
Cinnamic Acid Compounds (p-Coumaric, Ferulic, and p-Methoxycinnamic Acid) as Effective Antibacterial Agents Against Colistin-Resistant Acinetobacter baumannii


Abstract
1. Introduction
2. Results
2.1. Antibacterial Susceptibility
2.2. Evaluation of the Checkerboard Synergy Test Result
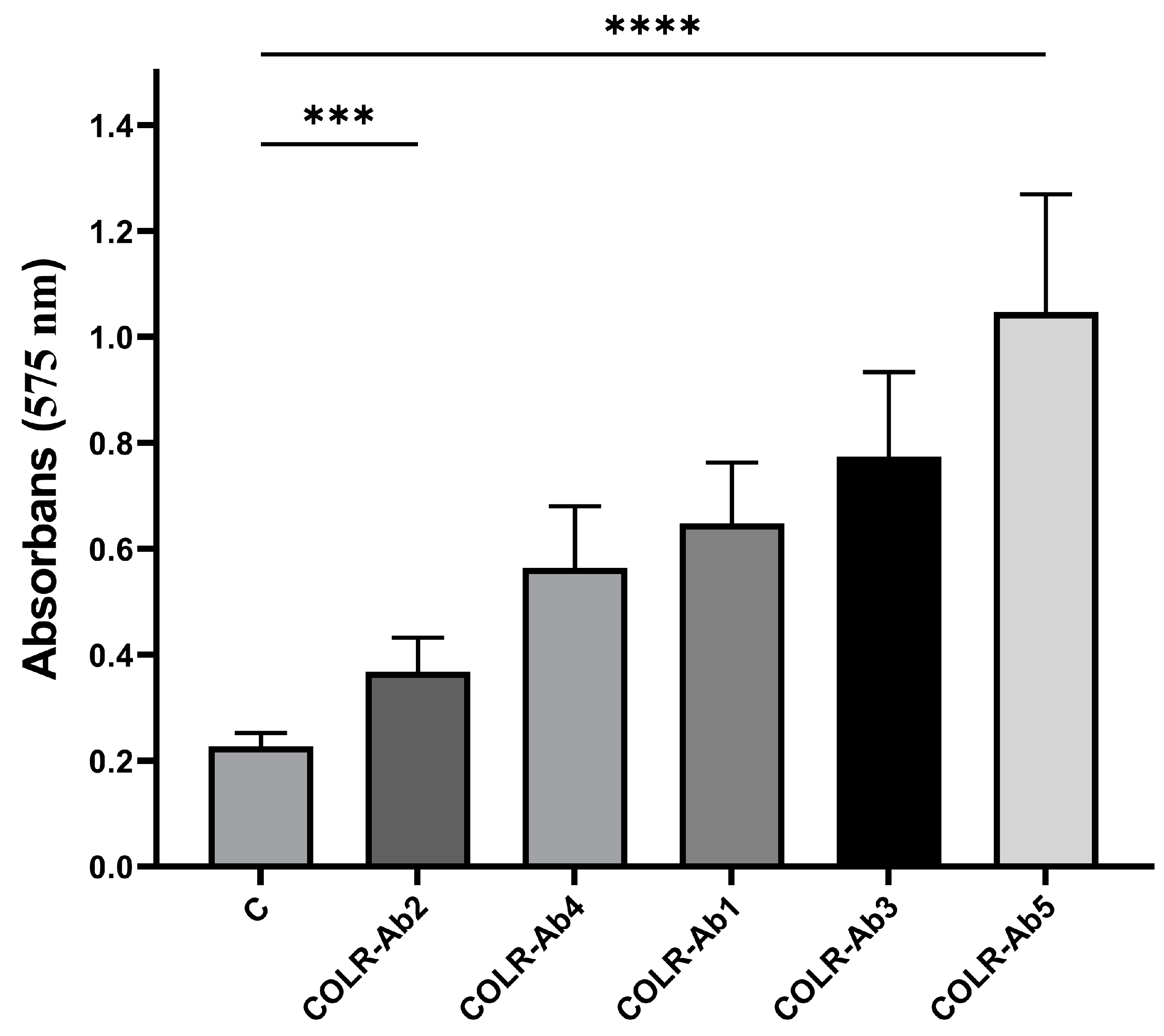
2.3. Detection of Biofilm Production
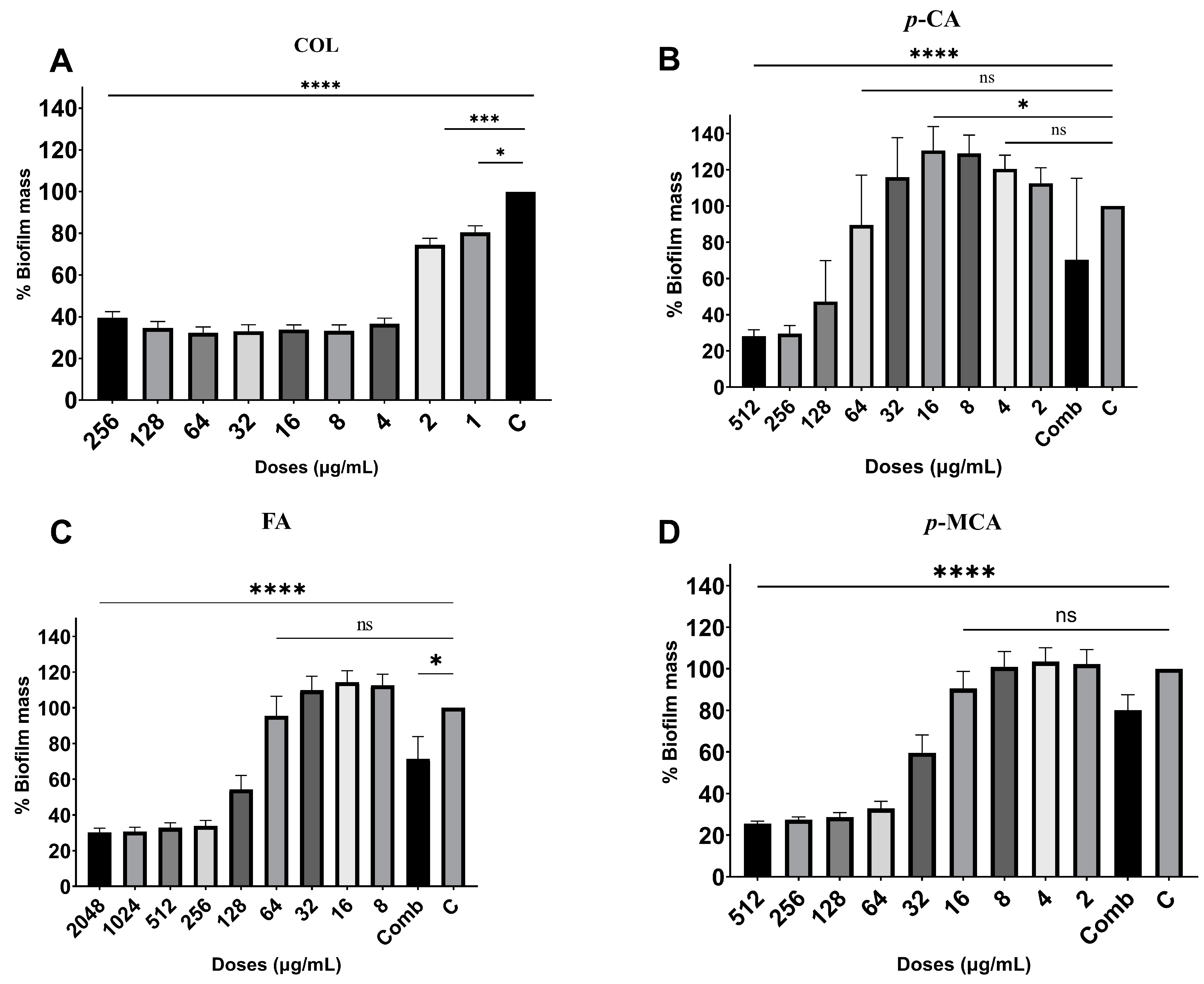
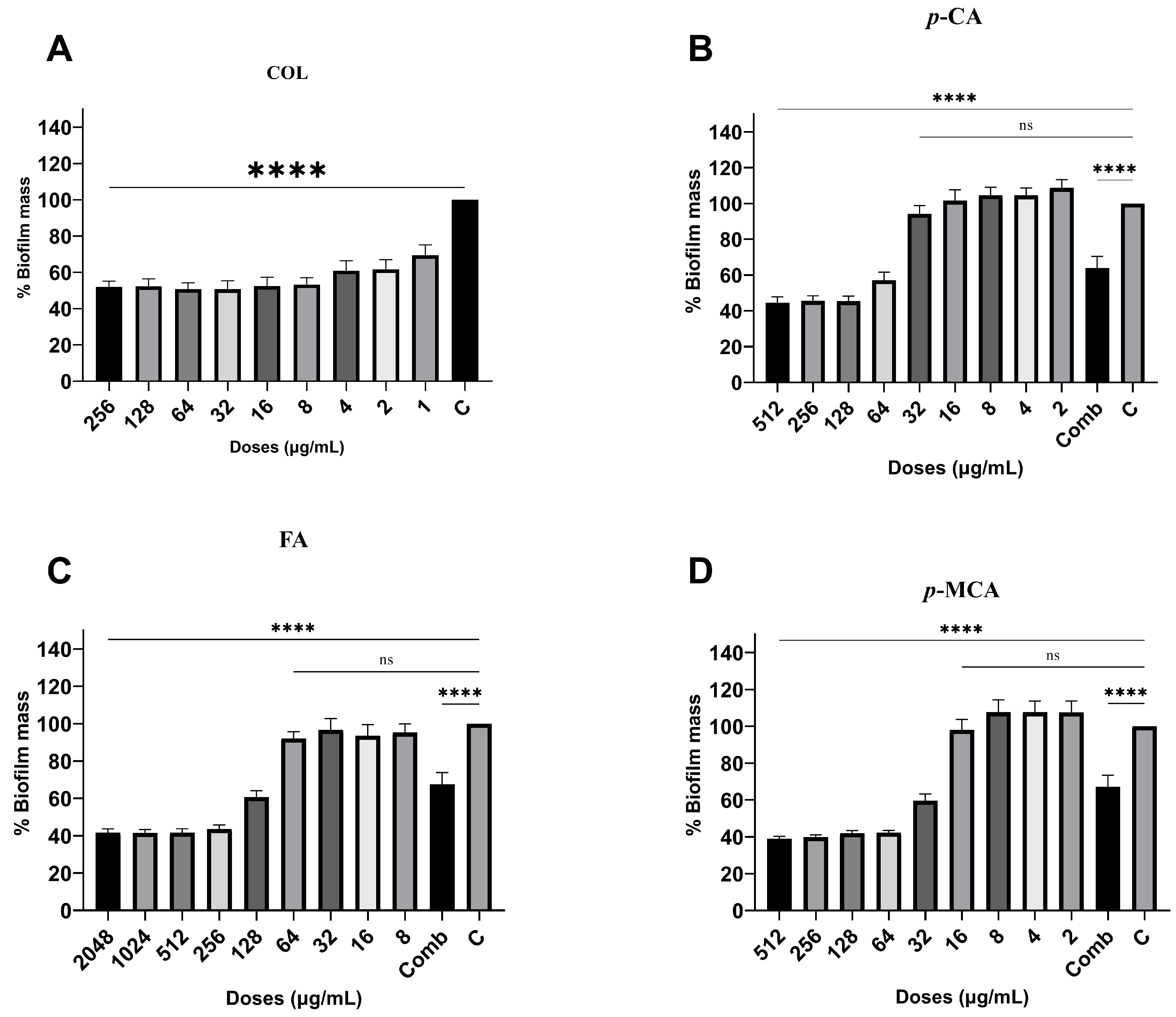
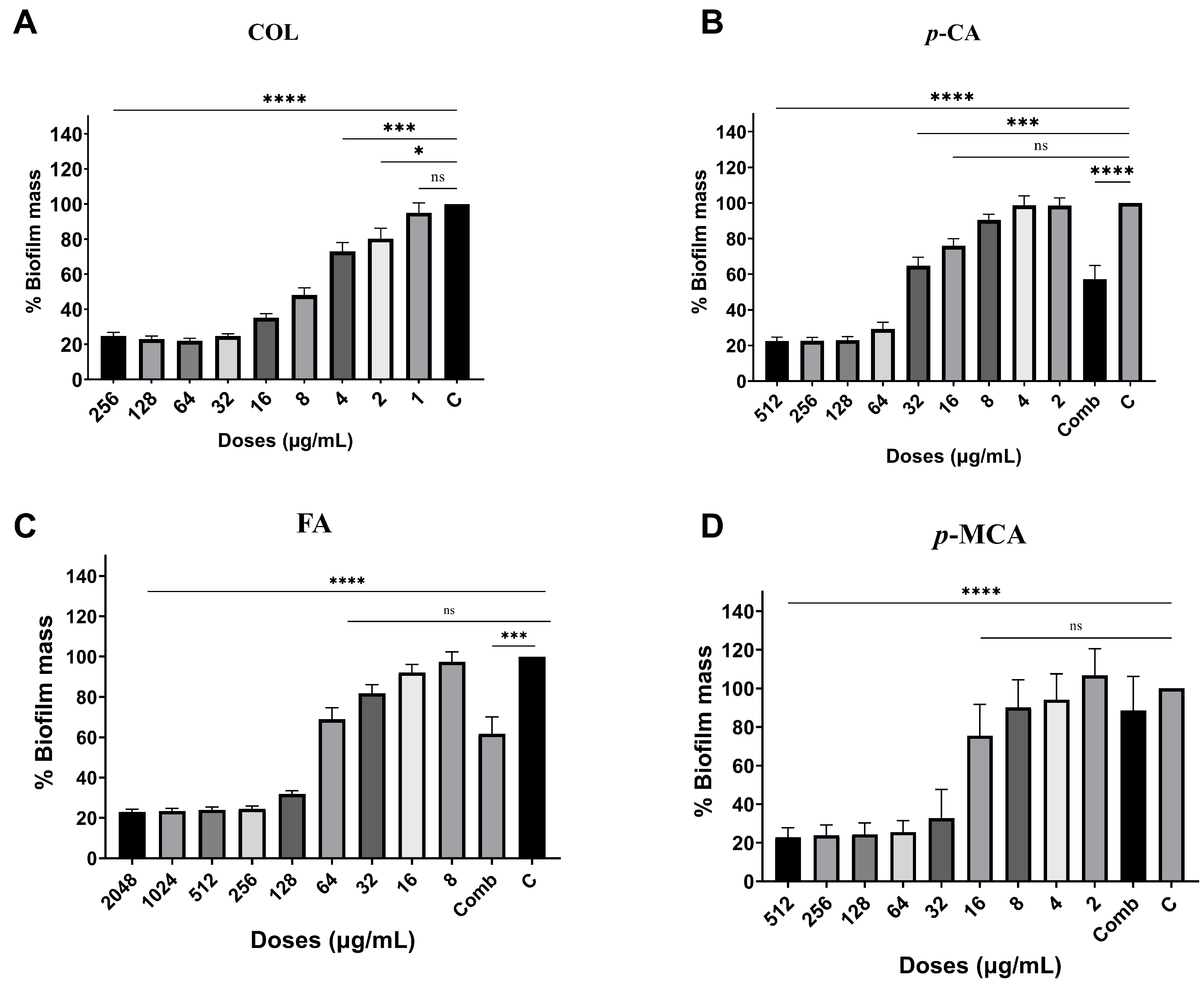
2.4. Antibiofilm Activity
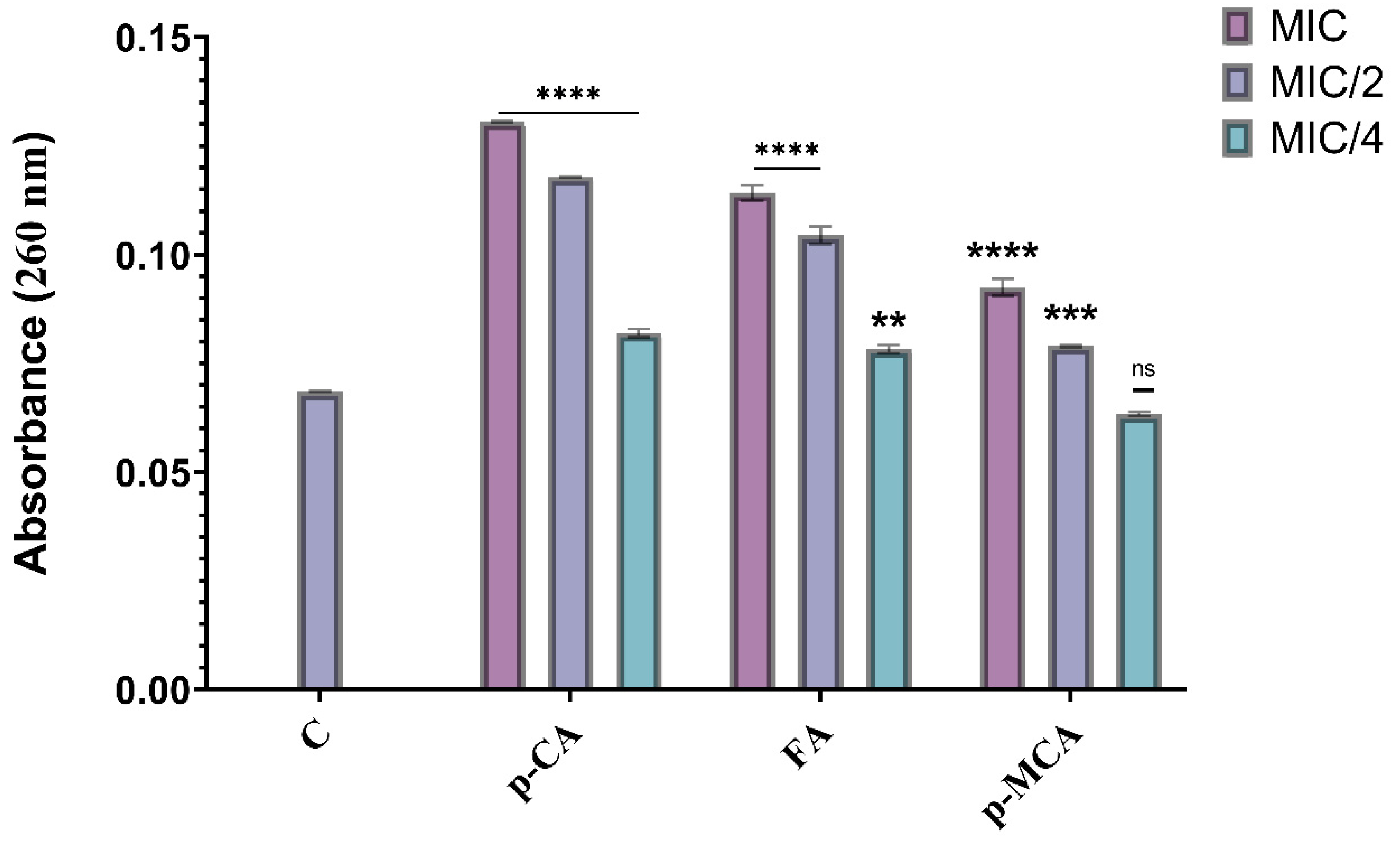
2.5. Measuring Cell Membrane Damage
2.6. Scanning Electron Microscopy
2.7. Results of Cytotoxic Activity
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Test Compounds
4.2. MIC Determination
4.3. Checkerboard Synergy Test
4.4. Detection of Biofilm Production
4.5. Antibiofilm Activity
4.6. Cytoplasmic Membrane Permeability Assay
4.7. Scanning Electron Microscopy
4.8. Cell Culture
4.9. Cell Viability Assay
4.10. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McMichael, A.J. Environmental and Social Influences on Emerging Infectious Diseases: Past, Present and Future. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2004, 359, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular Mechanisms of Antibiotic Resistance. Nat. Rev. Microbiol. 2014, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Darby, E.M.; Trampari, E.; Siasat, P.; Gaya, M.S.; Alav, I.; Webber, M.A.; Blair, J.M.A. Molecular Mechanisms of Antibiotic Resistance Revisited. Nat. Rev. Microbiol. 2023, 21, 280–295. [Google Scholar] [CrossRef] [PubMed]
- Baym, M.; Stone, L.K.; Kishony, R. Multidrug Evolutionary Strategies to Reverse Antibiotic Resistance. Science 2016, 351, aad3292. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 1317–1325. [Google Scholar] [CrossRef]
- Asif, M.; Alvi, I.A.; Rehman, S.U. Insight into Acinetobacter baumannii: Pathogenesis, Global Resistance, Mechanisms of Resistance, Treatment Options, and Alternative Modalities. Infect. Drug Resist. 2018, 11, 1249–1260. [Google Scholar] [CrossRef]
- AlSheikh, H.M.A.; Sultan, I.; Kumar, V.; Rather, I.A.; Al-Sheikh, H.; Tasleem Jan, A.; Haq, Q.M.R. Plant-Based Phytochemicals as Possible Alternative to Antibiotics in Combating Bacterial Drug Resistance. Antibiotics 2020, 9, 480. [Google Scholar] [CrossRef]
- Eliopoulos, G.M.; Maragakis, L.L.; Perl, T.M. Antimicrobial Resistance: Acinetobacter baumannii: Epidemiology, Antimicrobial Resistance, and Treatment Options. Clin. Infect. Dis. 2008, 46, 1254–1263. [Google Scholar] [CrossRef]
- Hazhirkamal, M.; Zarei, O.; Movahedi, M.; Karami, P.; Shokoohizadeh, L.; Taheri, M. Molecular Typing, Biofilm Production, and Detection of Carbapenemase Genes in Multidrug-Resistant Acinetobacter baumannii Isolated from Different Infection Sites Using ERIC-PCR in Hamadan, West of Iran. BMC Pharmacol. Toxicol. 2021, 22, 32. [Google Scholar] [CrossRef]
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and Pathophysiological Overview of Acinetobacter Infections: A Century of Challenges. Clin. Microbiol. Rev. 2017, 30, 409–447. [Google Scholar] [CrossRef]
- Pakharukova, N.; Tuittila, M.; Paavilainen, S.; Malmi, H.; Parilova, O.; Teneberg, S.; Knight, S.D.; Zavialov, A.V. Structural Basis for Acinetobacter baumannii Biofilm Formation. Proc. Natl. Acad. Sci. USA 2018, 115, 5558–5563. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Li, H.; Zhang, C.; Liang, B.; Li, J.; Wang, L.; Du, X.; Liu, X.; Qiu, S.; Song, H. Relationship between Antibiotic Resistance, Biofilm Formation, and Biofilm-Specific Resistance in Acinetobacter baumannii. Front. Microbiol. 2016, 7, 483. [Google Scholar] [CrossRef] [PubMed]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter baumannii Antibiotic Resistance Mechanisms. Pathogens 2021, 10, 373. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, G.J.; Domingues, S. Interplay between Colistin Resistance, Virulence and Fitness in Acinetobacter baumannii. Antibiotics 2017, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, S.S.; Dwibedy, S.K.; Padhy, I. Polymyxins, the Last-Resort Antibiotics: Mode of Action, Resistance Emergence, and Potential Solutions. J. Biosci. 2021, 46, 85. [Google Scholar] [CrossRef]
- Cai, Y.; Chai, D.; Wang, R.; Liang, B.; Bai, N. Colistin Resistance of Acinetobacter baumannii: Clinical Reports, Mechanisms and Antimicrobial Strategies. J. Antimicrob. Chemother. 2012, 67, 1607–1615. [Google Scholar] [CrossRef]
- Li, J.; Nation, R.L.; Turnidge, J.D.; Milne, R.W.; Coulthard, K.; Rayner, C.R.; Paterson, D.L. Colistin: The Re-Emerging Antibiotic for Multidrug-Resistant Gram-Negative Bacterial Infections. Lancet Infect. Dis. 2006, 6, 589–601. [Google Scholar] [CrossRef]
- Aghapour, Z.; Gholizadeh, P.; Ganbarov, K.; Bialvaei, A.Z.; Mahmood, S.S.; Tanomand, A.; Yousefi, M.; Asgharzadeh, M.; Yousefi, B.; Kafil, H.S. Molecular Mechanisms Related to Colistin Resistance in Enterobacteriaceae. Infect. Drug Resist. 2019, 12, 965–975. [Google Scholar] [CrossRef]
- Bialvaei, A.Z.; Samadi Kafil, H. Colistin, Mechanisms and Prevalence of Resistance. Curr. Med. Res. Opin. 2015, 31, 707–721. [Google Scholar] [CrossRef]
- Allen, H.K.; Trachsel, J.; Looft, T.; Casey, T.A. Finding Alternatives to Antibiotics. Ann. N. Y Acad. Sci. 2014, 1323, 91–100. [Google Scholar] [CrossRef]
- Kumar, M.; Sarma, D.K.; Shubham, S.; Kumawat, M.; Verma, V.; Nina, P.B.; Jp, D.; Kumar, S.; Singh, B.; Tiwari, R.R. Futuristic Non-Antibiotic Therapies to Combat Antibiotic Resistance: A Review. Front. Microbiol. 2021, 12, 16. [Google Scholar] [CrossRef] [PubMed]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive Review of Antimicrobial Activities of Plant Flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef]
- Savoia, D. Plant-Derived Antimicrobial Compounds: Alternatives to Antibiotics. Future Microbiol. 2012, 7, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Allemailem, K.S. Antimicrobial Potential of Naturally Occurring Bioactive Secondary Metabolites. J. Pharm. Bioallied Sci. 2021, 13, 155. [Google Scholar] [CrossRef]
- Ruwizhi, N.; Aderibigbe, B.A. Cinnamic Acid Derivatives and Their Biological Efficacy. Int. J. Mol. Sci. 2020, 21, 5712. [Google Scholar] [CrossRef]
- Sova, M. Antioxidant and Antimicrobial Activities of Cinnamic Acid Derivatives. Mini-Rev. Med. Chem. 2012, 12, 749–767. [Google Scholar] [CrossRef]
- Ververidis, F.; Trantas, E.; Douglas, C.; Vollmer, G.; Kretzschmar, G.; Panopoulos, N. Biotechnology of Flavonoids and Other Phenylpropanoid-Derived Natural Products. Part I: Chemical Diversity, Impacts on Plant Biology and Human Health. Biotechnol. J. 2007, 2, 1214–1234. [Google Scholar] [CrossRef]
- Ibitoye, O.B.; Ajiboye, T.O. Ferulic Acid Potentiates the Antibacterial Activity of Quinolone-Based Antibiotics against Acinetobacter baumannii. Microb. Pathog. 2019, 126, 393–398. [Google Scholar] [CrossRef]
- Nakazono, Y.; Watanabe, Y.; Hashinaga, F.; Tadera, K. Studies on Antimicrobial and Antioxidative Substance of Yuzu (Citrus junos Hort. Ex Tanaka) Seed. J. Biol. Sci. 2005, 6, 135–139. [Google Scholar] [CrossRef]
- Narasimhan, B.; Belsare, D.; Pharande, D.; Mourya, V.; Dhake, A. Esters, Amides and Substituted Derivatives of Cinnamic Acid: Synthesis, Antimicrobial Activity and QSAR Investigations. Eur. J. Med. Chem. 2004, 39, 827–834. [Google Scholar] [CrossRef]
- Shi, C.; Zhang, X.; Sun, Y.; Yang, M.; Song, K.; Zheng, Z.; Chen, Y.; Liu, X.; Jia, Z.; Dong, R.; et al. Antimicrobial Activity of Ferulic Acid Against Cronobacter sakazakii and Possible Mechanism of Action. Foodborne Pathog. Dis. 2016, 13, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Cueva, C.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Bills, G.; Vicente, M.F.; Basilio, A.; Rivas, C.L.; Requena, T.; Rodríguez, J.M.; Bartolomé, B. Antimicrobial Activity of Phenolic Acids against Commensal, Probiotic and Pathogenic Bacteria. Res. Microbiol. 2010, 161, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Du, C.; Beaman, H.T.; Monroe, M.B.B. Characterization of Phenolic Acid Antimicrobial and Antioxidant Structure–Property Relationships. Pharmaceutics 2020, 12, 419. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Maldonado, A.F.; Schieber, A.; Gänzle, M.G. Structure-Function Relationships of the Antibacterial Activity of Phenolic Acids and Their Metabolism by Lactic Acid Bacteria. J. Appl. Microbiol. 2011, 111, 1176–1184. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Rao, S.; Sun, J.; Ma, C.; Li, J. p-Coumaric Acid Kills Bacteria through Dual Damage Mechanisms. Food Control 2012, 25, 550–554. [Google Scholar] [CrossRef]
- Borges, A.; Saavedra, M.J.; Simões, M. The Activity of Ferulic and Gallic Acids in Biofilm Prevention and Control of Pathogenic Bacteria. Biofouling 2012, 28, 755–767. [Google Scholar] [CrossRef]
- Kang, J.; Liu, L.; Liu, Y.; Wang, X. Ferulic Acid Inactivates Shigella flexneri through Cell Membrane Destruction, Biofilm Retardation, and Altered Gene Expression. J. Agric. Food Chem. 2020, 68, 7121–7131. [Google Scholar] [CrossRef]
- Choudhury, D.; Talukdar, A.; Chetia, P.; Bhattacharjee, A.; Choudhury, M. Screening of Natural Products and Derivatives for the Identification of RND Efflux Pump Inhibitors. Comb. Chem. High Throughput Screen. 2016, 19, 705–713. [Google Scholar] [CrossRef]
- Ojha, D.; Patil, K.N. p-Coumaric Acid Inhibits the Listeria monocytogenes RecA Protein Functions and SOS Response: An Antimicrobial Target. Biochem. Biophys. Res. Commun. 2019, 517, 655–661. [Google Scholar] [CrossRef]
- Myszka, K.; Schmidt, M.T.; Białas, W.; Olkowicz, M.; Leja, K.; Czaczyk, K. Role of Gallic and p-Coumaric Acids in the AHL-Dependent Expression of FlgA Gene and in the Process of Biofilm Formation in Food-Associated Pseudomonas fluorescens KM120. J. Sci. Food Agric. 2016, 96, 4037–4047. [Google Scholar] [CrossRef]
- Bodini, S.F.; Manfredini, S.; Epp, M.; Valentini, S.; Santori, F. Quorum Sensing Inhibition Activity of Garlic Extract and p-Coumaric Acid. Lett. Appl. Microbiol. 2009, 49, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, N.A.; Elkhatib, W.F.; Sherif, M.M.; Abourehab, M.A.S.; Al-Rashood, S.T.; Eldehna, W.M.; Mostafa, N.M.; Elleboudy, N.S. In Silico Docking, Resistance Modulation and Biofilm Gene Expression in Multidrug-Resistant Acinetobacter baumannii via Cinnamic and Gallic Acids. Antibiotics 2022, 11, 870. [Google Scholar] [CrossRef] [PubMed]
- Kheiry, M.; Dianat, M.; Badavi, M.; Mard, S.A.; Bayati, V. p-Coumaric Acid Attenuates Lipopolysaccharide-Induced Lung Inflammation in Rats by Scavenging ROS Production: An In Vivo and In Vitro Study. Inflammation 2019, 42, 1939–1950. [Google Scholar] [CrossRef] [PubMed]
- Ou, S.; Kwok, K. Ferulic Acid: Pharmaceutical Functions, Preparation and Applications in Foods. J. Sci. Food Agric. 2004, 84, 1261–1269. [Google Scholar] [CrossRef]
- Płowuszyńska, A.; Gliszczyńska, A. Recent Developments in Therapeutic and Nutraceutical Applications of p-Methoxycinnamic Acid from Plant Origin. Molecules 2021, 26, 3827. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Aggarwal, R.; Mahajan, P.; Pandiya, S.; Bajaj, A.; Verma, S.K.; Yadav, P.; Kharat, A.S.; Khan, A.U.; Dua, M.; Johri, A.K. Antibiotic Resistance: A Global Crisis, Problems and Solutions. Crit. Rev. Microbiol. 2024, 50, 896–921. [Google Scholar] [CrossRef]
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare 2023, 11, 1946. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Gorlenko, C.L.; Kiselev, H.Y.; Budanova, E.V.; Zamyatnin, A.A.; Ikryannikova, L.N. Plant Secondary Metabolites in the Battle of Drugs and Drug-Resistant Bacteria: New Heroes or Worse Clones of Antibiotics? Antibiotics 2020, 9, 170. [Google Scholar] [CrossRef]
- Mumtaz, L.; Farid, A.; Yousef Alomar, S.; Ahmad, N.; Nawaz, A.; Andleeb, S.; Amin, A. Assesment of Polyphenolic Compounds against Biofilms Produced by Clinical Acinetobacter baumannii Strains Using in Silico and in Vitro Models. Saudi J. Biol. Sci. 2023, 30, 103743. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial Activity and Mode of Action of Ferulic and Gallic Acids Against Pathogenic Bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.M.; Ly, N.; Anderson, D.; Yang, J.C.; Macander, L.; Jarkowski, A.; Forrest, A.; Bulitta, J.B.; Tsuji, B.T. Resurgence of Colistin: A Review of Resistance, Toxicity, Pharmacodynamics, and Dosing. Pharmacotherapy 2010, 30, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Ordooei Javan, A.; Shokouhi, S.; Sahraei, Z. A Review on Colistin Nephrotoxicity. Eur. J. Clin. Pharmacol. 2015, 71, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Rychlíčková, J.; Kubíčková, V.; Suk, P.; Urbánek, K. Challenges of Colistin Use in ICU and Therapeutic Drug Monitoring: A Literature Review. Antibiotics 2023, 12, 437. [Google Scholar] [CrossRef]
- Ajiboye, T.O.; Skiebe, E.; Wilharm, G. Phenolic Acids Potentiate Colistin-Mediated Killing of Acinetobacter baumannii by Inducing Redox Imbalance. Biomed. Pharmacother. 2018, 101, 737–744. [Google Scholar] [CrossRef]
- Odabaş Köse, E.; Koyuncu Özyurt, Ö.; Bilmen, S.; Er, H.; Kilit, C.; Aydemir, E. Quercetin: Synergistic Interaction with Antibiotics against Colistin-Resistant Acinetobacter baumannii. Antibiotics 2023, 12, 739. [Google Scholar] [CrossRef]
- Gaddy, J.A.; Actis, L.A. Regulation of Acinetobacter baumannii Biofilm Formation. Future Microbiol. 2009, 4, 273–278. [Google Scholar] [CrossRef]
- Gedefie, A.; Demsiss, W.; Belete, M.A.; Kassa, Y.; Tesfaye, M.; Tilahun, M.; Bisetegn, H.; Sahle, Z. Acinetobacter baumannii Biofilm Formation and Its Role in Disease Pathogenesis: A Review. Infect. Drug Resist. 2021, 14, 3711–3719. [Google Scholar] [CrossRef]
- Bala, M.; Gupte, S.; Aggarwal, P.; Kaur, M.; Manhas, A. Biofilm Producing Multidrug Resistant Acinetobacter Species from a Tertiary Care Hospital: A Therapeutic Challenge. Int. J. Res. Med. Sci. 2016, 4, 3024–3026. [Google Scholar] [CrossRef]
- Zeighami, H.; Valadkhani, F.; Shapouri, R.; Samadi, E.; Haghi, F. Virulence Characteristics of Multidrug Resistant Biofilm Forming Acinetobacter baumannii Isolated from Intensive Care Unit Patients. BMC Infect. Dis. 2019, 19, 629. [Google Scholar] [CrossRef]
- Eales, M.G.; Ferrari, E.; Goddard, A.D.; Lancaster, L.; Sanderson, P.; Miller, C. Mechanistic and Phenotypic Studies of Bicarinalin, BP100 and Colistin Action on Acinetobacter baumannii. Res. Microbiol. 2018, 169, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Song, J.Y.; Cheong, H.J.; Noh, J.Y.; Kim, W.J. In Vitro Comparison of Anti-Biofilm Effects against Carbapenem-Resistant Acinetobacter baumannii: Imipenem, Colistin, Tigecycline, Rifampicin and Combinations. Infect. Chemother. 2015, 47, 27. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.-J.; Zhou, J.-W.; Zhang, P.-P.; Luo, H.-Z.; Tang, S.; Li, J.-J.; Deng, S.-M.; Jia, A.-Q. Quorum Sensing Inhibition and Tobramycin Acceleration in Chromobacterium violaceum by Two Natural Cinnamic Acid Derivatives. Appl. Microbiol. Biotechnol. 2020, 104, 5025–5037. [Google Scholar] [CrossRef] [PubMed]
- Kot, B.; Wicha, J.; Piechota, M.; Wolska, K.; Gruzewska, A. Antibiofilm Activity of Trans-Cinnamaldehyde, p-Coumaric, and Ferulic Acids on Uropathogenic Escherichia coli. Turk. J. Med. Sci. 2015, 45, 919–924. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-G.; Hu, H.-X.; Chen, J.-Y.; Xue, Y.-S.; Kodirkhonov, B.; Han, B.-Z. Comparative Study on Inhibitory Effects of Ferulic Acid and p-Coumaric Acid on Salmonella Enteritidis Biofilm Formation. World J. Microbiol. Biotechnol. 2022, 38, 136. [Google Scholar] [CrossRef] [PubMed]
- Hemaiswarya, S.; Kruthiventi, A.K.; Doble, M. Synergism between Natural Products and Antibiotics against Infectious Diseases. Phytomedicine 2008, 15, 639–652. [Google Scholar] [CrossRef]
- Li, J.; Liu, W.; Jiao, W.; Lian, Y.; Mi, S.; Chitrakar, B.; Sang, Y.; Wang, X. Effect of Ferulic Acid and P-coumaric Acid on Lowering Uric Acid through Network Pharmacology and in Vitro Studies. Food Saf. Health 2024, 2, 133–144. [Google Scholar] [CrossRef]
- Saenglee, S.; Jogloy, S.; Patanothai, A.; Leid, M.; Senawong, T. Cytotoxic Effects of Peanut Phenolics Possessing Histone Deacetylase Inhibitory Activity in Breast and Cervical Cancer Cell Lines. Pharmacol. Rep. 2016, 68, 1102–1110. [Google Scholar] [CrossRef]
- Sevimli-Gur, C.; Yesil-Celiktas, O. Cytotoxicity Screening of Supercritical Fluid Extracted Seaweeds and Phenylpropanoids. Mol. Biol. Rep. 2019, 46, 3691–3699. [Google Scholar] [CrossRef]
- Posadino, A.M.; Cossu, A.; Giordo, R.; Zinellu, A.; Sotgia, S.; Vardeu, A.; Hoa, P.T.; Deiana, L.; Carru, C.; Pintus, G. Coumaric Acid Induces Mitochondrial Damage and Oxidative-Mediated Cell Death of Human Endothelial Cells. Cardiovasc. Toxicol. 2013, 13, 301–306. [Google Scholar] [CrossRef]
- Kong, C.; Jeong, C.; Choi, J.; Kim, K.; Jeong, J. Antiangiogenic Effects of p-Coumaric Acid in Human Endothelial Cells. Phytother. Res. 2013, 27, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Moody, J.; Knapp, C. Tests to Assess Bactericidal Activity. In Clinical Microbiology Procedures Handbook; Garcia, L., Ed.; ASM Press: Washington, DC, USA, 2010. [Google Scholar]
- Kırmusaoğlu, S. The Methods for Detection of Biofilm and Screening Antibiofilm Activity of Agents. In Antimicrobials, Antibiotic Resistance, Antibiofilm Strategies and Activity Methods; Kırmusaoğlu, S., Ed.; IntechOpen: London, UK, 2019; p. 152. ISBN 978-1-78985-790-0. [Google Scholar]
- Haney, E.F.; Trimble, M.J.; Hancock, R.E.W. Microtiter Plate Assays to Assess Antibiofilm Activity against Bacteria. Nat. Prot. 2021, 16, 2615–2632. [Google Scholar] [CrossRef] [PubMed]
- Bendali, F.; Gaillard-Martinie, B.; Hebraud, M.; Sadoun, D. Kinetic of Production and Mode of Action of the Lactobacillus paracasei subsp. paracasei Anti-Listerial Bacteriocin, an Algerian Isolate. LWT Food Sci. Technol. 2008, 41, 1784–1792. [Google Scholar] [CrossRef]
- Kavakcıoğlu Yardımcı, B.; Mollaoğlu, Z. Antioxidant or Pro-Oxidant? The Effects of Boron Compounds on Saccharomyces Cerevisiae BY4741 Strain. Prep. Biochem. Biotechnol. 2021, 51, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Kustiati, U.; Ergün, S.; Karnati, S.; Nugrahaningsih, D.A.A.; Kusindarta, D.L.; Wihadmadyatami, H. Ethanolic Extract of Ocimum sanctum Linn. Inhibits Cell Migration of Human Lung Adenocarcinoma Cells (A549) by Downregulation of Integrin αvβ3, α5β1, and VEGF. Sci. Pharm. 2022, 90, 69. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | MIC (µg/mL) | Mean FICI for COL Combined with: | |||||
|---|---|---|---|---|---|---|---|
| COL | p-CA | FA | p-MCA | p-CA | FA | p-MCA | |
| COLR-Ab1 | 32 | 256 | 1024 | 512 | 0.16 (S) | 0.15 (S) | 0.09 (S) |
| COLR-Ab2 | 64 | 256 | 1024 | 512 | 0.19 (S) | 0.15 (S) | 0.09 (S) |
| COLR-Ab3 | 64 | 256 | 1024 | 256 | 0.16 (S) | 0.15 (S) | 0.27 (S) |
| COLR-Ab4 | 128 | 256 | 512 | 256 | 0.15 (S) | 0.51 (I) | 0.14 (S) |
| COLR-Ab5 | 64 | 128 | 1024 | 128 | 0.28 (S) | 0.12 (S) | 0.27 (S) |
| E. coli | 4 | - | - | - | - | - | - |
| Isolates | MIC (µg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| AMK | CIP | GEN | IPM | MEM | NET | SXT | COL * | |
| COLR-Ab1 | ≤4 (S) | ≤0.125 (S) | ≤1 (S) | ≤0.25 (S) | ≤0.125 (S) | 1 (S) | ≤1/19 (S) | >4 (R) |
| COLR-Ab2 | >16 (R) | >2 (R) | >4 (R) | >8 (R) | >8 (R) | >4 (R) | >4/76 (R) | >4 (R) |
| COLR-Ab3 | >16 (R) | >2 (R) | >4 (R) | 4 (I) | >8 (R) | >4 (R) | >4/76 (R) | >4 (R) |
| COLR-Ab4 | >16 (R) | >2 (R) | >4 (R) | >8 (R) | >8 (R) | >4 (R) | >4/76 (R) | >4 (R) |
| COLR-Ab5 | >16 (R) | >2 (R) | >4 (R) | >8 (R) | >8 (R) | >4 (R) | >4/76 (R) | >4 (R) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korkut, A.; Özkaya Gül, S.; Aydemir, E.; Er, H.; Odabaş Köse, E. Cinnamic Acid Compounds (p-Coumaric, Ferulic, and p-Methoxycinnamic Acid) as Effective Antibacterial Agents Against Colistin-Resistant Acinetobacter baumannii. Antibiotics 2025, 14, 71. https://doi.org/10.3390/antibiotics14010071
Korkut A, Özkaya Gül S, Aydemir E, Er H, Odabaş Köse E. Cinnamic Acid Compounds (p-Coumaric, Ferulic, and p-Methoxycinnamic Acid) as Effective Antibacterial Agents Against Colistin-Resistant Acinetobacter baumannii. Antibiotics. 2025; 14(1):71. https://doi.org/10.3390/antibiotics14010071
Chicago/Turabian StyleKorkut, Alaaddin, Serap Özkaya Gül, Esra Aydemir, Hakan Er, and Elif Odabaş Köse. 2025. "Cinnamic Acid Compounds (p-Coumaric, Ferulic, and p-Methoxycinnamic Acid) as Effective Antibacterial Agents Against Colistin-Resistant Acinetobacter baumannii" Antibiotics 14, no. 1: 71. https://doi.org/10.3390/antibiotics14010071
APA StyleKorkut, A., Özkaya Gül, S., Aydemir, E., Er, H., & Odabaş Köse, E. (2025). Cinnamic Acid Compounds (p-Coumaric, Ferulic, and p-Methoxycinnamic Acid) as Effective Antibacterial Agents Against Colistin-Resistant Acinetobacter baumannii. Antibiotics, 14(1), 71. https://doi.org/10.3390/antibiotics14010071