
Identification of a Potential High-Risk Clone and Novel Sequence Type of Carbapenem-Resistant Pseudomonas aeruginosa in Metro Manila, Philippines

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Results
2.1. Information on Sources of Isolates
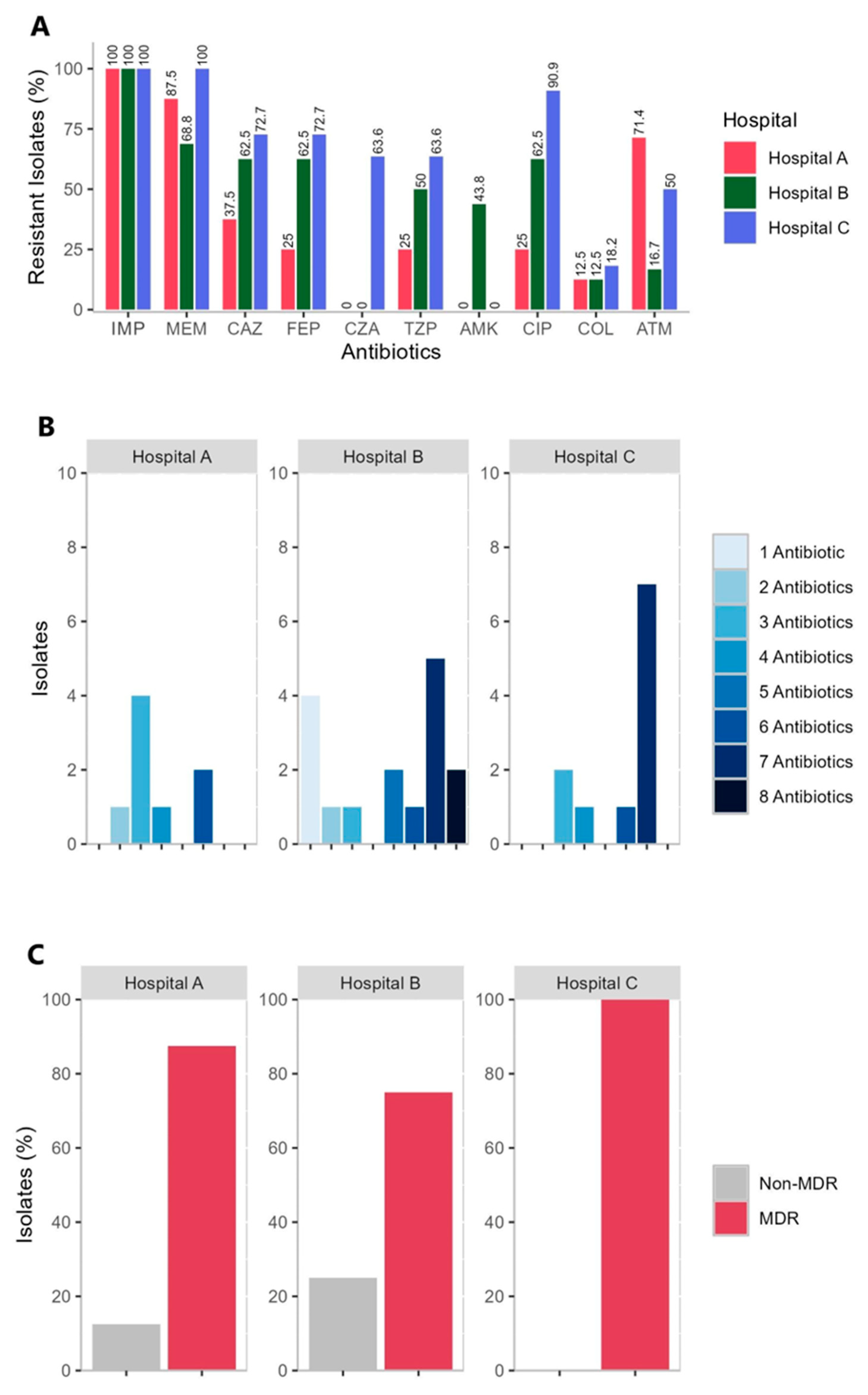
2.2. Antimicrobial Susceptibility of the CRPA Isolates
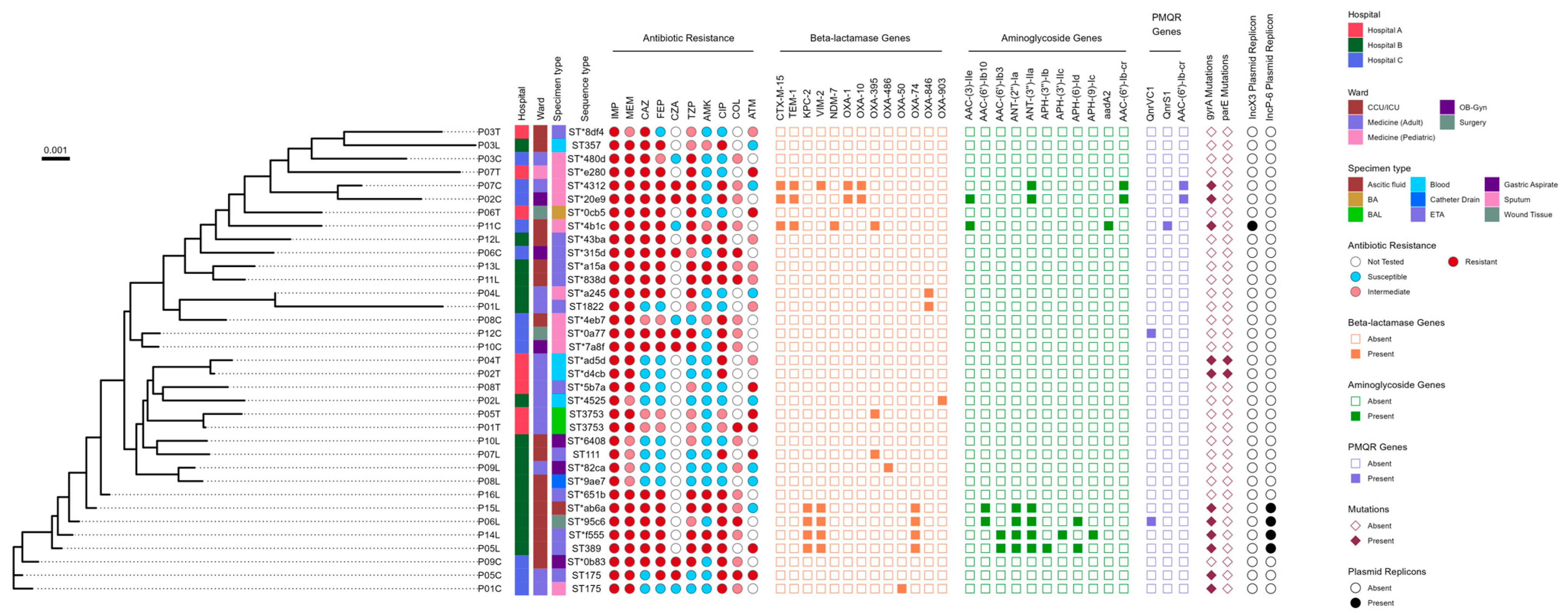
2.3. Genotyping Results Using MLST Analysis
2.4. The Phenotypic and Genotypic Profile of 35 CRPA Isolates
2.5. Presence of Beta-Lactamases
2.6. The Presence of Plasmid-Mediated Quinolone Resistance (PMQR) Genes and Mutations in the Quinolone Resistance-Determining Regions (QRDRs) of the gyrA and parE Genes
2.7. Presence of Aminoglycoside Resistance Genes
2.8. Antimicrobial Resistance Genes Flanked by Mobile Genetic Elements (MGEs)
2.9. Presence of OprD Mutations
3. Discussion
4. Materials and Methods
4.1. Isolation and Collection of P. aeruginosa
4.2. Antimicrobial Susceptibility Test
4.3. Whole-Genome Sequencing
4.4. Data Processing and Analytics
4.5. Identification of AMR Genes, Plasmid Replicons, and Flanking MGEs
4.6. Genotyping Using Traditional MLST Analysis
4.7. Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WHO Bacterial Priority Pathogen List; World Health Organization, Ed.; CC BY-NC-SA 3.0 IGO; WHO: Geneva, Switzerland, 2024. [Google Scholar]
- Raman, G.; Avendano, E.E.; Chan, J.; Merchant, S.; Puzniak, L. Risk factors for hospitalized patients with resistant or multidrug-resistant Pseudomonas aeruginosa infections: A systematic review and meta-analysis. Antimicrob. Resist. Infect. Control 2018, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Bogiel, T.; Prażyńska, M.; Kwiecińska-Piróg, J.; Gospodarek-Komkowska, E.; Mikucka, A. Carbapenem-resistant Pseudomonas aeruginosa strains-distribution of the essential enzymatic virulence factors genes. Antibiotics 2021, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Amsalu, A.; Sapula, S.A.; De Barros Lopes, M.; Hart, B.J.; Nguyen, A.H.; Drigo, B.; Turnidge, J.; Leong, L.E.; Venter, H. Efflux pump-driven antibiotic and biocide cross-resistance in Pseudomonas aeruginosa isolated from different ecological Niches: A case study in the development of multidrug resistance in environmental hotspots. Microorganisms 2020, 8, 1647. [Google Scholar] [CrossRef] [PubMed]
- Arzanlou, M.; Chai, W.C.; Venter, H. Intrinsic, adaptive and acquired antimicrobial resistance in Gram-negative bacteria. Essays Biochem. 2017, 61, 49–59. [Google Scholar]
- Li, H.; Luo, Y.F.; Williams, B.J.; Blackwell, T.S.; Xie, C.M. Structure and function of OprD protein in Pseudomonas aeruginosa: From antibiotic resistance to novel therapies. Int. J. Med. Microbiol. 2012, 302, 63–68. [Google Scholar] [CrossRef]
- Shu, J.C.; Kuo, A.J.; Su, L.H.; Liu, T.P.; Lee, M.H.; Su, I.N.; Wu, T.L. Development of carbapenem resistance in Pseudomonas aeruginosa is associated with OprD polymorphisms, particularly the amino acid substitution at codon 170. J. Antimicrob. Chemother. 2017, 72, 2489–2495. [Google Scholar] [CrossRef]
- Shariati, A.; Azimi, T.; Ardebili, A.; Chirani, A.S.; Bahramian, A.; Pormohammad, A.; Sadredinamin, M.; Erfanimanesh, S.; Bostanghadiri, N.; Shams, S.; et al. Insertional inactivation of oprD in carbapenem-resistant Pseudomonas aeruginosa strains isolated from burn patients in Tehran, Iran. New Microbes New Infect. 2018, 21, 75–80. [Google Scholar] [CrossRef]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile β-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef]
- Amsalu, A.; Sapula, S.A.; Whittall, J.J.; Hart, B.J.; Bell, J.M.; Turnidge, J.; Venter, H. Worldwide distribution and environmental origin of the Adelaide imipenemase (AIM-1), a potent carbapenemase in Pseudomonas aeruginosa. Microb. Genom. 2021, 7, 000715. [Google Scholar] [CrossRef]
- Sahuquillo-Arce, J.M. Carbapenemases: A worldwide threat to antimicrobial therapy. World J. Pharmacol. 2015, 4, 75. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A. Updated Functional Classification of β-Lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Sawa, T.; Kooguchi, K.; Moriyama, K. Molecular Diversity of Extended-Spectrum β-Lactamases and Carbapenemases, and Antimicrobial Resistance. J. Intensive Care. 2020, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Kocsis, B.; Gulyás, D.; Szabó, D. Diversity and distribution of resistance markers in Pseudomonas aeruginosa international high-risk clones. Microorganisms 2021, 9, 359. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.J.; Jeong, S.H. Mobile Carbapenemase Genes in Pseudomonas aeruginosa. Front. Microbiol. 2021, 12, 614058. [Google Scholar] [CrossRef]
- Wang, M.G.; Liu, Z.Y.; Liao, X.P.; Sun, R.Y.; Li, R.B.; Liu, Y.; Fang, L.X.; Sun, J.; Liu, Y.H.; Zhang, R.M. Retrospective data insight into the global distribution of carbapenemase-producing Pseudomonas aeruginosa. Antibiotics 2021, 10, 548. [Google Scholar] [CrossRef]
- Arowolo, M.T.; Orababa, O.Q.; Olaitan, M.O.; Osibeluwo, B.V.; Essiet, U.U.; Batholomew, O.H.; Ogunrinde, O.G.; Lagoke, O.A.; Soriwei, J.D.; Ishola, O.D.; et al. Prevalence of carbapenem resistance in Acinetobacter baumannii and Pseudomonas aeruginosa in sub-Saharan Africa: A systematic review and meta-analysis. PLoS ONE 2023, 18, e0287762. [Google Scholar] [CrossRef]
- Sihombing, B.; Bhatia, R.; Srivastava, R.; Aditama, T.Y.; Laxminarayan, R.; Rijal, S. Response to Antimicrobial Resistance in South-East Region. Lancet Reg. Health Southeast Asia 2023, 18, 100306. [Google Scholar] [CrossRef]
- Department of Health. Antimicrobial Resistance Surveillance Reference Laboratory 2022, Annual Report [Internet]. 2022. Available online: https://arsp.com.ph (accessed on 20 December 2023).
- Department of Health. Antimicrobial Resistance Surveillance Reference Laboratory 2023, Annual Report [Internet]. 2023. Available online: https://arsp.com.ph (accessed on 27 November 2024).
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef]
- Oliver, A.; Mulet, X.; López-Causapé, C.; Juan, C. The Increasing Threat of Pseudomonas aeruginosa High-Risk Clones. Drug Resistance Updates. Drug Resist. Updat. 2015, 21–22, 41–59. [Google Scholar] [CrossRef]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and treatment of multidrug-resistant and extensively drug-resistant Pseudomonas aeruginosa infections. Clin. Microbiol. Rev. 2019, 32, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Abordo, A.M.S.; Carascal, M.B.; Remenyi, R.; Dalisay, D.S.; Saludes, J.P. Clinically Isolated β-Lactam-Resistant Gram-Negative Bacilli in a Philippine Tertiary Care Hospital Harbor Multi-Class β-Lactamase Genes. Pathogens 2023, 12, 1019. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Nazareno, P.J.; Nakano, R.; Mondoy, M.; Nakano, A.; Bugayong, M.P.; Bilar, J.; Perez, M.; Medina, E.J.; Saito-Obata, M.; et al. Environmental Presence and Genetic Characteristics of Carbapenemase-Producing Enterobacteriaceae from Hospital Sewage and River Water in the Philippines. Appl. Env. Microbiol. 2019, 86, e01906-19. [Google Scholar] [CrossRef] [PubMed]
- Manohar, P.; Leptihn, S.; Lopes, B.S.; Nachimuthu, R. Dissemination of carbapenem resistance and plasmids encoding carbapenemases in Gram-negative bacteria isolated in India. JAC Antimicrob. Resist. 2021, 3, dlab015. [Google Scholar] [CrossRef] [PubMed]
- Reuland, E.A.; Halaby, T.; Hays, J.P.; de Jongh, D.M.; Snetselaar, H.D.; van Keulen, M.; Elders, P.J.; Savelkoul, P.H.; Vandenbroucke-Grauls, C.M.; Al Naiemi, N. Plasmid-mediated AmpC: Prevalence in community-acquired isolates in Amsterdam, the Netherlands, and risk factors for carriage. PLoS ONE 2015, 10, e0113033. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Peng, W.; Wu, Y.; Li, H.; Wang, Q.; Yi, H.; Zhang, R.; Shao, B.; Zhu, K. A Potential High-Risk Clone of Pseudomonas aeruginosa ST463. Front. Microbiol. 2021, 12, 670202. [Google Scholar] [CrossRef]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Nieto-Saucedo, J.R.; López-Jacome, L.E.; Franco-Cendejas, R.; Colín-Castro, C.A.; Hernández-Duran, M.; Rivera-Garay, L.R.; Zamarripa-Martinez, K.S.; Mosqueda-Gómez, J.L. Carbapenem-Resistant Gram-Negative Bacilli Characterization in a Tertiary Care Center from El Bajio, Mexico. Antibiotics 2023, 12, 1295. [Google Scholar] [CrossRef]
- Cabot, G.; Ocampo-Sosa, A.A.; Domínguez, M.A.; Gago, J.F.; Juan, C.; Tubau, F.; Rodríguez, C.; Moyà, B.; Peña, C.; Martínez-Martínez, L.; et al. Genetic markers of widespread extensively drug-resistant Pseudomonas aeruginosa high-risk clones. Antimicrob. Agents Chemother. 2012, 56, 6349–6357. [Google Scholar] [CrossRef]
- Pollini, S.; Maradei, S.; Pecile, P.; Olivo, G.; Luzzaro, F.; Docquier, J.D.; Rossolini, G.M. FIM-1, a new acquired metallo-β-lactamase from a Pseudomonas aeruginosa clinical isolate from Italy. Antimicrob. Agents Chemother. 2013, 57, 410–416. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Supplement M100, Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; CLSI: Wayne, PA, USA, 2022. [Google Scholar]
- Abulencia, M.F.; Tesalona, S.; Pineda-Cortel, M.R.; Manahan, E.; Lagamayo, E. GitHub—Abulenciamiguel/EpiTomas: Antimicrobial Resistance. GitHub. Available online: https://github.com/abulenciamiguel/EpiTomas (accessed on 26 February 2023).
- Pedersen, B.S.; Quinlan, A.R. Mosdepth: Quick coverage calculation for genomes and exomes. Bioinformatics 2018, 34, 867–868. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly Landes Biosci. 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Antipov, D.; Hartwick, N.; Shen, M.; Raiko, M.; Lapidus, A.; Pevzner, P.A. PlasmidSPAdes: Assembling plasmids from whole genome sequencing data. Bioinformatics 2016, 32, 3380–3387. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In Silico detection and typing of plasmids using plasmidfinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Tönnies, H.; Prior, K.; Harmsen, D.; Melman, A. Establishment and evaluation of a core genome multilocus sequence typing scheme for whole-genome sequence-based typing of Pseudomonas aeruginosa. J. Clin. Microbiol. 2021, 59, 10-1128. [Google Scholar] [CrossRef]
- Zhou, Z.; Alikhan, N.F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carriço, J.A.; Achtman, M. Grapetree: Visualization of core genomic relationships among 100,000 bacterial pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics | No. of Isolates at an MIC (mg/L) of: | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.125 | 0.250 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | Breakpoint(s) b | R (%) | |
| IMP d, | 35 +,a | ≤2/≥8 | 100.0 | ||||||||||
| MEM d | 6 | 3 +,a | 1, 25 +,a | ≤2/≥8 | 82.9 | ||||||||
| CAZ d | 11 | 3, 7 + | 1+,a | 13 +,a | ≤8/≥32 | 60.0 | |||||||
| FEP d | 5 * | 3 | 3 | 4, 6 + | 1 a | 13 +,a | ≤8/≥32 | 57.1 | |||||
| CZA e | 1 | 3 | 7 + | (≤8/4)/(≥16/4) | 63.6 | ||||||||
| TZP d | 4 * | 1 | 4 | 9 | 12 + | 5 +,a | (≤16/4)/(≥128/4) | 48.6 | |||||
| COL e | 9 * | 6 | 4 a | 1 +,a | ≤2/≥4 | 25.0 | |||||||
| CIP d | 1 * | 4 * | 6 * | 2 | 10 +,a | 11 +,a | 1 +,a | ≤0.5/≥2 | 62.9 | ||||
| AMK d | 10 * | 3 | 9 * | 3 | 3 | 7 +,a | <16/≥64 | 20.0 | |||||
| No. of isolates at zone diameter (mm) b of: | |||||||||||||
| 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | Breakpoint(s) c | ||
| ATM f | 1 a | 7 a | 1 | 1 | 4 | 7 | ≥22/≤15 | 38.1 | |||||
| OprD Type(s) | Resistance Phenotype(s) to IPM | Resistance Phenotype(s) to MEM | Isolates | OprD Type Classification | Mutations | Known to Cause Carbapenem Resistance | OprD Structure(s) Affected |
|---|---|---|---|---|---|---|---|
| T1 | Resistant | Resistant | P02T, P08C, P10C, P12C, P11L, P12L, P13L | Full-length type, Wild-type | None | None | None |
| T1 | Resistant | Intermediate | P07L, P09L | Full-length type, Wild-type | None | None | None |
| T1-I | Resistant | Resistant | P04L, P06C, P04T, P06T | Full-length type, without substitution of amino acid residue | None | None | None |
| T1-I | Resistant | Intermediate | P10L | Full-length type, without substitution of amino acid residue | None | None | None |
| T1-Ia | Resistant | Intermediate | P02L | Full-length type, substitution of 1 amino acid residue | Phe205Ile | Yes | Alters hydrophobicity and stability of the OprD channel |
| T1-Ia | Resistant | Intermediate | P03T | Full-length type, substitution of 1 amino acid residue | Val379Leu | Yes | Affects pore size and shape, reducing drug entry |
| T1-Ia | Resistant | Resistant | P16L | Full-length type, substitution of 1 amino acid residue | Ile628,629Ala | Yes | Impacts structural integrity and function of the OprD channel |
| T1-Ia | Resistant | Intermediate | P08L | Full-length type, substitution of 1 amino acid residue | Ile628,629Ala | Yes | Impacts structural integrity and function of the OprD channel |
| T1-II | Resistant | Resistant | P08T | Full-length type, with several polymorphisms | Lys344Thr, Phe508Leu | Unknown | None |
| T1-IIa | Resistant | Resistant | P05C | Full-length type, with several polymorphisms | Glu604Gln, Ile628,629Ala | Yes Yes | Affects L5, Impacts structural integrity and function of the OprD channel. |
| T1-IIb | Resistant | Resistant | P03C | Full-length type, with several polymorphisms | Glu688Lys, Ser719Thr | Yes Unknown | Affects L6 and electrostatic interactions, altering pore dynamics |
| T1-III | Resistant | Resistant | P07T, P09C, P01C | Full-length type, with several polymorphisms | Glu604Gln Ile628,629Ala Glu688Lys Ser719Thr | Yes Yes Yes Unknown | Affects L5, L6, structural integrity and function of the OprD channel, and electrostatic interactions, altering pore dynamics |
| T1-IIIa | Resistant | Resistant | P01L | Full-length type, with several polymorphisms | Thr308Ser Arg928,929Glu Ala944,945Gly Gly1,274Ala | Unknown Unknown Unknown Unknown | None |
| T1-IV | Resistant | Resistant | P03L | Full-length type, with several polymorphisms | Ser169,170,171Glu Ser175,177Arg Val379Leu Glu688Lys Ser719Thr Gln1,270Glu | Yes Yes Yes Yes Unknown Unknown | Affects L1, L3, L6, L8, altering pore characteristics |
| T2 | Resistant | Resistant | P01T, P05T | OprD deficient Type—Premature stop codon | Trp830 * (W830X) | Yes | Loss of porin function |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tesalona, S.D.; Abulencia, M.F.B.; Pineda-Cortel, M.R.B.; Sapula, S.A.; Venter, H.; Lagamayo, E.N. Identification of a Potential High-Risk Clone and Novel Sequence Type of Carbapenem-Resistant Pseudomonas aeruginosa in Metro Manila, Philippines. Antibiotics 2025, 14, 362. https://doi.org/10.3390/antibiotics14040362
Tesalona SD, Abulencia MFB, Pineda-Cortel MRB, Sapula SA, Venter H, Lagamayo EN. Identification of a Potential High-Risk Clone and Novel Sequence Type of Carbapenem-Resistant Pseudomonas aeruginosa in Metro Manila, Philippines. Antibiotics. 2025; 14(4):362. https://doi.org/10.3390/antibiotics14040362
Chicago/Turabian StyleTesalona, Sherill D., Miguel Francisco B. Abulencia, Maria Ruth B. Pineda-Cortel, Sylvia A. Sapula, Henrietta Venter, and Evelina N. Lagamayo. 2025. "Identification of a Potential High-Risk Clone and Novel Sequence Type of Carbapenem-Resistant Pseudomonas aeruginosa in Metro Manila, Philippines" Antibiotics 14, no. 4: 362. https://doi.org/10.3390/antibiotics14040362
APA StyleTesalona, S. D., Abulencia, M. F. B., Pineda-Cortel, M. R. B., Sapula, S. A., Venter, H., & Lagamayo, E. N. (2025). Identification of a Potential High-Risk Clone and Novel Sequence Type of Carbapenem-Resistant Pseudomonas aeruginosa in Metro Manila, Philippines. Antibiotics, 14(4), 362. https://doi.org/10.3390/antibiotics14040362