
Culex quinquefasciatus Mosquitoes Resist Acquisition of Methicillin-Resistant Staphylococcus aureus: Insights from Field and Laboratory Studies
 and
and
Abstract
1. Introduction
2. Results
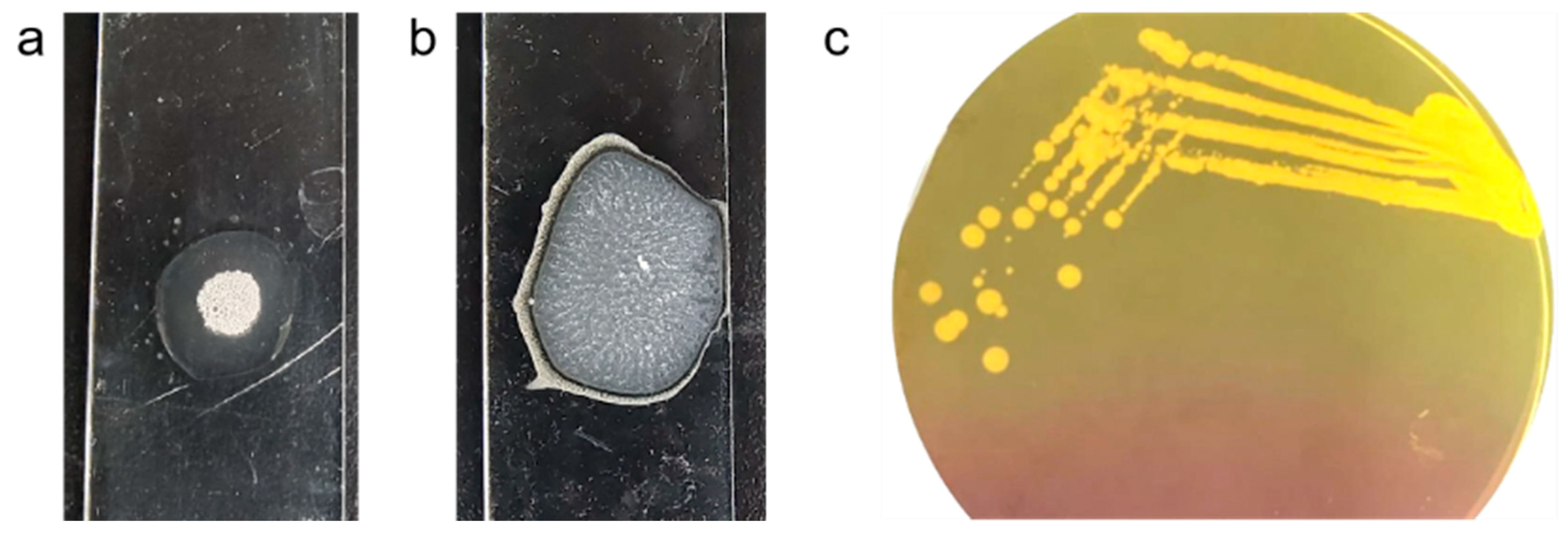
2.1. Phenotypic and Molecular Characterization of MRSA
2.2. The Inability of Mosquitoes to Acquire MRSA
2.3. Absence of MRSA in Field-Caught Mosquito Immatures
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. C. quinquefasciatus Colony
5.2. MRSA Culture
5.3. Phenotypic Characterization of MRSA
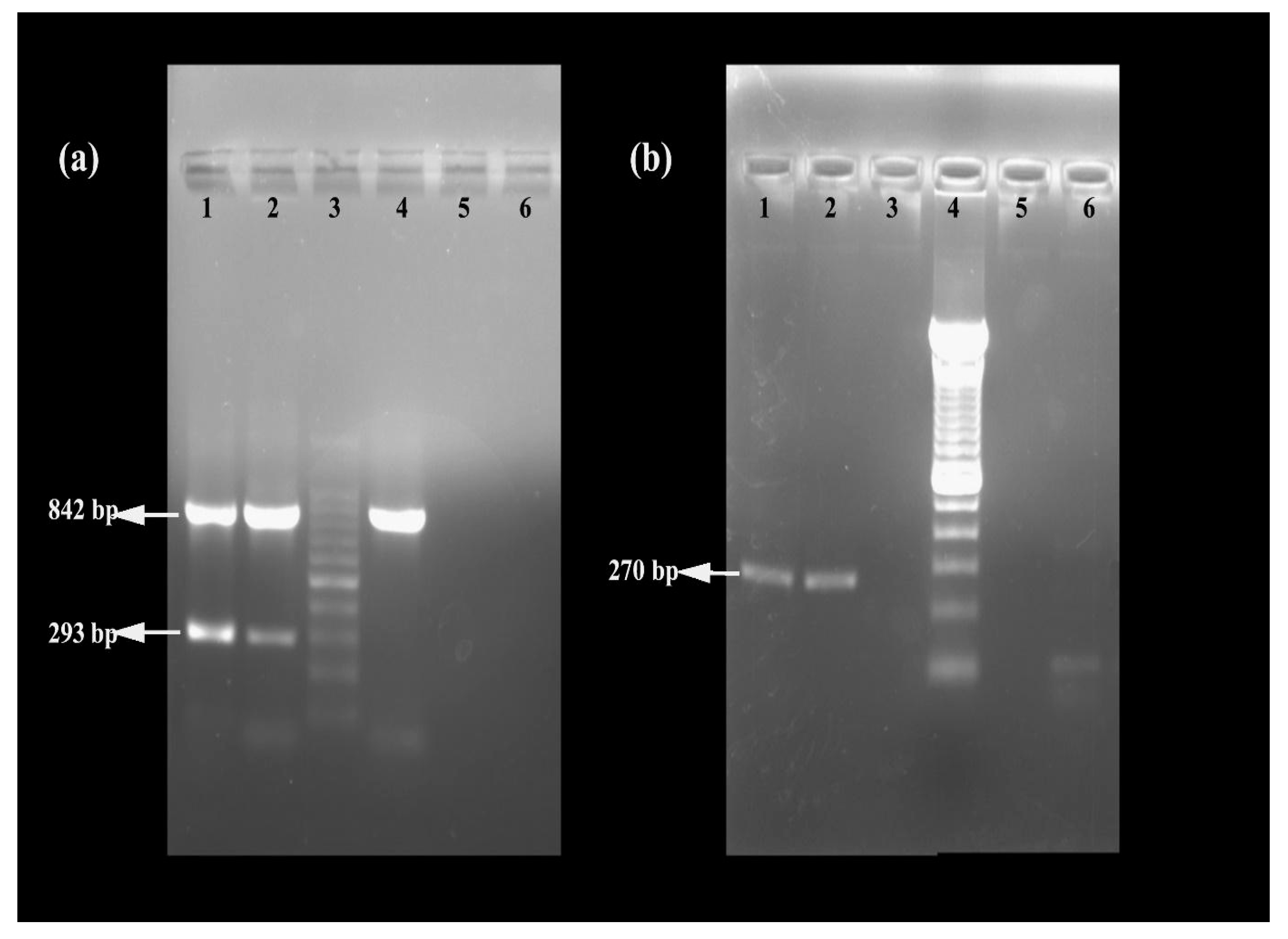
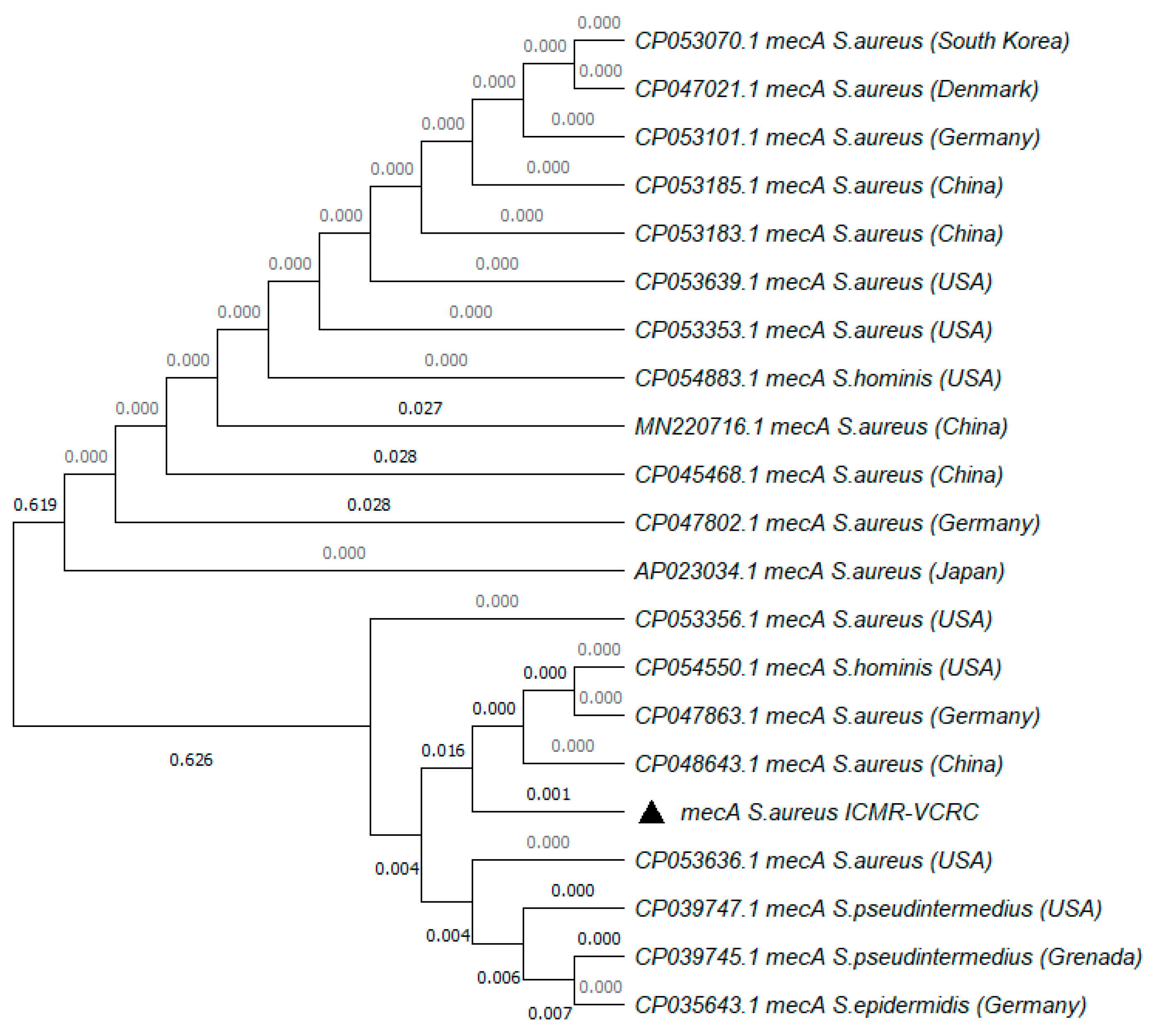
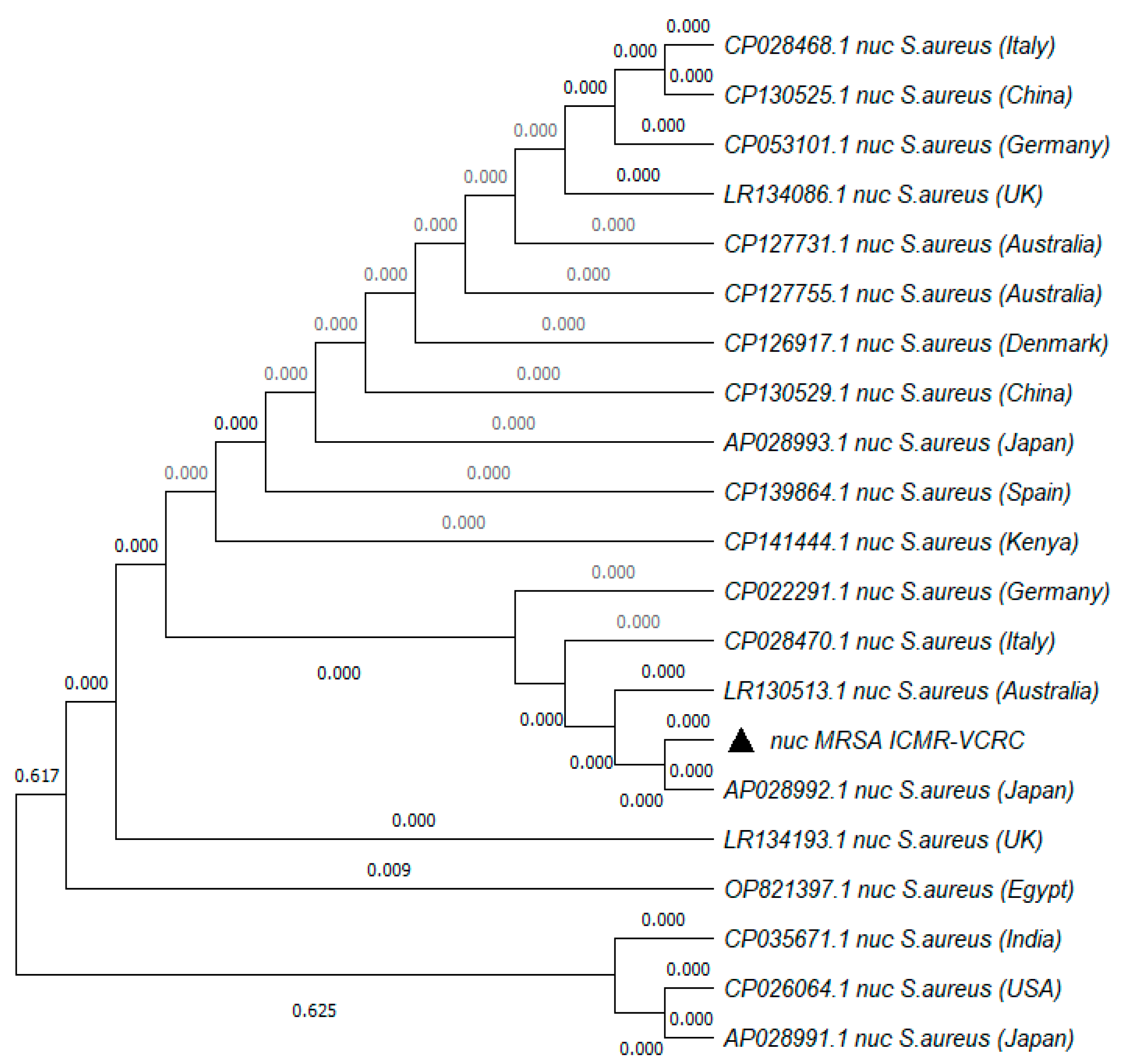
5.4. Molecular Characterization of MRSA
5.5. Dilution Plating
5.6. Experimental Exposure of MRSA to Larvae
5.7. Experimental Feeding of MRSA to Adults
5.8. Field Collection of Mosquito Immatures
5.9. Screening for MRSA
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Kraker, M.E.A.; Stewardson, A.J.; Harbarth, S. Will 10 Million People Die a Year Due to Antimicrobial Resistance by 2050? PLoS Med. 2016, 13, e1002184. [Google Scholar] [CrossRef] [PubMed]
- Larsson, D.G.J.; Flach, C.-F. Antibiotic Resistance in the Environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef]
- Gwenzi, W.; Chaukura, N.; Muisa-Zikali, N.; Teta, C.; Musvuugwa, T.; Rzymski, P.; Abia, A.L.K. Insects, Rodents, and Pets as Reservoirs, Vectors, and Sentinels of Antimicrobial Resistance. Antibiotics 2021, 10, 68. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, W.E.; Yang, Q. Clinical Perspective of Antimicrobial Resistance in Bacteria. Infect. Drug Resist. 2022, 15, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Gandra, S.; Ashok, A.; Caudron, Q.; Grenfell, B.T.; Levin, S.A.; Laxminarayan, R. Global Antibiotic Consumption 2000 to 2010: An Analysis of National Pharmaceutical Sales Data. Lancet Infect. Dis. 2014, 14, 742–750. [Google Scholar] [CrossRef]
- AMR Surveillance Network; Indian Council of Medical Research. Annual Report: 2021; Indian Council of Medical Research: New Delhi, India, 2021. [Google Scholar]
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare 2023, 11, 1946. [Google Scholar] [CrossRef]
- Foster, T. Medical Microbiology, 4th ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-Resistant Staphylococcus Aureus: An Overview of Basic and Clinical Research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Nandhini, P.; Kumar, P.; Mickymaray, S.; Alothaim, A.S.; Somasundaram, J.; Rajan, M. Recent Developments in Methicillin-Resistant Staphylococcus Aureus (MRSA) Treatment: A Review. Antibiotics 2022, 11, 606. [Google Scholar] [CrossRef] [PubMed]
- Kwiecinski, J.M.; Horswill, A.R. Staphylococcus Aureus Bloodstream Infections: Pathogenesis and Regulatory Mechanisms. Curr. Opin. Microbiol. 2020, 53, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Pramodhini, S.; Stephen, S.; Shanmugam, J.; Renuka, K. Prevalence and Antibiogram of MRSA Isolated from Patient Samples and Health Care Workers in a Tertiary Care Hospital, Puducherry, India. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1419–1424. [Google Scholar] [CrossRef]
- Jaradat, Z.W.; Ababneh, Q.O.; Sha’aban, S.T.; Alkofahi, A.A.; Assaleh, D.; Al Shara, A. Methicillin Resistant Staphylococcus Aureus and Public Fomites: A Review. Pathog. Glob. Health 2020, 114, 426–450. [Google Scholar] [CrossRef]
- Van Balen, J.; Mowery, J.; Piraino-Sandoval, M.; Nava-Hoet, R.C.; Kohn, C.; Hoet, A.E. Molecular Epidemiology of Environmental MRSA at an Equine Teaching Hospital: Introduction, Circulation and Maintenance. Vet. Res. 2014, 45, 31. [Google Scholar] [CrossRef]
- Yu, C.-H.; Shen, S.; Huang, K.-Y.A.; Huang, Y.-C. The Trend of Environmental and Clinical Methicillin-Resistant Staphylococcus Aureus in a Hospital in Taiwan: Impact of USA300. J. Microbiol. Immunol. Infect. 2022, 55, 241–248. [Google Scholar] [CrossRef]
- Börjesson, S.; Matussek, A.; Melin, S.; Löfgren, S.; Lindgren, P.-E. Methicillin-resistant Staphylococcus Aureus (MRSA) in Municipal Wastewater: An Uncharted Threat? J. Appl. Microbiol. 2010, 108, 1244–1251. [Google Scholar] [CrossRef]
- Ge, J.; Zhong, X.; Xiong, Y.; Qiu, M.; Huo, S.; Chen, X.; Mo, Y.; Cheng, M.; Chen, Q. Methicillin-Resistant Staphylococcus Aureus among Urban Rodents, House Shrews, and Patients in Guangzhou, Southern China. BMC Vet. Res. 2019, 15, 260. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Ponnusamy, L.; Mouhamadou, C.S.; Fodjo, B.K.; Sadia, G.C.; Affoue, F.P.K.; Deguenon, J.M.; Roe, R.M. Internal and External Microbiota of Home-Caught Anopheles Coluzzii (Diptera: Culicidae) from Côte d’Ivoire, Africa: Mosquitoes Are Filthy. PLoS ONE 2022, 17, e0278912. [Google Scholar] [CrossRef] [PubMed]
- Stelder, J.J.; Kjær, L.J.; Jensen, L.B.; Boklund, A.E.; Denwood, M.; Carlsen, M.; Bødker, R. Livestock-Associated MRSA Survival on House Flies (Musca domestica) and Stable Flies (Stomoxys calcitrans) after Removal from a Danish Pig Farm. Sci. Rep. 2021, 11, 3527. [Google Scholar] [CrossRef] [PubMed]
- Abdolmaleki, Z.; Mashak, Z.; Safarpoor Dehkordi, F. Phenotypic and Genotypic Characterization of Antibiotic Resistance in the Methicillin-Resistant Staphylococcus Aureus Strains Isolated from Hospital Cockroaches. Antimicrob. Resist. Infect. Control 2019, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Lowe, C.F.; Romney, M.G. Bedbugs as Vectors for Drug-Resistant Bacteria. Emerg. Infect. Dis. 2011, 17, 1132. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.-X.; Zhang, Z.-W.; Wang, C.; Yang, H.-W.; Zhang, Q.; Cheng, J. Evaluation of the CLSI Cefoxitin 30-Μg Disk-Diffusion Method for Detecting Methicillin Resistance in Staphylococci. Clin. Microbiol. Infect. 2006, 12, 1039–1042. [Google Scholar] [CrossRef]
- Goldstein, R.E.R.; Micallef, S.A.; Gibbs, S.G.; Davis, J.A.; He, X.; George, A.; Kleinfelter, L.M.; Schreiber, N.A.; Mukherjee, S.; Sapkota, A. Methicillin-Resistant Staphylococcus Aureus (MRSA) Detected at Four US Wastewater Treatment Plants. Environ. Health Perspect. 2012, 120, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Palaha, R.; Kaur, N.; Ratnakar, W.S.; Sodi, A.; Kaur, M.; Katiyar, R.; Sharma, M.; Kaur, C.; Kumar, V. Prevalence of Multidrug-Resistant, Coagulase-Positive Staphylococcus Aureus in Nasal Carriage, Food, Wastewater and Paper Currency in Jalandhar City (North-Western), an Indian State of Punjab. Environ. Monit. Assess. 2015, 187, 4134. [Google Scholar] [CrossRef]
- Lindh, J.M.; Terenius, O.; Faye, I. 16S rRNA Gene-Based Identification of Midgut Bacteria from Field-Caught Anopheles Gambiae Sensu Lato and A. Funestus Mosquitoes Reveals New Species Related to Known Insect Symbionts. Appl. Environ. Microbiol. 2005, 71, 7217–7223. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, M.P. Performance Standards for Antimicrobial Susceptibility Testing: Supplement M100, 30th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020; ISBN 978-1-68440-066-9. [Google Scholar]
- Hiramatsu, K.; Tomasz, A.; de Lencastre, H.; Perreten, V.; Holden, M.T.; Coleman, D.C. Guidelines for Reporting Novel mecA Gene Homologues. Ito T, International Working Group on the Classification of Staphylococcal Cassette Chromosome Elements (IWG-SCC). Antimicrob. Agents Chemother. 2012, 56, 4997–4999. [Google Scholar]
- Azuma, T.; Murakami, M.; Sonoda, Y.; Ozaki, A.; Hayashi, T. Occurrence and Quantitative Microbial Risk Assessment of Methicillin-Resistant Staphylococcus Aureus (MRSA) in a Sub-Catchment of the Yodo River Basin, Japan. Antibiotics 2022, 11, 1355. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al. Antimicrobials from Human Skin Commensal Bacteria Protect against Staphylococcus aureus and Are Deficient in Atopic Dermatitis. Sci. Transl. Med. 2017, 9, eaah4680. [Google Scholar] [CrossRef] [PubMed]
- Herrera, A.L.; Chaussee, M.S.; Pietri, J.E. Experimental Acquisition, Maintenance, and Transmission of Methicillin-Resistant Staphylococcus Aureus by the Common Bed Bug, Cimex Lectularius. J. Infect. Dis. 2024, 229, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Amerasinghe, F.P. Keys for the Identification of the Immature Stages of Genus Culex (Diptera: Culicidae) in Sri Lanka. J. Natn. Sci. Found. Sri Lanka 1996, 24, 37–50. [Google Scholar] [CrossRef]
- Foster, W.A.; Walker, E.D. Mosquitoes (Culicidae). In Medical and Veterinary Entomology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 261–325. [Google Scholar]
- Devanathan, N.; Mukhopadhyay, H.K.; Sihag, K.K.; Nathan, A.T.; Chakkaravarthi, A.; Srinivasan, L.; Srinivas, M.V.; Vasu, J.; Shanmugam, V.P.; Rahi, M.; et al. Synanthropic Rodents and Shrews Are Reservoirs of Zoonotic Bacterial Pathogens and Act as Sentinels for Antimicrobial Resistance Spillover in the Environment: A Study from Puducherry, India. One Health 2024, 17, 100759. [Google Scholar] [CrossRef]
- Baxtrom, C.; Mongkolpradit, T.; Kasimos, J.N.; Braune, L.M.; Wise, R.D.; Sierwald, P.; Ramsey, K.H. Common House Spiders Are Not Likely Vectors of Community-Acquired Methicillin-Resistant Staphylococcus Aureus Infections. J. Med. Entomol. 2006, 43, 962–965. [Google Scholar] [CrossRef]
- Wang, Z.; Yong, H.; Zhang, S.; Liu, Z.; Zhao, Y. Colonization Resistance of Symbionts in Their Insect Hosts. Insects 2023, 14, 594. [Google Scholar] [CrossRef] [PubMed]
- Hillyer, J.F.; Strand, M.R. Mosquito Hemocyte-Mediated Immune Responses. Curr. Opin. Insect Sci. 2014, 3, 14–21. [Google Scholar] [CrossRef]
- Kumar, A.; Srivastava, P.; Sirisena, P.; Dubey, S.K.; Kumar, R.; Shrinet, J.; Sunil, S. Mosquito Innate Immunity. Insects 2018, 9, 95. [Google Scholar] [CrossRef] [PubMed]
- Coon, K.L.; Vogel, K.J.; Brown, M.R.; Strand, M.R. Mosquitoes Rely on Their Gut Microbiota for Development. Mol. Ecol. 2014, 23, 2727–2739. [Google Scholar] [CrossRef]
- Beceiro, A.; Tomás, M.; Bou, G. Antimicrobial Resistance and Virulence: A Successful or Deleterious Association in the Bacterial World? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef]
- Abu-Hussien, S.H.; Hemdan, B.; Abd-Elhalim, B.T.; Aboul Fotouh, M.M.; Soliman, A.G.; Ghallab, Y.K.; Adly, E.; El-Sayed, S.M. Larvicidal Potential, Antimicrobial Properties and Molecular Docking Analysis of Egyptian Mint (Mentha rotundifolia) against Culex pipiens L. (Diptera: Culicidae) and Midgut-Borne Staphylococcus aureus. Sci. Rep. 2024, 14, 1697. [Google Scholar] [CrossRef]
- Queipo-Ortuño, M.I.; De Dios Colmenero, J.; Macias, M.; Bravo, M.J.; Morata, P. Preparation of Bacterial DNA Template by Boiling and Effect of Immunoglobulin G as an Inhibitor in Real-Time PCR for Serum Samples from Patients with Brucellosis. Clin. Vaccine Immunol. 2008, 15, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Brakstad, O.G.; Aasbakk, K.; Maeland, J.A. Detection of Staphylococcus Aureus by Polymerase Chain Reaction Amplification of the Nuc Gene. J. Clin. Microbiol. 1992, 30, 1654–1660. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, N.; Ganaie, F.; Mitra, S.; Tewari, R.; Dey, T.K.; Ojha, R.; Shome, R.; Shome, B.R. Development and Validation of Multiplex Polymerase Chain Reaction Assay for Concomitant Detection of Genus Staphylococcus and Clinically Relevant Methicillin Resistance Determinants. J. Appl. Biol. Biotechnol. 2020, 8, 1–6. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Blaize, J.F. Serial Dilutions and Plating: Microbial Enumeration; MyJoVE Corporation: Cambridge, MA, USA, 2024. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial | MIC (µg/mL) | Interpretation | Antimicrobial | MIC (µg/mL) | Interpretation |
|---|---|---|---|---|---|
| Cefoxitin Screen | ≥4 | R | Linezolid | 2 | S |
| Benzylpenicillin | ≥0.5 | R | Daptomycin | 0.5 | S |
| Oxacillin | ≥4 | R | Teicoplanin | ≤0.5 | S |
| Gentamicin | 4 | S | Tetracycline | ≤1 | S |
| Ciprofloxacin | ≥8 | R | Tigecycline | ≤0.12 | S |
| Levofloxacin | 4 | R | Nitrofurantoin | ≤16 | S |
| Inducible Clindamycin Resistance | + | + | Rifampicin | ≤0.03 | S |
| Erythromycin | ≥8 | R | Trimethoprim | 80 | R |
| Clindamycin | 0.25 | R | Vancomycin | ≤0.5 | S |
| Feeding | Day 1 | Day 3 | Day 5 | Day 7 | Day 14 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ex | Hm | Ex | Hm | Ex | Hm | Ex | Hm | Ex | Hm | |
| Larvae reared in MRSA-spiked water | + (1/3) | − | + (1/3) | − | − | − | − | − | − | − |
| Larvae reared in unspiked water | − | − | − | − | − | − | − | − | − | − |
| Adults fed with MRSA-spiked blood through a sterile membrane | − | + (1/3) | − | − | − | − | − | − | − | − |
| Adults fed with unspiked blood through MRSA-smeared membrane | − | + (1/3) | − | − | − | − | − | − | − | − |
| Adults fed with unspiked blood through a sterile membrane | − | − | − | − | − | − | − | − | − | − |
| S. No. | Sample | Larvae | Pupae | ||
|---|---|---|---|---|---|
| External Surface (n = 48 Pools) | Homogenate (n = 48 Pools) | External Surface (n = 48 Pools) | Homogenate (n = 48 Pools) | ||
| 1 | 16SrRNA+ | 26 | 6 | 19 | 1 |
| 2 | mec A+ | 0 | 0 | 0 | 0 |
| 3 | nuc+ | 0 | 0 | 0 | 0 |
| S. No | Genes | Oligonucleotide Sequence (5′–3′) | Cycling Conditions |
|---|---|---|---|
| 1 | 16S rRNA (842 bp) | FP: GTGATCGGCCACACTGGA RP:CAACTTAATGATGGCAACTAAGC | Initial denaturation: 94 °C, 5 min Denaturation: 94 °C, 1 min Annealing: 52.5 °C, 1 min Elongation: 72 °C, 1 min Final elongation: 72 °C, 10 min |
| 2 | mecA (293 bp) | FP: ACGAGTAGATGCTCAATATAA RP: CTTAGTTCTTTAGCGATTGC | |
| 3 | mecC (584 bp) | FP: GCTCCTAATGCTAATGCA RP: GGCTTAGAACGCCTCTATGA | |
| 4 | nuc (270 bp) | FP: GCGATTGATGGTGATACGGT RP:AGCCAAGCCTTGACGAACTAAAGC | Initial denaturation: 95 °C, 10 min Denaturation: 94 °C, 1 min Annealing: 55 °C, 30 s Elongation: 72 °C, 1.5 min Final elongation: 72 °C, 5 min |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arif, W.; Thangaraj, G.; Srinivasan, P.; Srirama, S.; Devaraju, P. Culex quinquefasciatus Mosquitoes Resist Acquisition of Methicillin-Resistant Staphylococcus aureus: Insights from Field and Laboratory Studies. Antibiotics 2024, 13, 618. https://doi.org/10.3390/antibiotics13070618
Arif W, Thangaraj G, Srinivasan P, Srirama S, Devaraju P. Culex quinquefasciatus Mosquitoes Resist Acquisition of Methicillin-Resistant Staphylococcus aureus: Insights from Field and Laboratory Studies. Antibiotics. 2024; 13(7):618. https://doi.org/10.3390/antibiotics13070618
Chicago/Turabian StyleArif, Waseema, Gowdham Thangaraj, Pushpa Srinivasan, Srikanth Srirama, and Panneer Devaraju. 2024. "Culex quinquefasciatus Mosquitoes Resist Acquisition of Methicillin-Resistant Staphylococcus aureus: Insights from Field and Laboratory Studies" Antibiotics 13, no. 7: 618. https://doi.org/10.3390/antibiotics13070618
APA StyleArif, W., Thangaraj, G., Srinivasan, P., Srirama, S., & Devaraju, P. (2024). Culex quinquefasciatus Mosquitoes Resist Acquisition of Methicillin-Resistant Staphylococcus aureus: Insights from Field and Laboratory Studies. Antibiotics, 13(7), 618. https://doi.org/10.3390/antibiotics13070618