Antitumor Potential of Seaweed Derived-Endophytic Fungi

 , , ,
, , ,
Abstract
1. Introduction
2. Endophytic Fungi
2.1. Ecological Role of Endophytic Fungi
2.2. Isolation of Endophytic Fungi
3. Cytotoxic Secondary Metabolites Produced by Endophytic Fungi
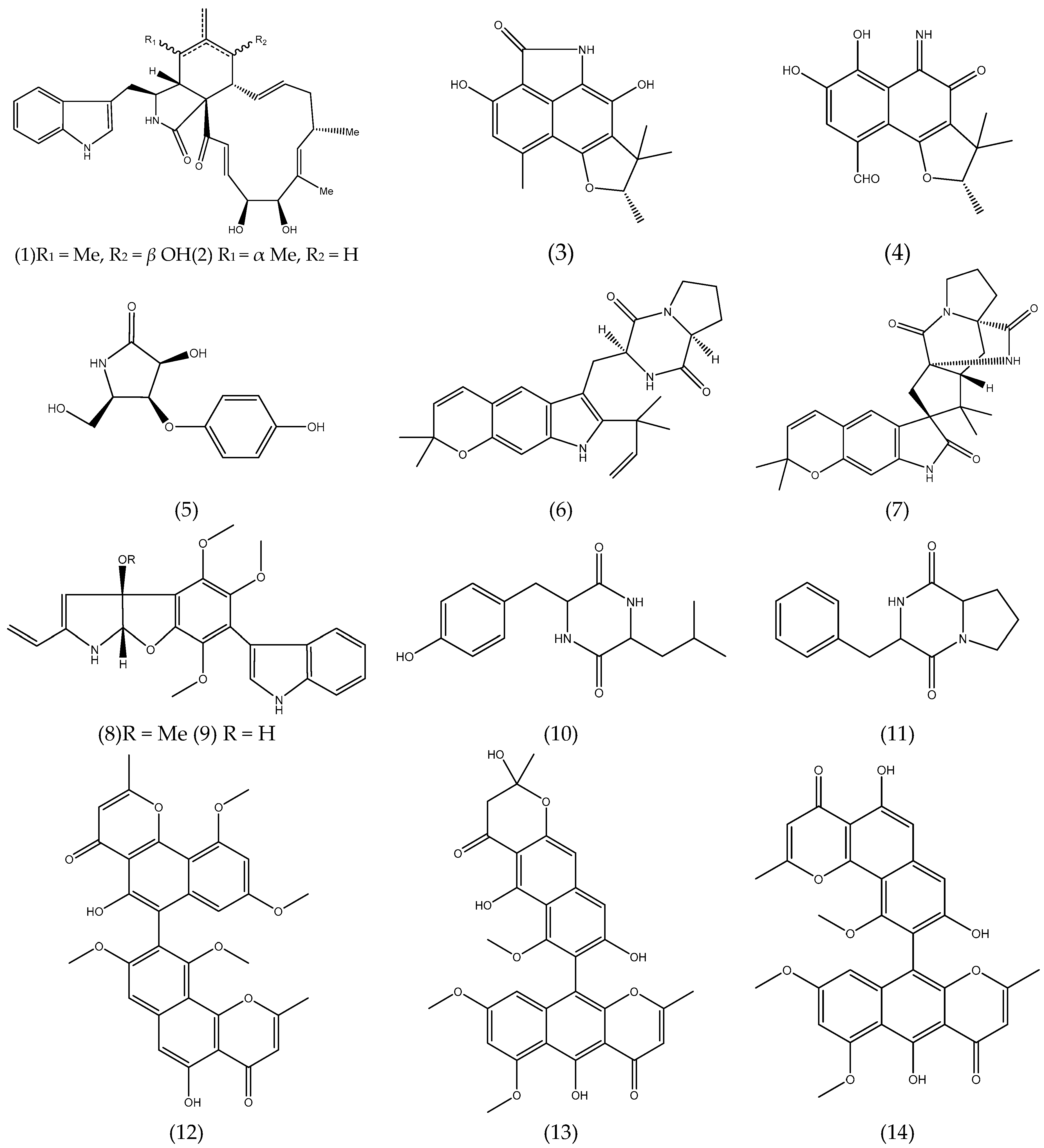
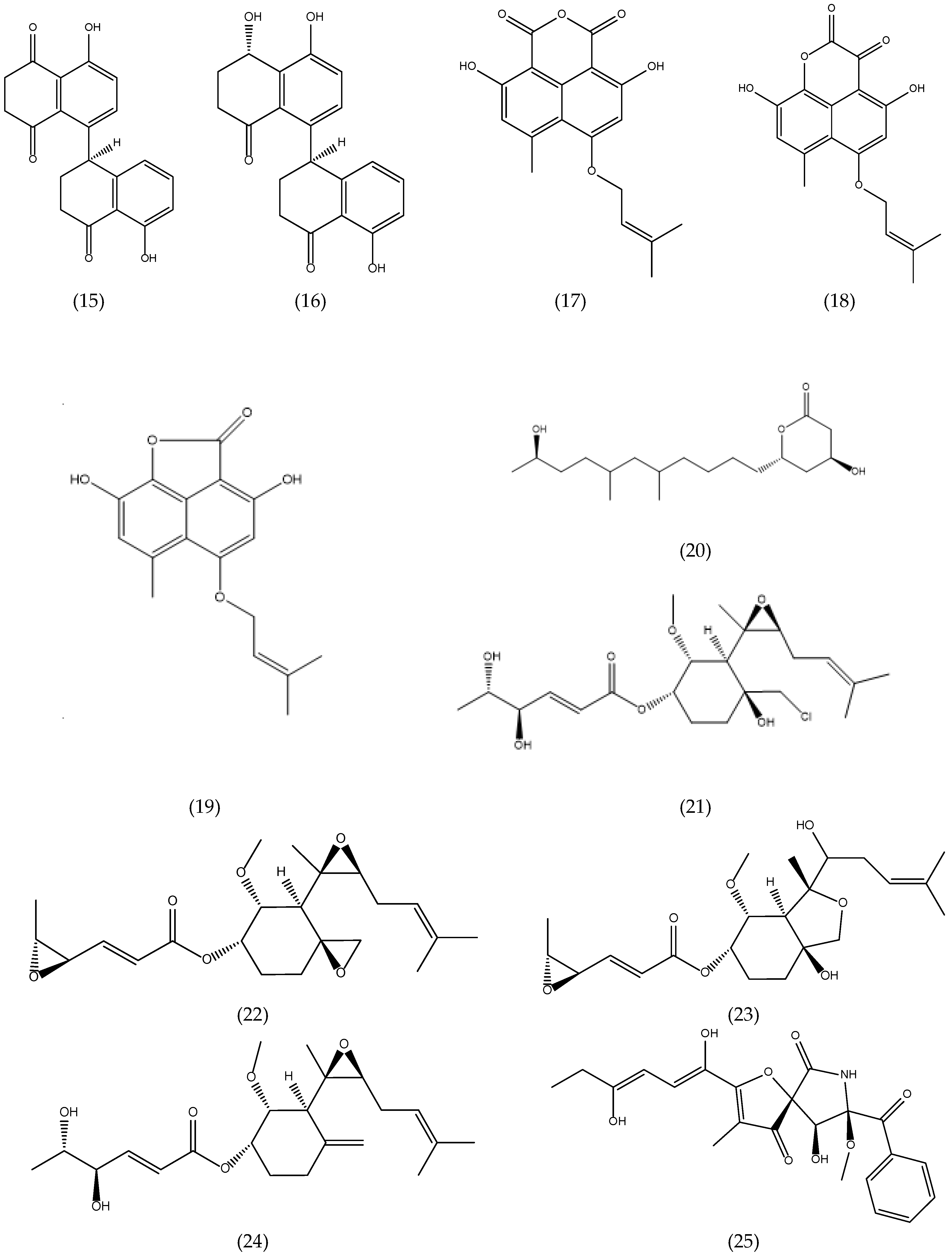
3.1. Alkaloids and Nitrogen-containing Heterocycles
3.2. Polyketides
3.3. Quinones
3.4. Terpenoids and Sterols
3.4.1. Terpenoids
3.4.2. Sterols
3.5. Others
4. Biotechnology of Marine Endophytic Fungi
Biotechnological Processes to Obtain Bioactive Metabolites of Endophytic Fungi
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nivetha, X.R.; Soundari, M.J.J.; Nadar, M.S.A.M.; Premnath, D.; Selvakumar, P.M.; Chang, R. An insight into cancer and anticancer drugs. Acta Sci. Med. Sci. 2019, 3, 32–43. [Google Scholar]
- Worldwide Cancer Data: Global Cancer Statistics for the Most Common Cancers. Available online: https://www.wcrf.org/dietandcancer/cancer-trends/worldwide-cancer-data (accessed on 31 July 2019).
- Rocha, D.H.A.; Seca, A.M.; Pinto, D.C.G.A. Seaweed secondary metabolites in vitro and in vivo anticancer activity. Mar. Drugs 2018, 16, 410. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer. World Health Organization. Available online: http://gco.iarc.fr/tomorrow/graphic-isotype?type=0&population=900&mode=population&sex=0&cancer=39&age_group=value&apc_male=0&apc_female=0 (accessed on 31 July 2019).
- Gerwick, W.H.; Moore, B.S. Lessons from the past and charting the future of Marine Natural Products drug discovery and chemical biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Newman, D.J. Natural products: Current Approaches and perspectives. In Natural Products as Source of Molecules with Therapeutic Potential; Cechinel, F.V., Ed.; Springer: Cham, Switzerland, 2018; pp. 309–331. [Google Scholar] [CrossRef]
- Jimenez, P.C.; Wilke, D.V.; Costa-Lotufo, L.V. Marine drugs for cancer: Surfacing biotechnological innovations from the oceans. Clinics 2018, 73, e482s. [Google Scholar] [CrossRef]
- Rayan, A.; Raiyn, J.; Falah, M. Nature is the best source of anticancer drugs: Indexing natural products for their anticancer bioactivity. PLoS ONE 2017, 12, e0187925. [Google Scholar] [CrossRef]
- Dyshlovoy, S.A.; Honecker, F. Marine compounds and cancer: 2017 updates. Mar. Drugs 2018, 16, 41. [Google Scholar] [CrossRef]
- Pavão, G.B.; Venâncio, V.P.; Oliveira, A.L.L.; Hernandes, L.C.; Almeida, M.R.; Antunes, L.M.G.; Debonsi, H.M. Differential genotoxicity and cytotoxicity of phomoxanthone A isolated from the fungus Phomopsis longicolla in HL60 cells and peripheral blood lymphocytes. Toxicol. In Vitro 2016, 37, 211–217. [Google Scholar] [CrossRef]
- Maciel, O.M.C.; Tavares, R.S.N.; Caluz, D.R.E.; Gaspar, L.R.; Debonsi, H.M. Photoprotective potential of metabolites isolated from algae-associated fungi Annulohypoxylon stygium. J. Photochem. Photobiol. B 2018, 178, 316–322. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Prakash, V.; Ranjan, N. Marine fungi: A source of potential anticancer compounds. Front. Microbiol. 2017, 8, 2536. [Google Scholar] [CrossRef]
- Jeewon, R.; Luckhun, A.B.; Bhoyroo, V.; Sadeer, N.B.; Mahomoodally, F.M.; Rampadarath, S.; Puchooa, D.; Sarma, V.; Kumar Durairajan, S.S.; Hyde, K.D. Pharmaceutical Potential of Marine Fungal Endophytes. In Endophytes and Secondary Metabolites; Jha, S., Ed.; Reference Series in Phytochemistry; Springer: Cham, Switzerland, 2019; pp. 1–23. [Google Scholar] [CrossRef]
- Li, T.; Wang, N.; Zhang, T.; Zhang, B.; Sajeevan, T.P.; Joseph, V.; Armstrong, L.; He, S.; Yan, X.; Naman, B. A Systematic Review of Recently Reported Marine Derived Natural Product Kinase Inhibitors. Mar. Drugs 2019, 17, 493. [Google Scholar] [CrossRef]
- Yanagihara, M.; Sasaki-Takahashi, N.; Sugahara, T.; Yamamoto, S.; Shinomi, M.; Yamashita, I.; Hayashida, M.; Yamanoha, B.; Numata, A.; Yamori, T.; et al. Leptosins isolated from marine fungus Leptoshaeria species inhibit DNA topoisomerases I and/or II and induce apoptosis by inactivation of Akt/protein kinase B. Cancer Sci. 2005, 96, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.H.; Debbab, A.; Kjer, J.; Proksch, P. Fungal endophytes from higher plants: A prolific source of phytochemicals and other bioactive natural products. Fungal Divers. 2010, 41, 1–16. [Google Scholar] [CrossRef]
- Petrini, O. Fungal Endophytes of Tree Leaves. In Microbial Ecology of Leaves; Andrews, J.H., Hirano, S.S., Eds.; Brock/Springer: New York, NY, USA, 1991; pp. 179–197. ISBN 978-1-4612-3168-4. [Google Scholar]
- Wilson, D. Endophyte: The Evolution of a Term, and Clarification of Its Use and Definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Singh, R.P.; Kumaru, P.; Reddy, C.R.K. Antimicrobial compounds from seaweeds-associated bacteria and fungi. Appl. Microbiol. Biotechnol. 2015, 99, 1571–1586. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef]
- Mani, V.M.; Soundari, A.P.G.; Karthiyaini, D.; Preeth, K. Bioprospecting endophytic fungi and their metabolites from medicinal tree Aegle marmelosin Western Ghats, India. Mycobiology 2015, 43, 303–310. [Google Scholar] [CrossRef]
- Hyde, K.D.; Soytong, K. The fungal endophyte dilemma. Fungal Divers. 2008, 33, 163–173. [Google Scholar]
- Rodriguez, R.J.; White, J.F., Jr.; Arnold, A.E.; Redman, A.R.A. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Ji, N.Y.; Wang, B.G. Mycochemistry of marine algicolous fungi. Fungal Divers. 2016, 80, 301–342. [Google Scholar] [CrossRef]
- Yan, J.F.; Broughton, S.J.; Yang, S.L.; Gange, A.C. Do endophytic fungi grow through their hosts systemically? Fungal Ecol. 2015, 13, 53–59. [Google Scholar] [CrossRef]
- Tan, R.X.; Zou, W.X. Endophytes: A rich source of functional metabolites. Nat. Prod. Rep. 2001, 18, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Debbab, A.; Aly, A.H.; Proksch, P. Endophytes and associated marine derived fungi—Ecological and chemical perspectives. Fungal Divers. 2012, 57, 45–83. [Google Scholar] [CrossRef]
- Brundrett, M.C. Understanding the roles of multifunctional mycorrhizal and endophytic fungi. In Microbial Root Endophytes; Schulz, B.J.E., Boyle, C.J.C., Sieber, T.N., Eds.; Springer: Berlin, Germany, 2006; pp. 281–293. [Google Scholar] [CrossRef]
- Felício, R.; Pavão, G.B.; Oliveira, A.L.; Erbert, C.; Conti, R.; Pupo, M.T.; Furtado, N.A.J.C.; Ferreira, E.G.; Costa-Lotufo, L.V.; Young, M.C.M.; et al. Antibacterial, antifungal and cytotoxic activities exhibited by endophytic fungi from the Brazilian marine red alga Bostrychia tenella (Ceramiales). Braz. J. Pharmacog. 2015, 25, 641–650. [Google Scholar] [CrossRef]
- Zhou, K.; Zhang, X.; Zhang, F.; Li, Z. Phylogenetically diverse cultivable fungal community and polyketide synthase (PKS), nonribosomal peptide synthase (NRPS) genes associated with the South China Sea sponges. Microb. Ecol. 2011, 62, 644–654. [Google Scholar] [CrossRef]
- Zuccaro, A.; Mitchell, J.I. Fungal communities of seaweeds. In The Fungal Community, 3rd ed.; Dighton, J., White, J.F., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 533–579. [Google Scholar] [CrossRef]
- Vallet, M.; Strittmatter, M.; Murúa, P.; Lacoste, S.; Dupont, J.; Hubas, C.; Prado, S. Chemically-mediated interactions between macroalgae, their fungal endophytes, and protistan pathogens. Front. Microbiol. 2018, 9, 3161. [Google Scholar] [CrossRef] [PubMed]
- Godinho, V.M.; Furbino, L.E.; Santiago, I.F.; Pellizzari, F.M.; Yokoya, N.; Pupo, D.; Cantrell, C.L. Diversity and bioprospecting of fungal communities associated with endemic and cold-adapted macroalgae in Antarctica. ISME J. 2013, 7, 14–34. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.D.; Hyde, K.D.; Liew, E.C. Detection and taxonomic placement of endophytic fungi within frond tissues of Livistona chinensis based on rDNA sequences. Mol. Phylogenet. Evol. 2001, 20, 1–13. [Google Scholar] [CrossRef]
- Schulz, B.; Guske, S.; Dammann, U.; Boyle, C. Endophyte-host interactions. II. Defining symbiosis of the endophyte-host interaction. Symbiosis 1998, 25, 213–227. [Google Scholar]
- Erbert, C.; Lopes, A.A.; Yokoya, N.S.; Furtado, N.A.J.C.; Conti, R.; Pupo, M.T.; Lopes, J.L.C.; Debonsi, H.M. Antibacterial compound from the endophytic fungus Phomopsis longicolla isolated from the tropical red seaweed Bostrychia radicans. Bot. Mar. 2012, 55, 435–440. [Google Scholar] [CrossRef]
- Kjer, J.; Debbab, A.; Aly, A.H.; Proksch, P. Methods for isolation of marine-derived endophytic fungi and their bioactive secondary products. Nat. Protoc. 2010, 5, 479–490. [Google Scholar] [CrossRef]
- Teixeira, T.R.; Santos, G.S.; Turatti, I.C.C.; Paziani, M.H.; Kress, M.R.Z.; Colepicolo, P.; Debonsi, H.M. Characterization of the lipid profile of Antarctic brown seaweeds and their endophytic fungi by gas chromatography–mass spectrometry (GC–MS). Polar Biol. 2019, 42, 1431–1444. [Google Scholar] [CrossRef]
- Cui, C.M.; Li, X.M.; Meng, L.; Li, C.S.; Huang, C.G.; Wang, B.G. 7-Nor-ergosterolide, a Pentalactone Containing Norsteroid and Related Steroids from the Marine-Derived Endophytic Aspergillus ochraceus EN-31. J. Nat. Prod. 2010, 73, 1780–1784. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Lin, X.; Zhou, X.; Wan, J.; Lu, X.; Yang, B.; Ai, W.; Lin, J.; Zhang, T.; Tu, Z.; et al. Cytotoxic and antiviral nitrobenzoyl sesquiterpenoids from the marine-derived fungus Aspergillus ochraceus Jcma1F17. Med. Chem. Commun. 2014, 5, 701–705. [Google Scholar] [CrossRef]
- Tan, Y.; Yang, B.; Lin, X.; Luo, X.; Pang, X.; Tang, L.; Liu, Y.; Li, X.; Zhou, X. Nitrobenzoyl Sesquiterpenoids with Cytotoxic Activities from a Marine-Derived Aspergillus ochraceus Fungus. J. Nat. Prod. 2018, 81, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.X.; Wang, X.F.; Ren, G.W.; Yuan, X.L.; Deng, N.; Ji, G.X.; Li, W.; Zhang, P. Prenylated Diphenyl Ethers from the Marine Algal-Derived Endophytic Fungus Aspergillus tennesseensis. Molecules 2018, 23, 2368. [Google Scholar] [CrossRef]
- Hawas, U.W.; El-Beih, A.A.; El-Halawany, A.M. Bioactive Anthraquinones from Endophytic Fungus Aspergillus versicolor Isolated from Red Sea Algae. Arch. Pharm. Res. 2012, 35, 1749–1756. [Google Scholar] [CrossRef]
- Sun, H.F.; Li, X.M.; Meng, L.; Cui, C.M.; Gao, S.S.; Li, C.S.; Huang, C.G.; Wang, B.G. Asperolides A−C, Tetranorlabdane Diterpenoids from the Marine Alga-Derived Endophytic Fungus Aspergillus wentii EN-48. J. Nat. Prod. 2012, 75, 148–152. [Google Scholar] [CrossRef]
- Zhang, Z.; Miao, L.; Sun, H.; Wei, S.; Wang, B.; Huang, C.; Jiao, B. Wentilactone B induces G2/M phase arrest and apoptosis via the Ras/Raf/MAPK signaling pathway inhuman hepatoma SMMC-7721 cells. Cell Death Dis. 2013, 4, e675. [Google Scholar] [CrossRef]
- Zhang, Z.; Miao, L.; Sun, W.; Jiao, B.; Wang, B.; Yao, L.; Huang, C. Wentilactone B from Aspergillus wentii Induces Apoptosis and Inhibits Proliferation and Migration of Human Hepatoma SMMC-7721 Cells. Biol. Pharm. Bull. 2012, 35, 1964–1971. [Google Scholar] [CrossRef]
- Lv, C.; Hong, Y.; Miao, L.; Li, C.; Xu, G.; Wei, S.; Wang, B.; Huang, C.; Jiao, B. Wentilactone A as a novel potential antitumor agent induces apoptosis and G2/M arrest of human lung carcinoma cells, and is mediated by HRas-GTP accumulation to excessively activate the Ras/Raf/ERK/p53-p21 pathway. Cell Death Dis. 2013, 4, e952. [Google Scholar] [CrossRef]
- Lv, C.; Sun, W.; Sun, H.; Wei, S.; Chen, R.; Wang, B.; Huang, C. Asperolide A, a Marine-Derived Tetranorditerpenoid, Induces G2/M Arrest in Human NCI-H460 Lung Carcinoma Cells, Is Mediated by p53-p21 Stabilization and Modulated by Ras/Raf/MEK/ERK Signaling Pathway. Mar. Drugs 2013, 11, 316–331. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Guo, C.; Meng, J.; Tian, H.; Guo, S.; Shi, D. Discovery of Natural Dimeric Naphthopyrones as Potential Cytotoxic Agents through ROS-Mediated Apoptotic Pathway. Mar. Drugs 2019, 17, 207. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.M.; Li, X.M.; Li, C.S.; Proksch, P.; Wang, B.G. Cytoglobosins A-G, Cytochalasans from a Marine-Derived Endophytic Fungus, Chaetomium globosum QEN-14. J. Nat. Prod. 2010, 73, 729–733. [Google Scholar] [CrossRef]
- Li, H.L.; Li, X.M.; Mándi, A.; Antus, S.; Li, X.; Zhang, P.; Liu, Y.; Kurtán, T.; Wang, B.G. Characterization of Cladosporols from the Marine Algal-Derived Endophytic Fungus Cladosporium cladosporioides EN-399 and Configurational Revision of the Previously Reported Cladosporol Derivatives. J. Org. Chem. 2017, 82, 9946–9954. [Google Scholar] [CrossRef] [PubMed]
- Elsebai, M.F.; Kehraus, S.; Lindequist, U.; Sasse, F.; Shaaban, S.; Gutschow, M.; Josten, M.; Sahl, H.G.; Konig, G.M. Antimicrobial phenalenone derivatives from the marine-derived fungus Coniothyrium cereale. Org. Biomol. Chem. 2011, 9, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.H.; Tang, X.Z.; Miao, F.P.; Ji, N.Y. A New Pyrrolidine Derivative and Steroids from an Algicolous Gibberella zeae Strain. Nat. Prod. Commun. 2011, 6, 1243–1246. [Google Scholar] [CrossRef]
- Wang, F.W. Bioactive metabolites from Guignardia sp., an endophytic fungus residing in Undaria pinnatifida . Chin. J. Nat. Med. 2012, 10, 72–76. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.M.; Wang, J.N.; Li, X.; Wang, B.-G. Prenylated indole alkaloids from the marine-derived fungus Paecilomyces variotii. Chin. Chem. Lett. 2015, 26, 313–316. [Google Scholar] [CrossRef]
- Zhang, P.; Li, X.M.; Mao, X.X.; Mándi, A.; Kurtàn, T.; Wang, B.G. Varioloid A, a new indolyl-6,10b-dihydro-5aH-[1]benzofuro[2 –b] indole derivative from the marine alga-derived endophytic fungus Paecilomyces variotii EN-291. Beilstein J. Org. Chem. 2016, 12, 2012–2018. [Google Scholar] [CrossRef]
- Gao, S.S.; Li, X.M.; Du, F.Y.; Li, C.S.; Proksch, P.; Wang, B.G. Secondary Metabolites from a Marine-Derived Endophytic Fungus Penicillium chrysogenum QEN-24S. Mar. Drugs. 2011, 9, 59–70. [Google Scholar] [CrossRef]
- Gao, S.S.; Li, X.M.; Li, C.S.; Proksch, P.; Wang, B.G. Penicisteroids A and B, antifungal and cytotoxic polyoxygenated steroids from the marine alga-derived endophytic fungus Penicillium chrysogenum QEN-24S. Bioorg. Med. Chem. Lett. 2011, 21, 2894–2897. [Google Scholar] [CrossRef] [PubMed]
- Gamal-Eldeen, A.M.; Abdel-Lateff, A.; Okino, T. Modulation of carcinogen metabolizing enzymes bychromanone A; a newchromone derivative from algicolous marine fungus Penicillium sp. Environ. Toxicol. Pharmacol. 2009, 28, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Wang, S.W.; Wang, G.J.; Pang, K.L.; Lee, C.K.; Kuo, Y.H.; Cha, H.J.; Lin, R.K.; Lee, T.H. Angiogenesis Inhibitors and Anti-Inflammatory Agents from Phoma sp. NTOU4195. J. Nat. Prod. 2016, 79, 2983–2990. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; Li, X.M.; Li, X.; Wang, C.Y.; Liu, H.; Kassack, M.U.; Meng, L.H.; Wang, B.G. Antioxidant Hydroanthraquinones from the Marine Algal-Derived Endophytic Fungus Talaromyces islandicus EN-501. J. Nat. Prod. 2017, 80, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Gunatilaka, A.A.L. Natural products from plant-associated microorganisms: Distribution, structural diversity, bioactivity, and implications of their occurrence. J. Nat. Prod. 2006, 69, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.S. New aspects of natural products in drug discovery. Trends Microbiol. 2007, 15, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.T.; Fischbach, M.A. Natural products version: 2.0: Connecting genes to molecules. J. Am. Chem. Soc. 2010, 132, 2469–2493. [Google Scholar] [CrossRef]
- Gomes, N.G.M.; Lefranc, F.; Kijjoa, A.; Kiss, R. Can Some Marine-Derived Fungal Metabolites Become Actual Anticancer Agents? Mar. Drugs 2015, 13, 3950–3991. [Google Scholar] [CrossRef]
- Evidente, A.; Kornienko, A.; Cimmino, A.; Andolfi, A.; Lefranc, F.; Mathieu, V.; Kiss, R. Fungal metabolites with anticancer activity. Nat. Prod. Rep. 2014, 31, 617–627. [Google Scholar] [CrossRef]
- Elsebai, M.F.; Natesan, L.; Kehraus, S.; Mohamed, I.E.; Schnakenburg, G.; Sasse, F.; Shaaban, S.; Gutschow, M.; Koning, G.M. HLE-Inhibitory Alkaloids with a Polyketide Skeleton from the Marine-Derived Fungus Coniothyrium cereale. J. Nat. Prod. 2011, 74, 2282–2285. [Google Scholar] [CrossRef]
- Zurlo, D.; Leone, C.; Assante, G.; Salzano, S.; Renzone, G.; Scaloni, A.; Foresta, C.; Colantuoni, V.; Lupo, A. Cladosporol a stimulates G1-phase arrest of the cell cycle by up-regulation of p21(waf1/cip1) expression in human colon carcinoma HT-29 cells. Mol. Carcinog. 2013, 52, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zurlo, D.; Assante, G.; Moricca, S.; Colantuoni, V.; Lupo, A. Cladosporol A, a new peroxisome proliferator-activated receptor γ (PPARγ) ligand, inhibits colorectal cancer cells proliferation through β-catenin/TCF pathway inactivation. Biophy. Acta Gen. Subj. 2014, 1840, 2361–2372. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Qiu, X.; Wang, R.; Duan, L.; Chen, S.; Luo, J.; Kong, L. Inhibition of Human Gastric Carcinoma Cell Growth in Vitro and in Vivo by Cladosporol Isolated from the Paclitaxel-Producing Strain Alternaria alternata var. monosporus. Biol. Pharm. Bull. 2009, 32, 2072–2074. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.X.; Jin, X.J.; Draskovic, M.; Crews, M.S.; Tenney, K.; Valeriote, F.A.; Yao, X.J.; Crews, P. Unraveling the Numerous Biosynthetic Products of the Marine Sediment-Derived Fungus, Aspergillus insulicola. Phytochem. Lett. 2012, 5, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Belofsky, G.N.; Jensen, P.R.; Renner, M.K.; Fenical, W. New cytotoxic sesquiterpenoid nitrobenzoyl esters from a marine isolate of the fungus Aspergillus versicolor. Tetrahedron Lett. 1998, 54, 1715–1724. [Google Scholar] [CrossRef]
- Zhao, H.Y.; Anbuchezhian, R.; Sun, W.; Shao, C.L.; Zhang, F.L.; Yin, Y.; Yu, Z.S.; Li, Z.Y.; Wang, C.Y. Cytotoxic Nitrobenzoyloxy-substituted Sesquiterpenes from Sponge derived Endozoic Fungus Aspergillus insulicola MD10-2. Curr. Pharm. Biotechnol. 2016, 17, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Silber, J.; Kramer, A.; Labes, A.; Tasdemir, D. From Discovery to Production: Biotechnology of Marine Fungi for the Production of New Antibiotics. Mar. Drugs 2016, 14, 137–157. [Google Scholar] [CrossRef]
- Strobel, G.A. Rainforest endophytes and bioactive products. Crit. Rev. Biotechnol. 2002, 22, 315–333. [Google Scholar] [CrossRef]
- Raghukumar, C. Marine fungal biotechnology: An ecological perspective. Fungal Divers. 2008, 31, 19–35. [Google Scholar]
- Chambergo, F.S.; Valencia, S.Y. Fungal biodiversity to biotechnology. Appl. Microbiol. Biotechnol. 2016, 100, 2567–2577. [Google Scholar] [CrossRef]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites-Strategies to activate silent gene clusters. Fungal Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B.; Cheung, R.C.; Wong, J.H.; Bekhit, A.A.; BekhitAel, D. Antibacterial products of marine organisms. Appl. Microbiol. Biotechnol. 2015, 99, 4145–4173. [Google Scholar] [CrossRef] [PubMed]
- Greco, K.; Keller, N.P.; Rokas, A. Unearthing fungal chemodiversity and prospects for drug discovery. Curr. Opin. Microbiol. 2019, 51, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F.; Aires-de-Sousa, J. Computational Methodologies in the Exploration of Marine Natural Product Leads. Mar. Drugs 2018, 16, 236–266. [Google Scholar] [CrossRef] [PubMed]
- Kildgaard, S.; Mansson, M.; Dosen, I.; Klitgaard, A.; Frisvad, J.C.; Larsen, T.O.; Nielsen, K.F. Accurate Dereplication of Bioactive Secondary Metabolites from Marine-Derived Fungi by UHPLC-DAD QTOFMS and a MS/HRMS Library. Mar. Drugs 2014, 12, 3681–3705. [Google Scholar] [CrossRef] [PubMed]
- El-Elimat, T.; Figueroa, M.; Ehrmann, B.M.; Cech, N.B.; Pearce, C.J.; Oberlies, N.H. High-Resolution MS, MS/MS, and UV Database of Fungal Secondary Metabolites as a Dereplication Protocol for Bioactive Natural Products. J. Nat. Prod. 2013, 76, 1709–1716. [Google Scholar] [CrossRef]
- Naman, C.B.; Rattan, R.; Nikoulina, S.E.; Lee, J.; Miller, B.W.; Moss, N.A.; Armstrong, L.; Boudreau, P.D.; Debonsi, H.M.; Valeriote, F.A.; et al. Integrating Molecular Networking and Biological Assays To Target the Isolation of a Cytotoxic Cyclic Octapeptide, Samoamide A, from an American Samoan Marine Cyanobacterium. J. Nat. Prod. 2017, 80, 625–633. [Google Scholar] [CrossRef]
- Naman, C.B.; Almaliti, J.; Armstrong, L.; Caro-Díaz, E.J.; Pierce, M.L.; Glukhov, E.; Fenner, A.; Spadafora, C.; Debonsi, H.M.; Dorrestein, P.C.; et al. Discovery and Synthesis of Caracolamide A, an Ion Channel Modulating Dichlorovinylidene Containing Phenethylamide from a Panamanian Marine Cyanobacterium cf. Symploca Species. J. Nat. Prod. 2017, 80, 2328–2334. [Google Scholar] [CrossRef]
- Philippus, A.C.; Zatelli, G.A.; Wanke, T.; Barros, M.G.A.; Kami, S.A.; Lhullier, C.; Armstrong, L.; Sandjo, L.P.; Falkenberg, M. Molecular networking prospection and characterization of terpenoids and C 15-acetogenins in Brazilian seaweed extracts. RSC Adv. 2018, 8, 29654–29661. [Google Scholar] [CrossRef]
- Linde, T.; Hansen, N.B.; Lübeck, M.; Lübeck, P.S. Fermentation in 24-well plates is an efficient screening platform for filamentous fungi. Lett. Appl. Microbiol. 2014, 59, 224–230. [Google Scholar] [CrossRef]
- Girarda, P.; Jordana, M.; Tsaob, M.; Wurma, F.M. Small-scale bioreactor system for process development and optimization. Biochem. Eng. J. 2001, 7, 117–119. [Google Scholar] [CrossRef]
- Kramer, A.; Paun, L.; Imhoff, J.F.; Kempken, F.; Labes, A. Development and validation of a fast and optimized screening method for enhanced production of secondary metabolites using the marine Scopulariopsis brevicaulis strain LF580 producing anti-cancer active scopularide A and B. PLoS ONE 2014, 9, e103320. [Google Scholar] [CrossRef] [PubMed]
- Duetz, W.A.; Ruedi, L.; Hermann, R.; O’Connor, K.; Buchs, J.; Witholt, B. Methods for intense aeration, growth, storage, and replication of bacterial strains in microtiter plates. Appl. Environ. Microbiol. 2000, 66, 2641–2646. [Google Scholar] [CrossRef]
- Samorski, M.; Müller-Newen, G.; Buchs, J. Quasi-continuous combined scattered light and fluorescence measurements: A novel measurement technique for shaken microtiter plates. Biotechnol. Bioeng. 2005, 92, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Kensy, F.; Zang, E.; Faulhammer, C.; Tan, R.K.; Buchs, J. Validation of a high-throughput fermentation system based on online monitoring of biomass and fluorescence in continuously shaken microtiter plates. Microb. Cell. Fact. 2009, 4, 1–17. [Google Scholar] [CrossRef]
- Joint, I.; Muhling, M.; Querellou, J. Culturing marine bacteria—An essential prerequisite for biodiscovery. Microb. Biotechnol. 2010, 3, 564–575. [Google Scholar] [CrossRef]
- Monaghan, R.L.; Barrett, J.F. Antibacterial drug discovery-Then, now and the genomics future. Biochem. Pharmacol. 2006, 71, 901–909. [Google Scholar] [CrossRef]
- Kniemeyer, O. Proteomics of eukaryotic microorganisms: The medically and biotechnologically important fungal genus Aspergillus. Proteomics 2011, 11, 3232–3243. [Google Scholar] [CrossRef]
- Kramer, A.; Beck, H.C.; Kumar, A.; Kristensen, L.P.; Imhoff, J.F.; Labes, A. Proteomic analysis of anti-cancerous scopularide production by a marine Microascus brevicaulis strain and its UV mutant. PLoS ONE 2015, 10, e0140047. [Google Scholar] [CrossRef]
- Bhadury, P.; Mohammad, B.T.; Wright, P.C. The current status of natural products from marine fungi and their potential as anti-infective agents. J. Ind. Microbiol. Biotechnol. 2006, 33, 325–337. [Google Scholar] [CrossRef]
- Lang, S.; Hüners, M.; Lurtz, V. Bioprocess engineering data on the cultivation of marine prokaryotes and fungi. Adv. Biochem. Eng. Biotechnol. 2005, 97, 29–62. [Google Scholar] [CrossRef]
- Xu, R.; Li, X.; Xu, G.; Wang, B. Optimizing production of asperolide A, a potential anti-tumor tetranorditerpenoid originally produced by the algal-derived endophytic fungus Aspergillus wentii EN-48*. Chin. J. Oceanol. Limn. 2017, 35, 658–663. [Google Scholar] [CrossRef]
- Zhou, W.; Cai, M.; Zhou, J.; Jiang, T.; Zhou, J.; Wang, M.; Zhou, X.; Zhang, Y. Nutrition and bioprocess development for efficient biosynthesis of an antitumor compound from marine-derived fungus. J. Ind. Microbiol. Biotechnol. 2013, 40, 1131–1142. [Google Scholar] [CrossRef]
- Zhou, W.; Cai, M.; Na, K.; Shen, C.; Zhang, X.; Zhou, X.; Zhao, W.; Zhang, Y. pH-Dependent accumulation of anticancer compound on mycelia in fermentation of marine fungus. J. Ind. Microbiol. Biotechnol. 2014, 41, 1169–1173. [Google Scholar] [CrossRef]
- Papagianni, M. Advances in citric acid fermentation by Aspergillus niger: Biochemical aspects, membrane transport and modeling. Biotechnol. Adv. 2007, 25, 244–263. [Google Scholar] [CrossRef]
- Schneider, P.; Misiek, M.; Hoffmeister, D. In vivo and in vitro production options for fungal secondary metabolites. Mol. Pharm. 2008, 5, 234–242. [Google Scholar] [CrossRef]
- Jesus, H.C.R.; Jeller, A.H.; Debonsi, H.M.; Alves, P.B.; Porto, A.L.M. Multiple Monohydroxylation Products from rac-Camphor by Marine Fungus Botryosphaeria sp. Isolated from Marine Alga Bostrychia radicans. J. Braz. Chem. Soc. 2017, 28, 498–504. [Google Scholar] [CrossRef]
- Xiong, Z.Q.; Wang, J.F.; Hao, Y.Y.; Wang, Y. Recent advances in the discovery and development of marine microbial natural products. Mar. Drugs 2013, 11, 700–717. [Google Scholar] [CrossRef]
- Rutledge, P.J.; Challis, G.L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Rev. Microbiol. 2015, 13, 509–523. [Google Scholar] [CrossRef]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R. The re-emergence of natural products for drug Discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef]
- Elander, R.P. Industrial production of β-lactam antibiotics. Appl. Microbiol. Biotechnol. 2003, 61, 385–392. [Google Scholar] [CrossRef]
- Jungbauer, A. Continuous downstream processing of biopharmaceuticals. Trends Biotechnol. 2013, 31, 479–492. [Google Scholar] [CrossRef]
- Zydney, A.L. Continuous downstream processing for high value biological products: A review. Biotechnol. Bioeng. 2016, 113, 465–475. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Endophytic Fungi | Sterilization Methods | Host Macroalgae | Compounds | Chemical Class | Cell line/Target Enzyme | Activity (IC50) | Rf |
|---|---|---|---|---|---|---|---|
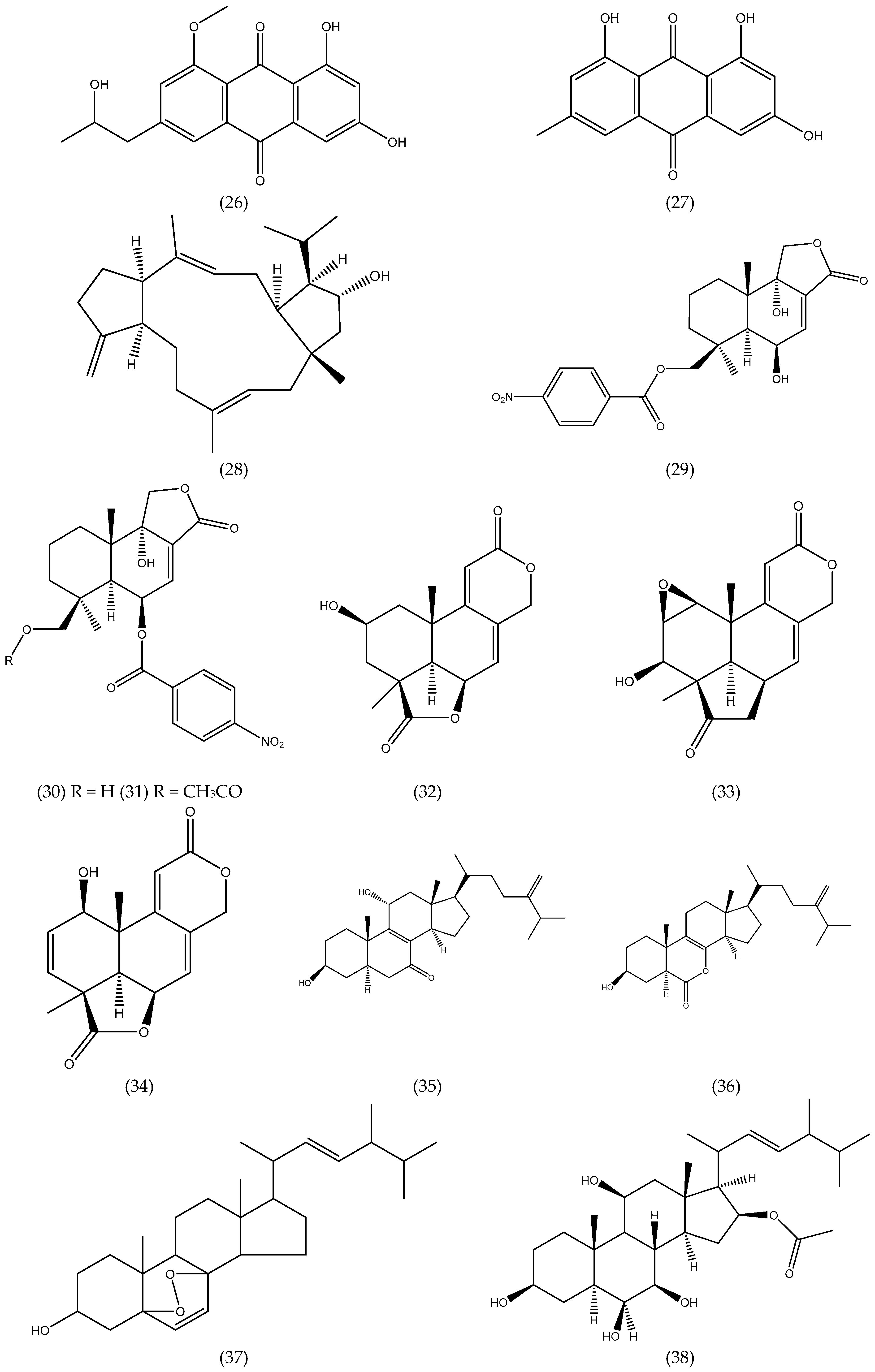
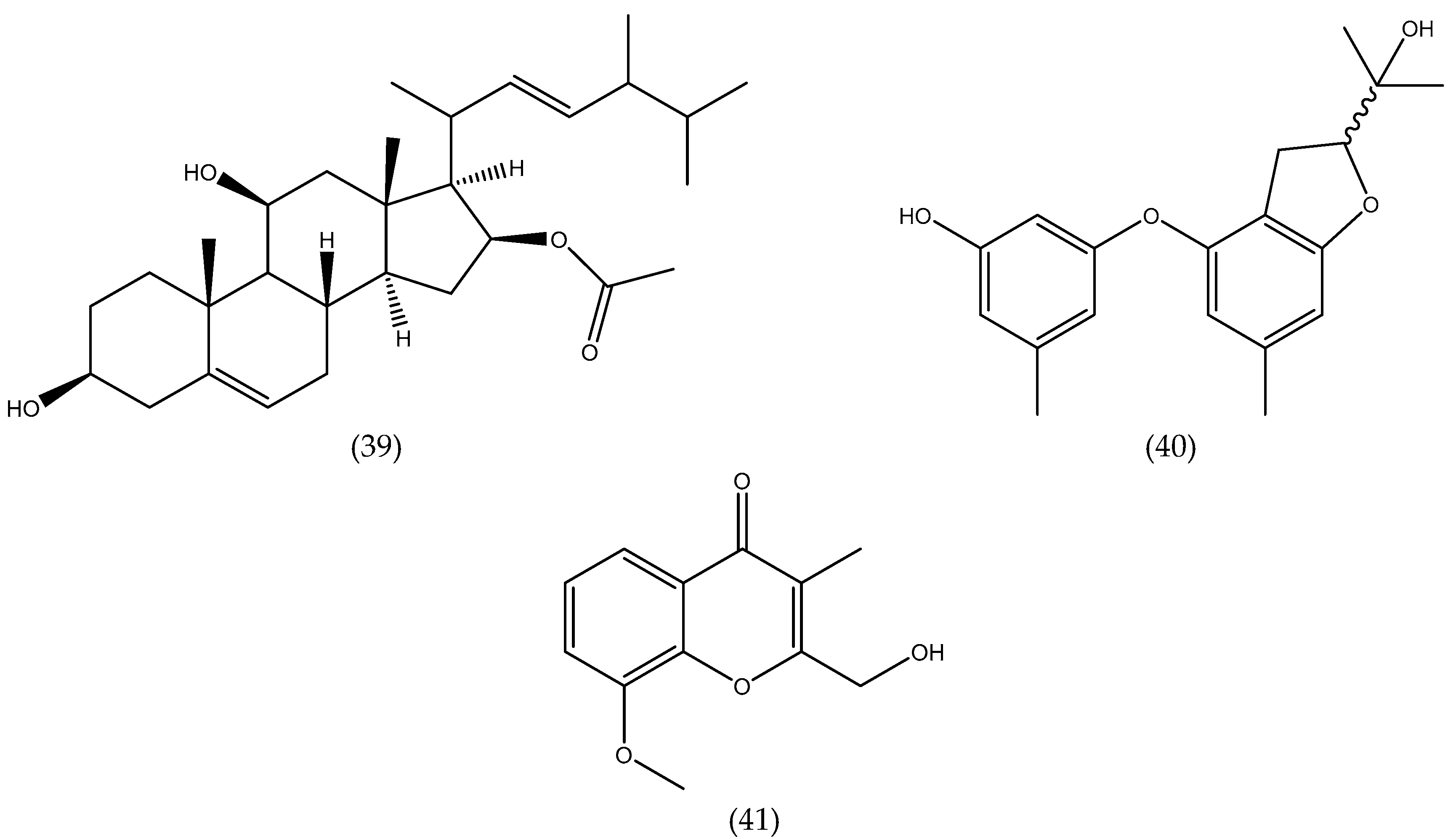
| Aspergillus ochraceus EM-31 | - | Sargassu kjellmanianum (B) | 7-nor-ergosterolide; 3β,11α-dihydroxyergosta-8,24(28)-dien-7-one; 3β-hydroxyergosta-8,24(28)-dien-7-one; (22E,24R)-3β,5α,9α-trihydroxyergosta-7,22- dien-6-one; (22E,24R)-3β,5α- dihydroxyergosta-7,22-dien-6-one;ergosterol; (22E,24R)-ergosta-4,6,8(14),22-tetraen-3-one; (22E,24R)-ergosta-7,22-diene- 3β,5α,6α-triol; (22E,24R)-ergosta-7,22-diene-6β- methoxy-3β,5α-diol; (22E,24R)-ergosta-7,22- diene-3β,6β-diol; (22E,24R)-ergosta-5α,6α- epoxide-8,22-diene-3β,7α-diol; (22E,24R)-5α,8α -epidioxyergosta-6,22-dien-3β-ol | steroids | NCI-H460, SW1990, SMMC-7721, HeLa, DU145, HepG2, MCF-7 | 5.00–28.00 µg mL−1 | [39] |
| Aspergillus ochraceus Jcma1F17 | (1) Rinsed 3x with sterile sea water; (2) 60–120 s 70% EtOH; (3) Rinsed with sterile artificial sea water. | Coelarthrum sp. (R) | 6β,9α-dihydroxy-14-p-nitrobenzoylcinnamolide; insulicolide A | terpenoids | H1975, U937, K562, BGC-823, Molt-4, MCF-7, A549, HeLa, HL60, Huh-7 | 1.95–9.40μM | [40] |
| Aspergillus ochraceus Jcma1F17 | (1) Rinsed 3x with sterile sea water; (2) 60–120 s 70% EtOH; (3) Rinsed with sterile artificial sea water. | Coelarthrum sp. (R) | insulicolide B; 14-O-acetylinsulicolide A; insulicolide C;6β,9α-dihydroxy-14-p- nitrobenzoylcinnamolide; insulicolide A; 9-deoxyinsulicolide A | terpenoids | ACHN, OS-RC-2, 786-O | 0.89–8.20 μM | [41] |
| Aspergillus tennesseensis | (1) 15 s 70% EtOH; (2) Rinsed in sterile water | Not identified | diorcinol L; (R)-diorcinol B; (S)-diorcinol B; 9-acetyldiorcinol B; diorcinol C; diorcinol D; diorcinol E; diorcinolJ; 3-(2-(1-hydroxy-1- methyl-ethyl)-6-methyl-2,3- dihydrobenzofuran-4-yloxy)-5-methylphenol | ethers | THP-1, A559, Du145 HeLa, MCF-7 MDA-MB-231 | 7.00–50.00 µg mL−1 | [42] |
| Aspergillus versicolor | - | Halimeda opuntia (G) | isorhodoptilometrin-1-methyl ether; emodin; 1-methyl emodin;evariquinone; 7-hydroxyemodin-6,8-methyl ether; siderin; arugosin C; variculanol | quinines | Murine L1210, CCRF-CEM, Murine colon 38, HCT-116, H-125, HepG2, CFU-GM | weak–mild | [43] |
| Aspergillus wentii EN-48 | (1) 15 s 70% EtOH; (2) Rinsed in sterile water | Sargassum sp. (B) | asperolides A−C; tetranorditerpenoid derivative; wentilactones A-B; botryosphaerin B; LL-Z1271-β | terpenoids | SMMC-7721, HeLa HepG2, MCF-7 MDA-MB-231 NCI-H460, SW1990 | 10.00–17.00 µM | [44] |
| Aspergillus wentii EN-48 | (1) 15 s 70% EtOH; (2) Rinsed in sterile water | Sargassum sp. (B) | wentilactone B | terpenoids | SMMC-7721, HepG2 Huh7, Hep3B | 18.96 µM (SMMC-7721) | [45] |
| Aspergillus wentii EN-48 | (1) 15 s 70% EtOH; (2) Rinsed in sterile water | Sargassum sp. (B) | wentilactone B | terpenoids | SMMC-7721 | - | [46] |
| Aspergillus wentii EN-48 | (1) 15 s 70% EtOH; (2) Rinsed in sterile water | Sargassum sp. (B) | wentilactone A | terpenoids | NCI-H460, NCI-H466 | - | [47] |
| Aspergillus wentii EN-48 | (1) 15 s 70% EtOH (2) Rinsed in sterile water | Sargassum sp. (B) | asperolide A | terpenoids | NCI-H460 | - | [48] |
| Aspergillus sp. XNM-4 | - | Leathesia nana (B) | (hydroxy(phenyl)methyl)-4H-pyran-4-one; 2-benzyl-4H-pyran-4-one; asperpyrone D; asperpyrone C; aurosperone B; fonsecinone B; asperpyrone B; dianhydro-aurasperone C; isoaurasperone A; aurasperone F; fonsecinone D; asperpyroneA; fonsecinone A; fonsecin; TMC 256 A1; flavasperone; carbonarone A; pestalamide A | polyketides | PANC-1, A549 MDA-MB-231, Caco-2, SK-OV-3 | 8.25 µM - potent on all cells | [49] |
| Chaetomium globosum QEN-14 | (1) 15 s 70% EtOH; (2) Rinsed with sterile water | Ulva pertusa(G) | cytoglobosins A-G; isochaetoglobosin D; chaetoglobosin Fex | alkaloids | A-549, P388, KB | 2.26–10.00 µM | [50] |
| Cladosporium cladosporioides EN-399 | (1) 15 s 70% EtOH; (2) Rinsed with sterile water | Laurencia okamurai (R) | cladosporols F−I; cladosporol C; cladosporol J | polyketides | A549, Huh7, LM3, H446 | 1.00–5.00 µM | [51] |
| Coniothyrium cereale | (1) Rinsed with sterile H2O (3x); (2) 15 s 70% EtOH; (3) Rinsed in sterile artificial seawater (ASW) | Enteromorpha sp. (G) | coniosclerodin; (Z)-coniosclerodinol; (E)- coniosclerodinol; (15S, 17S)-(-)-sclerodinol; conioscleroderolide; coniosclerodione; coniolactone; (-)-7,8-dihydro-3,6-dihydroxy- 1,7,7,8-tetramethyl-5H-furo-[2’,3’:5,6]naphtho [1,8-bc]furan-5-one; (-)-sclerodin A; lamellicolic anhydride; (-) scleroderolide; (-) sclerodione | polyketides | HTB-9, HLE | 7.20–20.00 µM | [52] |
| Gibberella zeae | - | Codium fragile (G) | 3-hydroxy-5-(hydroxymethyl)-4-(4’-hydroxy phenoxy)pyrrolidin-2-one; (22E,24R)-7β,8β-epoxy-3β,5α,9α-trihydroxy ergosta-22-en-6-one;(22E,24R)-3β,5α,9α-trihy droxyergosta-7,22-dien-6-one; (22E,24R)-3β, 5α-dihydroxyergosta-7,22-dien-6-one; (22E,24R)-ergosta-7,22-dien-3β,5α,6β-triol; (22E,24R)-ergosta-5,22-dien-3β-ol; (22E,24R)- 5α,8α-epidioxyergosta-6,22-dien-3β-ol; (22E,24R)-5α,8α-epidioxyergosta-6,9(11),22-trien-3β-ol; (22E,24R)-1(10→6)- abeoergosta-5,7,9,22-tetraen-3α-ol | alkaloids steroids | A-549 BEL-7402 | 17.60–61.80% | [53] |
| Guignardia sp. | (1) Washed with running tap water; (2) 1 min. 75% EtOH; (3) 5 min. 2.5% NaOCl; (4) Rinsed with sterile H2O (3x) | Undaria pinnatifida (B) | 6, 22-diene-5, 8-epidioxyergosta-3-ol; ergosterol; cyclo-(Tyr-Leu); cyclo-(Phe-Phe); cyclo-(Val-Leu); cyclo-(Phe-Pro); cyclo-(Leu-Ile) | steroids peptides | KB | 10.00–50.00 µg mL−1, | [54] |
| Paecilomyces variotii EN-291 | (1) 15 s 70% EtOH (2) Rinsed in sterile water | Not identified (R) | dihydrocarneamide A; iso-notoamide B | alkaloids | NCI-H460 | 55.90–69.30 µmol L−1 | [55] |
| Paecilomycesvariotii EN-291 | (1) 15 s 70% EtOH (2) Rinsed in sterile water | Grateloupia turuturu (R) | varioloid A; varioloid B | alkaloids | A549, HCT116, HepG2 | 2.50–8.20 µg mL−1 | [56] |
| Penicillium chrysogenum QEN-24S | (1) 15 s 70% EtOH (2) Rinsed in sterile water. | Laurencia sp. (R) | penicitides A-B; 2-(2,4-dihydroxy-6-methyl benzoyl)-glycerol; (2,4-dihydroxy-6-methyl benzoyl)-glycerol; penicimonoterpene | polyketides terpenoid | HepG2, NCI-H460, SMMC-7721, SW1990, DU145, Hela, MCF-7 | 32.00–40.00 µg mL−1 | [57] |
| Penicillium chrysogenum QEN-24S | - | Laurencia sp. (R) | penicisteroids A-B; anicequol; (22E, 24R)-ergosta-4,6,8(14),22-tetraen-3-one; (22E, 24R)-ergosta-7,22-dien-3,6-dione; (22E, 24R)-5α,8α-epidioxyergosta-6,22-dien-3β-ol; (22E, 24R)-ergosta-5α,6α-epoxide-8, 22-dien-3β,7α-diol; (22E, 24R)-ergosta-7,22-dien-3β,5α,6β-triol; (22E, 24R)-ergosta-7,22-dien-3β,6β-diol | steroids | HeLa SW1990 NCI-H460 | 15.00–40.00 µg mL−1 | [58] |
| Penicillium sp. | (1) 15 s 70% EtOH; (2) Rinsed in sterile water | Ulva sp. (G) | chromanone A | chromone | Hepa1c1c7, Cyp1A | 4.00 µg mL−1 | [59] |
| Phoma sp. NTOU4195 | - | Pterocladiella capillacea (R) | phomaketides A-E; pseurotins A3 and G; FR-111142, pseurotins A, A1, A2, D and F2, 14-norpseurotin A; A-carbonylcarbene; tyrosol; cyclo(-L-Pro-L-Leu); cyclo(-L-Pro-L-Phe) | polyketides alkaloids phenylethanoid peptides | Endothelial progenitor cells (EPCs) | 8.10–19.20 µM | [60] |
| Talaromyce sislandicus EN-501 | (1) 15 s 70% EtOH (2) Rinsed in sterile water | Laurencia okamurai (R) | 8-hydroxyconiothyrinone B; 8,11-dihydroxyconiothyrinone; 4R,8-dihydroxyconiothyrione B; 4S,8-dihydroxyconiothyrinone B; 4S,8-dihydroxy-10-O-methyldendryol E | quinines | A2780, A2780 CisR | <10.00 µM | [61] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, T.R.; Santos, G.S.d.; Armstrong, L.; Colepicolo, P.; Debonsi, H.M. Antitumor Potential of Seaweed Derived-Endophytic Fungi. Antibiotics 2019, 8, 205. https://doi.org/10.3390/antibiotics8040205
Teixeira TR, Santos GSd, Armstrong L, Colepicolo P, Debonsi HM. Antitumor Potential of Seaweed Derived-Endophytic Fungi. Antibiotics. 2019; 8(4):205. https://doi.org/10.3390/antibiotics8040205
Chicago/Turabian StyleTeixeira, Thaiz Rodrigues, Gustavo Souza dos Santos, Lorene Armstrong, Pio Colepicolo, and Hosana Maria Debonsi. 2019. "Antitumor Potential of Seaweed Derived-Endophytic Fungi" Antibiotics 8, no. 4: 205. https://doi.org/10.3390/antibiotics8040205
APA StyleTeixeira, T. R., Santos, G. S. d., Armstrong, L., Colepicolo, P., & Debonsi, H. M. (2019). Antitumor Potential of Seaweed Derived-Endophytic Fungi. Antibiotics, 8(4), 205. https://doi.org/10.3390/antibiotics8040205