
Engineered Lysin-Derived Peptide as a Potent Antimicrobial for Acne Vulgaris


Abstract
1. Introduction
2. Results
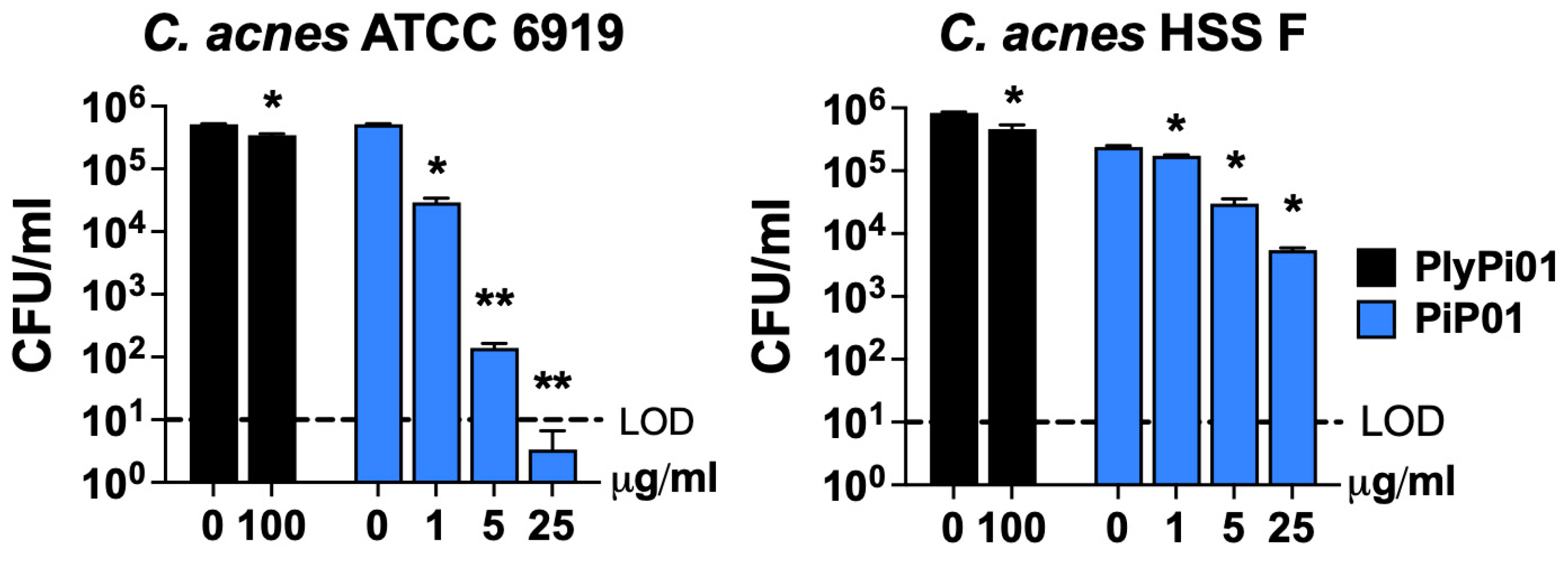
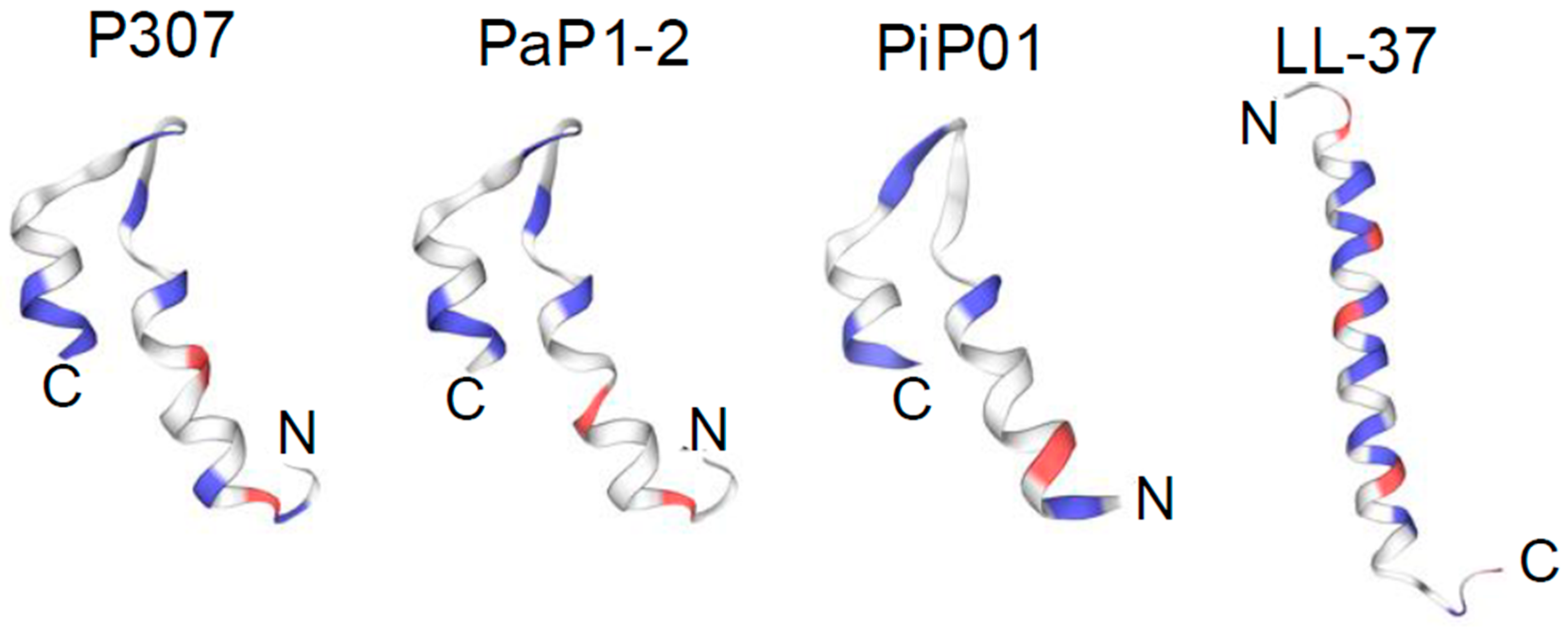
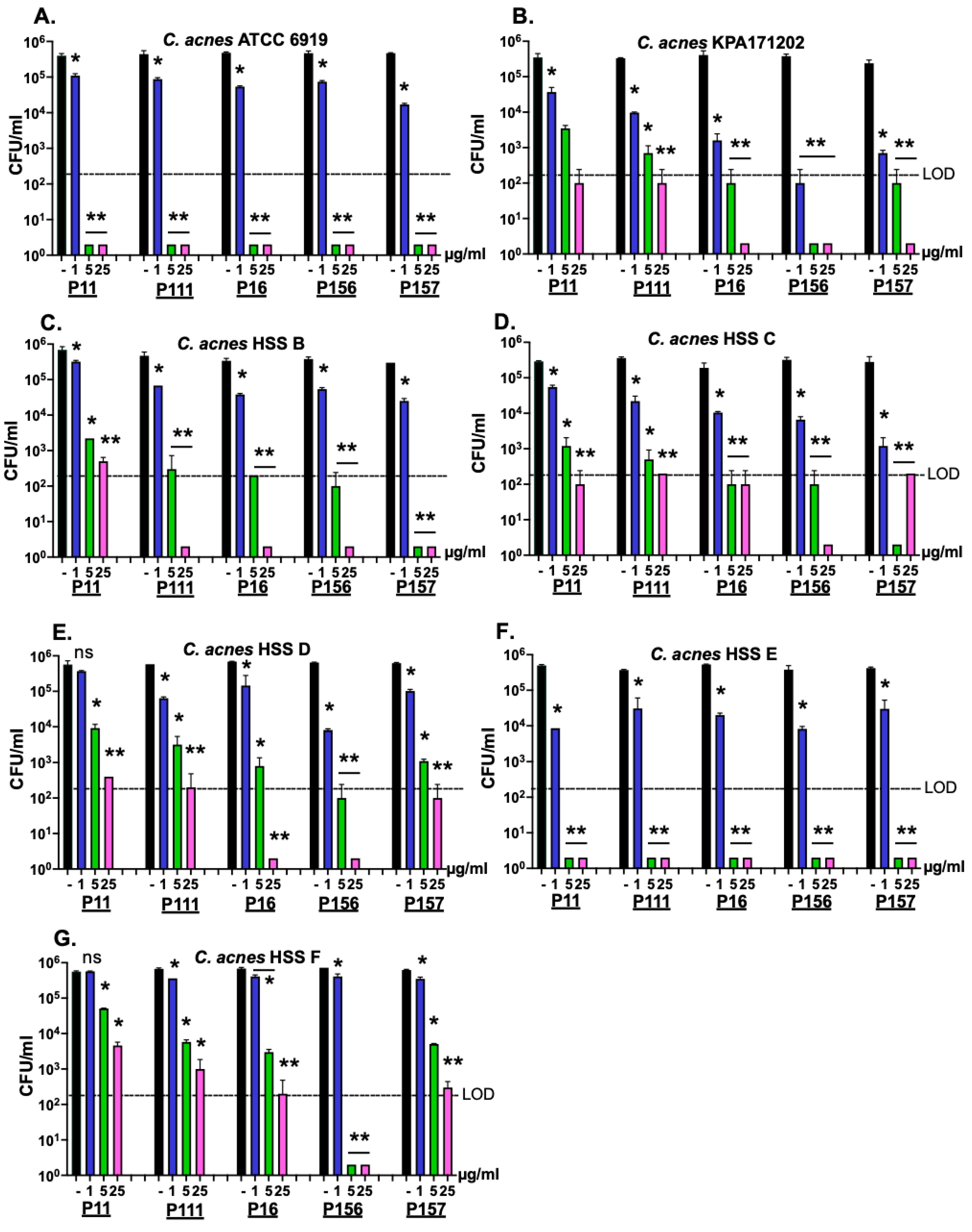
2.1. Modified PlyPi01-Derived Peptides Display Potent Bactericidal Activity
2.2. P156 Kills C. acnes Under Conditions Relevant to Skin Properties and Bacterial Growth
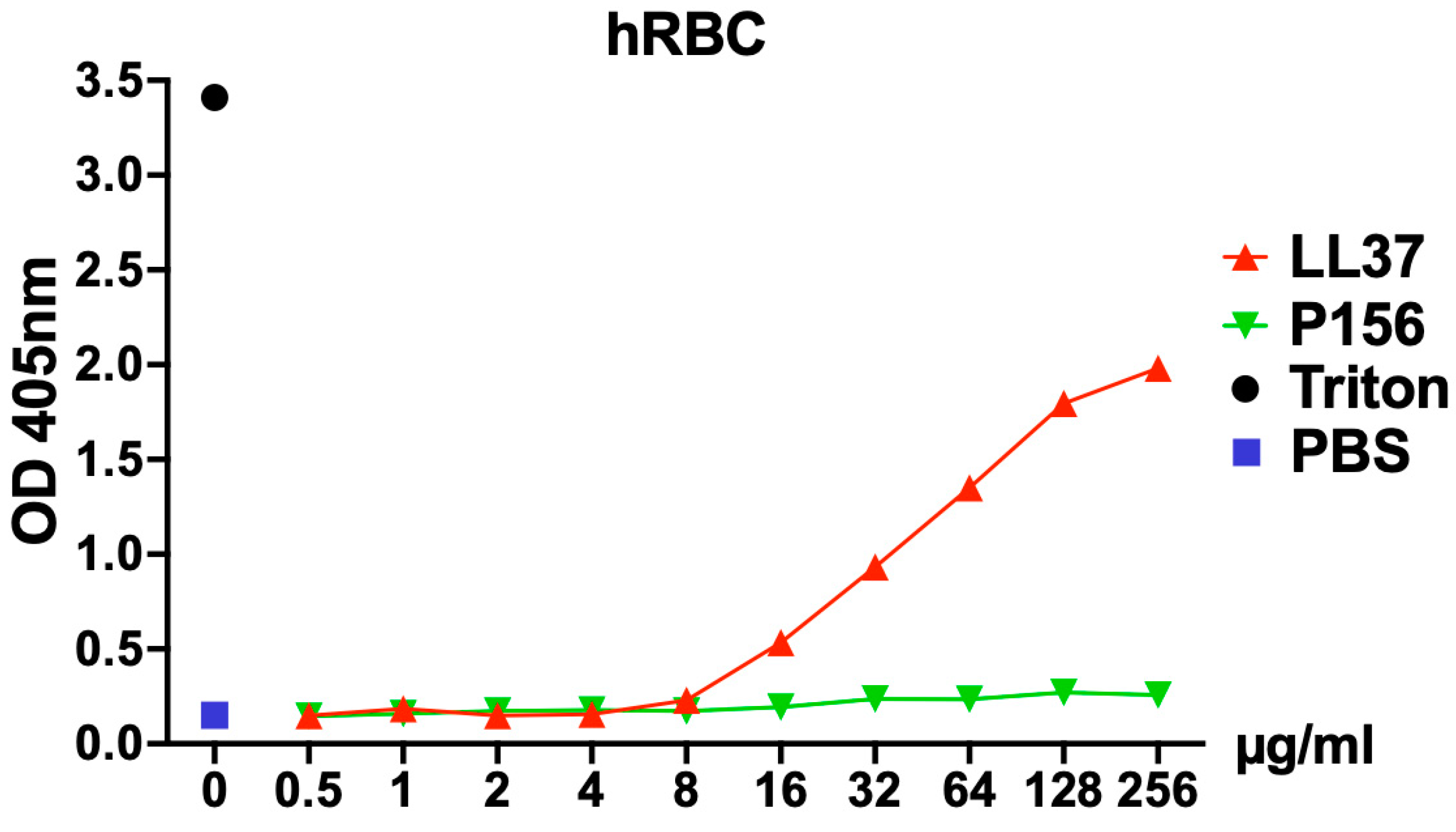
2.3. P156 Lacks Cytotoxicity Toward Human Cells
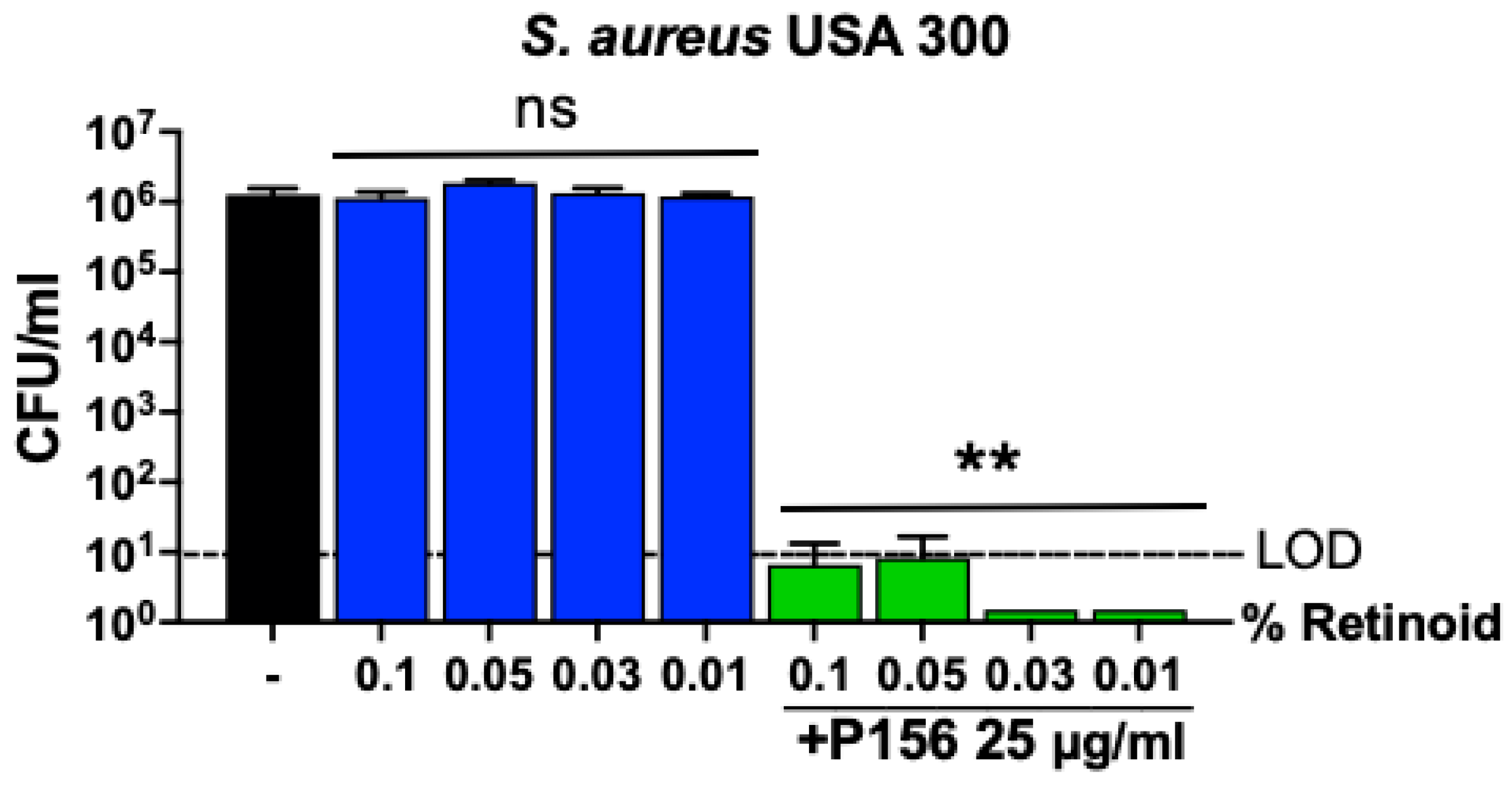
2.4. P156 Bactericidal Activity Is Preserved When Used in Combination with Current Common Acne Vulgaris Treatments
3. Discussion
4. Material and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. PlyPi01 Lysin Protein Expression and Purification
4.3. Peptide Synthesis and Properties
4.4. One-Hour Killing Assays
4.5. Time–Kill Assay
4.6. Cytotoxicity Assays
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zaenglein, A.L. Acne Vulgaris. N. Engl. J. Med. 2018, 379, 1343–1352. [Google Scholar] [PubMed]
- Williams, H.C.; Dellavalle, R.P.; Garner, S. Acne vulgaris. Lancet 2012, 379, 361–372. [Google Scholar] [PubMed]
- Dessinioti, C.; Katsambas, A. The Microbiome and Acne: Perspectives for Treatment. Dermatol. Ther. 2024, 14, 31–44. [Google Scholar]
- Moradi Tuchayi, S.; Makrantonaki, E.; Ganceviciene, R.; Dessinioti, C.; Feldman, S.R.; Zouboulis, C.C. Acne vulgaris. Nat. Rev. Dis. Primers 2015, 1, 15029. [Google Scholar]
- Tanghetti, E.A. The role of inflammation in the pathology of acne. J. Clin. Aesthet. Dermatol. 2013, 6, 27–35. [Google Scholar]
- Huang, L.; Yang, S.; Yu, X.; Fang, F.; Zhu, L.; Wang, L.; Zhang, X.; Yang, C.; Qian, Q.; Zhu, T. Association of different cell types and inflammation in early acne vulgaris. Front. Immunol. 2024, 15, 1275269. [Google Scholar]
- Hauk, L. Acne Vulgaris: Treatment Guidelines from the AAD. Am. Fam. Physician 2017, 95, 740–741. [Google Scholar]
- Reynolds, R.V.; Yeung, H.; Cheng, C.E.; Cook-Bolden, F.; Desai, S.R.; Druby, K.M.; Freeman, E.E.; Keri, J.E.; Stein Gold, L.F.; Tan, J.K.L.; et al. Guidelines of care for the management of acne vulgaris. J. Am. Acad. Dermatol. 2024, 90, 1006.e1–1006.e30. [Google Scholar]
- Totte, J.E.; van der Feltz, W.T.; Bode, L.G.; van Belkum, A.; van Zuuren, E.J.; Pasmans, S.G. A systematic review and meta-analysis on Staphylococcus aureus carriage in psoriasis, acne and rosacea. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1069–1077. [Google Scholar]
- Mayslich, C.; Grange, P.A.; Dupin, N. Cutibacterium acnes as an Opportunistic Pathogen: An Update of Its Virulence-Associated Factors. Microorganisms 2021, 9, 303. [Google Scholar] [CrossRef]
- Gajdacs, M.; Spengler, G.; Urban, E. Identification and Antimicrobial Susceptibility Testing of Anaerobic Bacteria: Rubik’s Cube of Clinical Microbiology? Antibiotics 2017, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Coates, P.; Vyakrnam, S.; Eady, E.A.; Jones, C.E.; Cove, J.H.; Cunliffe, W.J. Prevalence of antibiotic-resistant propionibacteria on the skin of acne patients: 10-year surveillance data and snapshot distribution study. Br. J. Dermatol. 2002, 146, 840–848. [Google Scholar] [PubMed]
- Dessinioti, C.; Katsambas, A. Propionibacterium acnes and antimicrobial resistance in acne. Clin. Dermatol. 2017, 35, 163–167. [Google Scholar]
- Dessinioti, C.; Katsambas, A. Antibiotics and Antimicrobial Resistance in Acne: Epidemiological Trends and Clinical Practice Considerations. Yale J. Biol. Med. 2022, 95, 429–443. [Google Scholar] [PubMed]
- Beig, M.; Shirazi, O.; Ebrahimi, E.; Banadkouki, A.Z.; Golab, N.; Sholeh, M. Prevalence of antibiotic-resistant Cutibacterium acnes (formerly Propionibacterium acnes) isolates, a systematic review and meta-analysis. J. Global Antimicrob. Resist. 2024, 39, 82–91. [Google Scholar]
- Fischetti, V.A. Bacteriophage lysins as effective antibacterials. Curr. Opin. Microbiol. 2008, 11, 393–400. [Google Scholar]
- Ghose, C.; Euler, C.W. Gram-Negative Bacterial Lysins. Antibiotics 2020, 9, 74. [Google Scholar] [CrossRef]
- Thandar, M.; Lood, R.; Winer, B.Y.; Deutsch, D.R.; Euler, C.W.; Fischetti, V.A. Novel Engineered Peptides of a Phage Lysin as Effective Antimicrobials against Multidrug-Resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 2671–2679. [Google Scholar]
- Heselpoth, R.D.; Euler, C.W.; Fischetti, V.A. PaP1, a Broad-Spectrum Lysin-Derived Cationic Peptide to Treat Polymicrobial Skin Infections. Front. Microbiol. 2022, 13, 817228. [Google Scholar]
- Rahman, M.; Wang, W.; Sun, O.; Ali Shah, J.; Li, C.; Sun, Y.; Li, Y.; Zhang, B.; Chen, W.; Wang, S. Endolysin, a Promising Solution against Antimicrobial Resistance. Antibiotics 2021, 10, 1277. [Google Scholar] [CrossRef]
- Lood, R.; Winer, B.Y.; Pelzek, A.J.; Diez-Martinez, R.; Thandar, M.; Euler, C.W.; Schuch, R.; Fischetti, V.A. Novel phage lysin capable of killing the multidrug-resistant gram-negative bacterium Acinetobacter baumannii in a mouse bacteremia model. Antimicrob. Agents Chemother. 2015, 59, 1983–1991. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Asif, F.; Zaman, S.U.; Arnab, M.K.H.; Rahman, M.M.; Hasan, M. Effect of charge on the antimicrobial activity of alpha-helical amphibian antimicrobial peptide. Curr. Res. Microb. Sci. 2023, 4, 100182. [Google Scholar] [PubMed]
- Douglas, H.C.; Gunter, S.E. The taxonomic position of Corynebacterium acnes. J. Bacteriol. 1946, 52, 15–23. [Google Scholar]
- Bruggemann, H.; Henne, A.; Hoster, F.; Liesegang, H.; Wiezer, A.; Strittmatter, A.; Hujer, S.; Durre, P.; Gottschalk, G. The complete genome sequence of Propionibacterium acnes, a commensal of human skin. Science 2004, 305, 671–673. [Google Scholar] [CrossRef]
- Novick, R. Properties of a cryptic high-frequency transducing phage in Staphylococcus aureus. Virology 1967, 33, 155–166. [Google Scholar] [CrossRef]
- McDougal, L.K.; Steward, C.D.; Killgore, G.E.; Chaitram, J.M.; McAllister, S.K.; Tenover, F.C. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus aureus isolates from the United States: Establishing a national database. J. Clin. Microbiol. 2003, 41, 5113–5120. [Google Scholar] [CrossRef]
- Voyich, J.M.; Otto, M.; Mathema, B.; Braughton, K.R.; Whitney, A.R.; Welty, D.; Long, R.D.; Dorward, D.W.; Gardner, D.J.; Lina, G.; et al. Is Panton-Valentine leukocidin the major virulence determinant in community-associated methicillin-resistant Staphylococcus aureus disease? J. Infect. Dis. 2006, 194, 1761–1770. [Google Scholar] [CrossRef]
- Nakamura, K.; Williams, M.R.; Kwiecinski, J.M.; Horswill, A.R.; Gallo, R.L. Staphylococcus aureus Enters Hair Follicles Using Triacylglycerol Lipases Preserved through the Genus Staphylococcus. J. Invest. Dermatol. 2021, 141, 2094–2097. [Google Scholar]
- Schlecht, S.; Freudenberg, M.A.; Galanos, C. Culture and biological activity of Propionibacterium acnes. Infection 1997, 25, 247–249. [Google Scholar] [CrossRef]
- Korting, H.C.; Lukacs, A.; Vogt, N.; Urban, J.; Ehret, W.; Ruckdeschel, G. Influence of the pH-value on the growth of Staphylococcus epidermidis, Staphylococcus aureus and Propionibacterium acnes in continuous culture. Zentralbl. Hyg. Umweltmed. 1992, 193, 78–90. [Google Scholar]
- Kaden, D.; Dahne, L.; Knorr, F.; Richter, H.; Lademann, J.; Meinke, M.C.; Patzelt, A.; Darvin, M.E.; Jung, S. Determination of the pH Gradient in Hair Follicles of Human Volunteers Using pH-Sensitive Melamine Formaldehyde-Pyranine Nile Blue Microparticles. Sensors 2020, 20, 5243. [Google Scholar] [CrossRef] [PubMed]
- Baker, L.B. Physiology of sweat gland function: The roles of sweating and sweat composition in human health. Temperature 2019, 6, 211–259. [Google Scholar] [CrossRef] [PubMed]
- Ciornei, C.D.; Tapper, H.; Bjartell, A.; Sternby, N.H.; Bodelsson, M. Human antimicrobial peptide LL-37 is present in atherosclerotic plaques and induces death of vascular smooth muscle cells: A laboratory study. BMC Cardiovasc. Disord. 2006, 6, 49. [Google Scholar] [CrossRef] [PubMed]
- Zaenglein, A.L.; Pathy, A.L.; Schlosser, B.J.; Alikhan, A.; Baldwin, H.E.; Berson, D.S.; Bowe, W.P.; Graber, E.M.; Harper, J.C.; Kang, S.; et al. Guidelines of care for the management of acne vulgaris. J. Am. Acad. Dermatol. 2016, 74, 945–973.e33. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Yan, Z.B.; Meng, Y.M.; Hong, X.Y.; Shao, G.; Ma, J.J.; Cheng, X.R.; Liu, J.; Kang, J.; Fu, C.Y. Antimicrobial peptides: Mechanism of action, activity and clinical potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef]
- Bechinger, B.; Gorr, S.U. Antimicrobial Peptides: Mechanisms of Action and Resistance. J. Dent. Res. 2017, 96, 254–260. [Google Scholar] [CrossRef]
- Zhang, L.; Dhillon, P.; Yan, H.; Farmer, S.; Hancock, R.E.W. Interactions of Bacterial Cationic Peptide Antibiotics with Outer and Cytoplasmic Membranes of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3317–3321. [Google Scholar] [CrossRef]
- McLaughlin, J.; Watterson, S.; Layton, A.M.; Bjourson, A.J.; Barnard, E.; McDowell, A. Propionibacterium acnes and Acne Vulgaris: New Insights from the Integration of Population Genetic, Multi-Omic, Biochemical and Host-Microbe Studies. Microorganisms 2019, 7, 128. [Google Scholar] [CrossRef]
- Mirshahpanah, P.; Maibach, H.I. Models in acnegenesis. Cutan. Ocul. Toxicol. 2007, 26, 195–202. [Google Scholar] [CrossRef]
- Prakash, C.; Bhargava, P.; Tiwari, S.; Majumdar, B.; Bhargava, R.K. Skin Surface pH in Acne Vulgaris: Insights from an Observational Study and Review of the Literature. J. Clin. Aesthet. Dermatol. 2017, 10, 33–39. [Google Scholar]
- Lim, L.M.; Ly, N.; Anderson, D.; Yang, J.C.; Macander, L.; Jarkowski, A.; Forrest, A.; Bulitta, J.B.; Tsuji, B.T. Resurgence of Colistin: A Review of Resistance, Toxicity, Pharmacodynamics, and Dosing. Pharmacotherapy 2010, 30, 1279–1291. [Google Scholar]
- Waaterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modeling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Amino Acid Sequence | pI | MW (kDa) | Net Charge | GRAVY |
|---|---|---|---|---|---|
| P156 | RRKAKAPRAEIYAQFNKWVYAGGKKLSGLVKRRRR | 11.90 | 4.15 | +11.1 | −1.149 |
| Species | Strain | Source | Notes | References |
|---|---|---|---|---|
| C. acnes | ATCC 6919 | ATCC | Type IA; source: facial acne | [23] |
| C. acnes | HSS B | HSS | Source: 2015 clinical isolate | This study |
| C. acnes | HSS C | HSS | Source: 2015 clinical isolate | This study |
| C. acnes | HSS D | HSS | Source: 2015 clinical isolate | This study |
| C. acnes | HSS E | HSS | Source: 2015 clinical isolate | This study |
| C. acnes | HSS F | HSS | Source: 2015 clinical isolate | This study |
| C. acnes | KPA171202 | RUBC | Type IB; source: skin | [24] |
| S. aureus | 8325 | RUBC | MSSA; source: blood | [25] |
| S. aureus | NRS384 | NARSA | USA300 pulsotype, MRSA; source: wound | [26] |
| S. aureus | MW2 BAA-1707 | ATCC | USA400 pulsotype, MRSA; source: blood | [27] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sela, U.; Heselpoth, R.D.; Fischetti, V.A. Engineered Lysin-Derived Peptide as a Potent Antimicrobial for Acne Vulgaris. Antibiotics 2025, 14, 344. https://doi.org/10.3390/antibiotics14040344
Sela U, Heselpoth RD, Fischetti VA. Engineered Lysin-Derived Peptide as a Potent Antimicrobial for Acne Vulgaris. Antibiotics. 2025; 14(4):344. https://doi.org/10.3390/antibiotics14040344
Chicago/Turabian StyleSela, Uri, Ryan D. Heselpoth, and Vincent A. Fischetti. 2025. "Engineered Lysin-Derived Peptide as a Potent Antimicrobial for Acne Vulgaris" Antibiotics 14, no. 4: 344. https://doi.org/10.3390/antibiotics14040344
APA StyleSela, U., Heselpoth, R. D., & Fischetti, V. A. (2025). Engineered Lysin-Derived Peptide as a Potent Antimicrobial for Acne Vulgaris. Antibiotics, 14(4), 344. https://doi.org/10.3390/antibiotics14040344