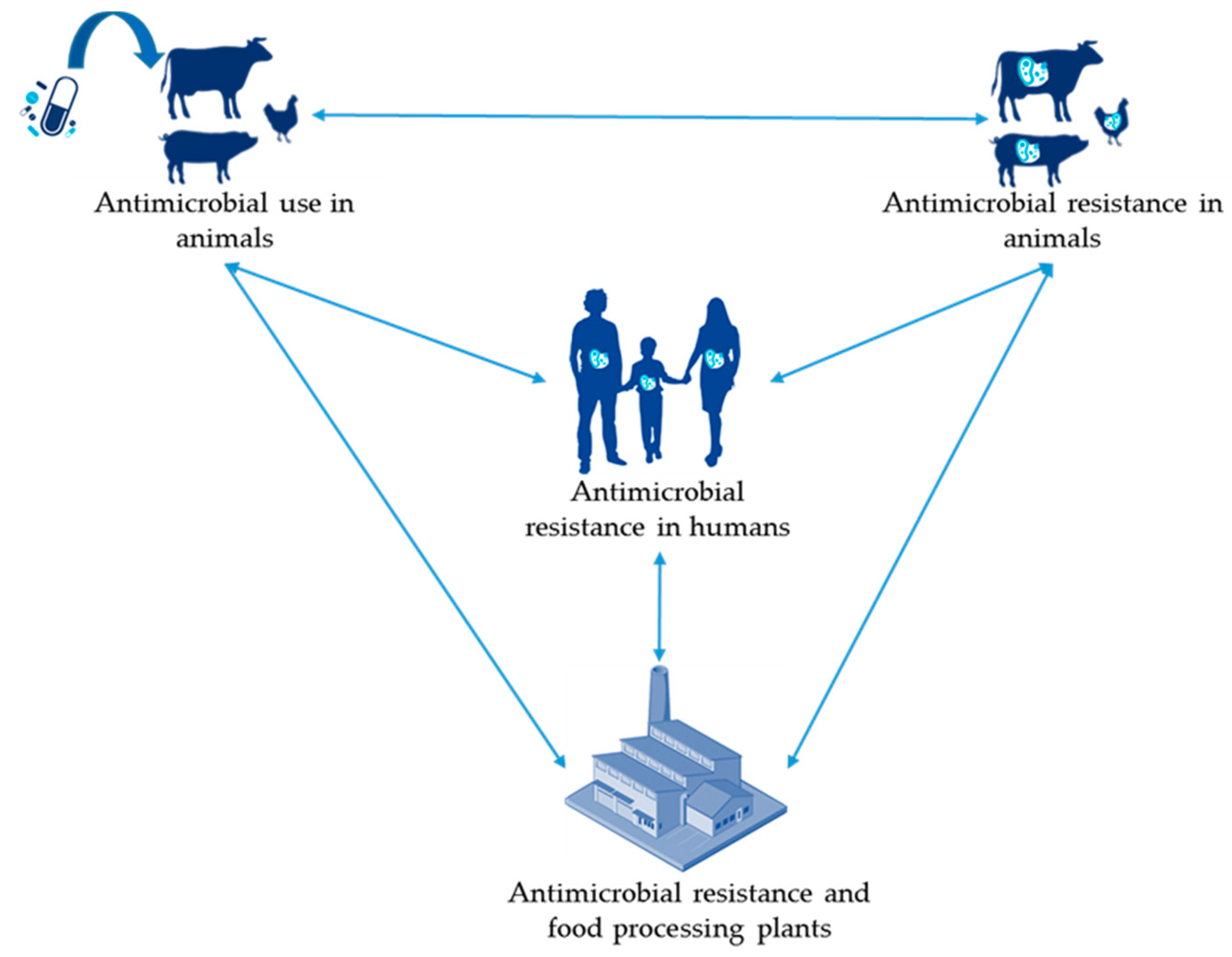
2.1. Antimicrobial Use and Antimicrobial Resistance in Animals
2.2. Antimicrobial Use in Animals and Antimicrobial Resistance in Humans
2.2.1. Molecular Studies Investigating Links between Resistant Bacteria or Genes in Animals and Their Transmission to Humans
2.2.2. Presence of AMR Bacteria in Food Products of Animal Origin
2.2.3. Association between Antimicrobial Use in Food Animals and Resistant Bacteria in Humans
2.2.4. Studies with Limited Evidence of an Association between AMR in Humans and Food-Producing Animals
2.3. Antimicrobial Resistance in Bacteria Linked to Food Processing Practices
3. Discussion
4. Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- O’Neill, J. Tackling dRug-Resistant Infections Globally: Final Report and Recommendations. 2016. Available online: https://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf (accessed on 15 May 2017).
- World Health Organization. Global Action Plan on Antimicrobial Resistance. 2015. Available online: https://apps.who.int/iris/bitstream/handle/10665/193736/9789241509763_eng.pdf?sequence=1&isAllowed=y (accessed on 15 May 2017).
- European Commission. A European One Health Action Plan Against Antimicrobial Resistance (amr). 2017. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:52017DC0339&from=EN (accessed on 25 April 2019).
- Boerlin, P.; Reid-Smith, R.J. Antimicrobial resistance: Its emergence and transmission. Anim. Health Res. Rev. 2008, 9, 115–126. [Google Scholar] [CrossRef]
- D’Costa, V.M.; King, C.E.; Kalan, L.; Morar, M.; Sung, W.W.; Schwarz, C.; Froese, D.; Zazula, G.; Calmels, F.; Debruyne, R.; et al. Antibiotic resistance is ancient. Nature 2011, 477, 457–461. [Google Scholar] [CrossRef]
- Aarestrup, F.M. The livestock reservoir for antimicrobial resistance: A personal view on changing patterns of risks, effects of interventions and the way forward. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140085. [Google Scholar] [CrossRef]
- Andremont, A.; Karp, B.; DeWaal, C.S.; Mitema, E.; Matar, G.; Woo, G.-J.; Scott, H.M.; Watanabe, H.; Fernandez, H.; Conly, J.; et al. World health organization ranking of antimicrobials according to their importance in human medicine: A critical step for developing risk management strategies to control antimicrobial resistance from food animal production. Clin. Infect. Dis. 2016, 63, 1087–1093. [Google Scholar]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism mcr-1 in animals and human beings in china: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Rushton, J.; Pinto Ferreira, J.; Stärk, K. Antimicrobial resistance: The use of antimicrobials in the livestock sector. OECD Food Agric. Fisheries Pap. 2014, 68, 6–12. [Google Scholar] [CrossRef]
- Castanon, J.I. History of the use of antibiotic as growth promoters in european poultry feeds. Poult. Sci. 2007, 86, 2466–2471. [Google Scholar] [CrossRef]
- Anon. Regulation (ec) no 1831/2003 of the european parliament and of the council of 22 september 2003 on additives for use in animal nutrition. Off. J. Eur. Union. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32003R1831&from=EN (accessed on 15 May 2017).
- Chantziaras, I.; Boyen, F.; Callens, B.; Dewulf, J. Correlation between veterinary antimicrobial use and antimicrobial resistance in food-producing animals: A report on seven countries. J. Antimicrob. Chemother. 2013, 69, 827–834. [Google Scholar] [CrossRef]
- ECDC/EFSA/EMA. First joint report on the integrated analysis of the consumption of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from humans and food-producing animals. Efsa J. 2015, 13. [Google Scholar]
- ECDC/EFSA/EMA. Second joint report on the integrated analysis of the consumption of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from humans and food-producing animals: Joint interagency antimicrobial consumption and resistance analysis (jiacra) report. Efsa J. 2017, 15. [Google Scholar]
- Smith, T.C.; Gebreyes, W.A.; Abley, M.J.; Harper, A.L.; Forshey, B.M.; Male, M.J.; Martin, H.W.; Molla, B.Z.; Sreevatsan, S.; Thakur, S.; et al. Methicillin-resistant Staphylococcus aureus in pigs and farm workers on conventional and antibiotic-free swine farms in the USA. PLoS ONE 2013, 8, e63704. [Google Scholar] [CrossRef]
- MARAN. Monitoring of Antimicrobial Resistance and Antibiotic Usage in Animals in the Netherlands in 2016. Available online: https://www.wur.nl/upload_mm/6/9/5/4f37c335-224c-4595-82e4-be6182c0a5e1_74ce6009-b112-428d-aeb7-99b95063aab6_Maran%20report%202017.pdf (accessed on 14 December 2017).
- European Medicines Agency. Trends in the Sales of Veterinary Antimicrobial Agents in Nine European Countries (2005–2009). Available online: https://www.ema.europa.eu/en/documents/report/trends-sales-veterinary-antimicrobial-agents-nine-european-countries_en.pdf (accessed on 15 May 2017).
- Dorado-Garcia, A.; Mevius, D.J.; Jacobs, J.J.; Van Geijlswijk, I.M.; Mouton, J.W.; Wagenaar, J.A.; Heederik, D.J. Quantitative assessment of antimicrobial resistance in livestock during the course of a nationwide antimicrobial use reduction in the netherlands. J. Antimicrob. Chemother. 2016, 71, 3607–3619. [Google Scholar] [CrossRef]
- Institute of Food Technologists. Antimicrobial resistance: Implications for the food system. Comp. Rev. Food Sci. Food Saf. 2006, 5, 71–137. [Google Scholar] [CrossRef]
- DANMAP. Use of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Bacteria from Food Animals, Food and Humans in Denmark. 2015. Available online: https://www.danmap.org/-/media/arkiv/projekt-sites/danmap/danmap-reports/danmap--2015/danmap-2015.pdf?la=en (accessed on 15 May 2017).
- Agersø, Y.; Aarestrup, F.M. Voluntary ban on cephalosporin use in Danish pig production has effectively reduced extended-spectrum cephalosporinase-producing Escherichia coli in slaughter pigs. J. Antimicrob. Chemother. 2013, 68, 569–572. [Google Scholar] [CrossRef]
- Callens, B.; Cargnel, M.; Sarrazin, S.; Dewulf, J.; Hoet, B.; Vermeersch, K.; Wattiau, P.; Welby, S. Associations between a decreased veterinary antimicrobial use and resistance in commensal Escherichia coli from Belgian livestock species (2011–2015). Prev. Vet. Med. 2018, 157, 50–58. [Google Scholar] [CrossRef]
- World Health Organization. Critically Important Antimicrobials for Human Medicines—5th Revision 2016. Available online: https://apps.who.int/iris/bitstream/handle/10665/255027/9789241512220-eng.pdf?sequence=1 (accessed on 14 December 2017).
- Hammerum, A.M.; Heuer, O.E.; Emborg, H.-D.; Bagger-Skjøt, L.; Jensen, V.F.; Rogues, A.-M.; Skov, R.L.; Agersø, Y.; Brandt, C.T.; Seyfarth, A.M.; et al. Danish integrated antimicrobial resistance monitoring and research program. Emerg. Infect. Dis. 2007, 13, 1632–1639. [Google Scholar] [CrossRef]
- Aarestrup, F.M.; Seyfarth, A.M.; Emborg, H.D.; Pedersen, K.; Hendriksen, R.S.; Bager, F. Effect of abolishment of the use of antimicrobial agents for growth promotion on occurrence of antimicrobial resistance in fecal Enterococci from food animals in Denmark. Antimicrob. Agents Chemother. 2001, 45, 2054–2059. [Google Scholar] [CrossRef]
- McCrackin, M.A.; Helke, K.L.; Galloway, A.M.; Poole, A.Z.; Salgado, C.D.; Marriott, B.P. Effect of antimicrobial use in agricultural animals on drug-resistant foodborne campylobacteriosis in humans: A systematic literature review. Crit. Rev. Food Sci. Nutr. 2016, 56, 2115–2132. [Google Scholar] [CrossRef]
- Dutil, L.; Irwin, R.; Finley, R.; Ng, L.K.; Avery, B.; Boerlin, P.; Bourgault, A.M.; Cole, L.; Daignault, D.; Desruisseau, A.; et al. Ceftiofur resistance in Salmonella enterica serovar heidelberg from chicken meat and humans, Canada. Emerg. Infect. Dis. 2010, 16, 48–54. [Google Scholar] [CrossRef]
- Lazarus, B.; Paterson, D.L.; Mollinger, J.L.; Rogers, B.A. Do human extraintestinal Escherichia coli infections resistant to expanded-spectrum cephalosporins originate from food-producing animals? A systematic review. Clin. Infect. Dis. 2015, 60, 439–452. [Google Scholar] [CrossRef]
- Mather, A.E.; Reid, S.W.; Maskell, D.J.; Parkhill, J.; Fookes, M.C.; Harris, S.R.; Brown, D.J.; Coia, J.E.; Mulvey, M.R.; Gilmour, M.W.; et al. Distinguishable epidemics of multidrug-resistant Salmonella typhimurium dt104 in different hosts. Science 2013, 341, 1514–1517. [Google Scholar] [CrossRef] [PubMed]
- Carroll, L.M.; Wiedmann, M.; den Bakker, H.; Siler, J.; Warchocki, S.; Kent, D.; Lyalina, S.; Davis, M.; Sischo, W.; Besser, T.; et al. Whole-genome sequencing of drug-resistant Salmonella enterica isolates from dairy cattle and humans in New York and Washington states reveals source and geographic associations. Appl. Environ. Microbiol. 2017, 83, e00140-17. [Google Scholar] [CrossRef] [PubMed]
- Pornsukarom, S.; van Vliet, A.H.M.; Thakur, S. Whole genome sequencing analysis of multiple Salmonella serovars provides insights into phylogenetic relatedness, antimicrobial resistance, and virulence markers across humans, food animals and agriculture environmental sources. BMC Genom. 2018, 19, 801. [Google Scholar] [CrossRef]
- Hald, T.; Lo Fo Wong, D.M.; Aarestrup, F.M. The attribution of human infections with antimicrobial resistant salmonella bacteria in denmark to sources of animal origin. Foodborne Pathog. Dis. 2007, 4, 313–326. [Google Scholar] [CrossRef]
- Armand-Lefevre, L.; Ruimy, R.; Andremont, A. Clonal comparison of staphylococcus aureus isolates from healthy pig farmers, human controls, and pigs. Emerg. Infect. Dis. 2005, 11, 711–714. [Google Scholar] [CrossRef]
- Larsen, J.; Petersen, A.; Larsen, A.R.; Sieber, R.N.; Stegger, M.; Koch, A.; Aarestrup, F.M.; Price, L.B.; Skov, R.L. Emergence of livestock-associated methicillin-resistant Staphylococcus aureus bloodstream infections in Denmark. Clin. Infect. Dis. 2017, 65, 1072–1076. [Google Scholar] [CrossRef]
- Oniciuc, E.-A.; Nicolau, A.I.; Hernández, M.; Rodríguez-Lázaro, D. Presence of methicillin-resistant Staphylococcus aureus in the food chain. Trends Food Sci. Technol. 2017, 61, 49–59. [Google Scholar] [CrossRef]
- Veterinary Medicines Directorate. Uk One Health Report—Joint Report on Antibiotic Use and Antibiotic Resistance, 2013–2017. New Haw, Addlestone. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/775075/One_Health_Report_2019_v45.pdf (accessed on 25 April 2019).
- Van Alen, S.; Ballhausen, B.; Peters, G.; Friedrich, A.W.; Mellmann, A.; Kock, R.; Becker, K. In the centre of an epidemic: Fifteen years of la-mrsa cc398 at the university hospital munster. Vet. Microbiol. 2017, 200, 19–24. [Google Scholar] [CrossRef]
- EFSA/ECDC. The european union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. Efsa J. 2019, 17. [Google Scholar]
- Duggett, N.A.; Sayers, E.; AbuOun, M.; Ellis, R.J.; Nunez-Garcia, J.; Randall, L.; Horton, R.; Rogers, J.; Martelli, F.; Smith, R.P.; et al. Occurrence and characterization of mcr-1-harbouring Escherichia coli isolated from pigs in Great Britain from 2013 to 2015. J. Antimicrob. Chemother. 2017, 72, 691–695. [Google Scholar]
- Venter, H.; Henningsen, M.L.; Begg, S.L. Antimicrobial resistance in healthcare, agriculture and the environment: The biochemistry behind the headlines. Essays Biochem. 2017, 61, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Anjum, M.F.; Duggett, N.A.; AbuOun, M.; Randall, L.; Nunez-Garcia, J.; Ellis, R.J.; Rogers, J.; Horton, R.; Brena, C.; Williamson, S.; et al. Colistin resistance in Salmonella and Escherichia coli isolates from a pig farm in Great Britain. J. Antimicrob. Chemother. 2016, 71, 2306–2313. [Google Scholar] [CrossRef] [PubMed]
- Irrgang, A.; Roschanski, N.; Tenhagen, B.-A.; Grobbel, M.; Skladnikiewicz-Ziemer, T.; Thomas, K.; Roesler, U.; Käsbohrer, A. Prevalence of mcr-1 in e. Coli from livestock and food in Germany, 2010–2015. PLoS ONE 2016, 11, e0159863. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency. Updated Advice on the Use of Colistin Products in Animals within the European Union: Development of Resistance and Possible Impact on Human and Animal Health. 2016. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/updated-advice-use-colistin-products-animals-within-european-union-development-resistance-possible_en-0.pdf (accessed on 25 April 2019).
- Yoon, E.J.; Hong, J.S.; Yang, J.W.; Lee, K.J.; Lee, H.; Jeong, S.H. Detection of mcr-1 plasmids in Enterobacteriaceae isolates from human specimens: Comparison with those in Escherichia coli isolates from livestock in Korea. Ann. Lab. Med. 2018, 38, 555–562. [Google Scholar] [CrossRef]
- Hasman, H.; Hammerum, A.M.; Hansen, F.; Hendriksen, R.S.; Olesen, B.; Agersø, Y.; Zankari, E.; Leekitcharoenphon, P.; Stegger, M.; Kaas, R.S.; et al. Detection of mcr-1 encoding plasmid-mediated colistin-resistant Escherichia coli isolates from human bloodstream infection and imported chicken meat, Denmark 2015. Eur. Surveill. 2015, 20, 30085. [Google Scholar] [CrossRef]
- Verraes, C.; Van Boxstael, S.; Van Meervenne, E.; Van Coillie, E.; Butaye, P.; Catry, B.; de Schaetzen, M.A.; Van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial resistance in the food chain: A review. Int. J. Environ. Res. Public Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef]
- Mateus, A.; Takahashi, E.A.; Abdelrazeq Elkholly, D.; Crotta, M.; Ekiri, A.; Wylie, C.; Guinat, C.; Patricio, M.; Marshall, L.; Pinto Ferreira, J.; et al. A Systematic Review to Assess the Significance of the Food Chain in the Context of Antimicrobial Resistance with Particular Respect to Retail Pork, Poultry Meat, Dairy Products, Seafood and Fresh Produce in the UK; Technical Report; Royal Veterinary College, University of London: London, UK, November 2016. [Google Scholar]
- Jans, C.; Sarno, E.; Collineau, L.; Meile, L.; Stark, K.D.C.; Stephan, R. Consumer exposure to antimicrobial resistant bacteria from food at swiss retail level. Front. Microbiol. 2018, 9, 362. [Google Scholar] [CrossRef]
- Food Standards Agency. Antimicrobial Resistance. Available online: https://www.food.gov.uk/business-guidance/antimicrobial-resistance-amr (accessed on 25 April 2019).
- Nelson, J.M.; Chiller, T.M.; Powers, J.H.; Angulo, F.J. Fluoroquinolone-resistant campylobacter species and the withdrawal of fluoroquinolones from use in poultry: A public health success story. Clin. Infect. Dis. 2007, 44, 977–980. [Google Scholar] [CrossRef]
- Tang, K.L.; Caffrey, N.P.; Nobrega, D.B.; Cork, S.C.; Ronksley, P.E.; Barkema, H.W.; Polachek, A.J.; Ganshorn, H.; Sharma, N.; Kellner, J.D.; et al. Restricting the use of antibiotics in food-producing animals and its associations with antibiotic resistance in food-producing animals and human beings: A systematic review and meta-analysis. Lancet Planet. Health 2017, 1, e316–e327. [Google Scholar] [CrossRef]
- Van Bunnik, B.A.D.; Woolhouse, M.E.J. Modelling the impact of curtailing antibiotic usage in food animals on antibiotic resistance in humans. R. Soc. Open Sci. 2017, 4, 161067. [Google Scholar] [CrossRef]
- SWEDRES-SVARM. Consumption of Antibiotics and Occurence of Antibiotic Resistance in Sweden. Uppsala, Sweden. 2014. Available online: https://www.sva.se/globalassets/redesign2011/pdf/om_sva/publikationer/swedres_svarm2014.pdf (accessed on 15 May 2017).
- Randall, L.P.; Lodge, M.P.; Elviss, N.C.; Lemma, F.L.; Hopkins, K.L.; Teale, C.J.; Woodford, N. Evaluation of meat, fruit and vegetables from retail stores in five United Kingdom regions as sources of extended-spectrum beta-lactamase (esbl)-producing and carbapenem-resistant Escherichia coli. Int. J. Food Microbiol. 2017, 241, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Ludden, C.; Raven, K.E.; Jamrozy, D.; Gouliouris, T.; Blane, B.; Coll, F.; de Goffau, M.; Naydenova, P.; Horner, C.; Hernandez-Garcia, J.; et al. One health genomic surveillance of Escherichia coli demonstrates distinct lineages and mobile genetic elements in isolates from humans versus livestock. mBio 2019, 10, e02693-18. [Google Scholar] [CrossRef] [PubMed]
- Dorado-Garcia, A.; Smid, J.H.; van Pelt, W.; Bonten, M.J.M.; Fluit, A.C.; van den Bunt, G.; Wagenaar, J.A.; Hordijk, J.; Dierikx, C.M.; Veldman, K.T.; et al. Molecular relatedness of esbl/ampc-producing Escherichia coli from humans, animals, food and the environment: A pooled analysis. J. Antimicrob. Chemother. 2018, 73, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Carmo, L.P.; Nielsen, L.R.; da Costa, P.M.; Alban, L. Exposure assessment of extended-spectrum beta-lactamases/ampc beta-lactamases-producing Escherichia coli in meat in Denmark. Infect. Ecol. Epidemiol. 2014, 4, 22924. [Google Scholar] [CrossRef]
- Evers, E.G.; Pielaat, A.; Smid, J.H.; van Duijkeren, E.; Vennemann, F.B.C.; Wijnands, L.M.; Chardon, J.E. Comparative exposure assessment of esbl-producing Escherichia coli through meat consumption. PLoS ONE 2017, 12, e0169589. [Google Scholar] [CrossRef]
- Gouliouris, T.; Raven, K.E.; Ludden, C.; Blane, B.; Corander, J.; Horner, C.S.; Hernandez-Garcia, J.; Wood, P.; Hadjirin, N.F.; Radakovic, M.; et al. Genomic surveillance of Enterococcus faecium reveals limited sharing of strains and resistance genes between livestock and humans in the United Kingdom. mBio 2018, 9, e01780-18. [Google Scholar] [CrossRef]
- Gantzhorn, M.R.; Pedersen, K.; Olsen, J.E.; Thomsen, L.E. Biocide and antibiotic susceptibility of Salmonella isolates obtained before and after cleaning at six danish pig slaughterhouses. Int. J. Food Microbiol. 2014, 181, 53–59. [Google Scholar] [CrossRef]
- Alonso-Hernando, A.; Capita, R.; Prieto, M.; Alonso-Calleja, C. Comparison of antibiotic resistance patterns in Listeria monocytogenes and Salmonella enterica strains pre-exposed and exposed to poultry decontaminants. Food Control 2009, 20, 1108–1111. [Google Scholar] [CrossRef]
- Condell, O.; Iversen, C.; Cooney, S.; Power, K.A.; Walsh, C.; Burgess, C.; Fanning, S. Efficacy of biocides used in the modern food industry to control Salmonella enterica, and links between biocide tolerance and resistance to clinically relevant antimicrobial compounds. Appl. Environ. Microbiol. 2012, 78, 3087–3097. [Google Scholar] [CrossRef]
- Poole, K. Bacterial stress responses as determinants of antimicrobial resistance. J. Antimicrob. Chemother. 2012, 67, 2069–2089. [Google Scholar] [CrossRef]
- McDonnell, L.; Armstrong, D.; Ashworth, M.; Dregan, A.; Malik, U.; White, P. National disparities in the relationship between antimicrobial resistance and antimicrobial consumption in europe: An observational study in 29 countries. J. Antimicrob. Chemother. 2017, 72, 3199–3204. [Google Scholar] [CrossRef] [PubMed]
- Goossens, H.; Ferech, M.; Vander Stichele, R.; Elseviers, M. Outpatient antibiotic use in Europe and association with resistance: A cross-national database study. Lancet 2005, 365, 579–587. [Google Scholar] [CrossRef]
- Zhang, X.F.; Doi, Y.; Huang, X.; Li, H.Y.; Zhong, L.L.; Zeng, K.J.; Zhang, Y.F.; Patil, S.; Tian, G.B. Possible transmission of mcr-1-harboring Escherichia coli between companion animals and human. Emerg. Infect. Dis. 2016, 22, 1679–1681. [Google Scholar] [CrossRef] [PubMed]
- EMA/EFSA. Joint scientific opinion on measures to reduce the need to use antimicrobial agents in animal husbandry in the European Union, and the resulting impacts on food safety (ronafa). Efsa J. 2017, 15. [Google Scholar]
- Anjum, M.F.; Zankari, E.; Hasman, H. Molecular methods for detection of antimicrobial resistance. Microbiol. Spectr. 2017, 5, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pires, S.M.; Duarte, A.S.; Hald, T. Source attribution and risk assessment of antimicrobial resistance. Microbiol. Spectr. 2018, 6, 1–7. [Google Scholar]


{kind=link}
| Antimicrobials | Hosts | Associated Bacteria | Reference |
|---|---|---|---|
| Cefotaxime (a 3rd generation cephalosporin) | Broilers | E. coli | [16] |
| Cephalosporin | Pigs | E. coli | [21] |
| Avoparcin (a glycopeptide) | Broilers | E. faecium | [24,25] |
| Tylosin (a macrolide) | Pigs | E. faecium and E. faecalis | [25] |
| Tylosin (a macrolide) | Broilers | E. faecium | [25] |
| Virginiamycin (a streptogramin) | Broilers | E. faecium | [25] |
| Avilamycin (an oligosaccharide) | Broilers | E. faecium | [25] |
| Ampicillin | Livestock | E. coli | [22] |
| Fluoroquinolones and other quinolones | Food producing animals | E. coli, Salmonella spp., Campylobacter jejuni and Campylobacter coli | [13,14] |
| Tetracyclines | Food producing animals | E. coli, Salmonella spp. and Campylobacter jejuni | [13,14] |
| Macrolides | Food producing animals | Campylobacter coli | [13,14] |
| Ceftiofur (a 3rd generation cephalosporin) | Chickens | E. coli and Salmonella enterica serovar Heidelberg | [27] |
| Bacterial Resistance Genes | Role in Antimicrobial Resistance |
|---|---|
| Genes encoding for extended-spectrum beta-lactamase (ESBL-) and AmpC beta-lactamase (AmpC-) in Enterobacteriaceae (for ex. CTX-M type) | Exhibit resistance to a wide range of β-lactam antibiotics, including penicillins and 3rd and 4th generation cephalosporins in Gram negative bacteria. They are one of the fastest emerging resistance problems in both humans, companion and production animals worldwide. Third and 4th generation cephalosporins are now classified as Highest Priority Critically Important Antimicrobials (HPCIAs) by WHO. |
| Carbapenemase-encoding genes in Enterobacteriaceae | Carbapenems are classified by WHO as HPCIAs. Resistance to carbapenems is emerging in humans and constitute a major public health concern. Although carbapenems are not licensed for use in animals, carbapenem resistance has been isolated in bacteria from companion, food-producing animals and wildlife. |
| Livestock-associated methicillin-resistant Staphylococcus aureus (LA-MRSA) Clonal Complex 3398 | Found in the skin and nose of pigs, cattle and horses without causing a disease and they can transmit it to humans. Exposed individuals can become colonised asymptomatically; LA-MRSA can cause skin infections and occasionally invasive disease (septicaemia). Veterinarians and farmers are at high risk of exposure through their occupations. |
| Plasmid mediated colistin resistance (mcr genes) | Colistin (Polymyxin E) is a last resort antibiotic in humans. This new gene is transmitted horizontally, which can cause a higher risk of spread between animals and humans. Colistin resistance has been identify in livestock (pigs and poultry) and animal-derived food and in humans. |
| Vancomycin resistance encoding genes in Enterococcus faecium | A major cause of nosocomial infection and is categorized as high priority by the WHO global priority list of antibiotic-resistant bacteria. Vancomycin-resistant Enterococci (VRE) have been isolated from food-producing animals as a result of the previous use of avoparcin as a growth promoter (Note: antibiotic growth promoters have been banned in Europe since January 2006 but are still used in other countries). |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bennani, H.; Mateus, A.; Mays, N.; Eastmure, E.; Stärk, K.D.C.; Häsler, B. Overview of Evidence of Antimicrobial Use and Antimicrobial Resistance in the Food Chain. Antibiotics 2020, 9, 49. https://doi.org/10.3390/antibiotics9020049
Bennani H, Mateus A, Mays N, Eastmure E, Stärk KDC, Häsler B. Overview of Evidence of Antimicrobial Use and Antimicrobial Resistance in the Food Chain. Antibiotics. 2020; 9(2):49. https://doi.org/10.3390/antibiotics9020049
Chicago/Turabian StyleBennani, Houda, Ana Mateus, Nicholas Mays, Elizabeth Eastmure, Katharina D. C. Stärk, and Barbara Häsler. 2020. "Overview of Evidence of Antimicrobial Use and Antimicrobial Resistance in the Food Chain" Antibiotics 9, no. 2: 49. https://doi.org/10.3390/antibiotics9020049
APA StyleBennani, H., Mateus, A., Mays, N., Eastmure, E., Stärk, K. D. C., & Häsler, B. (2020). Overview of Evidence of Antimicrobial Use and Antimicrobial Resistance in the Food Chain. Antibiotics, 9(2), 49. https://doi.org/10.3390/antibiotics9020049